
Increased In Vitro Intercellular Barrier Function of Lung Epithelial Cells Using Adipose-Derived Mesenchymal Stem/Stromal Cells



 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Culture of AD-MSCs
2.2. Isolation and Culture of BM-MSCs
2.3. Isolation of Lung Cells
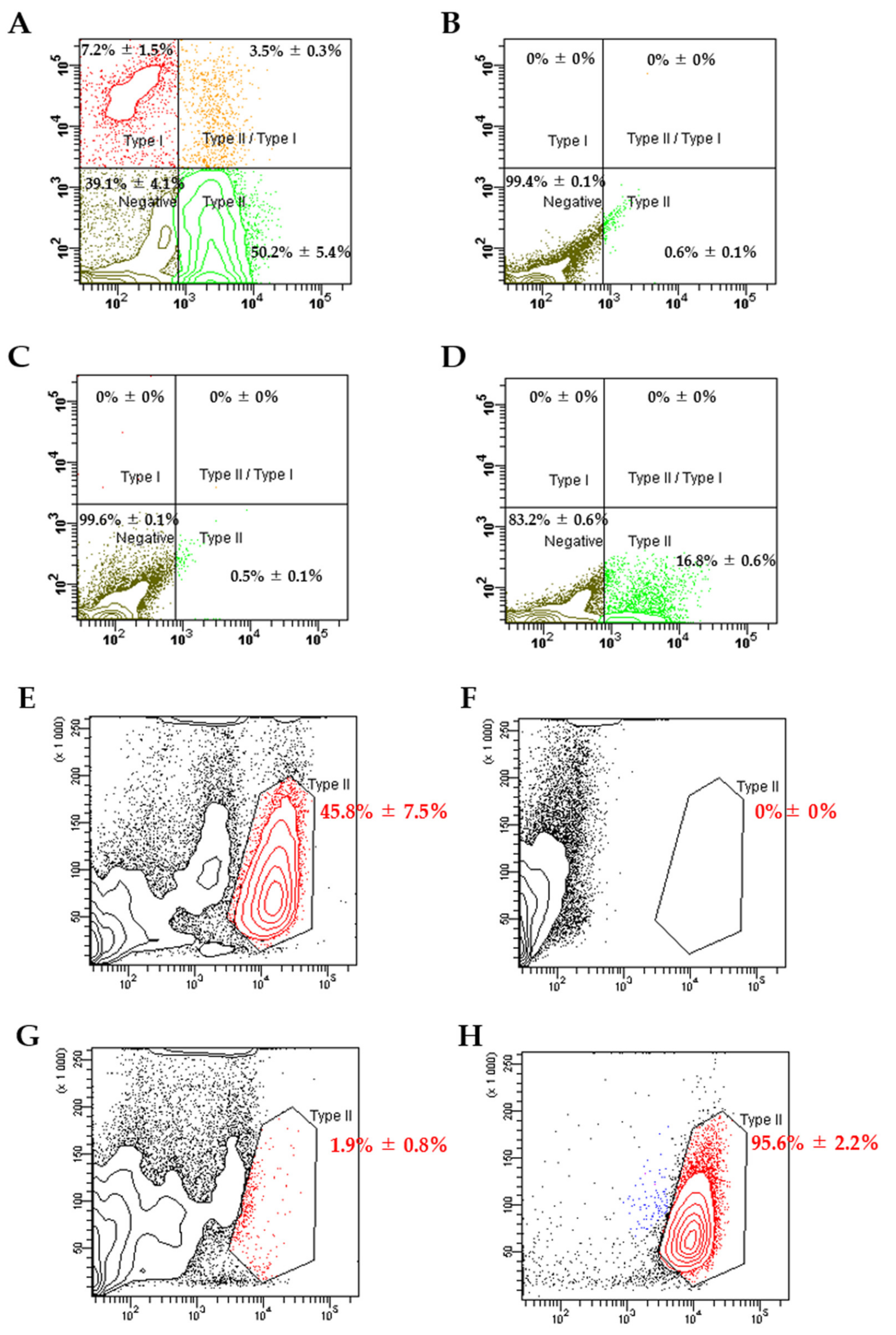
2.4. Flow Cytometry Analysis and Sorting of Alveolar Epithelial Type II (ATII) Cells
2.5. Transepithelial Electrical Resistance (TEER) Measurement
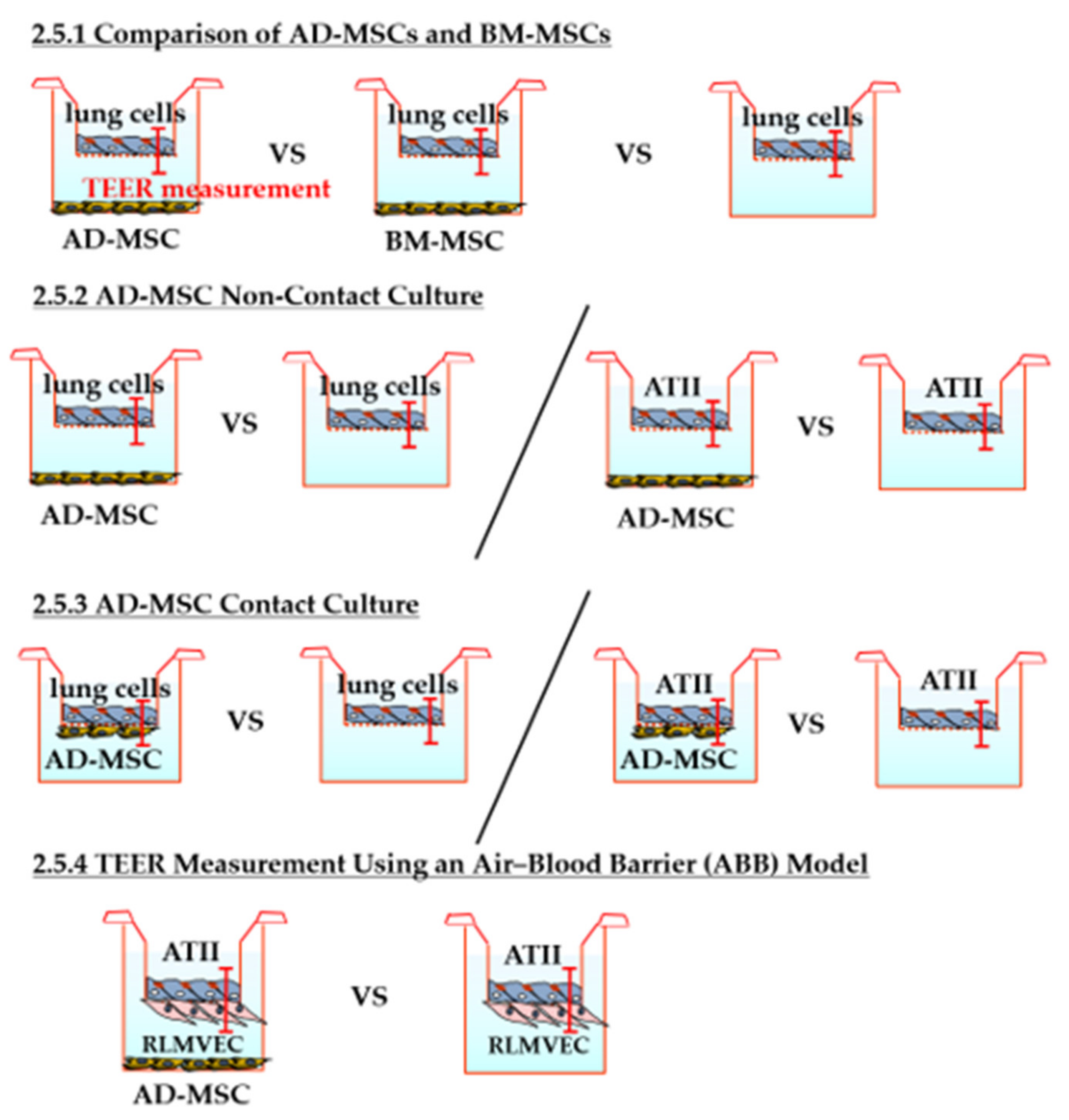
2.5.1. Comparison of AD-MSCs and BM-MSCs
2.5.2. TEER Measurement in AD-MSC Non-Contact Culture
2.5.3. TEER Measurement in AD-MSC Contact Culture
2.5.4. TEER Measurement Using an Air–Blood Barrier (ABB) Model
2.6. Permeability Assay
2.7. Hematoxylin and Eosin (HE) Staining of Insert Membranes
2.8. Immunostaining of Insert Membranes
2.9. Transmission Electron Microscopy
2.10. RNA Extraction, Reverse Transcription, and Quantitative Polymerase Chain Reaction (qPCR)
2.11. Statistical Analysis
3. Results
3.1. Flow Cytometry Analysis and Sorting of ATII Cells
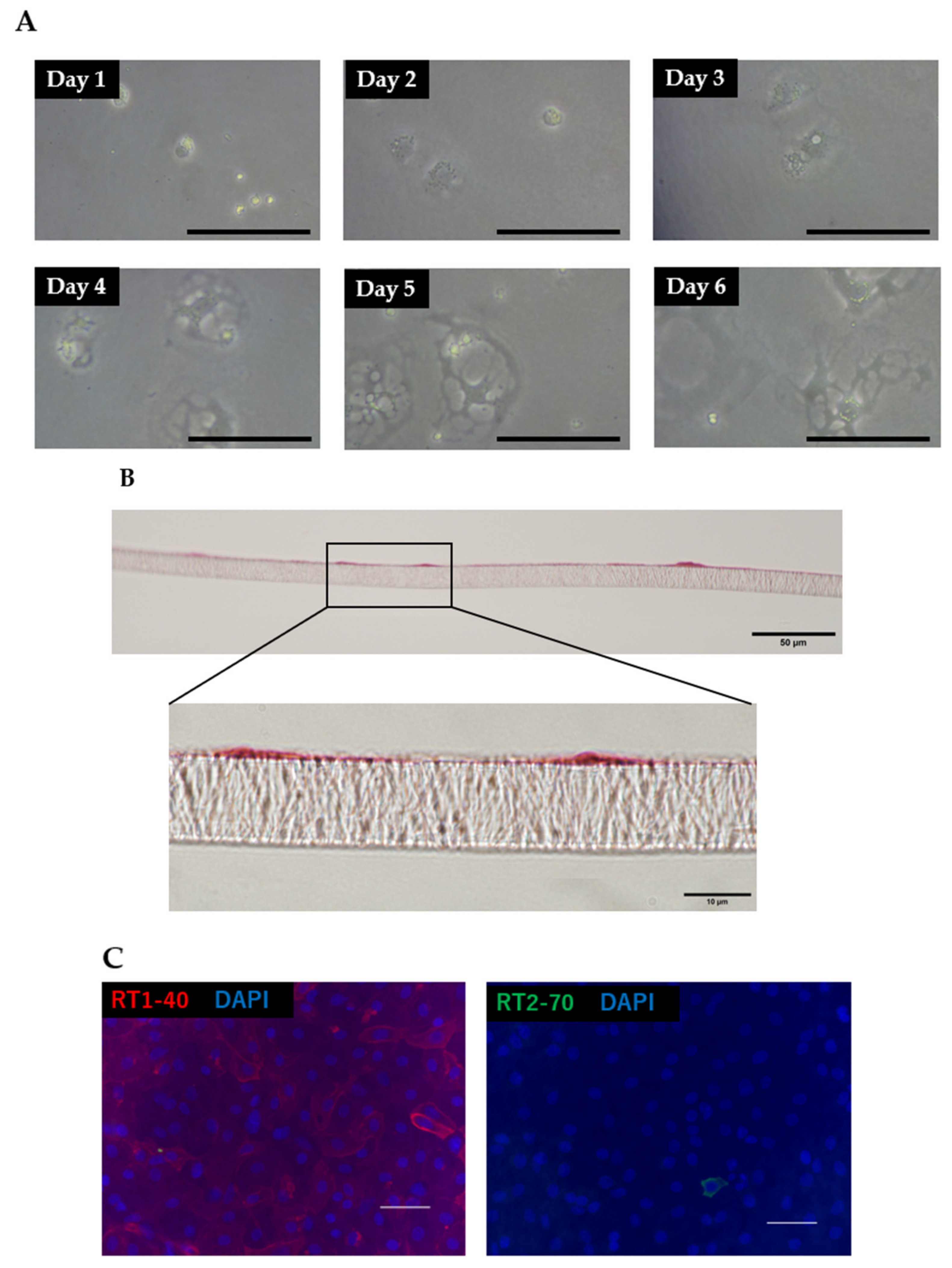
3.2. Morphology and Phenotypic Characteristics of ATII Cells
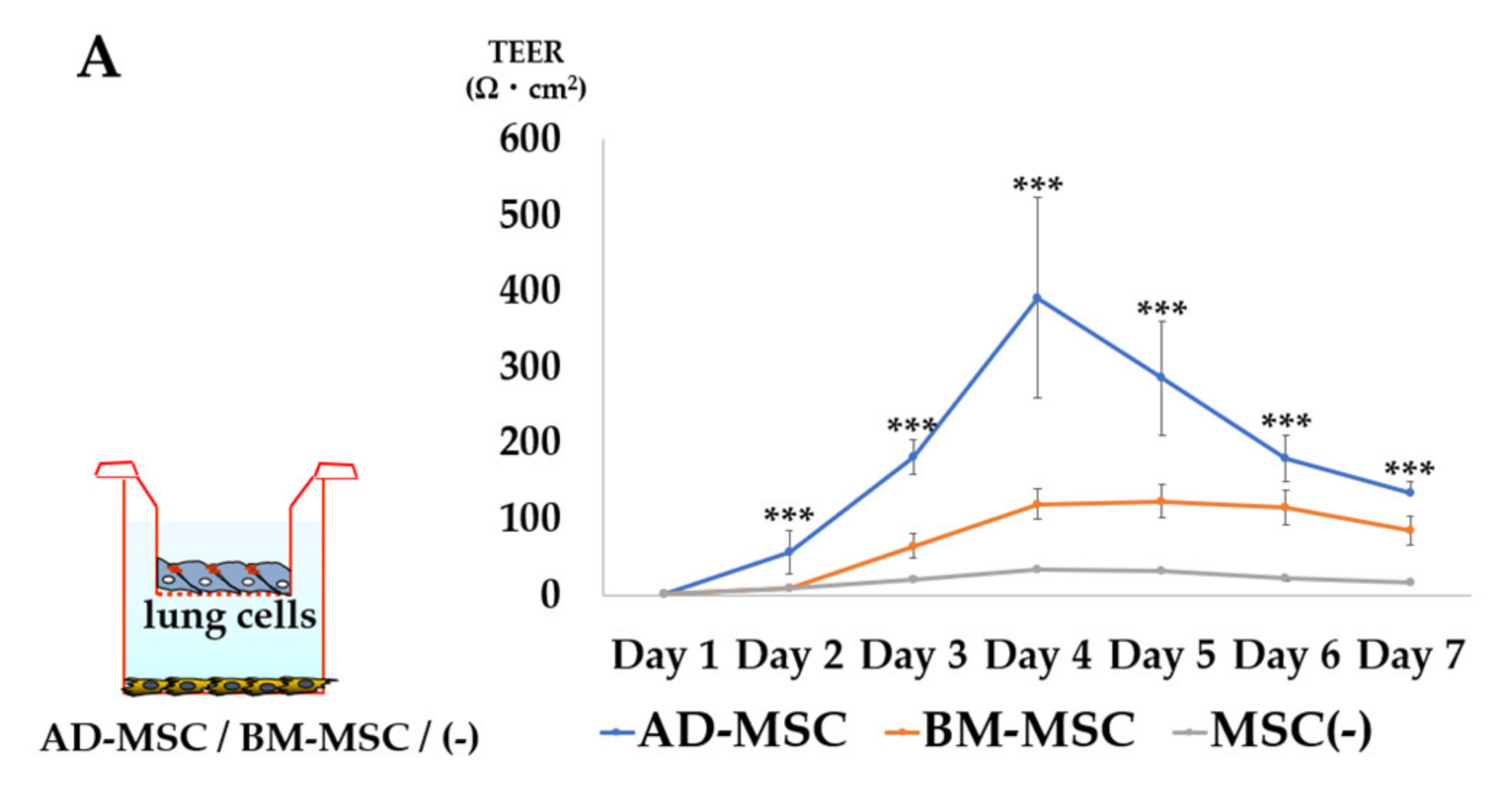
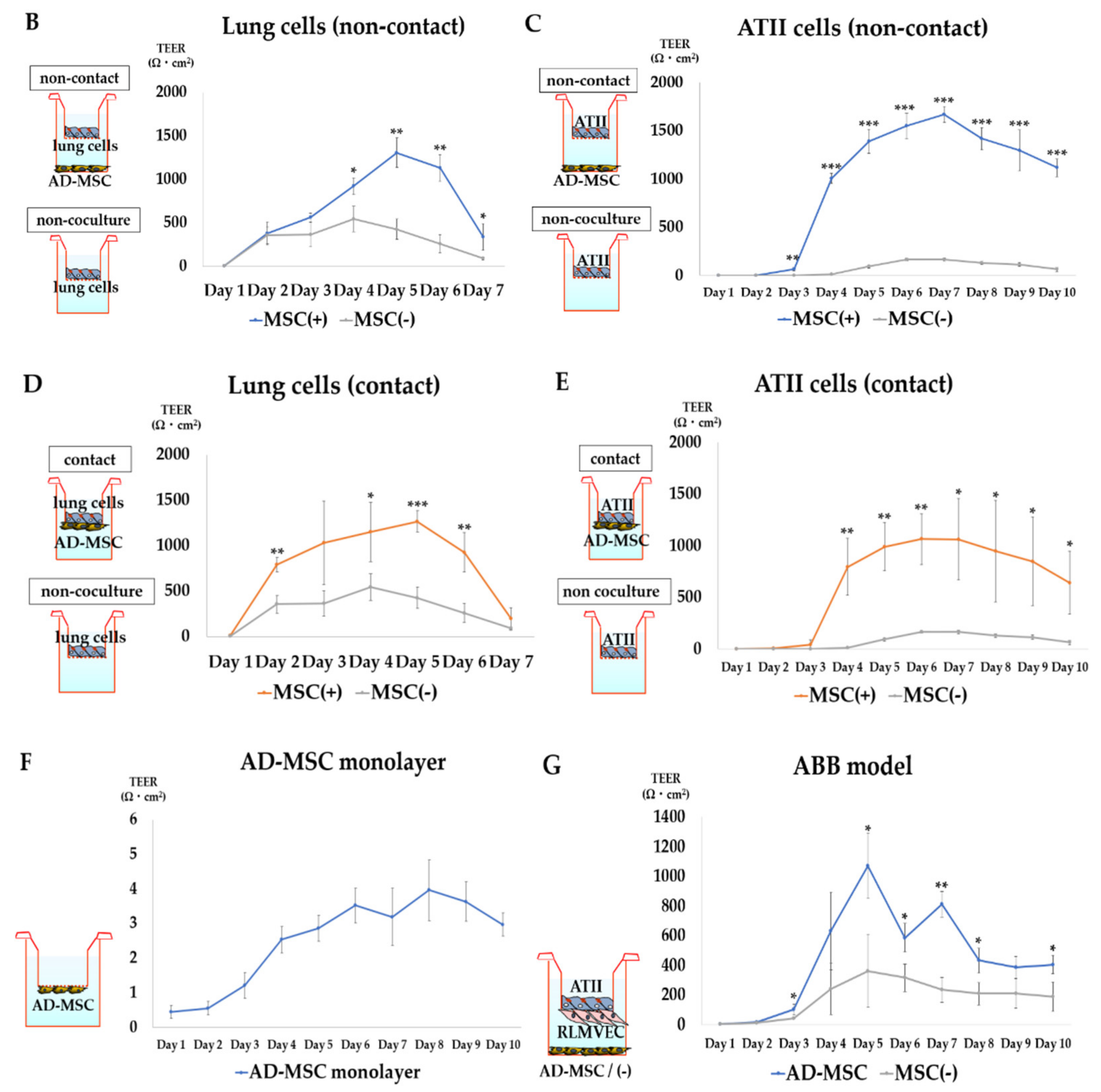
3.3. TEER Measurement
3.3.1. Comparison of AD-MSCs and BM-MSCs
3.3.2. TEER Measurement in AD-MSC Non-Contact Culture
3.3.3. TEER Measurement in AD-MSC Contact Culture
3.3.4. TEER Measurement in the ABB Model
3.4. Permeability Assay
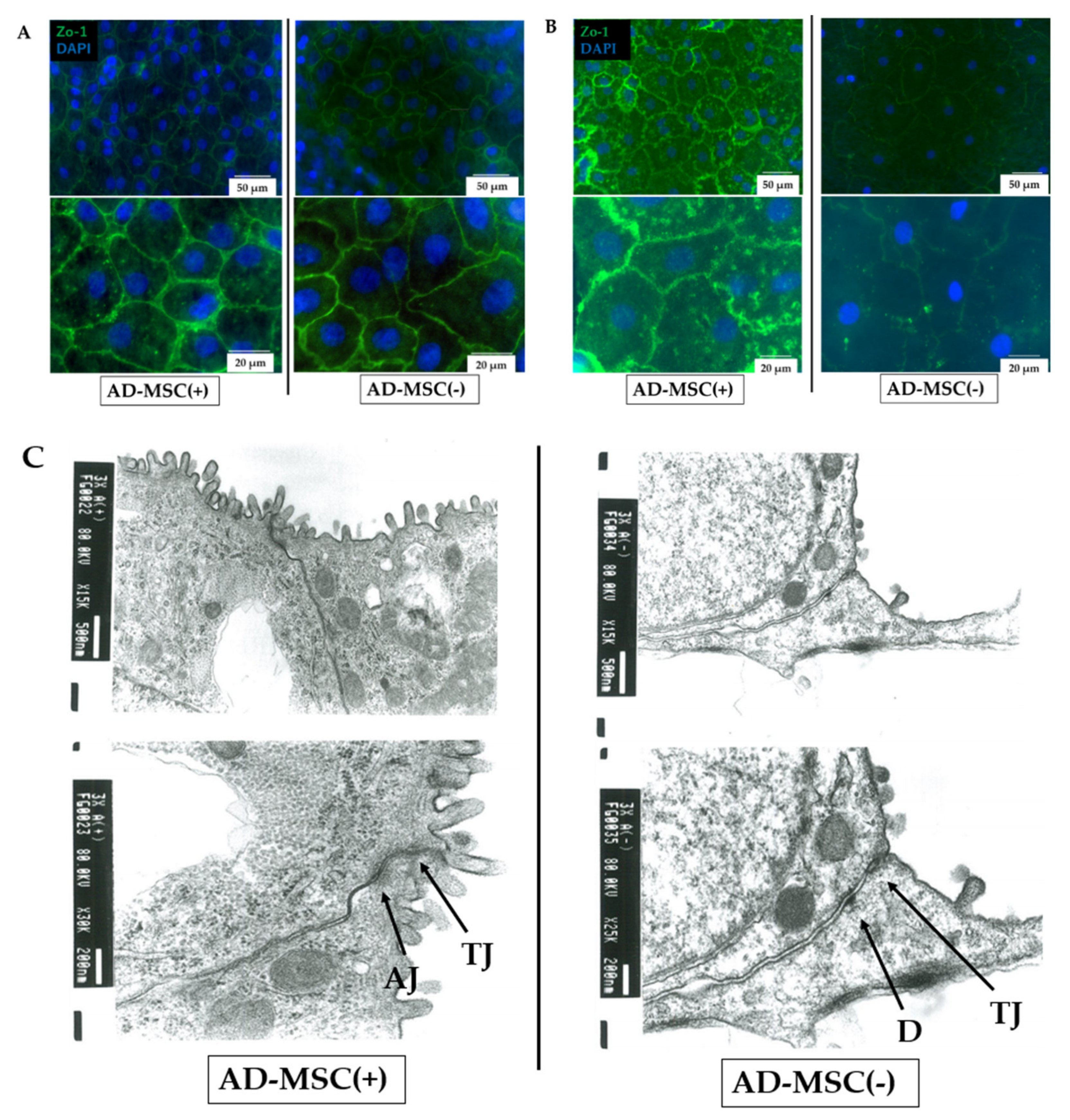
3.5. Immunostaining and Transmission Electron Microscopy of Cell Junctions
3.6. PCR Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Metcalfe, S.M. Mesenchymal stem cells and management of COVID-19 pneumonia. Med. Drug Discov. 2020, 5, 100019. [Google Scholar] [CrossRef] [PubMed]
- Bellani, G.; Laffey, J.G.; Pham, T.; Fan, E.; Brochard, L.; Esteban, A.; Gattinoni, L.; Van Haren, F.; Larsson, A.; McAuley, D.F.; et al. Epidemiology, Patterns of Care, and Mortality for Patients With Acute Respiratory Distress Syndrome in Intensive Care Units in 50 Countries. JAMA 2016, 315, 788–800. [Google Scholar] [CrossRef] [PubMed]
- Mac Sweeney, R.; McAuley, D.F. Acute respiratory distress syndrome. Lancet 2016, 388, 2416–2430. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Xiong, Y.; Xia, Y.; Zhang, R.; Tian, D.; Wang, T.; Dai, J.; Wang, L.; Yao, H.; Jiang, H.; et al. Therapeutic Effects of Human Umbilical Cord-Derived Mesenchymal Stem Cells in Acute Lung Injury Mice. Sci. Rep. 2017, 7, 39889. