
In Vivo Evaluation of an Antibody-Functionalized Lipoidal Nanosystem for Schistosomiasis Intervention



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Nanoliposome Preparation
2.2.2. Surface-Engineering of the Nanoliposomes with Anti-Calpain
2.2.3. Morphological Analysis
Scanning Electron Microscopy
Transmission Electron Microscopy
2.2.4. Particle Size Distribution, Zeta Potential, and Polydispersity Index (PDI) Analysis
2.2.5. Evaluation of the Drug Entrapment Efficiency of the PZQ-Loaded Nanoliposomes
2.2.6. Fourier-Transform Infrared Spectroscopy (FT-IR)
2.2.7. X-ray Powder Diffraction (XRPD) Analysis
2.2.8. Differential Scanning Calorimetry (DSC) Analysis
2.2.9. Thermogravimetric Analyzer (TGA) Analysis
2.2.10. In Vitro Release of the Drugs
2.2.11. In Vitro Cytotoxicity Assay (MTT Assay)
2.2.12. Cell Morphology Analysis
2.2.13. In Vivo Toxicity
2.2.14. Sample Preparation for Histopathological Analysis
2.2.15. In Vivo Antischistosomal Study
Infection of Animals
Experimental Design and Animals Grouping
- Group 1: Infected control;
- Group 2: Single dose of 250 mg/kg of Anti-calpain-PZQ-loaded nanoliposomes was given two weeks post infection;
- Group 3: A drug (PZQ) 250 mg/kg single dose was given two weeks post infection;
- Group 4: Single dose of 250 mg/kg of Anti-calpain-PZQ-loaded nanoliposomes was given four weeks post infection;
