Chikungunya Virus–Vector Interactions

1
Center for Vectorborne Diseases, School of Veterinary Medicine, University of California, Davis, CA 95616, USA
2
Department of Pathology, Microbiology and Immunology, School of Veterinary Medicine, University of California, Davis, CA 95616, USA
3
Department of Virology, Arboviruses and Insect Vectors, Institut Pasteur, 25-28 rue du Dr. Roux, 75724 Paris cedex 15, France
4
Institute for Human Infections and Immunity, Center for Tropical Diseases and Department of Pathology, University of Texas Medical Branch, Galveston, TX 77555, USA
*
Author to whom correspondence should be addressed.
Viruses 2014, 6(11), 4628-4663; https://0-doi-org.brum.beds.ac.uk/10.3390/v6114628
Submission received: 3 September 2014
/
Revised: 10 November 2014
/
Accepted: 10 November 2014
/
Published: 24 November 2014
(This article belongs to the Special Issue Interactions between Arboviruses and Arthropod Vectors)
Abstract
:Chikungunya virus (CHIKV) is a mosquito-borne alphavirus that causes chikungunya fever, a severe, debilitating disease that often produces chronic arthralgia. Since 2004, CHIKV has emerged in Africa, Indian Ocean islands, Asia, Europe, and the Americas, causing millions of human infections. Central to understanding CHIKV emergence is knowledge of the natural ecology of transmission and vector infection dynamics. This review presents current understanding of CHIKV infection dynamics in mosquito vectors and its relationship to human disease emergence. The following topics are reviewed: CHIKV infection and vector life history traits including transmission cycles, genetic origins, distribution, emergence and spread, dispersal, vector competence, vector immunity and microbial interactions, and co-infection by CHIKV and other arboviruses. The genetics of vector susceptibility and host range changes, population heterogeneity and selection for the fittest viral genomes, dual host cycling and its impact on CHIKV adaptation, viral bottlenecks and intrahost diversity, and adaptive constraints on CHIKV evolution are also discussed. The potential for CHIKV re-emergence and expansion into new areas and prospects for prevention via vector control are also briefly reviewed.
1. Introduction
Emerging and re-emerging arthropod-borne viruses (arboviruses) represent a significant threat to human and veterinary health worldwide. Chikungunya virus (CHIKV), a mosquito-borne alphavirus that causes chikungunya fever (CHIK), a severe, debilitating and often chronic arthralgia, is a prime example; since it was first isolated in 1952, the virus has been detected as the etiologic agent of sporadic epidemics in Africa and Asia and, since 2004, has expanded its geographic range to circulate on Indian Ocean islands, and in Italy, France, and the Americas. CHIKV has also re-emerged in Southeast Asia since 2006, causing an estimated 1.3 million human cases [1]. CHIKV cycles in urban settings between humans and two mosquito species found in the U.S., suggesting a potential for endemic establishment there. As evidence of this, 11 autochthonous cases of CHIK were detected in south Florida as of November 2014 [2,3]. Emergence of arboviruses like CHIKV underscores the interconnectedness of humans with their environments, and highlights our vulnerabilities to new disease threats posed by spreading viruses. Understanding how arboviruses like CHIKV emerge is critical to predict and prevent or mitigate human disease. Central to understanding emergence is knowledge of the natural ecology of CHIKV transmission and the dynamics of vector infections. Here we discuss current knowledge of CHIKV infection dynamics in mosquito vectors and its relationship to human disease emergence.
2. Chikungunya Infection and Vector Life History Traits
2.1. Chikungunya Virus Transmission Cycles and Genetic Origins
Chikungunya virus is endemic in countries of Sub-Saharan Africa, India and Southeast Asia. The virus circulates in an enzootic cycle in Africa between forest-dwelling mosquitoes and non-human primates [4]. Phylogenetic studies reveal the existence of two major enzootic CHIKV lineages in Africa: Western, and East/Central/South African (ECSA) [5]. In Asia, where the first outbreak was reported in 1958 in Thailand, CHIKV has historically been maintained in an urban cycle transmitted to humans by the mosquito Aedes (Stegomyia) aegypti and, to a lesser extent, Aedes (Stegomyia) albopictus [6].
2.2. Chikungunya Distribution, Emergence and Spread
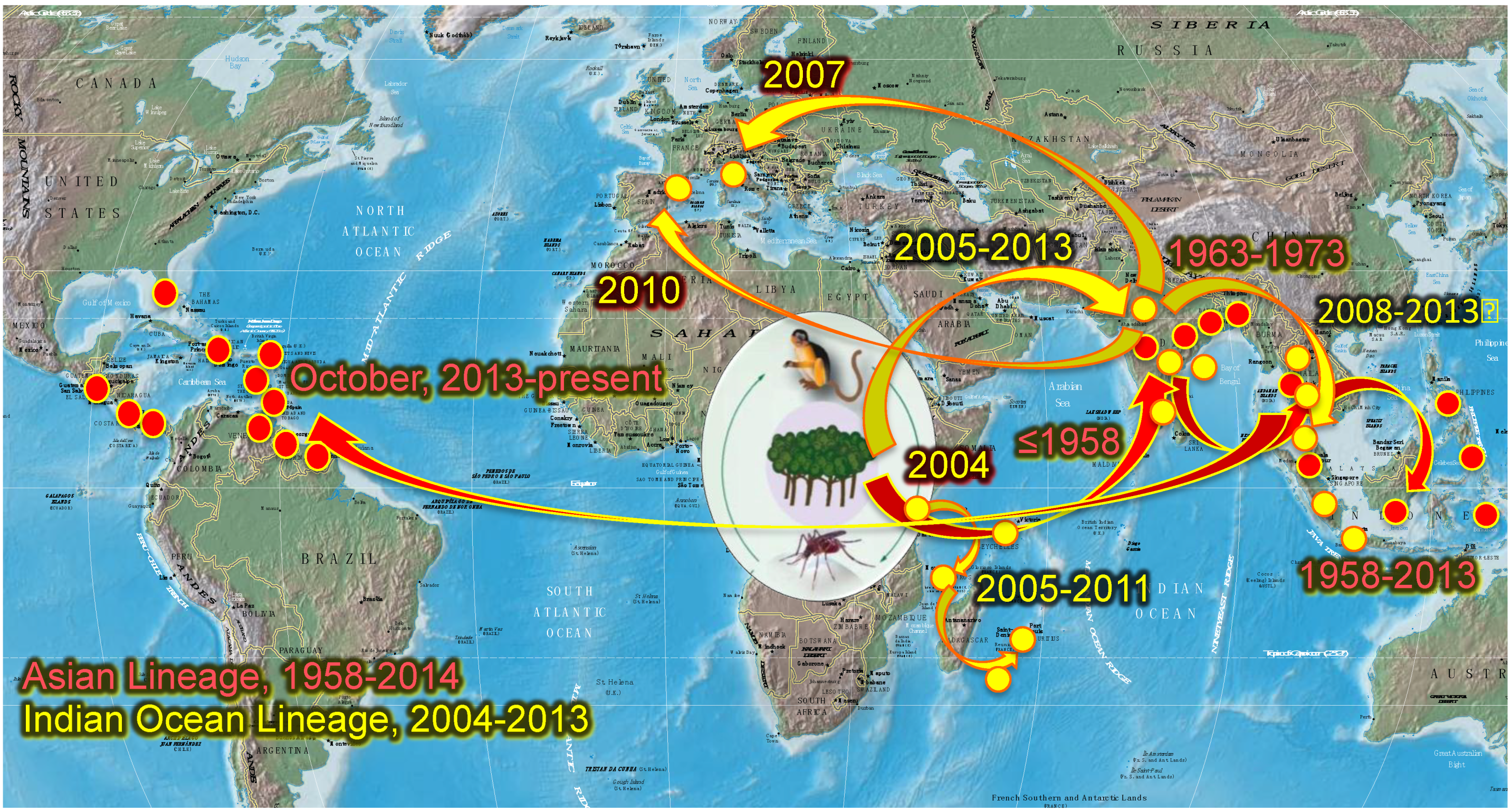
In 2004, CHIKV belonging to the ECSA lineage emerged from Lamu and Mombasa in coastal Kenya [7] and spread to Comoros and later, to other islands of the Indian Ocean including La Réunion (Figure 1). There, the predominant vector A. albopictus transmitted preferentially a CHIKV variant with a single amino acid change from an alanine (A) to valine (V) at E1 envelope glycoprotein amino acid 226 of the ECSA Indian Ocean lineage (IOL) genotype [8]. The E1-226V variant was more efficiently transmitted by A. albopictus [9,10], with roughly 40-fold more efficient initial infection of midgut epithelial cells [9,10,11]. The selection of the E1-226V variant occurs at the initial infection of the midgut of A. albopictus, leading to a higher viral dissemination and transmission of the IOL genotype by this mosquito [12]. A series of four additional adaptive mutations in the E2 gene have also been incriminated more recently in enhancing transmission by A. albopictus [6,9,13]. During 2005–2006, the virus spread to neighboring Indian Ocean islands including Mayotte, Mauritius and Madagascar, where CHIKV E1-A226V was mainly transmitted by A. albopictus [12,14,15,16]. Subsequently, the CHIKV IOL was introduced to India [17,18] and the surrounding islands, Sri Lanka [19] and the Maldives. In Africa, the CHIKV E1-226V variant was also implicated outbreaks in Cameroon [20], Gabon [21,22] and Congo [23]. This variant also caused the first European CHIKV outbreak in Italy in 2007 [24]. Since 2008, IOL CHIKV strains were also imported into Southeast Asia: Malaysia [25], Singapore [26], Thailand [27], China [28], Cambodia [29] and Bhutan [30]. Remarkably, the E1-226V variant was found preferentially in rural areas where A. albopictus was more abundant than A. aegypti, and presumably was the primary vector [31,32]. In September 2010, autochthonous cases of CHIKV were reported in southeast of France [33,34], again with A. albopictus as the vector [34,35,36]. In Southeast France, this species appears to behave differently compared to its tropical counterpart, as it efficiently transmits the E1-226A IOL variant detected in local circulation [34,36]. Aedes albopictus has been found in in 18 French departments [34,35,37] as well as 19 other countries in Europe [38,39].
Prior to December 2013, CHIKV transmission was not documented in the Americas, despite numerous introductions and the presence of conditions that are apparently suitable for its establishment [6]. In October 2013, two laboratory-confirmed, autochthonous CHIKV cases were detected in the French territory of Saint Martin Island, in the Caribbean Sea. Surprisingly, the CHIKV strain isolated belonged to the Asian genotype [40] rather than the IOL that emerged in 2004 in the Indian Ocean Basin and Asia [6]. This St. Martin strain was phylogenetically close to CHIKV identified in Indonesia in 2007, China in 2012 and Philippines in 2013 [40], but more distant from the Asian genotype that circulated in New Caledonia [41]. The only vector implicated in St. Martin, where A. albopictus has not been established, was A. aegypti. Very rapidly, an epidemic was established in the island and subsequently, CHIKV progressively spread throughout most of the Caribbean, and into Central and South America where human populations are mostly naïve to CHIKV [42].
Figure 1.
Geographic distribution and spread of Chikungunya virus (CHIKV) and its two urban vectors, A. aegypti and A. albopictus. Modified from reference [42], with permission.
Figure 1.
Geographic distribution and spread of Chikungunya virus (CHIKV) and its two urban vectors, A. aegypti and A. albopictus. Modified from reference [42], with permission.

American populations of A. aegypti and A. albopictus are susceptible to both ECSA/IOL and Asian genotypes of CHIKV with higher susceptibility observed for A. aegypti and the Asian CHIKV genotype, or A. albopictus and the ECSA/IOL genotype [43]. As of November 2014, 11 locally acquired cases of CHIKV have been documented in Florida, USA [44]. These recent findings and the history of CHIKV movement suggest that it will continue to spread throughout the Americas and other tropical and subtropical regions of the world wherever mosquito vectors occur.
3. Vectors
3.1. Chikungunya Vector Distribution
CHIKV is an African virus that circulates enzootically in sylvatic cycle between arboreal, canopy-dwelling mosquitoes and non-human primates. Enzootic strains from the West African and ECSA lineages have been isolated from diverse mosquito species in Senegal, Ivory Coast, Central African Republic, Uganda and South Africa [4,45,46,47]. In Senegal, CHIKV has been detected in vectors of yellow fever virus: Aedes (Diceromyia) furcifer, A. (Diceromyia) taylori, A. (Stegomyia) luteocephalus, A. (Stegomyia) africanus and A. (Stegomyia) neoafricanus [47]. In South Africa, A. (Diceromyia) cordellieri, was also involved in CHIKV transmission [48]. The urbanization of CHIKV, with epidemics occurring in peridomestic settings following the emergence of enzootic strains, coincides with the involvement of anthropophilic mosquitoes: A. (Stegomyia) aegypti (L.) and A. (Stegomyia) albopictus (Skuse), favoring inter-human transmission. Aedes aegypti, first described by Linné in 1862, originated in African forests and is today present in most tropical countries [49]. In Africa, A. aegypti is present in two genetic forms: (i) the dark and sylvatic A. aegypti formosus, found in forested habitats and using treeholes as larval development sites; and (ii) the pale and domestic A. aegypti aegypti, which is widespread in the tropics and subtropics and uses artificial larval habitats mainly in urban environments [50,51,52]. Aedes aegypti aegypti originated from the forest-dwelling formosus form, which may have spread from tropical African forests to North Africa where it probably became domesticated under pressure to use artificial water storage containers as larval habitats [53]. These mosquitoes progressively differentiated into domesticated populations known as A. aegypti aegypti. Human trading activities later introduced this subspecies globally throughout the most of the tropics and subtropics: the New World via the African slave trade from the 15th to 19th centuries, Asia in the 18th and 19th centuries, and the Pacific islands with troop movements during World War II.
Aedes aegypti can exist sympatrically with A. albopictus and also often shares larval habitats [16,54,55,56]. Aedes albopictus, originally described by Skuse in Calcutta, India in 1894 originated in forests of Southeast Asia [57], but is now is commonly found in peri-urban, rural and forested areas on five continents [58]. Aedes albopictus has no particular ecological specialization, colonizing both temperate and tropical regions [59]. Two types of populations are described [60]: (1) temperate populations imported to the U.S. from Japan [61,62] and then from the U.S. to Europe [63,64], where they are now established in 20 European countries [58]; these temperate populations are characterized by diapausing, cold-resistant eggs [65]; and (2) tropical populations [60]. Aedes albopictus is a competent vector for at least 26 arboviruses [66] and is implicated occasionally in DENV (e.g., on the Seychelles islands [67]; and more frequently in CHIKV transmission (e.g., on La Réunion Island [68]; see Table 1 of this review as well). Since the 2007 outbreak of CHIKV in Italy [24], Europe is considered vulnerable for transmission of several “tropical” arboviruses, particularly in regions where A. albopictus is present [34,69,70].
3.2. Vector Dispersal and Genetics
Aedes aegypti and A. albopictus have both spread beyond their native ranges via commercial trade and dessication-resistant eggs [71]. Aedes aegypti largely replaced A. albopictus in Southeast Asian cities in the first half of the 20th century [65] while the introduction of A. albopictus into the Americas during the 1980s was associated with a decline in the abundance of A. aegypti in some regions such as occurred in the Americas in the 1980s [59,72,73] and is now ongoing in Central Africa [74,75] and on islands of the Indian Ocean region [16,76]. Long distance spread of CHIKV urban vectors beyond their natural flight ranges, typically a few hundred meters to a few kilometres [77], is usually achieved through transportation of immature stages (i.e., larvae and eggs). From the 15th century onwards, successive waves of invasion by A. aegypti and, more recently, A. albopictus, have been facilitated by commercial transport.
Such mosquito invasions can be traced using molecular markers, which can now be developed from genome sequences. The A. aegypti genome sequence is complete [78] and genome annotations of A. albopictus are expected soon [79]. To assess mosquito phylogenetics, molecular markers used to define mosquito invasions have mainly been microsatellites (reviewed in [80]) and mtDNA [74,76,81,82,83,84]. Scenarios of invasions are more easily defined in island systems where mosquito populations have been geographically isolated with gene flow measurable through human displacements [85]. The involvement of human activities in reshuffling A. albopictus distributions has been demonstrated [86]. In Madagascar, A. albopictus dominant in the eastern coast and highland areas [87] are genetically distant from populations from South America and Southeast Asia [76]. This species continues to extend its geographic distribution in Madagascar, increasing its densities and progressively replacing A. aegypti, which is now present at residual levels in remote areas [76]. The decline in A. aegypti was also detected in the neighboring island of La Réunion [16,88].