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Danchuk, S.D.; Bonvillain, R.W.; Xu, B.; Scruggs, B.A.; Strong, A.; Semon, J.A.; Gimble, J.M.; Betancourt, A.M.; Sullivan, D.E.; et al. Interleukin 6 Mediates the Therapeutic Effects of Adipose-Derived Stromal/Stem Cells in Lipopolysaccharide-Induced Acute Lung Injury. Stem Cells 2014, 32, 1616–1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, J.D.; Lopes-Pacheco, M.; De Castro, L.L.; Kitoko, J.; Trivelin, S.A.; Amorim, N.R.; Capelozzi, V.L.; Morales, M.M.; Gutfilen, B.; De Souza, S.A.L.; et al. Eicosapentaenoic acid potentiates the therapeutic effects of adipose tissue-derived mesenchymal stromal cells on lung and distal organ injury in experimental sepsis. Stem Cell Res. Ther. 2019, 10, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-Y.; Kim, D.H.; Lee, D.; Jeon, H.B.; Kwon, S.-J.; Kim, S.M.; Yoo, Y.J.; Lee, E.H.; Choi, S.J.; Seo, S.W.; et al. Soluble intracellular adhesion molecule-1 secreted by human umbilical cord blood-derived mesenchymal stem cell reduces amyloid-β plaques. Cell Death Differ. 2011, 19, 680–691. [Google Scholar] [CrossRef] [Green Version]
- Danchuk, S.; Ylostalo, J.H.; Hossain, F.; Sorge, R.; Ramsey, A.; Bonvillain, R.W.; Lasky, J.; Bunnell, B.; Welsh, D.; Prockop, D.J.; et al. Human multipotent stromal cells attenuate lipopolysaccharide-induced acute lung injury in mice via secretion of tumor necrosis factor-α-induced protein 6. Stem Cell Res. Ther. 2011, 2, 27. [Google Scholar] [CrossRef] [Green Version]
- Németh, K.; Leelahavanichkul, A.; Yuen, P.; Mayer, B.; Parmelee, A.; Doi, K.; Robey, P.; Leelahavanichkul, K.; Koller, B.H.; Brown, J.M.; et al. Bone marrow stromal cells attenuate sepsis via prostaglandin E2–dependent reprogramming of host macrophages to increase their interleukin-10 production. Nat. Med. 2008, 15, 42–49. [Google Scholar] [CrossRef] [Green Version]
- El-Sayed, M.; El-Feky, M.A.; El-Amir, M.I.; Hasan, A.S.; Tag-Adeen, M.; Urata, Y.; Goto, S.; Luo, L.; Yan, C.; Li, T.-S. Immunomodulatory effect of mesenchymal stem cells: Cell origin and cell quality variations. Mol. Biol. Rep. 2019, 46, 1157–1165. [Google Scholar] [CrossRef]
