
Thermodynamic Stability Is a Strong Predictor for the Delivery of DARPins to the Cytosol via Anthrax Toxin

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Cloning
2.3. Protein Expression
2.4. Protein Purification
2.5. Biotin Ligase Uptake Assay
2.6. CD Spectroscopy and Equilibrium Unfolding
2.7. Surface Plasmon Resonance (SPR)
3. Results
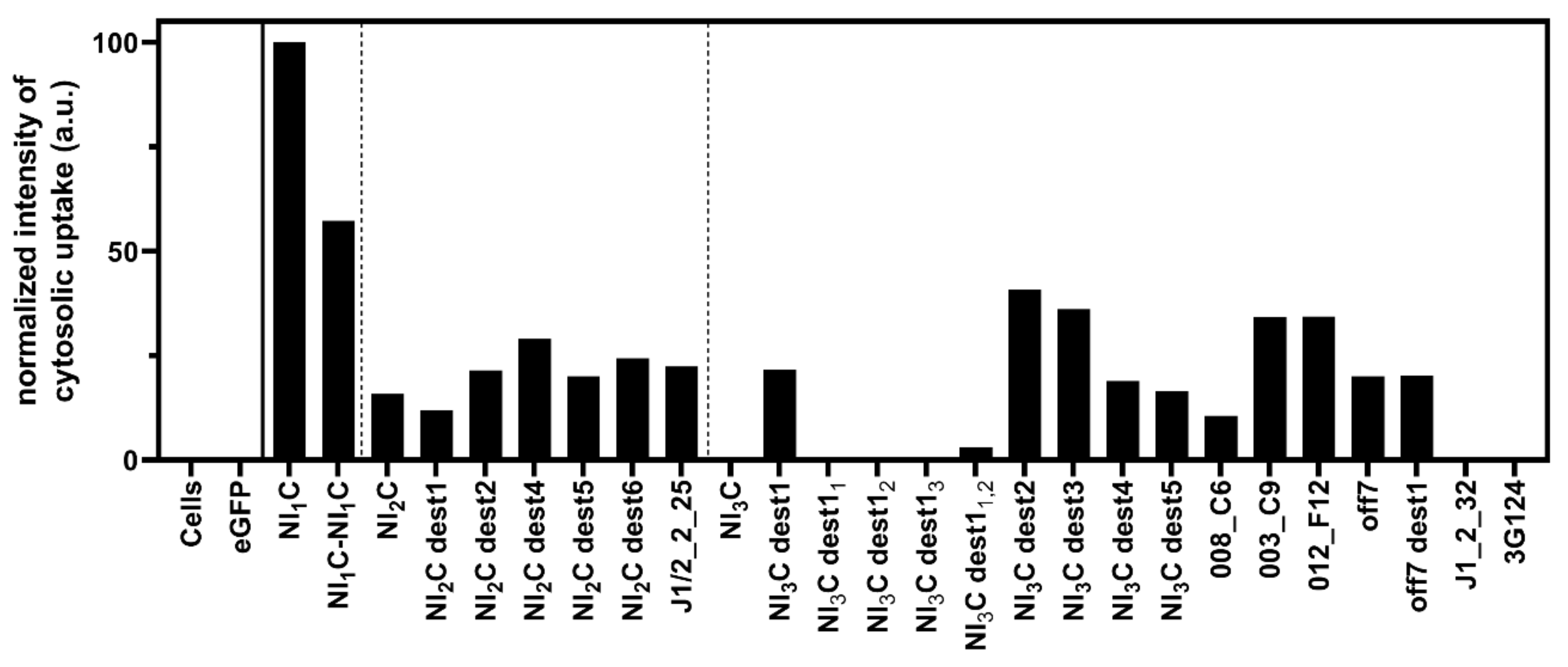
3.1. Cytosolic Translocation of LFN-DARPin Cargoes via PAwt-sANTXR-Ac2
3.2. Denaturant-induced Equilibrium Unfolding of DARPin Cargoes

{kind=link}
{kind=link}
| Protein | Framework Mutation a | IR b | MW | Dm | ΔG0 | m | CL c | Ref. |
|---|---|---|---|---|---|---|---|---|
| [kDa] | [M] | [kcal mol−1] | [kcal mol−1 M−1] | |||||
| LFN | - | - | 32.4 | 0.9 | 3.6 | 4.1 | Y | [27] |
| NI1C | - | 1 | 11.1 | 1.3 | 2.5 | 2.0 | Y | [28] |
| NI1C-NI1C | - | 1-1 | 21.0 | 1.4 | 2.5 | 1.8 | Y | - |
| NI2C | - | 2 | 14.6 | 3.3 | 7.5 | 2.3 | Y | [28] |
| NI2C dest2 | (L8A)1,2,3 | 2 | 14.5 | 1.7 | 4.2 | 2.5 | Y | [6] |
| J1/2_2_25 | - | 2 | 14.6 | 2.7 | 9.1 | 3.4 | Y | [25] |
| NI3C dest1 | (L24A)1,2,3 | 3 | 18.0 | 2.7 | 7.6 | 2.8 | Y | [6] |
| NI3C dest11 | (L24A)1 | 3 | 18.1 | 3.7 * | - | - | N | - |
| NI3C dest12 | (L24A)2 | 3 | 18.1 | 3.7 * | - | - | N | - |
| NI3C dest13 | (L24A)3 | 3 | 18.1 | 4.2 * | - | - | N | - |
| NI3C dest11,2 | (L24A)1,2 | 3 | 18.0 | 3.0 * | - | - | Y/N | - |
| NI3C dest12,3 | (L24A)2,3 | 3 | 18.0 | 3.6 * | - | - | N | - |
| NI3C dest2 | (L24G)1,2,3 | 3 | 17.9 | 1.6 | 6.7 | 4.2 | Y | [6] |
| NI3C dest3 | (L8A, L24A)1,2,3 | 3 | 17.8 | 1.6 | 6.4 | 4.1 | Y | [6] |
| NI3C dest4 | (L8G, L24A)1,2,3 | 3 | 17.8 | 0.7 | 2.6 | 3.5 | Y | [6] |
| 008_C6 | - | 3 | 18.2 | 2.1 * | - | - | Y | [29] |
| 003_C9 | - | 3 | 18.3 | 1.5 | 3.3 | 2.2 | Y | [29] |
| 012_F12 | - | 3 | 18.1 | 1.2 | 2.8 | 2.3 | Y | [29] |
| off7 | - | 3 | 18.1 | 3.6 * | - | - | Y | [21] |
| off7 dest1 | (L24A)1,2,3 | 3 | 18.0 | 1.8 | 6.7 | 3.7 | Y | - |
| NI3C | - | 3 | 18.1 | 4.8 * | - | - | N | [28] |
| J1_2_32 | - | 3 | 18.0 | 3.4 | 10.1 | 3.0 | N | [25] |
| 3G124 | - | 3 | 17.9 | 5.2* | - | - | N | [24] |
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| PA | protective antigen |
| DARPin(s) | designed ankyrin repeat protein(s) |
| CD | circular dichroism |
| LF | lethal factor |
| EF | edema factor |
| AEBSF | 4-(2-aminoethyl) benzenesulfonyl fluoride |
| IMAC | immobilized metal ion affinity chromatography |
| CV | column volume |
| MRE | mean residue ellipticity |
| Dm | denaturation midpoint |
| SPR | surface plasmon resonance |
| EpCAM | epithelial cell adhesion molecule |
| LFN | lethal factor 1–254 |
| BirA | biotin ligase derived from E. coli |
| dest | destabilized |
| JNKs | c-Jun N-terminal kinases |
| GdnHCl | guanidine hydrochloride |
| IR | internal repeats |
| CL | cytosolic localization |
References
- Kintzer, A.F.; Thoren, K.L.; Sterling, H.J.; Dong, K.C.; Feld, G.K.; Tang, I.I.; Zhang, T.T.; Williams, E.R.; Berger, J.M.; Krantz, B.A. The protective antigen component of anthrax toxin forms functional octameric complexes. J. Mol. Biol. 2009, 392, 614–629. [Google Scholar] [CrossRef]