The situation with these mosquitoes in Central Africa is slightly different. The invasion of A. albopictus occurred via several successive waves within a very short time frame. In the Central African Republic (CAR) A. albopictus first detected in 2008 [89], probably introduced from Cameroon where the species has been established since 2000 [90]. Aedes albopictus in Cameroon derives from multiple introductions from tropical sources that still need to be identified [74]. The polymorphism of mtDNA markers there is low, suggesting that A. albopictus were periodically introduced and that these events coincided with a decrease in A. aegypti densities [75].
3.3. Vector Competence
Vector competence, the ability of an arbovirus vector to acquire a pathogen and successfully transmit it to another susceptible host, is a complex process influenced by external factors including temperature, the availability of vertebrate hosts, vector population density and predation, as well as internal factors including mosquito survival and virus replication. For CHIKV, horizontal transmission through saliva that is injected when a mosquito probes or feeds on blood is the most common mechanism of transmission (Figure 2), although vertical transmission via infected eggs may also occur at a low rate [91]. Vector competence is typically evaluated experimentally using static laboratory-based methods, where assessment of virus replication is the primary endpoint. For CHIKV, mosquitoes from endemic locations are usually presented artificial bloodmeals loaded with known titers of virus from sympatric settings. Mosquitoes that imbibe infectious bloodmeals are incubated at a constant temperature for at least seven days and then killed to assess replication. The fractions of mosquitoes that become infected, develop infections that have disseminated from the initial infection site in the midgut into the hemocoel, and expectorate CHIKV in saliva are then measured to extrapolate vector competence for the population.
Figure 2.
Process of infection and transmission of CHIKV by mosquito vectors. Vector mosquitoes ingest CHIKV from viremic vertebrate hosts during bloodfeeding. During the extrinsic incubation period that occurs in the vector, CHIKV infects the midgut and disseminates through the body cavity to infect salivary glands for secretion into saliva. Re-feeding vectors transmit CHIKV to vertebrate hosts by expectorating virus in saliva. New vectors perpetuate the CHIKV cycle by ingesting virus during intrinsic incubation, a period of viremia in the vertebrate host. Vector competence assays test infection, dissemination, and transmission of CHIKV after extrinsic incubation.
Figure 2.
Process of infection and transmission of CHIKV by mosquito vectors. Vector mosquitoes ingest CHIKV from viremic vertebrate hosts during bloodfeeding. During the extrinsic incubation period that occurs in the vector, CHIKV infects the midgut and disseminates through the body cavity to infect salivary glands for secretion into saliva. Re-feeding vectors transmit CHIKV to vertebrate hosts by expectorating virus in saliva. New vectors perpetuate the CHIKV cycle by ingesting virus during intrinsic incubation, a period of viremia in the vertebrate host. Vector competence assays test infection, dissemination, and transmission of CHIKV after extrinsic incubation.

Table 1 summarizes experimental CHIKV vector competence estimates in arthropods from around the world. Most studies have focused on the two primary urban vectors, A. aegypti and A. albopictus, although other mosquitoes and one tick species have also been tested. Most studies have used relatively high bloodmeal titers exceeding 7 log10 plaque forming units (PFU)/mL (but that are still below peaks in infected humans [92]) and mosquitoes were typically sampled more than seven days post-feed, experimental designs that maximize the chances of CHIKV infection and transmission. Infection, dissemination, and transmission rates of A. aegypti and A. albopictus vary according to the geographic sources of mosquitoes tested. At bloodmeal titers of ≥7 log10 plaque-forming units (PFU)/mL, at least 80% of A. aegypti from all locations develop disseminated infections. More than half of A. albopictus that ingest bloodmeals ≥7 log10 PFU/mL also became infected or develop disseminated infections, with several exceptions. Infection and dissemination rates in A. albopictus from the US are dose-dependent and increase with the titer of the ingested bloodmeal [9,93]; dose response studies have not been published for mosquitoes from other regions. Aedes albopictus also exhibits strain-specific susceptibility; fewer mosquitoes from La Réunion Island, Mayotte, and colonized mosquitoes originally from Texas infected in parallel with a CHIKV isolate from early in the 2005–6 La Réunion Island outbreak developed disseminated infections than mosquitoes infected with strains isolated later in the outbreak (subsequent studies discussed below revealed that A to V substitution in amino acid 226 of the E1 protein (E1-A226V) mediates much of this differential infectivity [9,10,94]). In an attempt to exhaustively present CHIKV vector competence data, Table 1 shows all published data; however, comparing CHIKV vector competence over time in mosquitoes sampled from the same areas entails comparing data from experiments with unmatched bloodmeal titers. Differences could result from varied ingested doses rather than real differences in susceptibility to infection, dissemination and transmission. Despite this, studies from many locations worldwide show that A. aegypti and A. albopictus are generally highly susceptible to infection, dissemination and transmission of CHIKV. Alternate vector species, also represented in Table 1, may also serve as efficient CHIKV vectors in specific geographic settings; e.g., Opifex fuscus mosquitoes from New Zealand are highly competent at transmitting CHIKV from India.
3.4. Chikungunya Virus Vector Immunity and Microbial Interactions
While most arboviruses induce significant morbidity and/or mortality in some vertebrate hosts, infections of arthropod vectors are generally considered non-pathogenic. However, interactions between the replicating virus and the mosquito immune defense system produce an outcome that may influence subsequent viral dissemination or superinfection by other viruses (reviewed in [95]). Considerable progress has been achieved in understanding the innate defenses of the mosquito against arboviruses. The virus most intensively studied, DENV, triggers several immune pathways in A. aegypti: the Toll [96,97], JAK-STAT [98], and Imd/JNK pathways [99]. However, RNA interference (RNAi) appears to be the most significant innate antiviral immune response in mosquitoes [99,100,101,102,103,104,105]. So far, three major types of small RNA molecules have been identified in mosquito vectors: small interfering RNA (siRNA), microRNA (miRNA) and PIWI-interacting RNA (piRNA). These molecules have distinct roles in different cellular processes and virus-host interactions [106]. The antiviral siRNA pathway is triggered when long double-stranded RNA molecules are produced from secondary RNA structures and/or viral replication intermediates during infection of the vector. This leads to an activation of the RNA degradation machinery to the target viral RNA [100]. These pathways may act in a virus-specific manner.
Little is known about immune responses induced by CHIKV infection of mosquitoes. Viral replication is controlled via the exogenous RNAi pathway in mosquitoes [107]. The protein Ago-2 plays an important role in the antiviral RNAi response to CHIKV, similar to its role for other alphaviruses including, Sindbis (SINV) [102], Semliki Forest [108] and o’nyong-nyong viruses [101]. Viral replication under RNAi control may limit potential pathologic effects to favor mosquito survival [94].
The repeated use of insecticides to target mosquitoes as a means to control vector-borne diseases has found its limits due to the development of resistance [109], and alternative approaches are urgently needed. Recent strategies for controlling viral transmission have come from research on RNAi, such as the development of A. aegypti mosquitoes expressing small RNAs that render them resistant to viral infection; this approach is a promising mechanism for suppressing virus replication in mosquitoes [110,111,112]. Depending on the objectives, mosquito populations can also be reduced in density using various strategies including the Sterile Insect Technique (SIT), a SIT-like system called Release of Insects carrying a Dominant Lethal (RIDL) [113], or replaced by transmission-refractory mosquitoes [114]. For this latter strategy, Aedes mosquitoes have been successfully transformed using transposon vectors [115] or infections with the intracellular bacterium Wolbachia [116]. However, a system is required to spread the refractory genes into mosquito populations as well as to maintain the expression of the refractory phenotype through generations. Several gene-driver systems have been proposed [117] including the Medea system, which shows promise by coupling genes conferring disease refractoriness with a genetic mechanism for driving them through wild populations [118].
In addition to their ability to shorten mosquito life span [119], certain strains of Wolbachia are also able to reduce arboviral transmission [120,121]. Wolbachia induce various distortions of host reproduction via a form of sterility known as cytoplasmic incompatibility (CI), thereby promoting its spread into populations [122]. Therefore, certain strains of Wolbachia provide the double role of a gene driver system and carrier of a refractory phenotype. While the mosquito A. aegypti is free of natural endosymbiotic bacteria Wolbachia, some A. albopictus populations are naturally super-infected with two Wolbachia strains, wAlbA and wAlbB. These Wolbachia, present in mosquito midguts and salivary glands [123], do not affect CHIKV replication [124]. In contrast, Wolbachia are able to limit DENV in A. albopictus [125] but cannot completely block transmission [126]. Moreover, A. albopictus transfected with a heterologous Wolbachia isolated from the fruit fly Drosophila melanogaster (wMel) inhibit the transmission of CHIKV [127] as in A. aegypti [121,128]. Several mechanisms have been proposed to explain the molecular basis of the pathogen-blocking Wolbachia infection phenotype: upregulation of immune genes, production of reactive oxygen species, or competition for a limited resource such as cholesterol [129,130,131].
In their digestive tracts, wild populations of A. albopictus and A. aegypti have been shown to house Proteobacteria and Firmicutes, including the genera Acinetobacter, Asaia, Delftia, Pseudomonas, Wolbachia and Bacillus as well as members of the family Enterobacteriaceae [132]. A higher diversity of bacteria can be occasionally found in wild A. albopictus; 27 genera of cultivable bacteria have been detected in this species from Madagascar, with Pantea bacterium as the most prevalent [133]. Other bacterial members of the Alpha- and Gammaproteobacteria phyla, as well as Bacteroidetes, respond to CHIKV infection [132]. The abundance of bacteria belonging to the Enterobacteriaceae family increases with CHIKV infection [132], suggesting that cooperation or competition occurs within the host. Microbiota (including Wolbachia) endogenous to some mosquito species pre-activate the expression of basal genes in the immune response (i.e., immune priming), allowing the vector to be prepared for infection by pathogens. In A. aegypti infected with DENV, the arthropod microbiota elicit basal immune responses that act against the virus, and this response reduces the density of the microbial load in the mosquito midgut [134]. Antiviral activities can be induced by secreting antiviral compounds; Serratia odorifera in A. aegypti enhances susceptibility to CHIKV by interaction of its P40 protein with the mitochondrial protein porin present on the midgut brush border membrane of the mosquito midgut, thereby downregulating mosquito immune responses [135].
3.5. Vector co-Infection by Chikungunya and Other Arboviruses
Mosquitoes may feed several times during their lifespan and can ingest genetically distinct variants of the same virus species or even viruses from different families. Once a virus has infected the mosquito salivary glands, the mosquito becomes competent for transmission to the next vertebrate host, usually for the remainder of its life [136,137]. The mosquito can therefore host a collection of diverse viruses, playing a role in selecting genotypes involved in epidemics [138].
Because CHIKV circulates in DENV-endemic regions where the anthropophilic mosquitoes A. albopictus and A. aegypti can transmit both viruses, reports of co-infection in humans are increasing. Since the first observation in 1964 in South India [139], co-infections have been reported since the 2004 re-emergence: Sri-Lanka [140], India [141,142,143], Malaysia [144], Gabon [145], Madagascar [14], Singapore [146], and Angola [147]. The increasing number of reports of co-infections seems to coincide with introductions of A. albopictus [145]. In Gabon, between 2007 and 2010, 0.9% of 4287 acutely febrile patients were positive for both CHIKV and DENV, and more unexpectedly, co-infected A. albopictus were also collected [148]. Patients were possibly co-infected with the two viruses through the bite of a single mosquito, as has been demonstrated experimentally [149]. Considering the nearly worldwide circulation of DENV and CHIKV, co-infections may become more frequent. Concurrent infections may make diagnosis more challenging and could also result in different disease syndromes. Concurrent epidemics of yellow fever virus and CHIKV were also reported in Africa [150,151,152] and patients infected with the two viruses were suspected [153]. In the same way, Zika virus (ZIKV), typically transmitted in urban settings by A. aegypti, is also transmitted by A. albopictus, whose expanding distribution may favor the cocirculation of CHIKV and ZIKV [154].
4. Chikungunya Virus
4.1. Genetics of Vector Susceptibility and Host Range Changes
As described above, CHIKV circulates in two distinct transmission cycles: (1) enzootic transmission among nonhuman primates and perhaps other vertebrates by arboreal Aedes spp. mosquitoes in sub-Saharan African sylvatic foci, and; (2) urban transmission among humans by A. aegypti and/or A. albopictus. Like interactions between other arboviruses and their arthropod vectors, susceptibility to CHIKV infection of mosquitoes and their ability to transmit depend on the genetics of both, and differences in these properties can affect circulation and human exposure as described above. Enzootic vector-CHIKV interactions have received little experimental study [47]. Although population-based differences in urban vector competence may occur, these have not been addressed in enough detail to be conclusive or to begin to assess genetic components of vector susceptibility. Following the detection of the A. albopictus-adaptive A226V substitution in the E1 envelope glycoprotein during the 2005–2006 Réunion Island epidemic [8,9,10], the impact of CHIKV genetics on urban vector infection and transmission has received considerable study. Vector-adaptive evolution had been previously described for DENV [155,156] and the alphavirus Venezuelan equine encephalitis virus (VEEV) [157], the latter also involving a substitution in an envelope glycoprotein, in this case E2. Surprisingly, neither vector-adaptive alphavirus mutations has been shown to have much effect on infection of the previous or donor vector, A. aegypti in the case of CHIKV [9,12] and Culex (Melanoconion) taeniopus in the case of VEEV [158], challenging the hypothesis that most host-specific viral adaptations will have tradeoffs for fitness in other hosts. Further phylogenetic/reverse genetic studies of IOL CHIKV revealed an unprecedented series of four second-step, A. albopictus-adaptive mutations, each involving E2 substitutions, and one relying also on a synergistic effect of an E3 substitution [13,159]. Each of these mutations enhances initial infection of the mosquito midgut and has little or no effect on infection of A. aegypti. Furthermore, at least one combination of these independently acquired second-step mutations leads to an A. albopictus infection phenotype more efficient than that of any natural CHIKV strain studied to date, suggesting further vector-adaptive evolution and even more efficient circulation in regions where this mosquito is abundant. Structural modelling of these A. albopictus-adaptive envelope glycoprotein substitutions suggests that they alter the entry process in endosomes by affecting conformational changes required for E1 fusion with endosomes rather than directly affecting receptor binding [12,13].
4.2. Population Heterogeneity & Selection for Fittest Genomes
Alphaviruses like CHIKV exist as heterogeneous populations of viral RNAs called mutant swarms that arise from frequent nucleotide misincorporation during replication due to the inability of the viral RNA-dependent RNA polymerase (RdRp) to error correct. Given a ≈12 kb CHIKV genome and a mutation frequency of ≈10−4, each new RNA genome possesses one mutation on average. By comparison, double-stranded DNA virus mutation rates are several log10 lower [160]. Most mutant genomes are detrimental and removed from the swarm via purifying negative selection. By contrast, positive selection of a fit phenotype results in increased abundance of a genotype. Therefore, a dynamic mutation-selection balance determines the size and genetic diversity of a mutant swarm. Genetic diversity renders a population less prone to the consequences of negative selection pressures that target certain genotypes and renders a population more likely to contain variants with potential phenotypic advantages; these features can enhance plasticity and adaptability. A high fidelity CHIKV variant with a point mutation at amino acid position 483, a fidelity-determining locus in the RdRp, that was discovered experimentally after treatment with chemical mutagens, generates 30% fewer mutants than wild-type (wt) virus and is less fit in vectors and a mouse model [161,162]. This reduced fitness may result from the less mutated population containing fewer genotypes that can resist evolutionary pressures including negative selection or population bottlenecks. Complementary studies with low fidelity variants of CHIKV generated by mutagenizing the amino acid at 483 produce more errors than wt variants and are also attenuated in mosquito cells and mice [138]. Together these studies indicate that CHIKV maintains an intermediate mutation frequency to avoid detriments to fitness resulting from populations with too few or too many mutant genomes.
4.3. Dual Host Cycling & Chikungunya Virus Adaptation
Strong purifying selection of alphaviruses that cycle between mosquitoes and vertebrates results in less genetic variation than predicted by their high mutation rates [138,163,164,165]. Work with other alphaviruses including eastern equine encephalitis virus (EEEV), SINV, and VEEV shows that alternating between hosts poses conflicting challenges to replication that can limit adaptation to either host alone by imposing fitness costs where adaptations are antagonistic ([166] reviewed in [167]). When one host is artificially removed via experimental serial passage, the limitations of these trade-offs are evident; viruses serially passaged in a single host are more adaptable. CHIKV serially passaged in vertebrate or mosquito cells exhibits higher fitness when passaged in novel cell types and also showed enhanced neutralization escape and antiviral compound resistance. These changes are accompanied by increased genetic diversity. In contrast, alternating CHIKV passage between cell types restricts fitness and increases diversity, suggesting that only mutations beneficial or neutral in both host cells are maintained and that these variants retain fitness in alternating cycling [168].