- Zhan, X.-S.; El-Ashram, S.; Luo, D.-Z.; Luo, H.-N.; Wang, B.-Y.; Chen, S.-F.; Bai, Y.-S.; Chen, Z.-S.; Liu, C.-Y.; Ji, H.-Q. A Comparative Study of Biological Characteristics and Transcriptome Profiles of Mesenchymal Stem Cells from Different Canine Tissues. Int. J. Mol. Sci. 2019, 20, 1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, R.; Li, M.; Luo, M.; Teng, T.; Pan, Y.; Huang, H. Mesenchymal stem cells decrease blood–brain barrier permeability in rats with severe acute pancreatitis. Cell. Mol. Biol. Lett. 2019, 24, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Ma, Y.; Du, B.; Wang, Y.; Yang, G.-Y.; Bi, X. Mesenchymal Stem Cells Attenuated Blood-Brain Barrier Disruption via Downregulation of Aquaporin-4 Expression in EAE Mice. Mol. Neurobiol. 2020, 57, 3891–3901. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-Y.; Xu, Q.-W.; Xu, P.-Y.; Li, W.-M. MSC-derived exosomal miR-34a/c-5p and miR-29b-3p improve intestinal barrier function by targeting the Snail/Claudins signaling pathway. Life Sci. 2020, 257, 118017. [Google Scholar] [CrossRef]
- Wang, H.; Zheng, R.; Chen, Q.; Shao, J.; Yu, J.; Hualing, W. Mesenchymal stem cells microvesicles stabilize endothelial barrier function partly mediated by hepatocyte growth factor (HGF). Stem Cell Res. Ther. 2017, 8, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, S.; Guo, F.; Zhang, X.; Chang, W.; Peng, F.; Qiu, H.; Yang, Y. mTOR/STAT-3 pathway mediates mesenchymal stem cell–secreted hepatocyte growth factor protective effects against lipopolysaccharide-induced vascular endothelial barrier dysfunction and apoptosis. J. Cell. Biochem. 2019, 120, 3637–3650. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, Q.-H.; Liu, A.-R.; Xu, X.-P.; Han, J.-B.; Qiu, H.-B. Synergism of MSC-secreted HGF and VEGF in stabilising endothelial barrier function upon lipopolysaccharide stimulation via the Rac1 pathway. Stem Cell Res. Ther. 2015, 6, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, S.; Park, J.; Liu, A.; Lee, J.; Zhang, X.; Hao, Q.; Lee, J.-W. Mesenchymal Stem Cell Microvesicles Restore Protein Permeability Across Primary Cultures of Injured Human Lung Microvascular Endothelial Cells. Stem Cells Transl. Med. 2018, 7, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Matthay, M.A.; Thompson, B.T.; Read, E.J.; McKenna, D.H.; Liu, K.D.; Calfee, C.S.; Lee, J.W. Therapeutic Potential of Mesenchymal Stem Cells for Severe Acute Lung Injury. Chest 2010, 138, 965–972. [Google Scholar] [CrossRef] [Green Version]
- Doi, R.; Tsuchiya, T.; Mitsutake, N.; Nishimura, S.; Matsuu-Matsuyama, M.; Nakazawa, Y.; Ogi, T.; Akita, S.; Yukawa, H.; Baba, Y.; et al. Transplantation of bioengineered rat lungs recellularized with endothelial and adipose-derived stromal cells. Sci. Rep. 2017, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Wiener-Kronish, J.P.; Albertine, K.H.; Matthay, M. Differential responses of the endothelial and epithelial barriers of the lung in sheep to Escherichia coli endotoxin. J. Clin. Investig. 1991, 88, 864–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorin, A.B.; Stewart, P.A. Differential permeability of endothelial and epithelial barriers to albumin flux. J. Appl. Physiol. 1979, 47, 1315–1324. [Google Scholar] [CrossRef]