- Friebe, S.; van der Goot, F.; Bürgi, J. The ins and outs of anthrax toxin. Toxins 2016, 8, 69. [Google Scholar] [CrossRef] [Green Version]
- Feld, G.K.; Brown, M.J.; Krantz, B.A. Ratcheting up protein translocation with anthrax toxin. Protein Sci. 2012, 21, 606–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zielinski, R.; Lyakhov, I.; Jacobs, A.; Chertov, O.; Kramer-Marek, G.; Francella, N.; Stephen, A.; Fisher, R.; Blumenthal, R.; Capala, J. Affitoxin—A novel recombinant, HER2-specific, anticancer agent for targeted therapy of HER2-positive tumors. J. Immunother. 2009, 32, 817–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mechaly, A.; McCluskey, A.J.; Collier, R.J. Changing the receptor specificity of anthrax toxin. mBio 2012, 3, e00088-12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verdurmen, W.P.R.; Luginbühl, M.; Honegger, A.; Plückthun, A. Efficient cell-specific uptake of binding proteins into the cytoplasm through engineered modular transport systems. J. Control. Release 2015, 200, 13–22. [Google Scholar] [CrossRef] [PubMed]
- McCluskey, A.J.; Olive, A.J.; Starnbach, M.N.; Collier, R.J. Targeting HER2-positive cancer cells with receptor-redirected anthrax protective antigen. Mol. Oncol. 2013, 7, 440–451. [Google Scholar] [CrossRef]
- Varughese, M.; Chi, A.; Teixeira, A.V.; Nicholls, P.J.; Keith, J.M.; Leppla, S.H. Internalization of a Bacillus anthracis protective antigen-c-myc fusion protein mediated by cell surface anti-c-myc antibodies. Mol. Med. 1998, 4, 87–95. [Google Scholar] [CrossRef]
- Becker, L.; Verdurmen, W.P.R.; Plückthun, A. Reengineering anthrax toxin protective antigen for improved receptor-specific protein delivery. BMC Biol. 2020, 18, 100. [Google Scholar] [CrossRef] [PubMed]
- Rabideau, A.E.; Pentelute, B.L. Delivery of non-native cargo into mammalian cells using anthrax lethal toxin. ACS Chem. Biol. 2016, 11, 1490–1501. [Google Scholar] [CrossRef]
- Wesche, J.; Elliott, J.L.; Falnes, P.Ø.; Olsnes, S.; Collier, R.J. Characterization of membrane translocation by anthrax protective antigen. Biochemistry 1998, 37, 15737–15746. [Google Scholar] [CrossRef]
- Roderer, D.; Schubert, E.; Sitsel, O.; Raunser, S. Towards the application of Tc toxins as a universal protein translocation system. Nat. Commun. 2019, 10, 5263. [Google Scholar] [CrossRef] [Green Version]
- Thoren, K.L.; Worden, E.J.; Yassif, J.M.; Krantz, B.A. Lethal factor unfolding is the most force-dependent step of anthrax toxin translocation. Proc. Natl. Acad. Sci. USA 2009, 106, 21555–21560. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Finkelstein, A.; Collier, R.J. Evidence that translocation of anthrax toxin’s lethal factor is initiated by entry of its N terminus into the protective antigen channel. Proc. Natl. Acad. Sci. USA 2004, 101, 16756–16761. [Google Scholar] [CrossRef] [Green Version]