4.4. Viral Bottlenecks and Intrahost Diversity
The ability to circumvent bottlenecks within and between dynamic environments including switching between vector and vertebrate hosts impacts CHIKV evolution and is important for understanding changing population dynamics that ultimately cause human disease. Bottlenecks that reduce arbovirus population size can influence viral fitness by restricting phenotypic plasticity that stems from having genetic diversity. The evolutionary theory Muller’s ratchet asserts that asexual organisms with high mutation rates and small population sizes irreversibly accumulate deleterious mutations unless compensatory mutations restore mutation-free genomes to the population [169]. Studies with EEEV [170] validate this concept; fitness decreases after serial bottleneck passages can be rescued by subsequent large population passages, albeit with much replication needed to overcome the ratchet [171]. Anatomical barriers to productive alphavirus transmission by mosquito vectors are relatively well defined, although only one study has addressed CHIKV directly. As a first step, alphaviruses in a bloodmeal ingested by a mosquito must infect the midgut epithelium. Some VEEV studies suggest that only “portal” cells in the midgut epithelium are susceptible to infection [172]; other experiments show uniform susceptibility [173], possibly reflecting a longer virus-vector relationship for the latter. Secondary impediments to dissemination result in failure of the virus to escape from the midgut epithelium, infect salivary glands, and escape from salivary gland cells into saliva for transmission to vertebrates. The number of barcoded VEEV variants that successfully traverse these barriers is reduced at midgut escape and salivary gland infection compared to the ingested bloodmeal, and smaller initial bloodmeal populations are more prone to reductions in variant diversity [174], suggesting strong genetic bottlenecks that reduce diversity coincident with population size changes. These observed changes in genetic diversity of the mutant swarm in mosquitoes contrast with observations from other mosquito-borne arboviruses isolated from nature [175], as well as experimental in vivo passaging studies with VEEV that show the maintenance of genetic and phenotypic stability of the consensus (average sequence) [176]. The disparity in results between these studies may be explained by the re-establishment of genetic diversity after bottlenecks via subsequent replication. Studies with CHIKV support this idea; although population diversity in the midgut and salivary glands is reduced compared to the bloodmeal input or midgut population, respectively, it recovers downstream of each barrier and the consensus sequence remains unchanged [138].
Experimental studies showing that alternating hosts impose constraints on arboviruses including CHIKV indicate that inefficient transmission probably interrupts natural CHIKV cycling. However, the degree of natural extinction in vertebrate or vector hosts has not been directly addressed. Extinction in an individual mosquito-vertebrate-mosquito lineage is likely because most mosquitoes do not survive long enough in nature to feed more than once. The maintenance of consensus genetic stability in nature in the presence of intense circulation may be due to the extinction of most individual lineages, possibly via vector bottlenecks in individual mosquitoes. However, these extinctions must not be widespread enough; otherwise CHIKV cycling would be interrupted. No studies published on CHIKV or other alphaviruses have examined mutant swarms in naturally infected mosquitoes, humans, or other vertebrate hosts. Artificially generated, barcoded VEEV variants in mosquitoes that transmitted to laboratory mice were also observed in the brains of animals [174], suggesting that mosquito-to-vertebrate transmission of alphaviruses does not present a major bottleneck (although few (n = 3) infected mice were sampled). Virus doses expectorated by mosquitoes vary greatly, and are probably overestimated by standard laboratory salivation assays where infected mosquitoes eject more virus into tubes than in vivo [177]. Therefore, variance in transmitted doses, especially when few particles are transmitted, likely impacts the outcome of vertebrate infection and maintenance of alphavirus cycling in natural foci. Interruptions in seasonal transmission may represent another significant bottleneck imposed on CHIKV, especially if the virus is introduced into temperate climates [178] where vector survival and competence would decrease in colder temperatures [179]; however this phenomenon is complex [180] and has not been studied extensively.
4.5. Adaptive Constraints on Chikungunya Virus Evolution
Although the 2004 IOL CHIK emergence underscores the adaptive potential of RNA viruses including most arboviruses, it also provides examples of constraints on adaptive evolution that remain difficult to predict. Although the E1-A226V substitution was selected convergently after IOL strains reached locations with abundant A. albopictus, it surprisingly was not found in any CHIKV strains of the Asian lineage despite their circulation in regions of Asia native to this vector for more than 60 years [5]. This lack of adaptation to A. albopictus in Asia resulted from an epistatic interaction with E1 residue 98; Asian CHIKV strains, which have a threonine residue at position 98, show no increase in A. albopictus infectivity when the E1-A226V substitution is engineered into a cDNA clone derived from the Asian strain, while ECSA and IOL strains with 98A show a dramatic increase in infection. The lack of sequenced enzootic CHIKV strains with 98T suggests that this residue, which by itself has no detectable effect on infection of urban vectors or models for human infection, resulted from a founder effect when CHIKV was introduced into Asia sometime before 1958 [181].
Although variation among Asian strain CHIKV infection of A. aegypti has received little study, no evidence has been produced to support adaptive evolution in Asia since the 1950s. The above evidence indicating an adaptive constraint on Asian strains, including those now circulating in the Americas, for enhanced transmission by A. albopictus suggests that A. aegypti will remain the principal vectors as spread into American regions inhabited by both vectors continues [13]. However, additional studies of the Asian lineage and its potential to increase transmissibility by either mosquito are needed.
5. Future Prospects
5.1. Potential for Re-Emergence and Expansion into New Areas
Due to the immunologically almost completely naïve status of human populations and the widespread and abundant A. aegypti nearly throughout the western hemisphere tropics, CHIKV is expected to continue to spread and ultimately recapitulate the distribution of DENV in the Americas. The lack of historical evidence for a major role of A. albopictus in transmission of Asian lineage CHIKV strains and the adaptive constraint described above suggest that temperate American regions inhabited by A. albopictus but not A. aegypti may not be at as high a level of risk as regions where IOL strains are circulating. However, IOL strains continue to circulate in Asia and have a history of many importations into the Americas [182], so their risk of introduction into the Americas continues. Should both Asian and IOL strains cocirculate, as they have in Southeast Asia since 2007, even more geographic regions of the Americas could be at risk for CHIK.
5.2. Prospects for Prevention via Vector Control
The poor history of DENV control since the 1970s suggests that mitigation of CHIKV transmission via vector control will be highly challenging. Due to its tight association with artificial larval habitats, endophily of adult females, and daytime biting patterns, control of A. aegypti and DENV transmission is difficult, and A. albopictus presents similar challenges. The development of resistance to commonly used insecticides further complicates the control of these vectors. Although some novel strategies for A. aegypti control discussed above such as RIDL offer promise, they remain in the early stages of field testing [183]. Interim measures including the application of persistent insecticides to the interior of houses may be needed in some situations to reduce CHIKV and DENV transmission.
The long-term prospects for CHIKV maintenance in the endemic, urban cycle are not entirely clear. Following introduction of the Asian lineage into South and Southeast Asia during the 1950s or earlier, it became extinct in India after 1973 for unknown reasons, but continues to circulate in Southeast Asia today [167,184]. The only major human immune cross-reactivity known to affect CHIKV to a major extent is that from Ross River and Mayaro viruses, which like CHIKV are members of the Semliki Forest complex of alphaviruses. Ross River virus only occurs in Australia and Indonesia, and Mayaro virus in South America where seroprevalence is generally low in urban settings. These data suggest that CHIKV will continue to circulate indefinitely in Asia and probably in the Americas as well.

{kind=link}
{kind=link}
Table 1.
Chikungunya virus vector competence in arthropods. Infection was ascertained by detection of virus in bodies of bloodfed mosquitoes; dissemination was determined by detection of virus in legs or heads. Transmission was verified by detection of virus in saliva or by infection of vertebrates after re-feeding. CHIKV bloodmeal titers are expressed in in log10 cell culture infectious dose50/mosquito, plaque forming units/mL, or suckling mouse infectious culture lethal dose50/mL. Mosquitoes were held at 28 °C during the incubation period, except where noted: * indicates incubation at 24 °C and ** denotes incubation at 16 °C. For simplification, cohorts of the same species that fed on the same strain at similar bloodmeal titers are represented as one value and bloodmeal titers are shown as ranges.
| Mosquito Species | Source | Generation Number | Source of CHIKV, Isolation Year | Strain Name | Bloodmeal Titer | Incubation Period (Days) | % Infected (n) | % Disseminated (n) | % Transmitted (n) | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
| Aedes aegypti | Queensland, Australia | 1 | patient in Melbourne ex. Mauritius, March 2006 | not stated | 4 | 14-15 | 92 (23/25) | 92 (23/25) | 64 (16/25) | van den Hurk et al., 2010 [128] |
| New Caledonia | 1 | patient in New Caledonia ex. Indonesia, February 2011 | NC 2011-568 | 7 | 14 | n.d. | n.d. | 27 (10/37) | Dupont-Rouzeyrol et al., 2012 [41] | |
| patient in Reunion Island, 2006 | 6.21 | n.d. | n.d. | 75 (27/36) | ||||||
| Mayotte | 3 | patient in Reunion Island, 2005 (E1226V) | 6.21 | 7.5 | 14 | n.d. | 87 (54/62) | n.d. | Martin et al., 2010 [94] | |
| patient in Reunion Island, 2005 (E1226A) | 6.115 | n.d. | 78 (43/55) | n.d. | ||||||
| Cameroon | 1 | patient in Reunion Island, 2005 (E1226V) | 06.21 | 7 | 14 | n.d. | 89 (333/376) | n.d. | Paupy et al., 2010 [185] | |
| patient in Reunion Island, 2005 (E1226A) | 06.115 | n.d. | 97 (36/37) | n.d. | Vazeille et al., 2007 [10] | |||||
| patient in Reunion Island, 2005 (E1226V) | 06.21 | n.d | 65 (70/107) | n.d. | ||||||
| patient in Mayotte, 2006 | 06.111 | n.d. | 82 (68/82) | n.d. | ||||||
| patient in Democratic Republic of Congo, 2000 | 06.117 | n.d | 84 (56/66) | n.d. | ||||||
| Dakar, Senegal | 4 | patient in Thailand, 1962 | 15561 | 4.2–4.6 | 7 | 2 (1/45) | 0 (0/45) | n.d. | Turell et al., 1992 [186] | |
| Lagos, Nigeria | colony | patient in Calcutta, India, 1963 | 63-266 | 8 | 14 | 10 (2/20) | n.d. | 50 (1/2) | Shah et al., 1964 [187] | |
| French West Indies | 1-2 | patient in Reunion Island, 2006 | 06.21 | 7.5 | 14 | 98 (900/918) | n.d. | n.d. | Girod et al., 2011 [188] | |
| patient in Reunion Island, 2006 | 06.21 | 6 | 7 | 47 (301/634) | n.d. | n.d. | ||||
| Trinidad, West Indies | colony | patient in Calcutta, India, 1963 | 63-266 | 8 | 14 | 42 (3/7) | n.d. | 100 (1/1) | Shah et al., 1964 | |
| Aedes aegypti | Guadeloupe | 1 | patient in Reunion Island, 2005 (E1226V) | 6.21 | 7.5 | 14 | n.d. | 96 (346/358) | n.d. | Girod et al., 2011 |
| Martinique | 1 | patient in Reunion Island, 2005 (E1226V) | 6.21 | 7.5 | 14 | n.d. | 98 (285/290) | n.d. | Girod et al., 2011 | |
| French Guyana | 1 | patient in Reunion Island, 2005 (E1226V) | 6.21 | 7.5 | 14 | n.d. | 99 (269/270) | n.d. | Girod et al., 2011 | |
| Florida, USA | 1 | patient in France ex. Reunion Island, 2006 | LR2006-OPY1 | 6.1 | 6 | 58 (15/26) | 73 (11/15) | n.d. | Pesko et al., 2009 [93] | |
| patients in Reunion Island, 2005 | 06.21 and/or 06.115 | 7.5 | 10 | n.d. | 100 (48/48) | n.d. | Vega-Rua et al., 2014 [43] | |||
| Louisiana, USA | 4-5 | patient in Thailand, 1962 | 15561 | 4.2–4.6 | 7 | 5 (3/60) | 2 (1/60) | n.d. | Turell et al., 1992 | |
| Indiana, USA | 7 | patient in Thailand, 1962 | 15561 | 4.2–4.6 | 7 | 6 (2/35) | 6 (2/35) | n.d. | Turell et al., 1992 | |
| Puerto Rico, USA | 3 | patient in Thailand, 1962 | 15561 | 4.2–4.6 | 7 | 55 (9/55) | 11 (6/55) | n.d. | Turell et al., 1992 | |
| 5 | patient in Thailand, 1962 | 15561 | 5.3 | 7 | 33 (10/30) | 10 (3/30) | n.d. | |||
| Mexico | 1 | patients in Reunion Island, 2005 | 06.21 and/or 06.115 | 7.5 | 10 | n.d. | 97 (58/60) | n.d. | Vega-Rua et al., 2014 | |
| Panama | 1 | patients in Reunion Island, 2005 | 06.21 and/or 06.115 | 7.5 | 10 | n.d. | 97 (58/60) | n.d. | Vega-Rua et al., 2014 | |
| patient in New Caledonia, 2011 | NC/2011-568 | n.d. | 100 (30/30) | n.d. | ||||||
| Venezuela | 1 | patients in Reunion Island, 2005 | 06.21 and/or 06.115 | 7.5 | 10 | n.d. | 100 (51/51) | n.d. | Vega-Rua et al., 2014 | |
| Peru | 1 | patients in Reunion Island, 2005 | 06.21 and/or 06.115 | 7.5 | 10 | n.d. | 100 (89/89) | n.d. | Vega-Rua et al., 2014 | |
| Brazil | 1 | patients in Reunion Island, 2005 | 06.21 and/or 06.115 | 7.5 | 10 | n.d. | 98 (128/130) | n.d. | Vega-Rua et al., 2014 | |
| patient in New Caledonia, 2011 | NC/2011-568 | n.d. | 95 (57/60) | n.d. | Vega-Rua et al., 2014 | |||||
| Bolivia | 1 | patients in Reunion Island, 2005 | 06.21 and/or 06.115 | 7.5 | 10 | n.d. | 100 (60/60) | n.d. | Vega-Rua et al., 2014 | |
| Aedes aegypti | Paraguay | 1 | patients in Reunion Island, 2005 | 06.21 and/or 06.115 | 7.5 | 10 | n.d. | 99 (89/90) | n.d. | Vega-Rua et al., 2014 |
| Uruguay | 1 | patients in Reunion Island, 2005 | 06.21 and/or 06.115 | 7.5 | 10 | n.d. | 100 (60/60) | n.d. | Vega-Rua et al., 2014 | |
| Argentina | 1 | patients in Reunion Island, 2005 | 06.21 and/or 06.115 | 7.5 | 10 | n.d. | 99 (119/120) | n.d. | Vega-Rua et al., 2014 | |
| Bangkok, Thailand | 2 | patient in Thailand, 1962 | 15561 | 5.3 | 7 | 66 (20/30) | 66 (20/30) | n.d. | Turell et al., 1992 | |
| Ho Chi Minh City, Vietnam | colony | patient in Reunion Island, 2005 (E1226A) | 06.115 | 7 | 14 | n.d. | 66 (135/206) | n.d. | Vazeille et al., 2007 | |
| patient in Reunion Island, 2005 (E1226V) | 06.21 | n.d | 97 (227/234) | n.d. | ||||||
| patient in Mayotte, 2006 | 06.111 | n.d. | 92 (126/137) | n.d. | ||||||
| patient in Democratic Republic of Congo, 2000 | 06.117 | n.d | 78 (108/138) | n.d. | ||||||
| Higgs variant, Rexville D | colony | clone derived from patient in France ex. Reunion Island, 2006 E1226A | LR2006-OPY1 E1A226 | 5 | 7 | 20 (not stated) | n.d. | n.d. | Tsetsarkin et al., 2007 [9] | |