- Zuk, P.A.; Zhu, M.; Mizuno, H.; Huang, J.; Futrell, J.W.; Katz, A.J.; Benhaim, P.; Lorenz, H.P.; Hedrick, M.H. Multilineage Cells from Human Adipose Tissue: Implications for Cell-Based Therapies. Tissue Eng. 2001, 7, 211–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elbert, K.J.; Schäfer, U.F.; Schäfers, H.; Kim, K.; Lee, V.H.L.; Lehr, C. Monolayers of Human Alveolar Epithelial Cells in Primary Culture for Pulmonary Absorption and Transport Studies. Pharm. Res. 1999, 16, 601–608. [Google Scholar] [CrossRef]
- Dagenais, A.; Desjardins, J.; Shabbir, W.; Roy, A.; Filion, D.; Sauvé, R.; Berthiaume, Y. Loss of barrier integrity in alveolar epithelial cells downregulates ENaC expression and activity via Ca2+ and TRPV4 activation. Eur. J. Physiol. 2018, 470, 1615–1631. [Google Scholar] [CrossRef]
- Friedenstein, A.J.; Chailakhjan, R.K.; Lalykina, K.S. The development of fibroblast colonies in monolayer cultures of guinea-pig bone marrow and spleen cells. Cell Prolif. 1970, 3, 393–403. [Google Scholar] [CrossRef]
- Zuk, P.A.; Zhu, M.; Ashjian, P.; De Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P.; Hedrick, M.H. Human Adipose Tissue Is a Source of Multipotent Stem Cells. Mol. Biol. Cell 2002, 13, 4279–4295. [Google Scholar] [CrossRef] [PubMed]
- Miura, M.; Gronthos, S.; Zhao, M.; Lu, B.; Fisher, L.W.; Robey, P.; Shi, S. SHED: Stem cells from human exfoliated deciduous teeth. Proc. Natl. Acad. Sci. USA 2003, 100, 5807–5812. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.W.; Choi, J.; Yang, M.S.; Moon, Y.J.; Park, J.S.; Kim, H.C.; Kim, Y.J. Mesenchymal stem cells from cryopreserved human umbilical cord blood. Biochem. Biophys. Res. Commun. 2004, 320, 273–278. [Google Scholar] [CrossRef]
- In’t Anker, P.S.; Scherjon, S.A.; Der Keur, C.K.-V.; De Groot-Swings, G.M.; Claas, F.H.; Fibbe, W.E.; Kanhai, H.H. Isolation of Mesenchymal Stem Cells of Fetal or Maternal Origin from Human Placenta. Stem Cells 2004, 22, 1338–1345. [Google Scholar] [CrossRef]
- Prockop, D.J.; Kota, D.J.; Bazhanov, N.; Reger, R.L. Evolving paradigms for repair of tissues by adult stem/progenitor cells (MSCs). J. Cell. Mol. Med. 2010, 14, 2190–2199. [Google Scholar] [CrossRef] [PubMed]