- Pentelute, B.L.; Sharma, O.; Collier, R.J. Chemical dissection of protein translocation through the anthrax toxin pore. Angew. Chem. Int. Ed. Engl. 2011, 50, 2294–2296. [Google Scholar] [CrossRef]
- Rabideau, A.E.; Liao, X.; Akçay, G.; Pentelute, B.L. Translocation of non-canonical polypeptides into cells using protective antigen. Sci. Rep. 2015, 5, 11944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.; Pentelute, B.L.; Collier, R.J.; Zhou, Z.H. Atomic structure of anthrax protective antigen pore elucidates toxin translocation. Nature 2015, 521, 545–549. [Google Scholar] [CrossRef] [PubMed]
- Feld, G.K.; Thoren, K.L.; Kintzer, A.F.; Sterling, H.J.; Tang, I.I.; Greenberg, S.G.; Williams, E.R.; Krantz, B.A. Structural basis for the unfolding of anthrax lethal factor by protective antigen oligomers. Nat. Struct. Mol. Biol. 2010, 17, 1383–1390. [Google Scholar] [CrossRef]
- Beilhartz, G.L.; Sugiman-Marangos, S.N.; Melnyk, R.A. Repurposing bacterial toxins for intracellular delivery of therapeutic proteins. Biochem. Pharmacol. 2017, 142, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Verdurmen, W.P.R.; Mazlami, M.; Plückthun, A. A Biotin ligase-based assay for the quantification of the cytosolic delivery of therapeutic proteins. In Synthetic Antibodies; Tiller, T., Ed.; Springer: New York, NY, USA, 2017; Volume 1575, pp. 223–236. ISBN 978-1-4939-6855-8. [Google Scholar]
- Binz, H.K.; Amstutz, P.; Kohl, A.; Stumpp, M.T.; Briand, C.; Forrer, P.; Grütter, M.G.; Plückthun, A. High-affinity binders selected from designed ankyrin repeat protein libraries. Nat. Biotechnol. 2004, 22, 575–582. [Google Scholar] [CrossRef]
- Kohl, A.; Binz, H.K.; Forrer, P.; Stumpp, M.T.; Plückthun, A.; Grütter, M.G. Designed to be stable: Crystal structure of a consensus ankyrin repeat protein. Proc. Natl. Acad. Sci. USA 2003, 100, 1700–1705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steiner, D.; Forrer, P.; Plückthun, A. Efficient selection of DARPins with sub-nanomolar affinities using SRP phage display. J. Mol. Biol. 2008, 382, 1211–1227. [Google Scholar] [CrossRef] [Green Version]
- Brauchle, M.; Hansen, S.; Caussinus, E.; Lenard, A.; Ochoa-Espinosa, A.; Scholz, O.; Sprecher, S.G.; Plückthun, A.; Affolter, M. Protein interference applications in cellular and developmental biology using DARPins that recognize GFP and mCherry. Biol. Open 2014, 3, 1252–1261. [Google Scholar] [CrossRef] [Green Version]
- Parizek, P.; Kummer, L.; Rube, P.; Prinz, A.; Herberg, F.W.; Plückthun, A. Designed Ankyrin Repeat Proteins (DARPins) as novel isoform-specific intracellular inhibitors of c-Jun N-terminal kinases. ACS Chem. Biol. 2012, 7, 1356–1366. [Google Scholar] [CrossRef] [PubMed]
- Interlandi, G.; Wetzel, S.K.; Settanni, G.; Plückthun, A.; Caflisch, A. Characterization and further stabilization of designed ankyrin repeat proteins by combining molecular dynamics simulations and experiments. J. Mol. Biol. 2008, 375, 837–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arora, N.; Leppla, S.H. Residues 1-254 of anthrax toxin lethal factor are sufficient to cause cellular uptake of fused polypeptides. J. Biol. Chem. 1993, 268, 3334–3341. [Google Scholar] [CrossRef]