| clone derived from patient in France ex. Reunion Island, 2006 E1226V | LR2006-OPY1 E1V226 | 5 (not stated) | n.d. | n.d. | ||||||
| infectious clone from patient in West Africa, E1226A | 37997 E1A226 | 20 (not stated) | n.d. | n.d. | ||||||
| infectious clone from patient in West Africa, E1226V | 37997 E1V226 | 10 (not stated) | n.d. | n.d. | ||||||
| Aedes furcifer, Kadougou, Senegal, 1983 | 37997 | 8 | 14 | 100 (7/7) | 63 (5/8) | n.d. | Vanlandingham et al., 2005 [189] | |||
| not stated | colony | patient in Africa, not stated | not stated | 8.6-9.2 | 14 | 53 (24/45) | n.d. | 44 (20/45) | Mangiafico, 1971 [190] | |
| Rockefeller | colony | patient in Thailand, 1962 | 15561 | 4.2-4.6 | 7 | 18 (9/50) | 12 (6/50) | n.d. | Turell et al., 1992 | |
| Madeira Island, Spain | 1 | patient in Reunion Island, 2005 (E1226V) | 06.21 | 7 | 14 | n.d. | 100 (27/27) | 40 (4/10) | Vazeille et al., 2012 [191] | |
| Aedes albopictus | Queensland, Australia | 7 | patient in Melbourne ex. Mauritius, March 2006 | not stated | 3.9 | 14-15 | 92 (23/25) | 92 (23/25) | 32 (8/25) | van den Hurk et al., 2010 |
| Torres Strait, Australia | 7 | patient in Melbourne ex. Mauritius, March 2006 | not stated | 8 | 14 | 80 (4/5) | 80 (4/5) | 60 (3/5) | Nicholson et al., 2014 [192] | |
| Mauritius | 1 | patient in India, 1973 | Barsi, P0-731460 | 5.8 | 8-9 | 32 (13/41) | n.d. | n.d. | Tesh et al., 1976 [193] | |
| patient in Tanzania, 1953 | Ross, S-27 | 6.8 | 8-9 | 19 (13/67) | n.d. | n.d. | Tesh et al., 1976 | |||
| Mayotte | 1 | patient in Reunion Island, 2005 (E1226A) | 6.115 | 7 | 14 | n.d | 25 (115/462) | n.d. | Vazeille et al., 2007 | |
| 6 | patient in Reunion Island, 2005 (E1226A) | 6.115 | 7.5 | 14 | n.d. | 79 (45/57) | n.d. | Martin et al., 2010 [94] | ||
| 1 | patient in Reunion Island, 2005 (E1226V) | 6.21 | 7 | 14 | n.d | 91 (296/325) | n.d. | Vazeille et al., 2007 | ||
| 6 | patient in Reunion Island, 2005 (E1226V) | 6.21 | 7.5 | 14 | n.d. | 99 (64/65) | n.d. | Martin et al., 2010 | ||
| 1 | patient in Mayotte, 2006 | 06.111 | 7 | 14 | n.d. | 98 (48/49) | n.d. | Vazeille et al., 2007 | ||
| 1 | patient in Democratic Republic of Congo, 2000 | 06.117 | 7 | 14 | n.d | 73 (41/56) | n.d. | Vazeille et al., 2007 | ||
| Reunion Island | 2 | patient in Reunion Island, 2005 (E1226A) | 6.115 | 7.5 | 14 | n.d. | 90 (26/29) | n.d. | Martin et al., 2010 | |
| 1 | patient in Reunion Island, 2005 (E1226A) | 6.115 | 7 | 14 | n.d. | 25 (114/462) | n.d. | Vazeille et al., 2007 | ||
| 2 | patient in Reunion Island, 2005 (E1226V) | 6.21 | 7.5 | 14 | n.d. | 98 (55/56) | n.d. | Martin et al., 2010 | ||
| 1 | patient in Reunion Island, 2005 (E1226V) | 6.21 | 7 | 14 | n.d | 96 (391/409) | n.d. | Vazeille et al., 2007 | ||
| 2 | patient in Mayotte, 2006 | 6.111 | 7 | 14 | n.d. | 97 (91/94) | n.d. | Vazeille et al., 2007 | ||
| 2 | patient in Democratic Republic of Congo, 2000 | 6.117 | 7 | 14 | n.d | 80 (25/31) | n.d. | Vazeille et al., 2007 | ||
| Madagascar | 7 | patient in Thailand, 1962 | 15561 | 5.3 | 7 | 95 (19/20) | 35 (7/20) | n.d. | Turell et al., 1992 | |
| 1 | patient in India, 1973 | Barsi, P0-731460 | 6.2 | 8–9 | 87 (33/38) | n.d. | n.d. | Tesh et al., 1976 | ||
| 1 | patient in Tanzania, 1953 | Ross, S-27 | 6.8 | 8–9 | 39 (19/49) | n.d. | n.d. | Tesh et al., 1976 | ||
| 2-5 | patient in Reunion Island, 2006 | not stated | 7.5 | 14 | n.d. | 98 (497/503) | n.d. | Raharimalala et al., 2012 [76] | ||
| Aedes albopictus | Cameroon | 1 | patient in Reunion Island, 2006 | 06.21 | 7 | 14 | n.d. | 85 (187/218) | n.d. | Paupy et al., 2010 |
| patient in Reunion Island, 2005 (E1226V) | 06.21 | 7 | 14 | n.d | 68 (41/60) | n.d. | Vazeille et al., 2007 | |||
| patient in Mayotte, 2006 | 06.111 | 7 | 14 | n.d. | 44 (34/44) | n.d. | Vazeille et al., 2007 | |||
| patient in Democratic Republic of Congo, 2000 | 06.117 | 7 | 14 | n.d | 56 (22/39) | n.d. | Vazeille et al., 2007 | |||
| patient in Reunion Island, 2005 (E1226A) | 06.115 | 7 | 14 | n.d | 12 (11/90) | n.d. | Vazeille et al., 2007 | |||
| Virginia and Georgia, USA | 1 | mosquito pool, Comoros, 2005 strain COM125 | COM125 | 4.9 | 7 | 73 (83/114) | n.d. | 40 (33/83) | McTighe & Vaidyanathan, 2012 | |
| Hawaii, USA | 1 | patient in Tanzania, 1953 | Ross, S-27 | 7-7.5 | 8-9 | 69 (76/110) | n.d. | n.d. | Tesh et al., 1976 | |
| 1 | patient in India, 1973 | Barsi, P0-731460 | 5.8 | 8-9 | 97 (120/124) | n.d. | n.d. | Tesh et al., 1976 | ||
| colony | patient in Calcutta, India, 1963 | 63-266 | 8 | 14 | 100 (32/32) | n.d. | 34 (8/22) | Shah et al., 1964 | ||
| colony | patient in Thailand, 1962 | 15561 | 5.3 | 7 | 93 (28/30) | 60 (18/30) | n.d. | Turell et al., 1992 | ||
| Florida, USA | 1 | patient in France ex. Reunion Island, 2006 | LR2006-OPY1 | 6.1 | 6 | 100 (22/22) | 91 (20/22) | n.d. | Pesko et al., 2009 | |
| patients in Reunion Island, 2005 | 06.21 and/or 06.115 | 7.5 | 10 | n.d. | 83 (50/60) | n.d. | Vega-Rua et al., 2014 | |||
| 2 | patient in Thailand, 1962 | 15561 | 5.3 | 7 | 97 (29/30) | 37 (11/30) | n.d. | Turell et al., 1992 | ||
| Missouri, USA | 1 | patients in Reunion Island, 2005 | 06.21 and/or 06.115 | 7.5 | 10 | n.d. | 90 (54/60) | n.d. | Vega-Rua et al., 2014 | |
| Aedes albopictus | Texas, USA | colony | clone derived from patient in France ex. Reunion Island, 2006 E1226A | LR2006-OPY1 E1A226 | 5 | 7 | 31 (61/194) | 30 (not stated) | Tsetsarkin et al., 2007 | |
| clone derived from patient in France ex. Reunion Island, 2006 E1226V | LR2006-OPY1 E1V226 | 5 | 7 | 90 (241/269) | 65 (not stated) | Tsetsarkin et al., 2007 | ||||
| infectious clone from patient in West Africa, E1226A | 37997 E1A226 | 5 | 7 | 37 (97/226) | n.d. | n.d. | Tsetsarkin et al., 2007 | |||
| infectious clone from patient in West Africa, E1226V | 37997 E1V226 | 5 | 7 | 92 (253/274) | n.d. | n.d. | Tsetsarkin et al., 2007 | |||
| 9-10 | patient in Thailand, 1962 | 15561 | 5.3 | 7 | 75 (15/20) | 35 (7/20) | n.d. | Turell et al., 1992 | ||
| Louisiana, USA | 4-5 | patient in Thailand, 1962 | 15561 | 5.3 | 7 | 97 (29/30) | 80 (24/30) | n.d. | Turell et al., 1992 | |
| Mexico | 1 | patients in Reunion Island, 2005 | 06.21 and/or 06.115 | 7.5 | 10 | n.d. | 70 (42/60) | n.d. | Vega-Rua et al., 2014 | |
| Panama | 1 | patients in Reunion Island, 2005 | 06.21 and/or 06.115 | 7.5 | 10 | n.d. | 95 (57/60) | n.d. | Vega-Rua et al., 2014 | |
| patient in New Caledonia, 2011 | NC/2011-568 | 7.5 | 10 | n.d. | 97 (29/30) | n.d. | Vega-Rua et al., 2014 | |||
| Brazil | 6-7 | patient in Thailand, 1962 | 15561 | 5.3 | 7 | 73 (22/30) | 50 (15/30) | n.d. | Turell et al., 1992 | |
| 1 | patients in Reunion Island, 2005 | 06.21 and/or 06.115 | 7.5 | 10 | n.d. | 94 (301/320) | n.d. | Vega-Rua et al., 2014 | ||
| Argentina | 1 | patients in Reunion Island, 2005 | 06.21 and/or 06.115 | 7.5 | 10 | n.d. | 63 (35/56) | n.d. | Vega-Rua et al., 2014 | |
| patient in New Caledonia, 2011 | 06.21 and/or 06.115 | 7.5 | 10 | n.d. | 93 (28/30) | n.d. | Vega-Rua et al., 2014 | |||
| Israel | 1 | patient in Tanzania, 1953 | Ross, S-27 | 7.2 | 8-9 | 30 (14/47) | n.d. | n.d. | Tesh et al., 1976 | |
| patient in India, 1973 | Barsi, P0-731460 | 5.8 | 8-9 | 67 (37/55) | n.d. | n.d. | Tesh et al., 1976 | |||
| Lebanon | 1 | patient in Reunion Island, 2005 (E1226V) | 6.21 | 8 | 14 | n.d. | 29 (12/42) | n.d. | Haddad et al., 2012 [194] | |
| Aedes albopictus | Italy | 0 | patient in Reunion Island, 2005 (E1226V) | 06.21 | 7 | 14 | n.d. | 83 (52/63) | n.d. | Talbalaghi et al., 2010 [195] |
| France | 0 | patient in Reunion Island, 2005 (E1226V) | 6.21 | 7 | 14 | n.d. | 77.1 (27/35) | n.d. | Vazeille et al., 2008 [196] | |
| 13 | patient in France, 2010 (E1226A) | 1909 | 7.3 | 14 | n.d. | 96 (21/22) | 14 (3/21) | Vega-Rua et al., 2013 [36] | ||
| 13 | patient in France, 2010 (E1226A) | 1630 | 7.3 | 14 | n.d. | 90 (17/19) | 12 (2/17) | Vega-Rua et al., 2013 | ||
| Corsica, France | 0 | patient in Reunion Island, 2005 (E1226V) | 6.21 | 7.5 | 14 | n.d. | 94 (377/401) | n.d. | Moutailler et al., 2009 [197] | |
| Indonesia | 1 | patient in Tanzania, 1953 | Ross, S-27 | 7.1 | 8–9 | 64 34/53) | n.d. | n.d. | Tesh et al., 1976 | |
| Philippines | 1 | patient in Tanzania, 1953 | Ross, S-27 | 7.2 | 8–9 | 55 (21/38) | n.d. | n.d. | Tesh et al., 1976 | |
| India | 1 | patient in Tanzania, 1953 | Ross, S-27 | 7–7.2 | 8–9 | 38 (30/79) | n.d. | n.d. | Tesh et al., 1976 | |
| patient in India, 1973 | Barsi, P0-731460 | 5.7–5.9 | 8–9 | 71 (74/104) | n.d. | n.d. | Tesh et al., 1976 | |||
| Vietnam | 1 | patient in Tanzania, 1953 | Ross, S-27 | 7.7 | 8–9 | 44 (29/66) | n.d. | n.d. | Tesh et al., 1976 | |
| patient in India, 1978 | Barsi, P0-731460 | 5.7 | 8–9 | 49 (37/94) | n.d. | n.d. | Tesh et al., 1976 | |||
| Thailand | 1 | patient in Tanzania, 1953 | Ross, S-27 | 7.4 | 8–9 | 38 (12/32) | n.d. | n.d. | Tesh et al., 1976 | |
| patient in India, 1973 | Barsi, P0-731460 | 6 | 8–9 | 73 (24/33) | n.d. | n.d. | Tesh et al., 1976 | |||
| Malaysia | 1 | patient in Tanzania, 1953 | Ross, S-27 | 6.9 | 8–9 | 29 (15/51) | n.d. | n.d. | Tesh et al., 1976 | |
| patient in India, 1973 | Barsi, P0-731460 | 6 | 8–9 | 42 (27/64) | n.d. | n.d. | Tesh et al., 1976 | |||
| Taipei | 1 | patient in Tanzania, 1953 | Ross, S-27 | 7 | 8–9 | 25 (12/48) | n.d. | n.d. | Tesh et al., 1976 | |
| patient in India, 1973 | Barsi, P0-731460 | 5.8 | 8–9 | 28 (14/50) | n.d. | n.d. | Tesh et al., 1976 | |||
| Taiwan | 2 | patient in Thailand, 1962 | 15561 | 5.3 | 7 | 90 (27/30) | 20 (6/30) | n.d. | Turell et al., 1992 | |
| Okinawa, Japan | 5 | patient in Thailand, 1962 | 15561 | 5.3 | 7 | 72 (18/25) | 28 (7/25) | n.d. | Turell et al., 1992 | |
| Tokyo, Japan | 5 | patient in Thailand, 1962 | 15561 | 4.2–4.6 | 7 | 50 (14/28) | 14 (7/50 | n.d. | Turell et al., 1992 | |
| Jakarta | 1 | patient in India, 1975 | Barsi, P0-731460 | 5.8 | 8–9 | 70 (23/33) | n.d. | n.d. | Tesh et al., 1976 | |
| Philippines | 1 | patient in India, 1976 | Barsi, P0-731460 | 6 | 8–9 | 87 (33/38) | n.d. | n.d. | Tesh et al., 1976 | |
| Sabah, Malaysia | 5 | patient in Thailand, 1962 | 15561 | 4.2–4.6 | 7 | 43 (15/35) | 6 (2/43) | n.d. | Turell et al., 1992 | |
| Aedes albopictus | Hanoi, Vietnam | 3 | patient in Reunion Island, 2005 (E1226A) | 6.115 | 7 | 14 | n.d | 30 (16/54) | n.d. | Vazeille et al., 2007 |
| patient in Reunion Island, 2005 (E1226V) | 6.21 | 7 | 14 | n.d | 84 (105/126) | n.d. | Vazeille et al., 2007 | |||
| patient in Mayotte, 2006 | 6.111 | 7 | 14 | n.d. | 84 (105/126) | n.d. | Vazeille et al., 2007 | |||
| patient in Democratic Republic of Congo, 2000 | 6.117 | 7 | 14 | n.d | 47 (56/119) | n.d. | Vazeille et al., 2007 | |||
| not stated | colony | patient in Africa, not stated | not stated | 8.6–9.2 | 14 | 100 (50/50) | n.d. | 80 (40/50) | Mangiafico, 1971 | |
| Aedes antipodeus | North Auckland, New Zealand | 1 | patient in India | 91064A | 7.8 | 21** | 100 (15/15) | 73 (11/15) | 0 (0/15) | Kramer et al., 2011 [198] |
| Aedes caspius | France | 0 | patient in Reunion Island, 2005 (E1226V) | 06.21 | 7 | 14 | n.d. | 25 (4/16) | n.d. | Vazeille et al., 2008 |
| Aedes detritus | France | 0 | patient in Reunion Island, 2005 (E1226V) | 06.21 | 7 | 14 | n.d. | 67.3 (33/49) | n.d. | Vazeille et al., 2008 |
| Aedes fulgens | South Africa | 1 | not stated | H817 | 5.7 | 9–12 | 88 (29/33) | n.d. | 10 (3/29) | Jupp et al., 1981 [199] |
| Aedes furcifer | South Africa | colony or 1-4 | not stated | H817 | 5.7–6.9 | 8–29 | 71 (192/271) | n.d. | 30 (8/27) | Jupp et al., 1981 |
| Aedes hensilli | Micronesia | 12-15 | Mosquito in Comoros, 2005 | COM 125 | 5.7 | 8 | 63 (20/32) | 80 (16/20) | n.d. | Ledermann et al., 2014 [200] |
| Aedes notoscriptus | Auckland, New Zealand | 1 | patient in India | 91064A | 10.5 | 14* | 36 (8/32) | 75 (6/8) | 0 (0/8) | Kramer et al., 2011 |
| Aedes polynesiensis | Samoa | colony | patient in Calcutta, India, 1963 | 63-266 | 8 | 14 | 40 (4/10) | n.d. | n.d. | Shah et al., 1964 |
| Aedes togoi | not stated | colony | patient in Africa, not stated | not stated | 8.6–9.2 | 14 | 97 (42/43) | n.d. | 12 (5/43) | Mangiafico, 1971 |
| Aedes triseriatus | not stated | colony | patient in Africa, not stated | not stated | 8.6–9.2 | 14 | 100 (50/50) | n.d. | 84 (42/50) | Mangiafico, 1971 |
| Aedes vexans | Italy | 0 | patient in Reunion Island, 2005 (E1226V) | 06.21 | 7 | 14 | n.d. | 8 (2/26) | n.d. | Talbalaghi et al., 2010 |
| France | patient in Reunion Island, 2005 (E1226V) | 06.21 | 7 | 14 | n.d. | 0 (0/13) | n.d. | Vazeille et al., 2008 | ||
| Aedes vittatus | Senegal | 1 | mosquitoes, bats or humans in Senegal, 1962, '79 and '05 | ArD30237, CS13-288 or HD 180738 | 6 | 10 | 89 (41/46) | 54 (22/41) | 18 (4/22) | Diagne et al., 2014 [201] |
| 6-7 | 10 | 19 (19/98) | 37 (7/19) | 43 (3/7) | Diagne et al., 2014 | |||||
| Anopheles gambiae | G3 | colony | Aedes furcifer, Kadoug ou, Senegal, 1983 | 37997 | 8 | 14 | 0 (0/8) | 0 (0/8) | n.d. | Vanlandingham et al., 2005 |
| Anopheles maculipennis | Italy | 0 | patient in Reunion Island, 2005 (E1226V) | 06.21 | 7 | 14 | n.d. | 0 (0/10) | n.d. | Talbalaghi et al., 2010 |
| Culex fatigans | Philippines | colony | patient in Calcutta, India, 1963 | 63-266 | 8 | 14 | 0 (0/10) | n.d. | n.d. | Shah et al., 1964 |
| Culex horridus | South Africa | 1 | not stated | H817 | 4.6–5.4 | 14–25 | 6 (1/17) | n.d. | n.d. | Jupp et al., 1981 |
| Culex pipiens | Italy | 0 | patient in Reunion Island, 2005 (E1226V) | 06.21 | 7 | 14 | n.d. | 0 (0/45) | n.d. | Talbalaghi et al., 2010 |
| France | patient in Reunion Island, 2005 (E1226V) | 06.21 | 7 | 14 | n.d. | 0 (0/11) | n.d. | Vazeille et al., 2008 | ||
| Culex quinquefasciatus | Zimbabwe | 1 | not stated | H817 | 5.3 | 20–22 | 0 (0/19) | n.d. | 0 (0/19) | Jupp et al., 1981 |
| Eretmapodites chrysogaster | not stated | colony | patient in Africa, not stated | not stated | 8.6–9.2 | 14 | 80 (40/50) | n.d. | 36 (18/50) | Mangiafico, 1971 |
| Mansonia africana | Mozambique | 0 | not stated | H817 | 4.7–5.2 | 8–15 | 34 (23/67) | n.d. | n.d. | Jupp et al., 1981 |
| Opifex fuscus | Wellington, New Zealand | 1 | patient in India | 91064A | 6.2 | 11* | 98 (46/47) | 100 (46/46) | 100 (46/46) | Kramer et al., 2011 |
| Ornithodoros savignyi | South Africa | 0 | not stated | H817 | 6.6 | 50-61 | 0 (0/11) | n.d. | n.d. | Jupp et al., 1981 |
6. Conclusions
Chikungunya virus has caused explosive outbreaks of severe, debilitating and often chronic arthralgia since it emerged in the 1950s and later in 2004 from the enzootic ECSA lineage in Africa. The large populations of susceptible humans in many naïve regions and thriving populations of the two urban vectors, A. aegypti and A. albopictus, will probably facilitate endemicity throughout most regions of the tropics and subtropics for the foreseeable future, although the burden of disease is typically difficult to estimate because CHIKV and DENV infections are difficult to distinguish clinically. Further adaptation of CHIKV to these urban vectors is also suggested by recent findings [13]. Future emergences from enzootic African cycles will also remain a risk that is increasing with more and more air travel and international commerce. Although novel strategies to control vector populations and reduce transmission are in early stages of field testing, the deployment of one of several highly promising human vaccines probably offers the best hope for making a major impact in restricting CHIKV circulation and preventing human disease [184].