- Mohammadipoor, A.; Antebi, B.; Batchinsky, A.I.; Cancio, L.C. Therapeutic potential of products derived from mesenchymal stem/stromal cells in pulmonary disease. Respir. Res. 2018, 19, 1–14. [Google Scholar] [CrossRef]
- Antunes, M.; Abreu, S.C.; Cruz, F.F.; Teixeira, A.C.; Lopes-Pacheco, M.; Bandeira, E.; Olsen, P.; Diaz, B.L.; Takyia, C.M.; Freitas, I.P.R.G.; et al. Effects of different mesenchymal stromal cell sources and delivery routes in experimental emphysema. Respir. Res. 2014, 15, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abreu, S.C.; Antunes, M.A.; Xisto, D.G.; Cruz, F.; Branco, V.C.; Bandeira, E.; Kitoko, J.; De Araújo, A.F.; Dellatorre-Texeira, L.; Olsen, P.; et al. Bone Marrow, Adipose, and Lung Tissue-Derived Murine Mesenchymal Stromal Cells Release Different Mediators and Differentially Affect Airway and Lung Parenchyma in Experimental Asthma. Stem Cells Transl. Med. 2017, 6, 1557–1567. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Zhao, A. Mesenchymal stem cell therapy for acute respiratory distress syndrome: From basic to clinics. Protein Cell 2020, 11, 707–722. [Google Scholar] [CrossRef]
- Rogers, C.J.; Harman, R.J.; Bunnell, B.A.; Schreiber, M.A.; Xiang, C.; Wang, F.-S.; Santidrian, A.F.; Minev, B.R. Rationale for the clinical use of adipose-derived mesenchymal stem cells for COVID-19 patients. J. Transl. Med. 2020, 18, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Leng, Z.; Zhu, R.; Hou, W.; Feng, Y.; Yang, Y.; Han, Q.; Shan, G.; Meng, F.; Du, D.; Wang, S.; et al. Transplantation of ACE2- Mesenchymal Stem Cells Improves the Outcome of Patients with COVID-19 Pneumonia. Aging Dis. 2020, 11, 216–228. [Google Scholar] [CrossRef] [Green Version]
- Jackson, M.V.; Morrison, T.J.; Doherty, D.F.; McAuley, D.; Matthay, M.A.; Kissenpfennig, A.; O’Kane, C.; Krasnodembskaya, A.D. Mitochondrial Transfer via Tunneling Nanotubes is an Important Mechanism by Which Mesenchymal Stem Cells Enhance Macrophage Phagocytosis in the In Vitro and In Vivo Models of ARDS. Stem Cells 2016, 34, 2210–2223. [Google Scholar] [CrossRef] [Green Version]
- Alcayaga-Miranda, F.; Cuenca, J.; Martin, A.; Contreras, L.; Figueroa, F.E.; Khoury, M. Combination therapy of menstrual derived mesenchymal stem cells and antibiotics ameliorates survival in sepsis. Stem Cell Res. Ther. 2015, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Sung, D.K.; Chang, Y.S.; Sung, S.I.; Yoo, H.S.; Ahn, S.Y.; Park, W.S. Antibacterial effect of mesenchymal stem cells against Escherichia coli is mediated by secretion of beta- defensin- 2 via toll- like receptor 4 signalling. Cell. Microbiol. 2016, 18, 424–436. [Google Scholar] [CrossRef] [Green Version]
