
Bacteriocins in the Era of Antibiotic Resistance: Rising to the Challenge

 , , and
, , and
Abstract
:1. Introduction
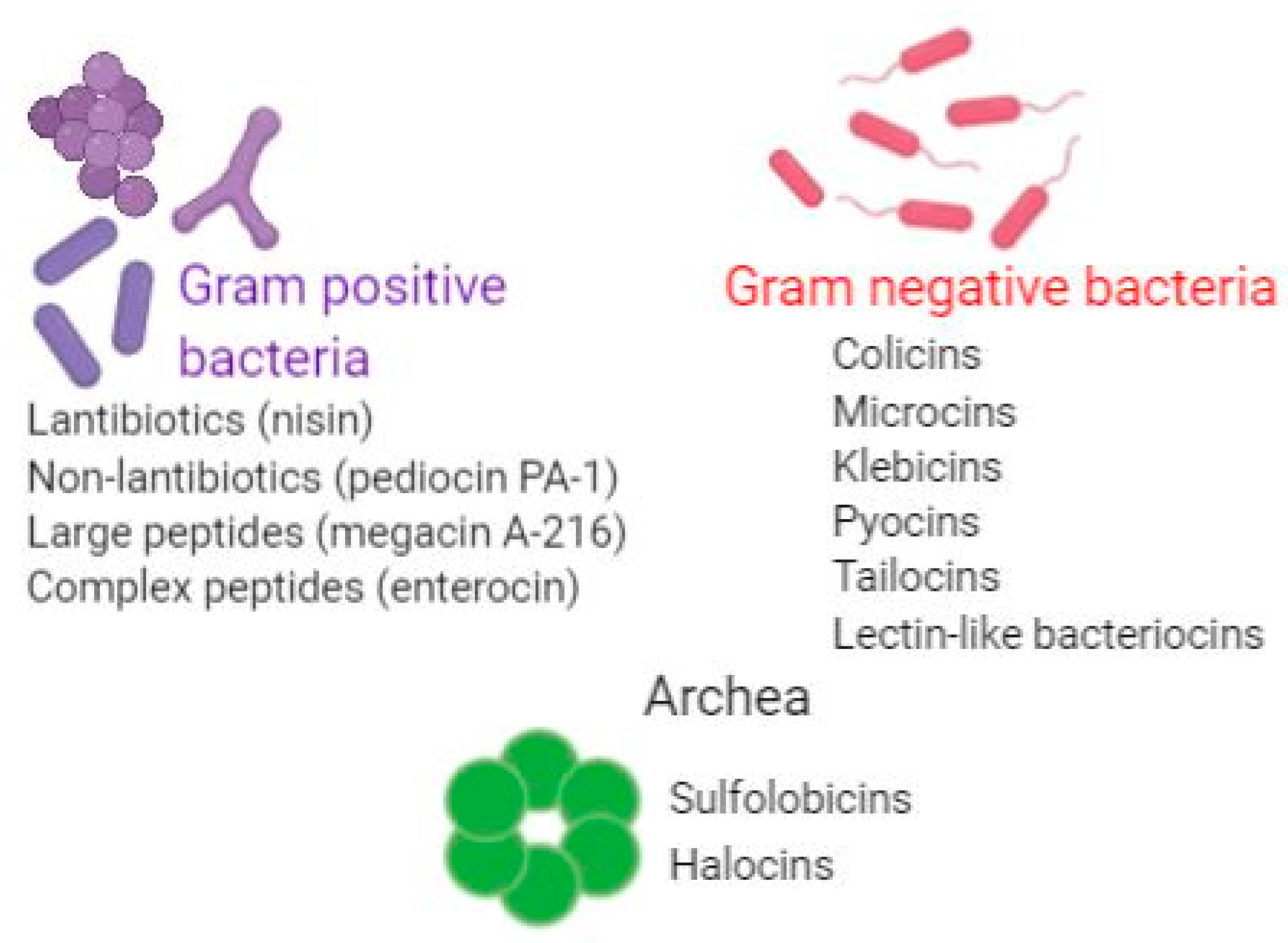
2. Bacteriocins: General Features
3. Bacteriocins to the Rescue in Microbial Infections
4. Bacteriocins Produced by Gram-Positive Bacteria
5. Bacteriocins Produced by Gram-Negative Bacteria
6. Bacteriocins Produced by Archaea
7. In Vivo Activity of Bacteriocins
8. Challenges and Opportunities
9. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Weist, K.; Diaz Hogberg, L. ECDC Publishes 2013 Surveillance Data on Antimicrobial Resistance and Antimicrobial Consumption in Europe. Eurosurveillance 2014, 19, 20962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santajit, S.; Indrawattana, N. Mechanisms of Antimicrobial Resistance in ESKAPE Pathogens. Biomed. Res. Int. 2016, 2016, 2475067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vivas, R.; Barbosa, T.; Dolabela, S.S.; Jain, S. Multidrug-Resistant Bacteria and Alternative Methods to Control Them: An Overview. Microb. Drug Resist. 2019, 25, 890–908. [Google Scholar] [CrossRef] [PubMed]
- Antibiotic Resistance Threats in the United States, 2019 (2019 AR Threats Report). Available online: www.cdc.gov/drugresistance/biggest-threats.html (accessed on 30 March 2020).