Conflicts of Interest
The authors declare no conflict of interest.
References and Notes
- Powers, A.M.; Logue, C.H. Changing patterns of chikungunya virus: Re-emergence of a zoonotic arbovirus. J. Gen. Virol. 2007, 88, 2363–2377. [Google Scholar] [PubMed]
- Kuehn, B.M. Chikungunya virus transmission found in the United States: US health authorities brace for wider spread. JAMA 2014, 312, 776–777. [Google Scholar] [PubMed]
- CDC. Chikungunya virus in the United States. Available online: http://www.cdc.gov/chikungunya/geo/united-states.html (accessed on 8 November 2014).
- Jupp, P.G.; McIntosh, B.M. Aedes furcifer and other mosquitoes as vectors of chikungunya virus at Mica, Northeastern Transvaal, South Africa. J. Am. Mosq. Control. Assoc. 1990, 6, 415–420. [Google Scholar] [PubMed]
- Volk, S.M.; Chen, R.; Tsetsarkin, K.A.; Adams, A.P.; Garcia, T.I.; Sall, A.A.; Nasar, F.; Schuh, A.J.; Holmes, E.C.; Higgs, S.; et al. Genome-scale phylogenetic analyses of chikungunya virus reveal independent emergences of recent epidemics and various evolutionary rates. J. Virol. 2010, 84, 6497–6504. [Google Scholar] [CrossRef] [PubMed]
- Tsetsarkin, K.A.; Chen, R.; Sherman, M.B.; Weaver, S.C. Chikungunya virus: Evolution and genetic determinants of emergence. Curr. Opin. Virol. 2011, 1, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Kariuki Njenga, M.; Nderitu, L.; Ledermann, J.P.; Ndirangu, A.; Logue, C.H.; Kelly, C.H.; Sang, R.; Sergon, K.; Breiman, R.; Powers, A.M. Tracking epidemic chikungunya virus into the Indian Ocean from East Africa. J. Gen. Virol. 2008, 89, 2754–2760. [Google Scholar]
- Schuffenecker, I.; Iteman, I.; Michault, A.; Murri, S.; Frangeul, L.; Vaney, M.C.; Lavenir, R.; Pardigon, N.; Reynes, J.M.; Pettinelli, F.; et al. Genome microevolution of chikungunya viruses causing the Indian Ocean outbreak. PLoS Med. 2006, 3, e263. [Google Scholar] [CrossRef] [PubMed]
- Tsetsarkin, K.A.; Vanlandingham, D.L.; McGee, C.E.; Higgs, S. A single mutation in chikungunya virus affects vector specificity and epidemic potential. PLoS Pathog. 2007, 3, e201. [Google Scholar] [CrossRef] [PubMed]
- Vazeille, M.; Moutailler, S.; Coudrier, D.; Rousseaux, C.; Khun, H.; Huerre, M.; Thiria, J.; Dehecq, J.S.; Fontenille, D.; Schuffenecker, I.; et al. Two chikungunya isolates from the outbreak of La Reunion (Indian Ocean) exhibit different patterns of infection in the mosquito, Aedes albopictus. PLoS One 2007, 2, e1168. [Google Scholar] [CrossRef] [PubMed]
- Tsetsarkin, K.A.; McGee, C.E.; Volk, S.M.; Vanlandingham, D.L.; Weaver, S.C.; Higgs, S. Epistatic roles of E2 glycoprotein mutations in adaption of chikungunya virus to Aedes albopictus and Ae. aegypti mosquitoes. PLoS One 2009, 4, e6835. [Google Scholar] [CrossRef] [PubMed]
- Arias-Goeta, C.; Mousson, L.; Rougeon, F.; Failloux, A.B. Dissemination and transmission of the E1–226V variant of chikungunya virus in Aedes albopictus are controlled at the midgut barrier level. PLoS One 2013, 8, e57548. [Google Scholar] [CrossRef] [PubMed]
- Tsetsarkin, K.A.; Chen, R.; Yun, R.; Rossi, S.L.; Plante, K.S.; Guerbois, M.; Forrester, N.; Perng, G.C.; Sreekumar, E.; Leal, G.; et al. Multi-peaked adaptive landscape for chikungunya virus evolution predicts continued fitness optimization in Aedes albopictus mosquitoes. Nat. Commun. 2014, 5, 4084. [Google Scholar] [CrossRef] [PubMed]
- Ratsitorahina, M.; Harisoa, J.; Ratovonjato, J.; Biacabe, S.; Reynes, J.M.; Zeller, H.; Raoelina, Y.; Talarmin, A.; Richard, V.; Louis Soares, J. Outbreak of dengue and chikungunya fevers, toamasina, Madagascar, 2006. Emerg. Infect. Dis. 2008, 14, 1135–1137. [Google Scholar] [CrossRef] [PubMed]
- De Lamballerie, X.; Leroy, E.; Charrel, R.N.; Ttsetsarkin, K.; Higgs, S.; Gould, E.A. Chikungunya virus adapts to tiger mosquito via evolutionary convergence: A sign of things to come? Virol. J. 2008, 5, 33. [Google Scholar] [CrossRef] [PubMed]
- Bagny, L.; Delatte, H.; Quilici, S.; Fontenille, D. Progressive decrease in Aedes aegypti distribution in reunion island since the 1900s. J. Med. Entomol. 2009, 46, 1541–1545. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.P.; Joseph, R.; Kamaraj, T.; Jambulingam, P. A226V mutation in virus during the 2007 chikungunya outbreak in Kerala, India. J. Gen. Virol. 2008, 89, 1945–1948. [Google Scholar] [CrossRef] [PubMed]
- Yergolkar, P.N.; Tandale, B.V.; Arankalle, V.A.; Sathe, P.S.; Sudeep, A.B.; Gandhe, S.S.; Gokhle, M.D.; Jacob, G.P.; Hundekar, S.L.; Mishra, A.C. Chikungunya outbreaks caused by African genotype, India. Emerg. Infect. Dis. 2006, 12, 1580–1583. [Google Scholar] [CrossRef] [PubMed]
- Kularatne, S.A.; Gihan, M.C.; Weerasinghe, S.C.; Gunasena, S. Concurrent outbreaks of chikungunya and dengue fever in Kandy, Sri Lanka, 2006–07: A comparative analysis of clinical and laboratory features. Postgrad. Med. J. 2009, 85, 342–346. [Google Scholar] [CrossRef] [PubMed]
- Peyrefitte, C.N.; Rousset, D.; Pastorino, B.A.; Pouillot, R.; Bessaud, M.; Tock, F.; Mansaray, H.; Merle, O.L.; Pascual, A.M.; Paupy, C.; et al. Chikungunya virus, Cameroon, 2006. Emerg. Infect. Dis. 2007, 13, 768–771. [Google Scholar] [CrossRef] [PubMed]
- Peyrefitte, C.N.; Bessaud, M.; Pastorino, B.A.; Gravier, P.; Plumet, S.; Merle, O.L.; Moltini, I.; Coppin, E.; Tock, F.; Daries, W.; et al. Circulation of chikungunya virus in Gabon, 2006–2007. J. Med. Virol. 2008, 80, 430–433. [Google Scholar] [CrossRef] [PubMed]
- Paupy, C.; Kassa Kassa, F.; Caron, M.; Nkoghe, D.; Leroy, E.M. A chikungunya outbreak associated with the vector Aedes albopictus in remote villages of Gabon. Vector Borne Zoonotic Dis. 2012, 12, 167–169. [Google Scholar] [CrossRef] [PubMed]
- Mombouli, J.V.; Bitsindou, P.; Elion, D.O.; Grolla, A.; Feldmann, H.; Niama, F.R.; Parra, H.J.; Munster, V.J. Chikungunya virus infection, Brazzaville, Republic of Congo, 2011. Emerg. Infect. Dis. 2013, 19, 1542–1543. [Google Scholar] [CrossRef] [PubMed]
- Rezza, G.; Nicoletti, L.; Angelini, R.; Romi, R.; Finarelli, A.C.; Panning, M.; Cordioli, P.; Fortuna, C.; Boros, S.; Magurano, F.; et al. Infection with chikungunya virus in Italy: An outbreak in a temperate region. Lancet 2007, 370, 1840–1846. [Google Scholar] [CrossRef] [PubMed]
- Sam, I.C.; Chan, Y.F.; Chan, S.Y.; Loong, S.K.; Chin, H.K.; Hooi, P.S.; Ganeswrie, R.; Abubakar, S. Chikungunya virus of Asian and Central/East African genotypes in Malaysia. J. Clin. Virol. 2009, 46, 180–183. [Google Scholar] [CrossRef] [PubMed]
- Ng, L.C.; Tan, L.K.; Tan, C.H.; Tan, S.S.; Hapuarachchi, H.C.; Pok, K.Y.; Lai, Y.L.; Lam-Phua, S.G.; Bucht, G.; Lin, R.T.; et al. Entomologic and virologic investigation of chikungunya, Singapore. Emerg. Infect. Dis. 2009, 15, 1243–1249. [Google Scholar] [CrossRef]
- Theamboonlers, A.; Rianthavorn, P.; Praianantathavorn, K.; Wuttirattanakowit, N.; Poovorawan, Y. Clinical and molecular characterization of chikungunya virus in South Thailand. Jpn. J. Infect. Dis. 2009, 62, 303–305. [Google Scholar] [PubMed]
- Wu, D.; Wu, J.; Zhang, Q.; Zhong, H.; Ke, C.; Deng, X.; Guan, D.; Li, H.; Zhang, Y.; Zhou, H.; et al. Chikungunya outbreak in Guangdong Province, China, 2010. Emerg. Infect. Dis. 2012, 18, 493–495. [Google Scholar] [CrossRef] [PubMed]
- Duong, V.; Andries, A.C.; Ngan, C.; Sok, T.; Richner, B.; Asgari-Jirhandeh, N.; Bjorge, S.; Huy, R.; Ly, S.; Laurent, D.; et al. Reemergence of chikungunya virus in Cambodia. Emerg. Infect. Dis. 2012, 18, 2066–2069. [Google Scholar] [CrossRef] [PubMed]
- Wangchuk, S.; Chinnawirotpisan, P.; Dorji, T.; Tobgay, T.; Dorji, T.; Yoon, I.K.; Fernandez, S. Chikungunya fever outbreak, Bhutan, 2012. Emerg. Infect. Dis. 2013, 19, 1681–1684. [Google Scholar] [CrossRef] [PubMed]
- Hapuarachchi, H.C.; Bandara, K.B.; Sumanadasa, S.D.; Hapugoda, M.D.; Lai, Y.L.; Lee, K.S.; Tan, L.K.; Lin, R.T.; Ng, L.F.; Bucht, G.; et al. Re-emergence of chikungunya virus in south-east Asia: Virological evidence from Sri Lanka and Singapore. J. Gen. Virol. 2010, 91, 1067–1076. [Google Scholar] [CrossRef] [PubMed]
- Rianthavorn, P.; Prianantathavorn, K.; Wuttirattanakowit, N.; Theamboonlers, A.; Poovorawan, Y. An outbreak of chikungunya in Southern Thailand from 2008 to 2009 caused by African strains with A226V mutation. Int. J. Infect. Dis. 2010, 14, e161–e165. [Google Scholar] [CrossRef] [PubMed]
- Gould, E.A.; Gallian, P.; De Lamballerie, X.; Charrel, R.N. First cases of autochthonous dengue fever and chikungunya fever in France: From bad dream to reality! Clin. Microbiol. Infect. 2010, 16, 1702–1704. [Google Scholar] [CrossRef] [PubMed]
- Grandadam, M.; Caro, V.; Plumet, S.; Thiberge, J.M.; Souares, Y.; Failloux, A.B.; Tolou, H.J.; Budelot, M.; Cosserat, D.; Leparc-Goffart, I.; et al. Chikungunya virus, Southeastern France. Emerg. Infect. Dis. 2011, 17, 910–913. [Google Scholar] [CrossRef] [PubMed]
- Delaunay, P.; Jeannin, C.; Schaffner, F.; Marty, P. News on the presence of the tiger mosquito Aedes albopictus in metropolitan France. Arch. Pediatr. 2009, 16, S66–S71. [Google Scholar] [CrossRef] [PubMed]
- Vega-Rua, A.; Zouache, K.; Caro, V.; Diancourt, L.; Delaunay, P.; Grandadam, M.; Failloux, A.B. High efficiency of temperate Aedes albopictus to transmit chikungunya and dengue viruses in the Southeast of France. PLoS One 2013, 8, e59716. [Google Scholar] [CrossRef] [PubMed]
- Paty, M.; Six, C.; Charlet, F.; Heuze, G.; Cochet, A.; Wiegandt, A.; Chappert, J.; Dejour-Salamanca, D.; Guinard, A.; Soler, P.; et al. Large number of imported chikungunya cases in mainland France, 2014: A challenge for surveillance and response. Euro Surveill. 2014, 19, 20856. [Google Scholar] [PubMed]
- Tomasello, D.; Schlagenhauf, P. Chikungunya and dengue autochthonous cases in Europe, 2007–2012. Travel Med. Infect. Dis. 2013, 11, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Napoli, C.; Salcuni, P.; Pompa, M.G.; Declich, S.; Rizzo, C. Estimated imported infections of chikungunya and dengue in Italy, 2008 to 2011. J. Travel Med. 2012, 19, 294–297. [Google Scholar] [CrossRef] [PubMed]
- Leparc-Goffart, I.; Nougairede, A.; Cassadou, S.; Prat, C.; de Lamballerie, X. Chikungunya in the Americas. Lancet 2014, 383, 514. [Google Scholar] [CrossRef] [PubMed]
- Dupont-Rouzeyrol, M.; Caro, V.; Guillaumot, L.; Vazeille, M.; DʼOrtenzio, E.; Thiberge, J.M.; Baroux, N.; Gourinat, A.C.; Grandadam, M.; Failloux, A.B. Chikungunya virus and the mosquito vector Aedes aegypti in New Caledonia (South Pacific region). Vector borne and Zoonotic Dis. 2012, 12, 1036–1041. [Google Scholar] [CrossRef]
- Weaver, S.C. Arrival of chikungunya virus in the new world: Prospects for spread and impact on public health. PLoS Negl. Trop. Dis. 2014, 8, e2921. [Google Scholar] [CrossRef] [PubMed]
- Vega-Rua, A.; Zouache, K.; Girod, R.; Failloux, A.B.; Lourenco-de-Oliveira, R. High level of vector competence of Aedes aegypti and Aedes albopictus from ten American countries as a crucial factor in the spread of chikungunya virus. J. Virol. 2014, 88, 6294–6306. [Google Scholar] [CrossRef] [PubMed]
- ARBOnet. Chikungunya Virus Disease—United States, 2014 Provisional Data Reported to Arbonet; Centers for Disease Control Arboviral Diseases Branch: Fort Collins, CO, USA, 2014.