- Wetzel, S.K.; Settanni, G.; Kenig, M.; Binz, H.K.; Plückthun, A. Folding and unfolding mechanism of highly stable full-consensus ankyrin repeat proteins. J. Mol. Biol. 2008, 376, 241–257. [Google Scholar] [CrossRef] [Green Version]
- Schilling, J.; Schöppe, J.; Plückthun, A. From DARPins to LoopDARPins: Novel LoopDARPin design allows the selection of low picomolar binders in a single round of ribosome display. J. Mol. Biol. 2014, 426, 691–721. [Google Scholar] [CrossRef] [PubMed]
- Binz, H.K.; Kohl, A.; Plückthun, A.; Grütter, M.G. Crystal structure of a consensus-designed ankyrin repeat protein: Implications for stability. Proteins 2006, 65, 280–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, X.; Rabideau, A.E.; Pentelute, B.L. Delivery of antibody mimics into mammalian cells via anthrax toxin protective antigen. ChemBioChem 2014, 15, 2458–2466. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Craggs, T.D.; Christodoulou, J.; Jackson, S.E. Stable intermediate states and high energy barriers in the unfolding of GFP. J. Mol. Biol. 2007, 370, 356–371. [Google Scholar] [CrossRef] [PubMed]
- Krantz, B.A.; Trivedi, A.D.; Cunningham, K.; Christensen, K.A.; Collier, R.J. Acid-induced unfolding of the amino-terminal domains of the lethal and edema factors of anthrax toxin. J. Mol. Biol. 2004, 344, 739–756. [Google Scholar] [CrossRef] [PubMed]
- Lo, S.Y.; Säbel, C.E.; Mapletoft, J.P.J.; Siemann, S. Influence of chemical denaturants on the activity, fold and zinc status of anthrax lethal factor. Biochem. Biophys. Rep. 2015, 1, 68–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, P.; Singh, S.; Tiwari, A.; Bhat, R.; Bhatnagar, R. Effect of pH on stability of anthrax lethal factor: Correlation between denaturation and activity. Biochem. Biophys. Res. Commun. 2001, 284, 568–573. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Becker, L.; Singh Badwal, J.; Brandl, F.; Verdurmen, W.P.R.; Plückthun, A. Thermodynamic Stability Is a Strong Predictor for the Delivery of DARPins to the Cytosol via Anthrax Toxin. Pharmaceutics 2021, 13, 1285. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13081285
Becker L, Singh Badwal J, Brandl F, Verdurmen WPR, Plückthun A. Thermodynamic Stability Is a Strong Predictor for the Delivery of DARPins to the Cytosol via Anthrax Toxin. Pharmaceutics. 2021; 13(8):1285. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13081285
Chicago/Turabian StyleBecker, Lukas, Jasleen Singh Badwal, Fabian Brandl, Wouter P. R. Verdurmen, and Andreas Plückthun. 2021. "Thermodynamic Stability Is a Strong Predictor for the Delivery of DARPins to the Cytosol via Anthrax Toxin" Pharmaceutics 13, no. 8: 1285. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics13081285