- Giedraitienė, A.; Vitkauskienė, A.; Naginienė, R.; Pavilonis, A. Antibiotic Resistance Mechanisms of Clinically Important Bacteria. Med. Kaunas 2011, 47, 137–146. [Google Scholar] [CrossRef]
- Navidinia, M. The Clinical Importance of Emerging ESKAPE Pathogens in Nosocomial Infections. J. Paramed Sci. 2016, 7, 2008–4978. [Google Scholar]
- Pitout, J.D.D.; Nordmann, P.; Poirel, L. Carbapenemase-Producing Klebsiella Pneumoniae, a Key Pathogen Set for Global Nosocomial Dominance. Antimicrob. Agents Chemother. 2015, 59, 5873–5884. [Google Scholar] [CrossRef] [Green Version]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, Research, and Development of New Antibiotics: The WHO Priority List of Antibiotic-Resistant Bacteria and Tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Rolain, J.-M.; Abat, C.; Jimeno, M.-T.; Fournier, P.-E.; Raoult, D. Do We Need New Antibiotics? Clin. Microbiol. Inf. 2016, 22, 408–415. [Google Scholar] [CrossRef]
- Fair, R.J.; Tor, Y. Antibiotics and Bacterial Resistance in the 21st Century. Perspect. Med. Chem. 2014, 6, 25–64. [Google Scholar] [CrossRef] [Green Version]
- Riley, M.A.; Robinson, S.M.; Roy, C.M.; Dorit, R.L. Rethinking the Composition of a Rational Antibiotic Arsenal for the 21st Century. Future Med. Chem. 2013, 5, 1231–1242. [Google Scholar] [CrossRef]
- Ghodhbane, H.; Elaidi, S.; Sabatier, J.M.; Achour, S.; Benhmida, J.; Regaya, I. Bacteriocins Active Against Multi-Resistant Gram—Negative Bacteria Implicated in Nosocomial Infections. Infect. Disord. Drug Targets 2015, 15, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Drissi, F.; Buffet, S.; Raoult, D.; Merhej, V. Common Occurrence of Antibacterial Agents in Human Intestinal Microbiota. Front. Microbiol. 2015, 6, 441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ditu, L.M.; Chifiriuc, M.C.; Pelinescu, D.; Avram, I.; Pircalabioru, G.; Mihaescu, G. Class I and II Bacteriocins: Structure, Biosynthesis and Drug Delivery Systems for the Improvement of Their Antimicrobial Activity. Curr. Proteom. 2014, 11, 121–127. [Google Scholar] [CrossRef]
- Ibrahim, O.O. Classification of Antimicrobial Peptides Bacteriocins, and the Nature of Some Bacteriocins with Potential Applications in Food Safety and Bio-Pharmaceuticals. EC Microbiol. 2019, 15, 591–608. [Google Scholar]
- Hols, P.; Ledesma-García, L.; Gabant, P.; Mignolet, J. Mobilization of Microbiota Commensals and Their Bacteriocins for Therapeutics. Trends Microbiol. 2019, 27, 690–702. [Google Scholar] [CrossRef] [Green Version]
- Belguesmia, Y.; Naghmouchi, K.; Chihib, N.-E.; Drider, D. Class IIa Bacteriocins: Current Knowledge and Perspectives. Prokaryotic Antimicrob. Pept. 2011, 171–195. [Google Scholar] [CrossRef]
- O’Shea, E.F.; O’Connor, P.M.; O’Sullivan, O.; Cotter, P.D.; Ross, R.P.; Hill, C. Bactofencin A, a New Type of Cationic Bacteriocin with Unusual Immunity. mBio 2013, 4, e00498–e00513. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Gutierrez, E.; Mayer, M.J.; Cotter, P.D.; Narbad, A. Gut Microbiota as a Source of Novel Antimicrobials. Gut Microbes 2019, 10, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Kumariya, R.; Kumari, G.; Raiput, Y.S.; Akhtar, N.; Patel, S. Bacteriocins: Classification, Synthesis, Mechanism of Action and Resistance Development in Food Spoilage Causing Bacteria. Microb. Pathog. 2019, 128, 171–177. [Google Scholar] [CrossRef]