- Weinbren, M.P.; Haddow, A.J.; Williams, M.C. The occurrence of chikungunya virus in Uganda. I. Isolation from mosquitoes. Trans. R. Soc. Trop. Med. Hygiene 1958, 52, 253–257. [Google Scholar] [CrossRef]
- McCarthy, M.C.; Haberberger, R.L.; Salib, A.W.; Soliman, B.A.; El-Tigani, A.; Khalid, I.O.; Watts, D.M. Evaluation of arthropod-borne viruses and other infectious disease pathogens as the causes of febrile illnesses in the Khartoum Province of Sudan. J. Med. Virol. 1996, 48, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Diallo, M.; Thonnon, J.; Traore-Lamizana, M.; Fontenille, D. Vectors of chikungunya virus in Senegal: Current data and transmission cycles. The American journal of tropical medicine and hygiene 1999, 60, 281–286. [Google Scholar] [PubMed]
- Jupp, P.G.; Kemp, A. What is the potential for future outbreaks of chikungunya, dengue and yellow fever in Southern Africa? South. Afr. Med. J. 1996, 86, 35–37. [Google Scholar]
- Gubler, D.J. Epidemic dengue/dengue hemorrhagic fever as a public health, social and economic problem in the 21st century. Trends Microbiol. 2002, 10, 100–103. [Google Scholar] [CrossRef] [PubMed]
- McClelland, G.A.H. A worldwide survey of variation in scale pattern of the abdominal tergum of Aedes aegypti (l.) (diptera: Culicidae). Trans. R. Entomol. Soc. Lond. 1974, 126, 239–259. [Google Scholar] [CrossRef]
- Tabachnick, W.J.; Powell, J.R. A world-wide survey of genetic variation in the yellow fever mosquito, Aedes Aegypti. Genet. Res. 1979, 34, 215–229. [Google Scholar] [CrossRef] [PubMed]
- Failloux, A.B.; Vazeille, M.; Rodhain, F. Geographic genetic variation in populations of the dengue virus vector Aedes aegypti. J. Mol. Evolut. 2002, 55, 653–663. [Google Scholar] [CrossRef]
- Powell, J.R.; Tabachnick, W.J. History of domestication and spread of Aedes aegypti—A review. Memorias do Instituto Oswaldo Cruz 2013, 108, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Juliano, S.A.; Lounibos, L.P.; OʼMeara, G.F. A field test for competitive effects of Aedes albopictus on Aedes aegypti in South Florida: Differences between sites of coexistence and exclusion? Oecologia 2004, 139, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Braks, M.A.; Honorio, N.A.; Lourencqo-De-Oliveira, R.; Juliano, S.A.; Lounibos, L.P. Convergent habitat segregation of Aedes aegypti and Aedes albopictus (diptera: Culicidae) in Southeastern Brazil and Florida. J. Med. Entomol. 2003, 40, 785–794. [Google Scholar] [CrossRef] [PubMed]
- Simard, F.; Nchoutpouen, E.; Toto, J.C.; Fontenille, D. Geographic distribution and breeding site preference of Aedes albopictus and Aedes aegypti (diptera: Culicidae) in Cameroon, Central Africa. J. Med. Entomol. 2005, 42, 726–731. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.E. The history of dengue in tropical Asia and its probable relationship to the mosquito Aedes aegypti. J. Trop. Med. Hygiene 1956, 59, 243–251. [Google Scholar]
- Medlock, J.M.; Hansford, K.M.; Schaffner, F.; Versteirt, V.; Hendrickx, G.; Zeller, H.; Van Bortel, W. A review of the invasive mosquitoes in Europe: Ecology, public health risks, and control options. Vector borne Zoonotic Dis. 2012, 12, 435–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lounibos, L.P. Invasions by insect vectors of human disease. Annu. Rev. Entomol. 2002, 47, 233–266. [Google Scholar] [CrossRef] [PubMed]
- Rai, K.S. Aedes albopictus in the Americas. Annu. Rev. Entomol. 1991, 36, 459–484. [Google Scholar] [CrossRef] [PubMed]
- Hawley, W.A.; Reiter, P.; Copeland, R.S.; Pumpuni, C.B.; Craig, G.B., Jr. Aedes albopictus in North America: Probable introduction in used tires from Northern Asia. Science 1987, 236, 1114–1116. [Google Scholar] [CrossRef] [PubMed]
- Sprenger, D.; Wuithiranyagool, T. The discovery and distribution of Aedes albopictus in Harris County, Texas. J. Am. Mosq. Control Assoc. 1986, 2, 217–219. [Google Scholar] [PubMed]
- Adhami, J.; Reiter, P. Introduction and establishment of Aedes (Stegomyia) albopictus skuse (diptera: Culicidae) in Albania. J. Am. Mosq. Control Assoc. 1998, 14, 340–343. [Google Scholar] [PubMed]
- Sabatini, A.; Raineri, V.; Trovato, G.; Coluzzi, M. Aedes albopictus in Italy and possible diffusion of the species into the Mediterranean area. Parassitologia 1990, 32, 301–304. [Google Scholar] [PubMed]
- Hawley, W.A. The biology of Aedes albopictus. J. Am. Mosq. Control Association. Suppl. 1988, 1, 1–39. [Google Scholar]
- Paupy, C.; Delatte, H.; Bagny, L.; Corbel, V.; Fontenille, D. Aedes albopictus, an arbovirus vector: From the darkness to the light. Microbes Infect. 2009, 11, 1177–1185. [Google Scholar] [CrossRef] [PubMed]
- Metselaar, D.; Grainger, C.R.; Oei, K.G.; Reynolds, D.G.; Pudney, M.; Leake, C.J.; Tukei, P.M.; D’Offay, R.M.; Simpson, D.I. An outbreak of type 2 dengue fever in the Seychelles, probably transmitted by Aedes albopictus (skuse). Bull. World Health Organ. 1980, 58, 937–943. [Google Scholar] [PubMed]
- Delatte, H.; Dehecq, J.S.; Thiria, J.; Domerg, C.; Paupy, C.; Fontenille, D. Geographic distribution and developmental sites of Aedes albopictus (diptera: Culicidae) during a chikungunya epidemic event. Vector Borne Zoonotic Dis. 2008, 8, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Gjenero-Margan, I.; Aleraj, B.; Krajcar, D.; Lesnikar, V.; Klobucar, A.; Pem-Novosel, I.; Kurecic-Filipovic, S.; Komparak, S.; Martic, R.; Duricic, S.; et al. Autochthonous dengue fever in Croatia, August-September 2010. Euro Surveillance 2011, 16, 9. [Google Scholar]
- La Ruche, G.; Souares, Y.; Armengaud, A.; Peloux-Petiot, F.; Delaunay, P.; Despres, P.; Lenglet, A.; Jourdain, F.; Leparc-Goffart, I.; Charlet, F.; et al. First two autochthonous dengue virus infections in metropolitan France, September 2010. Euro Surveill. 2010, 15, 19676. [Google Scholar] [PubMed]
- Juliano, S.A.; O'Meara, G.F.; Morrill, J.R.; Cutwa, M.M. Desiccation and thermal tolerance of eggs and the coexistence of competing mosquitoes. Oecologia 2002, 130, 458–469. [Google Scholar] [CrossRef] [PubMed]
- O’Meara, G.F.; Evans, L.F., Jr.; Gettman, A.D.; Cuda, J.P. Spread of Aedes albopictus and decline of Ae. aegypti (diptera: Culicidae) in Florida. J. Med. Entomol. 1995, 32, 554–562. [Google Scholar] [PubMed]
- Juliano, S.A.; Lounibos, L.P. Ecology of invasive mosquitoes: Effects on resident species and on human health. Ecol. Lett. 2005, 8, 558–574. [Google Scholar] [PubMed]
- Kamgang, B.; Brengues, C.; Fontenille, D.; Njiokou, F.; Simard, F.; Paupy, C. Genetic structure of the tiger mosquito, Aedes albopictus, in Cameroon (Central Africa). PLoS One 2011, 6, e20257. [Google Scholar] [CrossRef] [PubMed]
- Kamgang, B.; Ngoagouni, C.; Manirakiza, A.; Nakoune, E.; Paupy, C.; Kazanji, M. Temporal patterns of abundance of Aedes aegypti and Aedes albopictus (diptera: Culicidae) and mitochondrial DNA analysis of Ae. albopictus in the Central African Republic. PLoS Negl. Trop. Dis. 2013, 7, e2590. [Google Scholar] [CrossRef] [PubMed]
- Raharimalala, F.N.; Ravaomanarivo, L.H.; Ravelonandro, P.; Rafarasoa, L.S.; Zouache, K.; Tran-Van, V.; Mousson, L.; Failloux, A.B.; Hellard, E.; Moro, C.V.; et al. Biogeography of the two major arbovirus mosquito vectors, Aedes aegypti and Aedes albopictus (diptera, culicidae), in Madagascar. Parasites Vectors 2012, 5, 56. [Google Scholar] [CrossRef] [PubMed]
- Reiter, P.; Sprenger, D. The used tire trade: A mechanism for the worldwide dispersal of container breeding mosquitoes. J. Am. Mosq. Control Assoc. 1987, 3, 494–501. [Google Scholar] [PubMed]
- Nene, V.; Wortman, J.R.; Lawson, D.; Haas, B.; Kodira, C.; Tu, Z.J.; Loftus, B.; Xi, Z.; Megy, K.; Grabherr, M.; et al. Genome sequence of Aedes aegypti, a major arbovirus vector. Science 2007, 316, 1718–1723. [Google Scholar] [PubMed]
- Bonizzoni, M.; Gasperi, G.; Chen, X.; James, A.A. The invasive mosquito species Aedes albopictus: Current knowledge and future perspectives. Trends Parasitol. 2013, 29, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Urdaneta-Marquez, L.; Failloux, A.B. Population genetic structure of Aedes aegypti, the principal vector of dengue viruses. Infect. Genetics Evol. 2011, 11, 253–261. [Google Scholar] [CrossRef]
- Birungi, J.; Munstermann, L.E. Genetic structure of Aedes albopictus (diptera: Culicidae) populations based on mitochondrial nd5 sequences: Evidence for an independent invasion into Brazil and United States. Ann. Entomol. Soc. Am. 2002, 95, 125–132. [Google Scholar] [CrossRef]
- Maia, R.T.; Scarpassa, V.M.; Maciel-Litaiff, L.H.; Tadei, W.P. Reduced levels of genetic variation in Aedes albopictus (diptera: Culicidae) from Manaus, Amazonas State, Brazil, based on analysis of the mitochondrial DNA nd5 gene. Genetics Mol. Res. 2009, 8, 998–1007. [Google Scholar] [CrossRef]
- Mousson, L.; Dauga, C.; Garrigues, T.; Schaffner, F.; Vazeille, M.; Failloux, A.B. Phylogeography of Aedes (stegomyia) aegypti (l.) and Aedes (stegomyia) albopictus (skuse) (diptera: Culicidae) based on mitochondrial DNA variations. Genet. Res. 2005, 86, 1–11. [Google Scholar] [PubMed]
- Usmani-Brown, S.; Cohnstaedt, L.; Munstermann, L.E. Population genetics of Aedes albopictus (diptera: Culicidae) invading populations, using mitochondrial nicotinamide adenine dinucleotide dehydrogenase subunit 5 sequences. Ann. Entomol. Soc. Am. 2009, 102, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, R.G.; Roderick, G.K. Arthropods on islands: Colonization, speciation, and conservation. Annu. Rev. Entomol. 2002, 47, 595–632. [Google Scholar] [CrossRef] [PubMed]
- Beebe, N.W.; Ambrose, L.; Hill, L.A.; Davis, J.B.; Hapgood, G.; Cooper, R.D.; Russell, R.C.; Ritchie, S.A.; Reimer, L.J.; Lobo, N.F.; et al. Tracing the tiger: Population genetics provides valuable insights into the Aedes (stegomyia) albopictus invasion of the Australasian region. PLoS Negl. Trop. Dis. 2013, 7, e2361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontenille, D.; Rodhain, F. Biology and distribution of Aedes albopictus and Aedes aegypti in Madagascar. J. Am. Mosq. Control Assoc. 1989, 5, 219–225. [Google Scholar] [PubMed]
- Bagny, L.; Arnoux, S.; Delatte, H.; Lajoie, G.; Fontenille, D. Spread of invasive Aedes albopictus and decline of resident Aedes aegypti in urban areas of Mayotte 2007–2010. Biol. Invasions 2012, 14, 1623–1633. [Google Scholar] [CrossRef]
- Diallo, M.; Laganier, R.; Nangouma, A. First record of Ae. albopictus (skuse 1894), in Central African Republic. Trop. Med. Int. Health 2010, 15, 1185–1189. [Google Scholar] [CrossRef] [PubMed]
- Fontenille, D.; Toto, J.C. Aedes (stegomyia) albopictus (skuse), a potential new dengue vector in Southern Cameroon. Emerg. Infect. Dis. 2001, 7, 1066–1067. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Dash, P.K.; Singh, A.K.; Sharma, S.; Gopalan, N.; Rao, P.V.; Parida, M.M.; Reiter, P. Evidence of experimental vertical transmission of emerging novel ECSA genotype of chikungunya virus in Aedes aegypti. PLoS Negl. Trop. Dis. 2014, 8, e2990. [Google Scholar] [CrossRef] [PubMed]
- Thiberville, S.D.; Boisson, V.; Gaudart, J.; Simon, F.; Flahault, A.; de Lamballerie, X. Chikungunya fever: A clinical and virological investigation of outpatients on Reunion Island, South-West Indian Ocean. PLoS Negl. Trop. Dis. 2013, 7, e2004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pesko, K.; Westbrook, C.J.; Mores, C.N.; Lounibos, L.P.; Reiskind, M.H. Effects of infectious virus dose and bloodmeal delivery method on susceptibility of Aedes aegypti and Aedes albopictus to chikungunya virus. J. Med. Entomol. 2009, 46, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Martin, E.; Moutailler, S.; Madec, Y.; Failloux, A.B. Differential responses of the mosquito Aedes albopictus from the Indian Ocean region to two chikungunya isolates. BMC Ecol. 2010, 10, 8. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Vargas, I.; Travanty, E.A.; Keene, K.M.; Franz, A.W.; Beaty, B.J.; Blair, C.D.; Olson, K.E. RNA interference, arthropod-borne viruses, and mosquitoes. Virus Res. 2004, 102, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Xi, Z.; Ramirez, J.L.; Dimopoulos, G. The Aedes aegypti toll pathway controls dengue virus infection. PLoS Pathog. 2008, 4, e1000098. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, J.L.; Dimopoulos, G. The toll immune signaling pathway control conserved anti-dengue defenses across diverse Ae. aegypti strains and against multiple dengue virus serotypes. Dev. Compar. Immunol. 2010, 34, 625–629. [Google Scholar] [CrossRef]
- Souza-Neto, J.A.; Sim, S.; Dimopoulos, G. An evolutionary conserved function of the jak-stat pathway in anti-dengue defense. Proc. Natl. Acad. Sci. USA 2009, 106, 17841–17846. [Google Scholar] [CrossRef] [PubMed]
- Sanders, H.R.; Foy, B.D.; Evans, A.M.; Ross, L.S.; Beaty, B.J.; Olson, K.E.; Gill, S.S. Sindbis virus induces transport processes and alters expression of innate immunity pathway genes in the midgut of the disease vector, Aedes aegypti. Insect Biochem. Mol. Biol. 2005, 35, 1293–1307. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Vargas, I.; Scott, J.C.; Poole-Smith, B.K.; Franz, A.W.; Barbosa-Solomieu, V.; Wilusz, J.; Olson, K.E.; Blair, C.D. Dengue virus type 2 infections of Aedes aegypti are modulated by the mosquito's RNA interference pathway. PLoS Pathog. 2009, 5, e1000299. [Google Scholar] [CrossRef] [PubMed]