- Morofuji, Y. Drug Development for Central Nervous System Diseases Using In vitro Blood-brain Barrier Models and Drug Repositioning. Curr. Pharm. Des. 2020, 26, 1466–1485. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Qiu, X.; Jiao, C.; Lu, M.; Zhao, X.; Li, X.; Li, J.; Ma, J.; Zhang, H. Candida albicans SC5314 inhibits NLRP3/NLRP6 inflammasome expression and dampens human intestinal barrier activity in Caco-2 cell monolayer model. Cytokine 2020, 126, 154882. [Google Scholar] [CrossRef] [PubMed]
- Bengalli, R.; Gualtieri, M.; Capasso, L.; Urani, C.; Camatini, M. Impact of zinc oxide nanoparticles on an in vitro model of the human air-blood barrier. Toxicol. Lett. 2017, 279, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Kuehn, A.; Kletting, S.; de Souza Carvalho-Wodarz, C.; Repnik, U.; Griffiths, G.; Fischer, U.; Meese, E.; Huwer, H.; Wirth, D.; May, T.; et al. Human alveolar epithelial cells expressing tight junctions to model the air-blood barrier. ALTEX 2016, 33, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Matsukawa, Y.; Lee, V.H.L.; Crandall, E.D.; Kim, K.-J. Size-Dependent Dextran Transport across Rat Alveolar Epithelial Cell Monolayers. J. Pharm. Sci. 1997, 86, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Michaut, P.; Planes, C.; Escoubet, B.; Clement, A.; Amiel, C.; Clerici, C. Rat lung alveolar type II cell line maintains sodium transport characteristics of primary culture. J. Cell. Physiol. 1996, 169, 78–86. [Google Scholar] [CrossRef]
- Dobbs, L.G. Isolation and culture of alveolar type II cells. Am. J. Physiol. Cell. Mol. Physiol. 1990, 258, L134–L147. [Google Scholar] [CrossRef]
- Travaglini, K.J.; Nabhan, A.N.; Penland, L.; Sinha, R.; Gillich, A.; Sit, R.V.; Chang, S.; Conley, S.D.; Mori, Y.; Seita, J.; et al. A molecular cell atlas of the human lung from single-cell RNA sequencing. Nat. Cell Biol. 2020, 587, 619–625. [Google Scholar] [CrossRef]
- Raredon, M.S.B.; Adams, T.S.; Suhail, Y.; Schupp, J.C.; Poli, S.; Neumark, N.; Leiby, K.L.; Greaney, A.M.; Yuan, Y.; Horien, C.; et al. Single-cell connectomic analysis of adult mammalian lungs. Sci. Adv. 2019, 5, eaaw3851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlingmann, B.; Molina, S.A.; Koval, M. Claudins: Gatekeepers of lung epithelial function. Semin. Cell Dev. Biol. 2015, 42, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Tsukita, S.; Furuse, M. Claudin-based barrier in simple and stratified cellular sheets. Curr. Opin. Cell Biol. 2002, 14, 531–536. [Google Scholar] [CrossRef]