- Suganthi, V.; Mohanasrinivasan, V. Optimization Studies for Enhanced Bacteriocin Production by Pediococcus Pentosaceus KC692718 Using Response Surface Methodology. J. Food Sci. Technol. 2015, 52, 3773–3783. [Google Scholar] [CrossRef]
- Duquesne, S.; Destoumieux-Garzon, D.; Peduzzi, J.; Rebuffat, S. Microcins, Gene-Encoded Antibacterial Peptides from Enterobacteria. Nat. Prod. Rep. 2007, 24, 708–734. [Google Scholar] [CrossRef] [PubMed]
- Baquero, F.; Lanza, V.F.; Baquero, M.-R.; del Campo, R.; Bravo-Vazquez, D.A. Microcins in Enterobacteriaceae: Peptide Antimicrobials in the Eco-Active Intestinal Chemosphere. Front. Microbiol. 2019, 10, 2261. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Li, N.; Zeng, X.; Liu, L.; Wang, Y.; Wang, G.; Cai, S.; Huang, S.; Ding, X.; Song, Q.; et al. A Comprehensive Antimicrobial Activity Evaluation of the Recombinant Microcin J25 Against the Foodborne Pathogens Salmonella and E. Coli O157:H7 by Using a Matrix of Conditions. Front. Microbiol. 2019, 10, 1954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abanoz, H.S.; Kunduhoglu, B. Antimicrobial Activity of a Bacteriocin Produced by Enterococcus Faecalis KT11 against Some Pathogens and Antibiotic-Resistant Bacteria. Korean J. Food Sci. Anim. Resour. 2018, 38, 1064–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etayash, H.; Azmi, S.; Dangeti, R.; Kaur, K. Peptide Bacteriocins–Structure Activity Relationships. Curr. Top. Med. Chem. 2015, 16, 220–241. [Google Scholar] [CrossRef]
- Dobson, A.; Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocin Production: A Probiotic Trait? Appl. Environ. Microbiol. 2012, 78, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Chikindas, M.L.; Weeks, R.; Drider, D.; Chistyakov, V.A.; Dicks, L.M. Functions and Emerging Applications of Bacteriocins. Curr. Opin. Biotechnol. 2017, 49, 23–38. [Google Scholar] [CrossRef]
- Weinstock, G.M. A Glimpse of Microbial Power in Preventive Medicine. JAMA Pediatr. 2016, 170, 11. [Google Scholar] [CrossRef]
- Meade, E.; Slattery, M.A.; Garvey, M. Bacteriocins, Potent Antimicrobial Peptides and the Fight against Multi Drug Resistant Species: Resistance Is Futile? Antibiotics 2020, 9, 32. [Google Scholar] [CrossRef] [Green Version]
- Field, D.; Begley, M.; O’Connor, P.M.; Daly, K.M.; Hugenholtz, F.; Cotter, P.D.; Hill, C.; Ross, R.P. Bioengineered Nisin A Derivatives with Enhanced Activity against Both Gram—Positive and Gram—Negative Pathogens. PLoS ONE 2012, 7, e46884. [Google Scholar] [CrossRef] [Green Version]
- Field, D.; Seisling, N.; Cotter, P.D.; Ross, R.P.; Hill, C. Synergistic Nisin-Polymyxin Combinations for the Control of Pseudomonas Biofilm Formation. Front. Microbiol. 2016, 7, 1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasamiravaka, T.; Labtani, Q.; Duez, P.; El-Jaziri, M. The Formation of Biofilms by Pseudomonas Aeruginosa: A Review of the Natural and Synthetic Compounds Interfering with Control Mechanisms. Biomed. Res. Int. 2015, 2015, 759348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giacometti, A.; Cirioni, O.; Barchiesi, F.; Scalise, G. In-Vitro Activity and Killing Effect of Polycationic Peptides on Methicillin-Resistant Staphylococcus Aureus and Interactions with Clinically Used Antibiotics. Diagn. Microbiol. Infect. Dis. 2000, 38, 115–118. [Google Scholar] [CrossRef]