- Keene, K.M.; Foy, B.D.; Sanchez-Vargas, I.; Beaty, B.J.; Blair, C.D.; Olson, K.E. RNA interference acts as a natural antiviral response to o’nyong-nyong virus (alphavirus; Togaviridae) infection of Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2004, 101, 17240–17245. [Google Scholar] [CrossRef] [PubMed]
- Campbell, C.L.; Keene, K.M.; Brackney, D.E.; Olson, K.E.; Blair, C.D.; Wilusz, J.; Foy, B.D. Aedes aegypti uses RNA interference in defense against sindbis virus infection. BMC Microbiol. 2008, 8, 47. [Google Scholar] [CrossRef] [PubMed]
- Myles, K.M.; Wiley, M.R.; Morazzani, E.M.; Adelman, Z.N. Alphavirus-derived small RNAs modulate pathogenesis in disease vector mosquitoes. Proc. Natl. Acad. Sci. USA 2008, 105, 19938–19943. [Google Scholar] [CrossRef] [PubMed]
- Cirimotich, C.M.; Scott, J.C.; Phillips, A.T.; Geiss, B.J.; Olson, K.E. Suppression of RNA interference increases alphavirus replication and virus-associated mortality in Aedes aegypti mosquitoes. BMC Microbiol. 2009, 9, 49. [Google Scholar] [CrossRef] [PubMed]
- Khoo, C.C.; Piper, J.; Sanchez-Vargas, I.; Olson, K.E.; Franz, A.W. The RNA interference pathway affects midgut infection- and escape barriers for sindbis virus in Aedes aegypti. BMC Microbiol. 2010, 10, 130. [Google Scholar] [CrossRef] [PubMed]
- Donald, C.L.; Kohl, A.; Schnettler, E. New insights into control of arbovirus replication and spread by insect RNA interference pathways. Insects 2012, 3, 511–531. [Google Scholar]
- McFarlane, M.; Arias-Goeta, C.; Martin, E.; OʼHara, Z.; Lulla, A.; Mousson, L.; Rainey, S.M.; Misbah, S.; Schnettler, E.; Donald, C.L.; et al. Characterization of Aedes aegypti innate-immune pathways that limit chikungunya virus replication. PLoS Negl. Trop. Dis. 2014, 8, e2994. [Google Scholar] [CrossRef] [PubMed]
- Schnettler, E.; Donald, C.L.; Human, S.; Watson, M.; Siu, R.W.; McFarlane, M.; Fazakerley, J.K.; Kohl, A.; Fragkoudis, R. Knockdown of piRNA pathway proteins results in enhanced semliki forest virus production in mosquito cells. J. Gen. Virol. 2013, 94, 1680–1689. [Google Scholar] [CrossRef] [PubMed]
- McGraw, E.A.; OʼNeill, S.L. Beyond insecticides: New thinking on an ancient problem. Nat. Rev. Microbiol. 2013, 11, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Franz, A.W.; Sanchez-Vargas, I.; Adelman, Z.N.; Blair, C.D.; Beaty, B.J.; James, A.A.; Olson, K.E. Engineering RNA interference-based resistance to dengue virus type 2 in genetically modified Aedes aegypti. Proc. Natl. Acad. Sci. USA 2006, 103, 4198–4203. [Google Scholar] [CrossRef] [PubMed]
- Mathur, G.; Sanchez-Vargas, I.; Alvarez, D.; Olson, K.E.; Marinotti, O.; James, A.A. Transgene-mediated suppression of dengue viruses in the salivary glands of the yellow fever mosquito, Aedes aegypti. Insect Mol. Biol. 2010, 19, 753–763. [Google Scholar] [CrossRef] [PubMed]
- Travanty, E.A.; Adelman, Z.N.; Franz, A.W.; Keene, K.M.; Beaty, B.J.; Blair, C.D.; James, A.A.; Olson, K.E. Using RNA interference to develop dengue virus resistance in genetically modified Aedes aegypti. Insect Biochem. Mol. Biol. 2004, 34, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Alphey, L.; McKemey, A.; Nimmo, D.; Neira Oviedo, M.; Lacroix, R.; Matzen, K.; Beech, C. Genetic control of Aedes mosquitoes. Pathog. Glob. Health 2013, 107, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Alphey, L. Natural and engineered mosquito immunity. J. Biol. 2009, 8, 40. [Google Scholar] [CrossRef] [PubMed]
- Fraser, M.J., Jr. Insect transgenesis: Current applications and future prospects. Annu. Rev. Entomol. 2012, 57, 267–289. [Google Scholar] [CrossRef] [PubMed]
- Walker, T.; Johnson, P.H.; Moreira, L.A.; Iturbe-Ormaetxe, I.; Frentiu, F.D.; McMeniman, C.J.; Leong, Y.S.; Dong, Y.; Axford, J.; Kriesner, P.; et al. The wMel wolbachia strain blocks dengue and invades caged Aedes aegypti populations. Nature 2011, 476, 450–453. [Google Scholar] [CrossRef] [PubMed]
- Sinkins, S.P.; Gould, F. Gene drive systems for insect disease vectors. Nat. Rev. Genet. 2006, 7, 427–435. [Google Scholar] [CrossRef]
- Chen, C.H.; Huang, H.; Ward, C.M.; Su, J.T.; Schaeffer, L.V.; Guo, M.; Hay, B.A. A synthetic maternal-effect selfish genetic element drives population replacement in Drosophila. Science 2007, 316, 597–600. [Google Scholar] [CrossRef] [PubMed]
- McMeniman, C.J.; Lane, R.V.; Cass, B.N.; Fong, A.W.; Sidhu, M.; Wang, Y.F.; OʼNeill, S.L. Stable introduction of a life-shortening wolbachia infection into the mosquito Aedes aegypti. Science 2009, 323, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Hedges, L.M.; Brownlie, J.C.; O’Neill, S.L.; Johnson, K.N. Wolbachia and virus protection in insects. Science 2008, 322, 702. [Google Scholar] [CrossRef] [PubMed]
- Moreira, L.A.; Iturbe-Ormaetxe, I.; Jeffery, J.A.; Lu, G.; Pyke, A.T.; Hedges, L.M.; Rocha, B.C.; Hall-Mendelin, S.; Day, A.; Riegler, M.; et al. A wolbachia symbiont in Aedes aegypti limits infection with dengue, chikungunya, and plasmodium. Cell 2009, 139, 1268–1278. [Google Scholar] [CrossRef] [PubMed]
- Hancock, P.A.; Sinkins, S.P.; Godfray, H.C. Strategies for introducing wolbachia to reduce transmission of mosquito-borne diseases. PLoS Negl. Trop. Dis. 2011, 5, e1024. [Google Scholar] [CrossRef] [PubMed]
- Zouache, K.; Voronin, D.; Tran-Van, V.; Mousson, L.; Failloux, A.B.; Mavingui, P. Persistent wolbachia and cultivable bacteria infection in the reproductive and somatic tissues of the mosquito vector Aedes albopictus. PLoS One 2009, 4, e6388. [Google Scholar] [CrossRef] [PubMed]
- Mousson, L.; Martin, E.; Zouache, K.; Madec, Y.; Mavingui, P.; Failloux, A.B. Wolbachia modulates chikungunya replication in Aedes albopictus. Mol. Ecol. 2010, 19, 1953–1964. [Google Scholar] [PubMed]
- Mousson, L.; Zouache, K.; Arias-Goeta, C.; Raquin, V.; Mavingui, P.; Failloux, A.B. The native wolbachia symbionts limit transmission of dengue virus in Aedes albopictus. PLoS Negl. Trop. Dis. 2012, 6, e1989. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Bian, G.; Pan, X.; Xi, Z. Wolbachia induces density-dependent inhibition to dengue virus in mosquito cells. PLoS Negl. Trop. Dis. 2012, 6, e1754. [Google Scholar] [PubMed]
- Blagrove, M.S.; Arias-Goeta, C.; Di Genua, C.; Failloux, A.B.; Sinkins, S.P. A wolbachia wMel transinfection in Aedes albopictus is not detrimental to host fitness and inhibits chikungunya virus. PLoS Negl. Trop. Dis. 2013, 7, e2152. [Google Scholar] [CrossRef] [PubMed]
- van den Hurk, A.F.; Hall-Mendelin, S.; Pyke, A.T.; Frentiu, F.D.; McElroy, K.; Day, A.; Higgs, S.; OʼNeill, S.L. Impact of wolbachia on infection with chikungunya and yellow fever viruses in the mosquito vector Aedes aegypti. PLoS Negl. Trop. Dis. 2012, 6, e1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rances, E.; Ye, Y.H.; Woolfit, M.; McGraw, E.A.; OʼNeill, S.L. The relative importance of innate immune priming in wolbachia-mediated dengue interference. PLoS Pathog. 2012, 8, e1002548. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Zhou, G.; Wu, J.; Bian, G.; Lu, P.; Raikhel, A.S.; Xi, Z. Wolbachia induces reactive oxygen species (ros)-dependent activation of the toll pathway to control dengue virus in the mosquito Aedes aegypti. Proc. Natl. Acad. Sci. USA 2012, 109, E23–E31. [Google Scholar] [CrossRef] [PubMed]
- Caragata, E.P.; Rances, E.; Hedges, L.M.; Gofton, A.W.; Johnson, K.N.; OʼNeill, S.L.; McGraw, E.A. Dietary cholesterol modulates pathogen blocking by wolbachia. PLoS Pathog. 2013, 9, e1003459. [Google Scholar] [CrossRef] [PubMed]
- Zouache, K.; Raharimalala, F.N.; Raquin, V.; Tran-Van, V.; Raveloson, L.H.; Ravelonandro, P.; Mavingui, P. Bacterial diversity of field-caught mosquitoes, Aedes albopictus and Aedes aegypti, from different geographic regions of Madagascar. FEMS Microbiol. Ecol. 2011, 75, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Minard, G.; Tran, F.H.; Raharimalala, F.N.; Hellard, E.; Ravelonandro, P.; Mavingui, P.; Valiente Moro, C. Prevalence, genomic and metabolic profiles of acinetobacter and asaia associated with field-caught Aedes albopictus from Madagascar. FEMS Microbiol. Ecol. 2013, 83, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, J.L.; Souza-Neto, J.; Torres Cosme, R.; Rovira, J.; Ortiz, A.; Pascale, J.M.; Dimopoulos, G. Reciprocal tripartite interactions between the Aedes aegypti midgut microbiota, innate immune system and dengue virus influences vector competence. PLoS Negl. Trop. Dis. 2012, 6, e1561. [Google Scholar] [CrossRef] [PubMed]
- Apte-Deshpande, A.D.; Paingankar, M.S.; Gokhale, M.D.; Deobagkar, D.N. Serratia odorifera mediated enhancement in susceptibility of Aedes aegypti for chikungunya virus. Indian J. Med. Res. 2014, 139, 762–768. [Google Scholar] [PubMed]
- Black, W.C.; Bennett, K.E.; Gorrochotegui-Escalante, N.; Barillas-Mury, C.V.; Fernandez-Salas, I.; de Lourdes Munoz, M.; Farfan-Ale, J.A.; Olson, K.E.; Beaty, B.J. Flavivirus susceptibility in Aedes aegypti. Arch. Med. Res. 2002, 33, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Bennett, K.E.; Flick, D.; Fleming, K.H.; Jochim, R.; Beaty, B.J.; Black, W.C.T. Quantitative trait loci that control dengue-2 virus dissemination in the mosquito Aedes aegypti. Genetics 2005, 170, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Stapleford, K.A.; Coffey, L.L.; Lay, S.; Borderia, A.V.; Duong, V.; Isakov, O.; Rozen-Gagnon, K.; Arias-Goeta, C.; Blanc, H.; Beaucourt, S.; et al. Emergence and transmission of arbovirus evolutionary intermediates with epidemic potential. Cell. Host Microbe 2014, 15, 706–716. [Google Scholar] [CrossRef] [PubMed]
- Myers, R.M.; Carey, D.E. Concurrent isolation from patient of two arboviruses, chikungunya and dengue type 2. Science 1967, 157, 1307–1308. [Google Scholar] [CrossRef] [PubMed]
- Hapuarachchi, H.A.; Bandara, K.B.; Hapugoda, M.D.; Williams, S.; Abeyewickreme, W. Laboratory confirmation of dengue and chikungunya co-infection. Ceylon Med. J. 2008, 53, 104–105. [Google Scholar] [PubMed]
- Ezzedine, K.; Cazanave, C.; Pistone, T.; Receveur, M.C.; Neau, D.; Ragnaud, J.M.; Malvy, D. Dual infection by chikungunya virus and other imported infectious agent in a traveller returning from India. Travel Med. Infect. Dis. 2008, 6, 152–154. [Google Scholar] [CrossRef] [PubMed]
- Chahar, H.S.; Bharaj, P.; Dar, L.; Guleria, R.; Kabra, S.K.; Broor, S. Co-infections with chikungunya virus and dengue virus in Delhi, India. Emerg. Infect. Dis. 2009, 15, 1077–1080. [Google Scholar] [CrossRef] [PubMed]
- Schilling, S.; Emmerich, P.; Gunther, S.; Schmidt-Chanasit, J. Dengue and chikungunya virus co-infection in a German traveller. J. Clin. Virol. 2009, 45, 163–164. [Google Scholar] [CrossRef] [PubMed]
- Nayar, S.K.; Noridah, O.; Paranthaman, V.; Ranjit, K.; Norizah, I.; Chem, Y.K.; Mustafa, B.; Chua, K.B. Co-infection of dengue virus and chikungunya virus in two patients with acute febrile illness. Med. J. Malays. 2007, 62, 335–336. [Google Scholar]
- Leroy, E.M.; Nkoghe, D.; Ollomo, B.; Nze-Nkogue, C.; Becquart, P.; Grard, G.; Pourrut, X.; Charrel, R.; Moureau, G.; Ndjoyi-Mbiguino, A.; et al. Concurrent chikungunya and dengue virus infections during simultaneous outbreaks, Gabon, 2007. Emerg. Infect. Dis. 2009, 15, 591–593. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.F.; Su, C.L.; Shu, P.Y.; Yang, C.F.; Liao, T.L.; Cheng, C.H.; Hu, H.C.; Huang, J.H. Concurrent isolation of chikungunya virus and dengue virus from a patient with coinfection resulting from a trip to Singapore. J. Clin. Microbiol. 2010, 48, 4586–4589. [Google Scholar] [CrossRef] [PubMed]
- Parreira, R.; Centeno-Lima, S.; Lopes, A.; Portugal-Calisto, D.; Constantino, A.; Nina, J. Dengue virus serotype 4 and chikungunya virus coinfection in a traveller returning from Luanda, Angola, January 2014. Euro Surveill. 2014, 19, 10. [Google Scholar]
- Caron, M.; Paupy, C.; Grard, G.; Becquart, P.; Mombo, I.; Nso, B.B.; Kassa Kassa, F.; Nkoghe, D.; Leroy, E.M. Recent introduction and rapid dissemination of chikungunya virus and dengue virus serotype 2 associated with human and mosquito coinfections in Gabon, Central Africa. Clin. Infect. Dis. 2012, 55, e45–e53. [Google Scholar] [CrossRef] [PubMed]
- Vazeille, M.; Mousson, L.; Martin, E.; Failloux, A.B. Orally co-infected Aedes albopictus from La Reunion Island, Indian Ocean, can deliver both dengue and chikungunya infectious viral particles in their saliva. PLoS Negl. Trop. Dis. 2010, 4, e706. [Google Scholar] [CrossRef] [PubMed]
- Courtois, G.; Osterrieth, P.; Blanes Ridaura, G. Isolation of the yellow fever virus in the Belgian Congo. Ann. Soc. Belge Med. Tropicale 1960, 40, 29–60. [Google Scholar]
- Osterrieth, P.; Rathe, E.; Deleplanque-Liegeois, P. Simultaneous isolation of the yellow fever and chikungunya viruses at Bili (Haut-Uele, Belgian Congo). Ann. Soc. Belge Med. Tropicale 1961, 41, 207–212. [Google Scholar]