- Lal-Nag, M.; Morin, P.J. The claudins. Genome Biol. 2009, 10, 235. [Google Scholar] [CrossRef] [Green Version]
- Günzel, D.; Fromm, M. Claudins and Other Tight Junction Proteins. Compr. Physiol. 2012, 2, 1819–1852. [Google Scholar] [CrossRef] [PubMed]
- Soini, Y. Claudins in lung diseases. Respir. Res. 2011, 12, 70. [Google Scholar] [CrossRef] [Green Version]
- Lappi-Blanco, E.; Soini, Y. Claudin-7 expression and its association with tumor progression in systemic malignancies--reply. Hum. Pathol. 2013, 44, 1958–1959. [Google Scholar] [CrossRef] [PubMed]
- LaFemina, M.J.; Sutherland, K.M.; Bentley, T.; Gonzales, L.W.; Allen, L.; Chapin, C.J.; Rokkam, D.; Sweerus, K.A.; Dobbs, L.G.; Ballard, P.L.; et al. Claudin-18 Deficiency Results in Alveolar Barrier Dysfunction and Impaired Alveologenesis in Mice. Am. J. Respir. Cell Mol. Biol. 2014, 51, 550–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Flodby, P.; Luo, J.; Kage, H.; Sipos, A.; Gao, D.; Ji, Y.; Beard, L.L.; Marconett, C.N.; DeMaio, L.; et al. Knockout Mice Reveal Key Roles for Claudin 18 in Alveolar Barrier Properties and Fluid Homeostasis. Am. J. Respir. Cell Mol. Biol. 2014, 51, 210–222. [Google Scholar] [CrossRef] [Green Version]
- LaFemina, M.J.; Rokkam, D.; Chandrasena, A.; Pan, J.; Bajaj, A.; Johnson, M.; Frank, J.A. Keratinocyte growth factor enhances barrier function without altering claudin expression in primary alveolar epithelial cells. Am. J. Physiol. Cell. Mol. Physiol. 2010, 299, L724–L734. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, L.A.; Overgaard, C.E.; Ward, C.; Margulies, S.S.; Koval, M. Differential effects of claudin-3 and claudin-4 on alveolar epithelial barrier function. Am. J. Physiol. Cell. Mol. Physiol. 2011, 301, L40–L49. [Google Scholar] [CrossRef]
- Wray, C.; Mao, Y.; Pan, J.; Chandrasena, A.; Piasta, F.; Frank, J.A. Claudin-4 augments alveolar epithelial barrier function and is induced in acute lung injury. Am. J. Physiol. Cell. Mol. Physiol. 2009, 297, L219–L227. [Google Scholar] [CrossRef] [Green Version]
- Barczyk, M.; Carracedo, S.; Gullberg, D. Integrins. Cell Tissue Res. 2009, 339, 269–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campiglio, C.; Figliuzzi, M.; Silvani, S.; Tironi, M.; Conti, S.; Boschetti, F.; Remuzzi, A. Influence of Culture Substrates on Morphology and Function of Pulmonary Alveolar Cells In Vitro. Biomolecules 2021, 11, 675. [Google Scholar] [CrossRef] [PubMed]