- Brumfitt, W.; Salton, M.R.J.; Hamilton-Miller, J.M.T. Nisin, Alone and Combined with Peptidoglycan-Modulating Antibiotics: Activity against Methicillin-Resistant Staphylococcus Aureus and Vancomycin-Resistant Enterococci. J. Antimicrob. Chemother. 2002, 50, 731–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, J.M.; Gwak, J.W.; Kamarajan, P.; Fenno, J.C.; Rickard, A.H.; Kapila, Y.L. Biomedical Applications of Nisin. J. Appl. Microbiol. 2016, 120, 1449–1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Sullivan, J.N.; O’Connor, P.M.; Rea, M.C.; O’Sullivan, O.; Walsh, C.J.; Healy, B.; Mathur, H.; Field, D.; Hill, C.; Ross, R.P. Nisin J, a Novel Natural Nisin Variant, Is Produced by Staphylococcus Capitis Sourced from the Human Skin Microbiota Julie. J. Bacteriol. 2020, 202, e00639-19. [Google Scholar] [CrossRef] [Green Version]
- Sashihara, T.; Kimura, H.; Higuchi, T.; Adachi, A.; Matsusaki, H.; Sonomoto, K.; Ishizaki, A. A Novel Lantibiotic, Nukacin ISK-1, of Staphylococcus Warneri ISK-1: Cloning of the Structural Gene and Identification of the Structure. Biosci. Biotechnol. Biochem. 2000, 64, 2420–2428. [Google Scholar] [CrossRef]
- Okuda, K.; Zendo, T.; Sugimoto, S.; Iwase, T.; Tajima, A.; Yamada, S.; Sonomoto, K.; Mizunoe, Y. Effects of Bacteriocins on Methicillin-Resistant Staphylococcus Aureus. Antimicr. Agents Chemother. 2013, 57, 5572–5579. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, J.M.; Martinez, M.I.; Kok, J. Pediocin PA-1, a Wide-Spectrum Bacteriocin from Lactic Acid Bacteria. Crit. Rev. Food Sci. Nutr. 2002, 42, 91–121. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, J.N.; Rea, M.C.; O’Connor, P.M.; Hill, C.; Ross, R.P. Human Skin Microbiota Is a Rich Source of Bacteriocin-Producing Staphylococci That Kill Human Pathogens. FEMS Microbiol. Ecol. 2019, 95, fiy241. [Google Scholar] [CrossRef]
- Ansari, A.; Zohra, R.R.; Tarar, O.M.; Ali, S.; Qader, U.; Aman, A. Screening, Purification and Characterization of Thermostable, Protease Resistant Bacteriocin Active against Methicillin Resistant Staphylococcus Aureus ( MRSA). BMC Microbiol. 2018, 18, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Biswas, K.; Upadhayay, S.; Rapsang, G.F.; Joshi, S.R. Antibacterial and Synergistic Activity Against b -Lactamase-Producing Nosocomial Bacteria by Bacteriocin of LAB Isolated From Lesser Known Traditionally Fermented Products of India. HAYATI J. Biosci. 2017, 24, 87–95. [Google Scholar] [CrossRef]
- LeBlanc, J.G.; LeBlanc, A.M.; de Souza Oliviera, R.P.; Todorov, S.D. Use of Synbiotics (Probiotics and Prebiotics) to Improve the Safety of Foods. In Practical Food Safety-Contemporary Issues and Future Directions; Bhat, R., Gomez-Lopez, V.M., Eds.; Wiley: Chichester, UK, 2014; pp. 497–531. [Google Scholar]
- Hahn-Löbmann, S.; Stephan, A.; Schulz, S.; Schneider, T.; Shaverskyi, A.; Tusé, D.; Giritch, A.; Gleba, Y. Colicins and Salmocins—New Classes of Plant-Made Non-Antibiotic Food Antibacterials. Front. Plant Sci. 2019, 10, 437. [Google Scholar] [CrossRef] [PubMed]
- Cameron, A.; Zaheer, R.; Adator, E.H.; Barbieri, R.; Reuter, T.; McAllister, T.A. Bacteriocin Occurrence and Activity in Escherichia Coli Isolated from Bovines and Wastewater. Toxins 2019, 11, 475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behrens, H.M.; Six, A.; Walker, D.; Kleanthous, C. The Therapeutic Potential of Bacteriocins as Protein Antibiotics. Emerg. Top. Life Sci. 2017, 1, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Rendueles, O.; Beloin, C.; Latour-Lambert, P.; Ghigo, J.-M. A New Biofilm-Associated Colicin with Increased Efficiency against Biofilm Bacteria. ISME J. 2014, 8, 1275–1288. [Google Scholar] [CrossRef] [Green Version]