- Filipe, A.F.; Pinto, M.R. Arbovirus studies in Luanda, Angola. 2. Virological and serological studies during an outbreak of dengue-like disease caused by the chikungunya virus. Bull. World Health Organ. 1973, 49, 37–40. [Google Scholar] [PubMed]
- Gould, L.H.; Osman, M.S.; Farnon, E.C.; Griffith, K.S.; Godsey, M.S.; Karch, S.; Mulenda, B.; El Kholy, A.; Grandesso, F.; de Radigues, X.; et al. An outbreak of yellow fever with concurrent chikungunya virus transmission in South Kordofan, Sudan, 2005. Trans. R. Soc. Trop. Med. Hygiene 2008, 102, 1247–1254. [Google Scholar] [CrossRef]
- Grard, G.; Caron, M.; Mombo, I.M.; Nkoghe, D.; Mboui Ondo, S.; Jiolle, D.; Fontenille, D.; Paupy, C.; Leroy, E.M. Zika virus in Gabon (Central Africa)—2007: A new threat from Aedes albopictus? PLoS Negl. Trop. Dis. 2014, 8, e2681. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, P.M.; Rico-Hesse, R. Differential susceptibility of Aedes aegypti to infection by the American and Southeast Asian genotypes of dengue type 2 virus. Vector Borne Zoonotic Dis. 2001, 1, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Cologna, R.; Armstrong, P.M.; Rico-Hesse, R. Selection for virulent dengue viruses occurs in humans and mosquitoes. J. Virol. 2005, 79, 853–859. [Google Scholar] [CrossRef] [PubMed]
- Brault, A.C.; Powers, A.M.; Ortiz, D.; Estrada-Franco, J.G.; Navarro-Lopez, R.; Weaver, S.C. Venezuelan equine encephalitis emergence: Enhanced vector infection from a single amino acid substitution in the envelope glycoprotein. Proc. Natl. Acad. Sci. USA 2004, 101, 11344–11349. [Google Scholar] [CrossRef] [PubMed]
- Deardorff, E.R.; Weaver, S.C. Vector competence of Culex (Melanoconion) taeniopus for equine-virulent subtype ie strains of venezuelan equine encephalitis virus. Am. J. Trop. Med. Hygiene 2010, 82, 1047–1052. [Google Scholar] [CrossRef]
- Tsetsarkin, K.A.; Weaver, S.C. Sequential adaptive mutations enhance efficient vector switching by chikungunya virus and its epidemic emergence. PLoS Pathog. 2011, 7, e1002412. [Google Scholar] [CrossRef] [PubMed]
- Holmes, E.C. The Evolution and Emergence of RNA Viruses; Oxford University Press: New York, NY, USA, 2009. [Google Scholar]
- Rozen-Gagnon, K.; Stapleford, K.A.; Mongelli, V.; Blanc, H.; Failloux, A.B.; Saleh, M.C.; Vignuzzi, M. Alphavirus mutator variants present host-specific defects and attenuation in mammalian and insect models. PLoS Pathog. 2014, 10, e1003877. [Google Scholar] [CrossRef] [PubMed]
- Coffey, L.L.; Beeharry, Y.; Borderia, A.V.; Blanc, H.; Vignuzzi, M. Arbovirus high fidelity variant loses fitness in mosquitoes and mice. Proc. Natl. Acad. Sci. USA 2011, 108, 16038–16043. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.T.; Ebel, G.D.; Lanciotti, R.S.; Brault, A.C.; Guzman, H.; Siirin, M.; Lambert, A.; Parsons, R.E.; Beasley, D.W.; Novak, R.J.; et al. Phylogenetic analysis of North American West Nile virus isolates, 2001–2004: Evidence for the emergence of a dominant genotype. Virology 2005, 342, 252–265. [Google Scholar] [CrossRef] [PubMed]
- Ebel, G.D.; Carricaburu, J.; Young, D.; Bernard, K.A.; Kramer, L.D. Genetic and phenotypic variation of West Nile virus in New York, 2000–2003. Am. J. Trop. Med. Hygiene 2004, 71, 493–500. [Google Scholar]
- Parameswaran, P.; Charlebois, P.; Tellez, Y.; Nunez, A.; Ryan, E.M.; Malboeuf, C.M.; Levin, J.Z.; Lennon, N.J.; Balmaseda, A.; Harris, E.; et al. Genome-wide patterns of intrahuman dengue virus diversity reveal associations with viral phylogenetic clade and interhost diversity. J. Virol. 2012, 86, 8546–8558. [Google Scholar] [CrossRef] [PubMed]
- Turner, P.E.; Morales, N.M.; Alto, B.W.; Remold, S.K. Role of evolved host breadth in the initial emergence of an RNA virus. Evolution 2010, 64, 3273–3286. [Google Scholar] [CrossRef] [PubMed]
- Coffey, L.L.; Forrester, N.; Tsetsarkin, K.; Vasilakis, N.; Weaver, S.C. Factors shaping the adaptive landscape for arboviruses: Implications for the emergence of disease. Future Microbiol. 2013, 8, 155–176. [Google Scholar] [CrossRef] [PubMed]
- Coffey, L.L.; Vignuzzi, M. Host alternation of chikungunya virus increases fitness while restricting population diversity and adaptability to novel selective pressures. J. Virol. 2011, 85, 1025–1035. [Google Scholar] [CrossRef] [PubMed]
- Muller, H.J. The relation of recombination to mutational advance. Mutat. Res. 1964, 1, 2–9. [Google Scholar]
- Weaver, S.C.; Brault, A.C.; Kang, W.; Holland, J.J. Genetic and fitness changes accompanying adaptation of an arbovirus to vertebrate and invertebrate cells. J. Virol. 1999, 73, 4316–4326. [Google Scholar] [PubMed]
- Duarte, E.A.; Clarke, D.K.; Moya, A.; Elena, S.F.; Domingo, E.; Holland, J. Many-trillionfold amplification of single RNA virus particles fails to overcome the Mullerʼs ratchet effect. J. Virol. 1993, 67, 3620–3623. [Google Scholar] [PubMed]
- Smith, D.R.; Adams, A.P.; Kenney, J.L.; Wang, E.; Weaver, S.C. Venezuelan equine encephalitis virus in the mosquito vector Aedes taeniorhynchus: Infection initiated by a small number of susceptible epithelial cells and a population bottleneck. Virology 2008, 372, 176–186. [Google Scholar] [CrossRef]
- Kenney, J.L.; Adams, A.P.; Gorchakov, R.; Leal, G.; Weaver, S.C. Genetic and anatomic determinants of enzootic venezuelan equine encephalitis virus infection of Culex (Melanoconion) taeniopus. PLoS Negl. Trop. Dis. 2012, 6, e1606. [Google Scholar] [CrossRef] [PubMed]
- Forrester, N.L.; Guerbois, M.; Seymour, R.L.; Spratt, H.; Weaver, S.C. Vector-borne transmission imposes a severe bottleneck on an RNA virus population. PLoS Pathog. 2012, 8, e1002897. [Google Scholar] [CrossRef] [PubMed]
- Brackney, D.E.; Pesko, K.N.; Brown, I.K.; Deardorff, E.R.; Kawatachi, J.; Ebel, G.D. West Nile virus genetic diversity is maintained during transmission by Culex pipiens quinquefasciatus mosquitoes. PLoS One 2011, 6, e24466. [Google Scholar] [CrossRef] [PubMed]
- Coffey, L.L.; Vasilakis, N.; Brault, A.C.; Powers, A.M.; Tripet, F.; Weaver, S.C. Arbovirus evolution in vivo is constrained by host alternation. Proc. Natl. Acad. Sci. USA 2008, 105, 6970–6975. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.R.; Aguilar, P.V.; Coffey, L.L.; Gromowski, G.D.; Wang, E.; Weaver, S.C. Venezuelan equine encephalitis virus transmission and effect on pathogenesis. Emerg. Infect. Dis. 2006, 12, 1190–1196. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Moreno, D.; Vargas, I.S.; Olson, K.E.; Harrington, L.C. Modeling dynamic introduction of chikungunya virus in the United States. PLoS Negl. Trop. Dis. 2012, 6, e1918. [Google Scholar] [CrossRef] [PubMed]
- Eisen, L.; Monaghan, A.J.; Lozano-Fuentes, S.; Steinhoff, D.F.; Hayden, M.H.; Bieringer, P.E. The impact of temperature on the bionomics of Aedes (stegomyia) aegypti, with special reference to the cool geographic range margins. J. Med. Entomol. 2014, 51, 496–516. [Google Scholar] [CrossRef] [PubMed]
- Zouache, K.; Fontaine, A.; Vega-Rua, A.; Mousson, L.; Thiberge, J.M.; Lourenco-De-Oliveira, R.; Caro, V.; Lambrechts, L.; Failloux, A.B. Three-way interactions between mosquito population, viral strain and temperature underlying chikungunya virus transmission potential. Proc. Biol. Sci. 2014, 281, 1–8. [Google Scholar] [CrossRef]
- Tsetsarkin, K.A.; Chen, R.; Leal, G.; Forrester, N.; Higgs, S.; Huang, J.; Weaver, S.C. Chikungunya virus emergence is constrained in Asia by lineage-specific adaptive landscapes. Proc. Natl. Acad. Sci. USA 2011, 108, 7872–7877. [Google Scholar] [CrossRef] [PubMed]
- Lanciotti, R.S.; Kosoy, O.L.; Laven, J.J.; Panella, A.J.; Velez, J.O.; Lambert, A.J.; Campbell, G.L. Chikungunya virus in US travelers returning from India, 2006. Emerg. Infect. Dis. 2007, 13, 764–767. [Google Scholar] [CrossRef] [PubMed]
- Wilke, A.B.; Marrelli, M.T. Genetic control of mosquitoes: Population suppression strategies. Rev. Inst. Med. Trop. Sao Paulo 2012, 54, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.C.; Osorio, J.E.; Livengood, J.A.; Chen, R.; Stinchcomb, D.T. Chikungunya virus and prospects for a vaccine. Expert Rev. Vaccines 2012, 11, 1087–1101. [Google Scholar] [CrossRef] [PubMed]
- Paupy, C.; Ollomo, B.; Kamgang, B.; Moutailler, S.; Rousset, D.; Demanou, M.; Herve, J.P.; Leroy, E.; Simard, F. Comparative role of Aedes albopictus and Aedes aegypti in the emergence of dengue and chikungunya in Central Africa. Vector Borne Zoonotic Dis. 2010, 10, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Turell, M.J.; Beaman, J.R.; Tammariello, R.F. Susceptibility of selected strains of Aedes aegypti and Aedes albopictus (diptera: Culicidae) to chikungunya virus. J. Med. Entomol. 1992, 29, 49–53. [Google Scholar] [PubMed]
- Shah, K.V.; Gilotra, S.K.; Gibbs, C.J., Jr.; Rozeboom, L.E. Laboratory studies of transmission of chikungunya virus by mosquitoes: A preliminary report. Indian J. Med. Res. 1964, 52, 703–709. [Google Scholar] [PubMed]
- Girod, R.; Gaborit, P.; Marrama, L.; Etienne, M.; Ramdini, C.; Rakotoarivony, I.; Dollin, C.; Carinci, R.; Issaly, J.; Dusfour, I.; et al. High susceptibility to chikungunya virus of Aedes aegypti from the French West Indies and French Guiana. Trop. Med. Int. Health 2011, 16, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Vanlandingham, D.L.; Hong, C.; Klingler, K.; Tsetsarkin, K.; McElroy, K.L.; Powers, A.M.; Lehane, M.J.; Higgs, S. Differential infectivities of oʼnyong-nyong and chikungunya virus isolates in Anopheles gambiae and Aedes aegypti mosquitoes. Am. J. Trop. Med. Hygiene 2005, 72, 616–621. [Google Scholar]
- Mangiafico, J.A. Chikungunya virus infection and transmission in five species of mosquito. Am. J. Trop. Med. Hygiene 1971, 20, 642–645. [Google Scholar]
- Vazeille, M.; Yebakima, A.; Lourenco-de-Oliveira, R.; Andriamahefazafy, B.; Correira, A.; Rodrigues, J.M.; Veiga, A.; Moreira, A.; Leparc-Goffart, I.; Grandadam, M.; et al. Oral receptivity of Aedes aegypti from Cape Verde for yellow fever, dengue, and chikungunya viruses. Vector Borne Zoonotic Dis. 2013, 13, 37–40. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, J.; Ritchie, S.A.; van den Hurk, A.F. Aedes albopictus (diptera: Culicidae) as a potential vector of endemic and exotic arboviruses in Australia. J. Med. Entomol. 2014, 51, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Tesh, R.B.; Gubler, D.J.; Rosen, L. Variation among geographic strains of Aedes albopictus in susceptibility to infection with chikungunya virus. Am. J. Trop. Med. Hygiene 1976, 25, 326–335. [Google Scholar]
- Haddad, N.; Mousson, L.; Vazeille, M.; Chamat, S.; Tayeh, J.; Osta, M.A.; Failloux, A.B. Aedes albopictus in Lebanon, a potential risk of arboviruses outbreak. BMC Infect. Dis. 2012, 12, 300. [Google Scholar] [CrossRef] [PubMed]
- Talbalaghi, A.; Moutailler, S.; Vazeille, M.; Failloux, A.B. Are Aedes albopictus or other mosquito species from Northern Italy competent to sustain new arboviral outbreaks? Med. Vet. Entomol. 2010, 24, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Vazeille, M.; Jeannin, C.; Martin, E.; Schaffner, F.; Failloux, A.B. Chikungunya: A risk for Mediterranean countries? Acta Trop. 2008, 105, 200–202. [Google Scholar] [CrossRef] [PubMed]
- Moutailler, S.; Barre, H.; Vazeille, M.; Failloux, A.B. Recently introduced Aedes albopictus in Corsica is competent to chikungunya virus and in a lesser extent to dengue virus. Trop. Med. Int. Health 2009, 14, 1105–1109. [Google Scholar] [CrossRef] [PubMed]
- Kramer, L.D.; Chin, P.; Cane, R.P.; Kauffman, E.B.; Mackereth, G. Vector competence of New Zealand mosquitoes for selected arboviruses. Am. J. Trop. Med. Hygiene 2011, 85, 182–189. [Google Scholar] [CrossRef]
- Jupp, P.G.; McIntosh, B.M.; Dos Santos, I.; DeMoor, P. Laboratory vector studies on six mosquito and one tick species with chikungunya virus. Trans. R. Soc. Trop. Med. Hygiene 1981, 75, 15–19. [Google Scholar] [CrossRef]
- Ledermann, J.P.; Guillaumot, L.; Yug, L.; Saweyog, S.C.; Tided, M.; Machieng, P.; Pretrick, M.; Marfel, M.; Griggs, A.; Bel, M.; et al. Aedes hensilli as a potential vector of chikungunya and zika viruses. PLoS Negl. Trop. Dis. 2014, 8, e3188. [Google Scholar] [CrossRef] [PubMed]
- Diagne, C.T.; Faye, O.; Guerbois, M.; Knight, R.; Diallo, D.; Faye, O.; Ba, Y.; Dia, I.; Faye, O.; Weaver, S.C.; et al. Vector competence of Aedes aegypti and Aedes vittatus (diptera: Culicidae) from Senegal and Cape Verde archipelago for West African lineages of chikungunya virus. Am. J. Trop. Med. Hygiene 2014, 91, 635–641. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Coffey, L.L.; Failloux, A.-B.; Weaver, S.C. Chikungunya Virus–Vector Interactions. Viruses 2014, 6, 4628-4663. https://0-doi-org.brum.beds.ac.uk/10.3390/v6114628
AMA Style
Coffey LL, Failloux A-B, Weaver SC. Chikungunya Virus–Vector Interactions. Viruses. 2014; 6(11):4628-4663. https://0-doi-org.brum.beds.ac.uk/10.3390/v6114628
Chicago/Turabian StyleCoffey, Lark L., Anna-Bella Failloux, and Scott C. Weaver. 2014. "Chikungunya Virus–Vector Interactions" Viruses 6, no. 11: 4628-4663. https://0-doi-org.brum.beds.ac.uk/10.3390/v6114628