- Campbell, L.; Hollins, A.; Al-Eid, A.; Newman, G.R.; von Ruhland, C.; Gumbleton, M. Caveolin-1 Expression and Caveolae Biogenesis during Cell Transdifferentiation in Lung Alveolar Epithelial Primary Cultures. Biochem. Biophys. Res. Commun. 1999, 262, 744–751. [Google Scholar] [CrossRef]
- Fang, X.; Neyrinck, A.P.; Matthay, M.A.; Lee, J.W. Allogeneic Human Mesenchymal Stem Cells Restore Epithelial Protein Permeability in Cultured Human Alveolar Type II Cells by Secretion of Angiopoietin-1*. J. Biol. Chem. 2010, 285, 26211–26222. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Description | Fold Change | Related Cell Junction |
|---|---|---|---|
| Cav2 | Caveolin2 | 2.29 | Focal adhesions |
| Cldn15 | Claudin15 | 1.75 | Tight junctions |
| Cldn4 | Claudin4 | 1.89 | Tight junctions |
| Cldn6 | Claudin6 | 4.13 | Tight junctions |
| Dsc2 | Desmocollin2 | 1.81 | Desmosomes |
| Itga5 | Integrin, alpha 5 | 2.15 | Focal adhesions |
| Itgal | Integrin, alpha L | 1.87 | Focal adhesions |
| Itgam | Integirn, alpha M | 3.5 | Focal adhesions |
| Itgax | Integirn, alpha X | 2.78 | Focal adhesions |
| Itgb2 | Integrin, beta 2 | 1.82 | Focal adhesions |
| Itgb3 | Integrin, beta 3 | 1.89 | Focal adhesions |
| Itgb6 | Integrin, beta 6 | 2.15 | Focal adhesions |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishii, M.; Tsuchiya, T.; Doi, R.; Morofuji, Y.; Fujimoto, T.; Muto, H.; Suematsu, T.; Mori, R.; Matsumoto, K.; Miyazaki, T.; et al. Increased In Vitro Intercellular Barrier Function of Lung Epithelial Cells Using Adipose-Derived Mesenchymal Stem/Stromal Cells. Pharmaceutics 2021, 13, 1264. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13081264
Ishii M, Tsuchiya T, Doi R, Morofuji Y, Fujimoto T, Muto H, Suematsu T, Mori R, Matsumoto K, Miyazaki T, et al. Increased In Vitro Intercellular Barrier Function of Lung Epithelial Cells Using Adipose-Derived Mesenchymal Stem/Stromal Cells. Pharmaceutics. 2021; 13(8):1264. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13081264
Chicago/Turabian StyleIshii, Mitsutoshi, Tomoshi Tsuchiya, Ryoichiro Doi, Yoichi Morofuji, Takashi Fujimoto, Hideki Muto, Takashi Suematsu, Ryoichi Mori, Keitaro Matsumoto, Takuro Miyazaki, and et al. 2021. "Increased In Vitro Intercellular Barrier Function of Lung Epithelial Cells Using Adipose-Derived Mesenchymal Stem/Stromal Cells" Pharmaceutics 13, no. 8: 1264. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13081264