- Ghequire, M.G.K.; Öztürk, B.; De Mot, R. Lectin-Like Bacteriocins. Front Microbiol. 2018, 2706. [Google Scholar] [CrossRef] [Green Version]
- Yao, G.W.; Duarte, I.; Le, T.T.; Carmody, L.; LiPuma, J.J.; Young, R.G.C.F. A Broad-Host-Range Tailocin from Burkholderia Cenocepacia. Appl. Env. Microbiol. 2017, 83, e03414-16. [Google Scholar] [CrossRef] [Green Version]
- Ghequire, M.G.K.; De Mot, R. LlpB Represents a Second Subclass of Lectin-like Bacteriocins. Microb. Biotechnol. 2019, 12, 567–573. [Google Scholar] [CrossRef]
- Karthikeyan, P.; Bhat, S.G.; Chandrasekaran, M. Halocin SH10 Production by an Extreme Haloarchaeon Natrinema Sp. BTSH10 Isolated from Salt Pans of South India. Saudi J. Biol. Sci. 2013, 20, 205–212. [Google Scholar] [CrossRef] [Green Version]
- Charlesworth, J.C.; Burns, B.P. Untapped Resources: Biotechnological Potential of Peptides and Secondary Metabolites in Archaea. Archaea 2015, 2015, 282035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juturu, V.; Wu, J.C. Microbial Production of Bacteriocins: Latest Research Development and Applications. Biotechnol. Adv. 2018, 36, 2187–2200. [Google Scholar] [CrossRef] [PubMed]
- Jabe´s, D.; Brunati, C.; Candiani, G.; Riva, S.; Romano, G.; Donadio, S. Efficacy of the New Lantibiotic NAI-107 in Experimental Infections Induced by Multidrug Resistant Gram—Positive Pathogens. Antimicrob. Agents Chemother. 2011, 55, 1671–1676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piper, C.; Draper, L.A.; Cotter, P.D.; Paul Ross, R.; Hill, C. A Comparison of the Activities of Lacticin 3147 and Nisin against Drug-Resistant Staphylococcus Aureus and Enterococcus Species. J. Antimicrob. Chemother. 2009, 64, 546–551. [Google Scholar] [CrossRef] [Green Version]
- Corr, S.C.; Li, Y.; Riedel, C.U.; O’Toole, P.W.; Hill, C.; Gahan, C.G. Bacteriocin Production as a Mechanism for the Antiinfective Activity of Lactobacillus Salivarius UCC118. Proc. Natl. Acad. Sci. USA 2007, 104, 7617–7621. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Wang, Q.; Zeng, X.; Ye, Q.; Huang, S.; Yu, H.; Yang, T.; Qiao, S. Use of the Antimicrobial Peptide Sublancin with Combined Antibacterial and Immunomodulatory Activities to Protect against Methicillin-Resistant Staphylococcus Aureus Infection in Mice. J. Agric. Food Chem. 2017, 65, 8595–8605. [Google Scholar] [CrossRef]
- De Kwaadsteniet, M.; Doeschate, K.T.; Dicks, L.M. Nisin F in the Treatment of Respiratory Tract Infections Caused by Staphylococcus Aureus. Lett. Appl. Microbiol. 2009, 48, 65–70. [Google Scholar] [CrossRef]
- Scholl, D.; Martin, D.W., Jr. Antibacterial Efficacy of R-Type Pyocins towards Pseudomonas Aeruginosa in a Murine Peritonitis Model. Antimicrob. Agents Chemother. 2008, 52, 1647–1652. [Google Scholar] [CrossRef] [Green Version]
- Kommineni, S.; Bretl, D.J.; Lam, V.; Chakraborty, R.; Hayward, M.; Simpson, P.; Cao, Y.; Bousounis, P.; Kristich, C.J.; Salzman, N.H. Bacteriocin Production Augments Niche Competition by Enterococci in the Mammalian Gastrointestinal Tract. Nature 2015, 526, 719–722. [Google Scholar] [CrossRef] [Green Version]
- Gebhart, D.; Lok, S.; Clare, S.; Tomas, M.; Stares, M.; Scholl, D.; Donskey, C.J.; Lawley, T.D.; Govoni, G.R. A Modified R-Type Bacteriocin Specifically Targeting Clostridium Difficile Prevents Colonization of Mice without Affecting Gut Microbiota Diversity. mBio 2015, 6, e02368-14. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Mao, R.; Zhang, Y.; Teng, D.; Wang, X.; Xi, D.; Huang, J.; Wang, J. Biotechnical Paving of Recombinant Enterocin A as the Candidate of Anti-Listeria Agent. BMC Microbiol. 2014, 408, 22. [Google Scholar]
- Song, D.F.; Li, X.; Zhang, Y.H.; Zhu, M.Y.; Gu, Q. Mutational Analysis of Positively Charged Residues in the N-Terminal Region of the Class IIa Bacteriocin Pediocin PA-1. Lett. Appl. Microbiol. 2014, 376, 356–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Song, H.; Zheng, W. Improvement of Antimicrobial Activity of Pediocin PA-1 by Site- Directed Mutagenesis in C-Terminal Domain. Protein Pept. Lett. 2015, 22, 1007–1012. [Google Scholar] [CrossRef]
- Acuña, L.; Picariello, G.; Sesma, F.; Morero, R.D.; Bellomio, A. A New Hybrid Bacteriocin, Ent35–MccV, Displays Antimicrobial Activity against Pathogenic Gram—Positive and Gram—Negative Bacteria. FEBS Open Bio 2012, 2, 12–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francino, M.P. Antibiotics and the Human Gut Microbiome: Dysbioses and Accumulation of Resistances. Front. Microbiol. 2016, 6, 1543. [Google Scholar] [CrossRef] [Green Version]
- Inglis, R.F.; Scanlan, P.; Buckling, A. Iron Availability Shapes the Evolution of Bacteriocin Resistance in Pseudomonas Aeruginosa. ISME J. 2016, 10, 2060–2066. [Google Scholar] [CrossRef]
- Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocins—A Viable Alternative to Antibiotics? Nat. Rev. Microbiol. 2013, 11, 95–105. [Google Scholar] [CrossRef]
- Chifiriuc, M.C.; Grumezescu, A.M.; Lazar, V.; Bolocan, A.; Triaridis, S.; Grigore, R.; Bertesteanu, S. Contribution of Antimicrobial Peptides to the Development of New and Efficient Antimicrobial Strategies. Curr. Proteom. 2014, 11, 98–107. [Google Scholar] [CrossRef]
- do Carmo de Freire Bastos, M. Resistance to Bacteriocins Produced by Gram—Positive Bacteria. Microbiology 2014, 161, 683–700. [Google Scholar] [CrossRef] [Green Version]
- Fimland, G.; Eijsink, V.G.H.; Nissen-Mayer, J. Comparative Studies of Immunity Proteins of Pediocin-like Bacteriocins. Microbiology 2002, 148, 3661–3670. [Google Scholar] [CrossRef] [Green Version]
- Jarvis, B. Resistance to nisin and production of nisin-inactivating enzymes by several Bacillus species. J. Gen. Microbiol. 1967, 47, 33–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dicks, L.; Dreyer, L.; Smith, C.; van Staden, A.D. A Review: The Fate of Bacteriocins in the Human Gastro-Intestinal Tract: Do They Cross the Gut-Blood Barrier? Front. Microbiol. 2018, 9, 2297. [Google Scholar] [CrossRef] [PubMed]
- Maher, S.; McClean, S. Investigation of the Cytotoxicity of Eukaryotic and Prokaryotic Antimicrobial Peptides in Intestinal Epithelial Cells in Vitro. Biochem. Pharm. 2006, 71, 1289–1298. [Google Scholar] [CrossRef] [PubMed]
- Jasniewski, J.; Cailliez-Grimal, C.; Chevalot, I.; Milliere, J.B.; Revol-Junelles, A.M. Interactions between Two Carnobacteriocins Cbn BM1 and Cbn B2 from Carnobacterium Maltaromaticum CP5 on Target Bacteria and Caco-2 Cells. Food Chem. Toxicol. 2009, 47, 893–897. [Google Scholar] [CrossRef] [PubMed]
- Tabata, A.; Yamada, T.; Ohtani, H.; Ohkura, K.; Tomoyasu, T.; Nagamune, H. β-Hemolytic Streptococcus Anginosus Subsp. Anginosus Causes Streptolysin S-Dependent Cytotoxicity to Human Cell Culture Lines in Vitro. J. Oral. Microbiol. 2019, 11, 1609839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hetz, C.; Bono, M.R.; Barros, L.F.; Lagos, R. Microcin E492, a Channel-Forming Bacteriocin from Klebsiella Pneumoniae, Induces Apoptosis in Some Human Cell Lines. Proc. Natl. Acad. Sci. USA 2002, 99, 2696–2701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaucher, R.A.; Teixeira, M.L.; Brandelli, A. Investigation of the Cytotoxicity of Antimicrobial Peptide P40 on Eukaryotic Cells. Curr. Microbiol. 2010, 60, 1–5. [Google Scholar] [CrossRef]
- Bird, T.J.; Grieble, H.G. Pyocin Antibiosis in Chick Embryos. Antimicrob. Agents Chemother. 1969, 9, 495–498. [Google Scholar]
- Urbán, P.; Valle-Delgado, J.J.; Moles, E.; Marques, J.; Díez, C.; Fernandez-Busquets, X. Nanotools for the Delivery of Antimicrobial Peptides. Curr. Drug Targets 2012, 13, 1158–1172. [Google Scholar] [CrossRef]
- Fahim, H.A.; Khairalla, A.S.; El-Gendy, A.O. Nanotechnology: A Valuable Strategy to Improve Bacteriocin Formulations. Front. Microbiol. 2016, 7, 1385. [Google Scholar] [CrossRef] [Green Version]
- Brandelli, A. Nanostructures as Promising Tools for Delivery of Antimicrobial Peptides. Mini Rev. Med. Chem. 2012, 12, 731–741. [Google Scholar] [CrossRef] [PubMed]
- Vemuri, S.; Rhodes, C.T. Preparation and Characterization of Liposomes as Therapeutic Delivery Systems: A Review. Pharm. Acta Helv. 1995, 70, 95–111. [Google Scholar] [CrossRef]
- Mugabe, C.; Azghani, A.O.; Omri, A. Liposome-Mediated Gentamicin Delivery: Development and Activity against Resistant Strains of Pseudomonas Aeruginosa Isolated from Cystic Fibrosis Patients. J. Antimicrob. Chemother. 2005, 55, 269–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malheiros Pda, S.; Sant’Anna, V.; Barbosa Mde, S.; Brandelli, A.; Franco, B.D. Effect of Liposome-Encapsulated Nisin and Bacteriocin-like Substance P34 on Listeria Monocytogenes Growth in Minas Frescal Cheese. Int. J. Food Microbiol. 2012, 156, 272–277. [Google Scholar] [CrossRef]
- Colas, J.-C.; Shi, W.; Malleswara Rao, V.S.N.; Omri, A.; Reza Mozafari, M.; Singh, H. Microscopical Investigations of Nisin-Loaded Nanoliposomes Prepared by Mozafari Method and Their Bacterial Targeting. Micron 2007, 38, 841–847. [Google Scholar] [CrossRef]
- de Mello, M.B.; da Silva Malheiros, P.; Brandelli, A.; Pesce da Silveira, N.; Jantzen, M.M.; de Sousa da Motta, A. Characterization and Antilisterial Effect of Phosphatidylcholine Nanovesicles Containing the Antimicrobial Peptide Pediocin. Probiotics Antimicrob. Proteins 2013, 5, 43–50. [Google Scholar] [CrossRef]
- Feng, L.; Mumper, R.J. A Critical Review of Lipid-Based Nanoparticles for Taxane Delivery. Cancer Lett. 2013, 334, 157–175. [Google Scholar] [CrossRef] [Green Version]
- Prombutara, P.; Kulwatthanasal, Y.; Supaka, N.; Sramala, I.; Chareonpornwattana, S. Production of Nisin-Loaded Solid Lipid Nanoparticles for Sustained Antimicrobial Activity. Food Control 2012, 24, 184–190. [Google Scholar] [CrossRef]
- Zohri, M.; Shafiee Alavidjeh, M.; Haririan, I.; Ardestani, M.S.; Ebrahimi, S.E.S.; Sani, H.T.; Sadjadi, S.K. A Comparative Study between the Antibacterial Effect of Nisin and Nisin-Loaded Chitosan/Alginate Nanoparticles on the Growth of Staphylococcus Aureus in Raw and Pasteurized Milk Samples. Probiotics Antimicrob. Proteins 2010, 2, 258–266. [Google Scholar] [CrossRef]
- Zohri, M.; Shafiee Alavidjeh, M.; Mirdamadi, S.S.; Behmadi, H.; Hossaini Nasr, S.; Eshghi Gonbaki, S.; Shafiee Ardestani, M.; Jabbari Arabzadeh, A. Nisin−Loaded Chitosan/Alginate Nanoparticles: A Hopeful Hybrid Biopreservative. J. Food Saf. 2013, 33, 40–49. [Google Scholar] [CrossRef]
- Heunis, T.D.; Smith, C.; Dicks, L. Evaluation of a Nisin-Eluting Nanofiber Scaffold to Treat Staphylococcus Aureus-Induced Skin Infections in Mice. Antimicrob. Agents Chemother. 2013, 57, 3928–3935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahire, J.J.; Dicks, L.M. Nisin Incorporated with 2, 3-Dihydroxybenzoic Acid in Nanofibers Inhibits Biofilm Formation by a Methicillin-Resistant Strain of Staphylococcus Aureus. Probiotics Antimicrob. Proteins 2015, 7, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Dawson, M.J.; Scott, R.W. New Horizons for Host Defense Peptides and Lantibiotics. Curr. Opin Pharm. 2012, 12, 545–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Class | Subclass | Examples | Characteristics | Antimicrobial Spectrum | Mechanisms of Action | References |
|---|---|---|---|---|---|---|
| I | Ia (lantibiotics) | Nisin, lacticin 481, lactosin S, carnocin U149, subtilin subtilosin A Mersacidin | small membrane-active, proteolysis- and heat-resistant peptides (<5 kDa) | MRSA, Listeria spp., Streptococcus sp. Clostridium difficile, Bacillus, Enterococcus, C. albicans | Pore formation Cell wall synthesis | [16] |
| Ib(labyrinthopeptins) | ||||||
| Ic (sanctibiotics) | ||||||
| II | IIa (pediocin-like bacteriocins), IIb (two-peptides unmodified bacteriocins), IIc (circular bacteriocins) IId (unmodified, linear, nonpediocin-like bacteriocins) | Pediocin PA-I, pediocin AcH, enterocin A Uberolysin, carnocyclin, circularin A and AS-48, Grassericin A/reutericin A | heat-stable, pH- resistant, nonmodified, small peptides (<10 kDa) | E. coli, Listeria monocytogenes, Staphylococcus epidermidis, Serratia marcescens, K. pneumoniae, MRSA | Pore formation | [14,15,16,17] |
| III | Gram positive: lysostaphin, lactacin A and B helveticin V-1829, helveticin J, helveticin M acidophilus A | large heat-labile proteins (with a molecular weight higher than 10 kDa | S. aureus, S. saprophyticus, Enterobacter cloacae Gardnerella vaginalis, Streptococcus agalactiae, P. aeruginosa | Pore formation | [15,16,19,20] | |
| Gram negative: pyocin salmocins | P. aeruginosa Salmonella sp. STEC | Pore formation | [22,23,24] | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gradisteanu Pircalabioru, G.; Popa, L.I.; Marutescu, L.; Gheorghe, I.; Popa, M.; Czobor Barbu, I.; Cristescu, R.; Chifiriuc, M.-C. Bacteriocins in the Era of Antibiotic Resistance: Rising to the Challenge. Pharmaceutics 2021, 13, 196. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13020196
Gradisteanu Pircalabioru G, Popa LI, Marutescu L, Gheorghe I, Popa M, Czobor Barbu I, Cristescu R, Chifiriuc M-C. Bacteriocins in the Era of Antibiotic Resistance: Rising to the Challenge. Pharmaceutics. 2021; 13(2):196. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13020196
Chicago/Turabian StyleGradisteanu Pircalabioru, Gratiela, Laura Ioana Popa, Luminita Marutescu, Irina Gheorghe, Marcela Popa, Ilda Czobor Barbu, Rodica Cristescu, and Mariana-Carmen Chifiriuc. 2021. "Bacteriocins in the Era of Antibiotic Resistance: Rising to the Challenge" Pharmaceutics 13, no. 2: 196. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13020196