- Group 5: A drug (PZQ) 250 mg/kg single dose was given four weeks post infection.
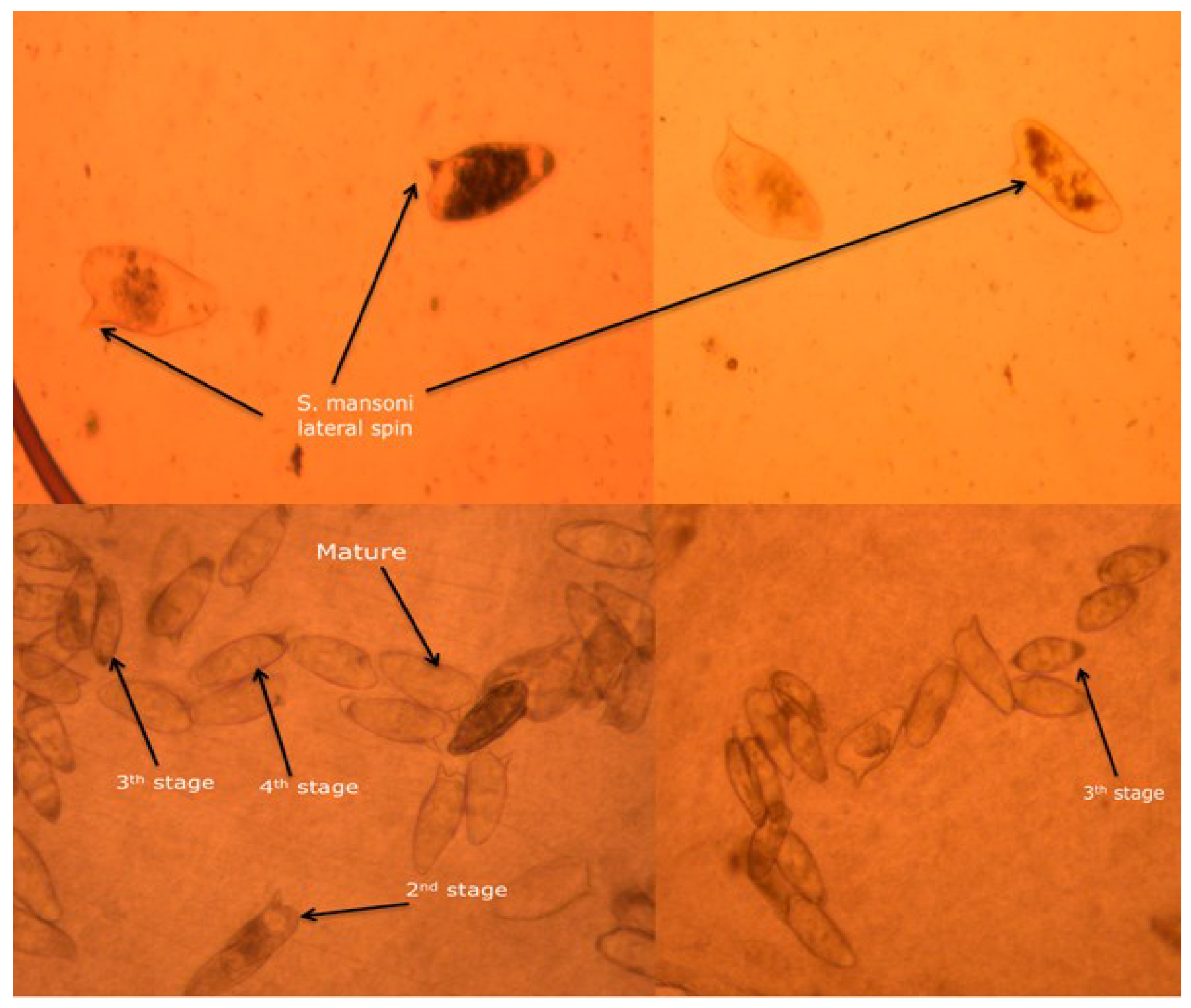
Assessment of Parasitological Criteria of Cure
2.2.16. Statistical Analysis
3. Results
3.1. Physicochemical Characterization of the Nanoliposomes
3.1.1. Surface Morphology and Shape Analysis of the Formulated Nanoliposomes
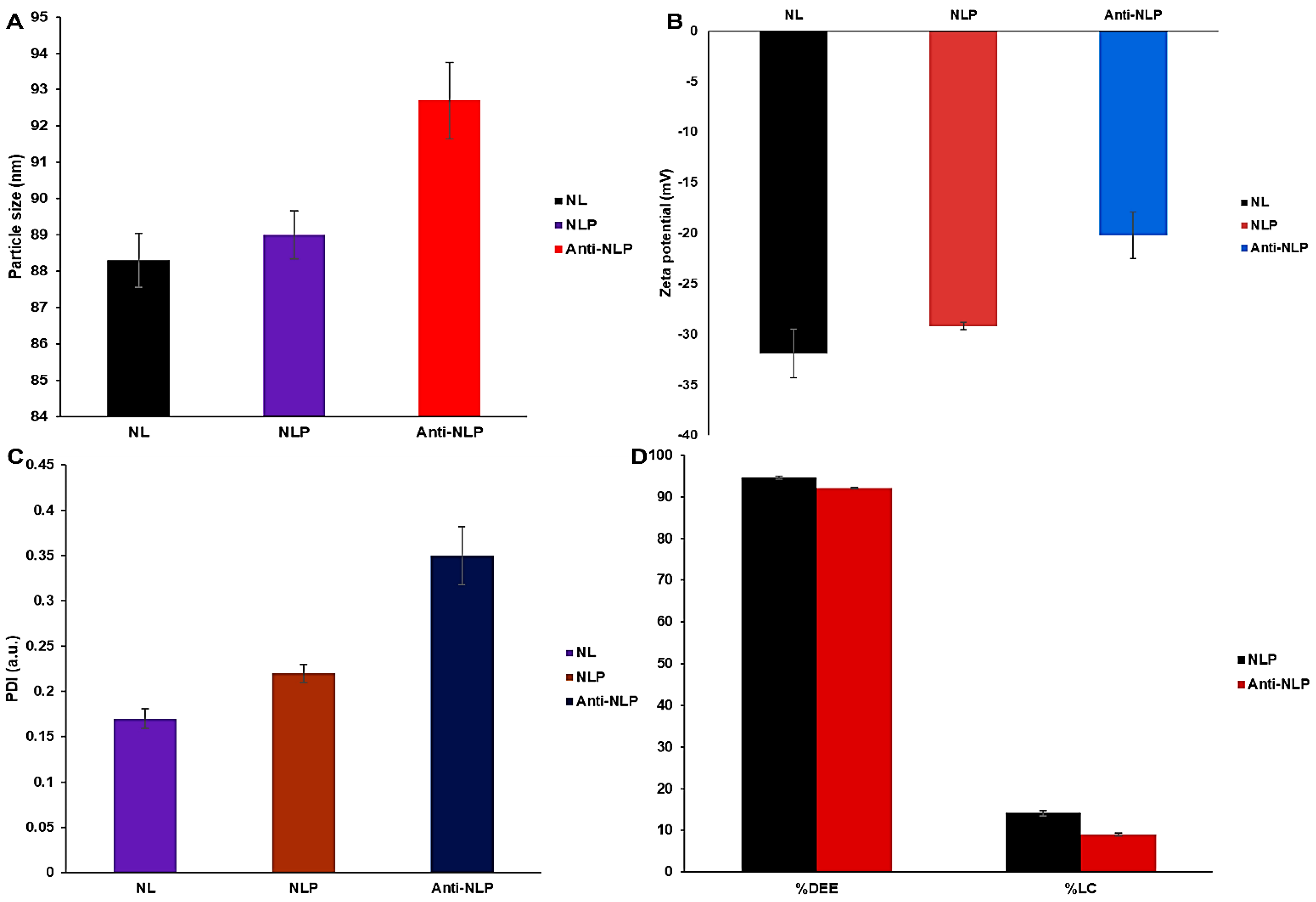
3.1.2. Particle Size Distribution, PDI, Zeta Potential, Drug Entrapment Efficacy, and Drug Loading Capacity Analysis
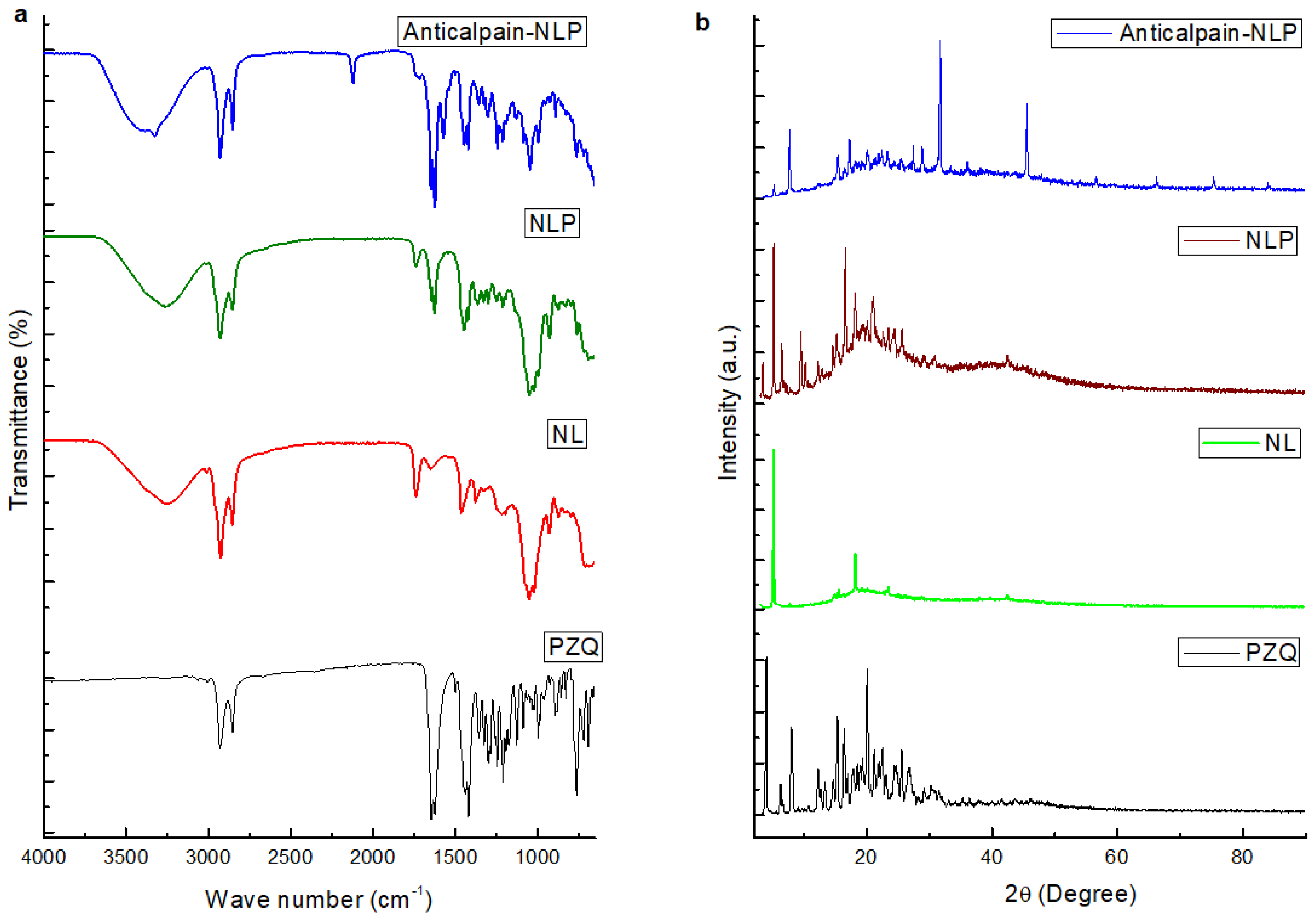
3.1.3. Molecular Vibrational Transitions and X-ray Powder Diffraction of the Formulated Nanoliposomes Evaluation
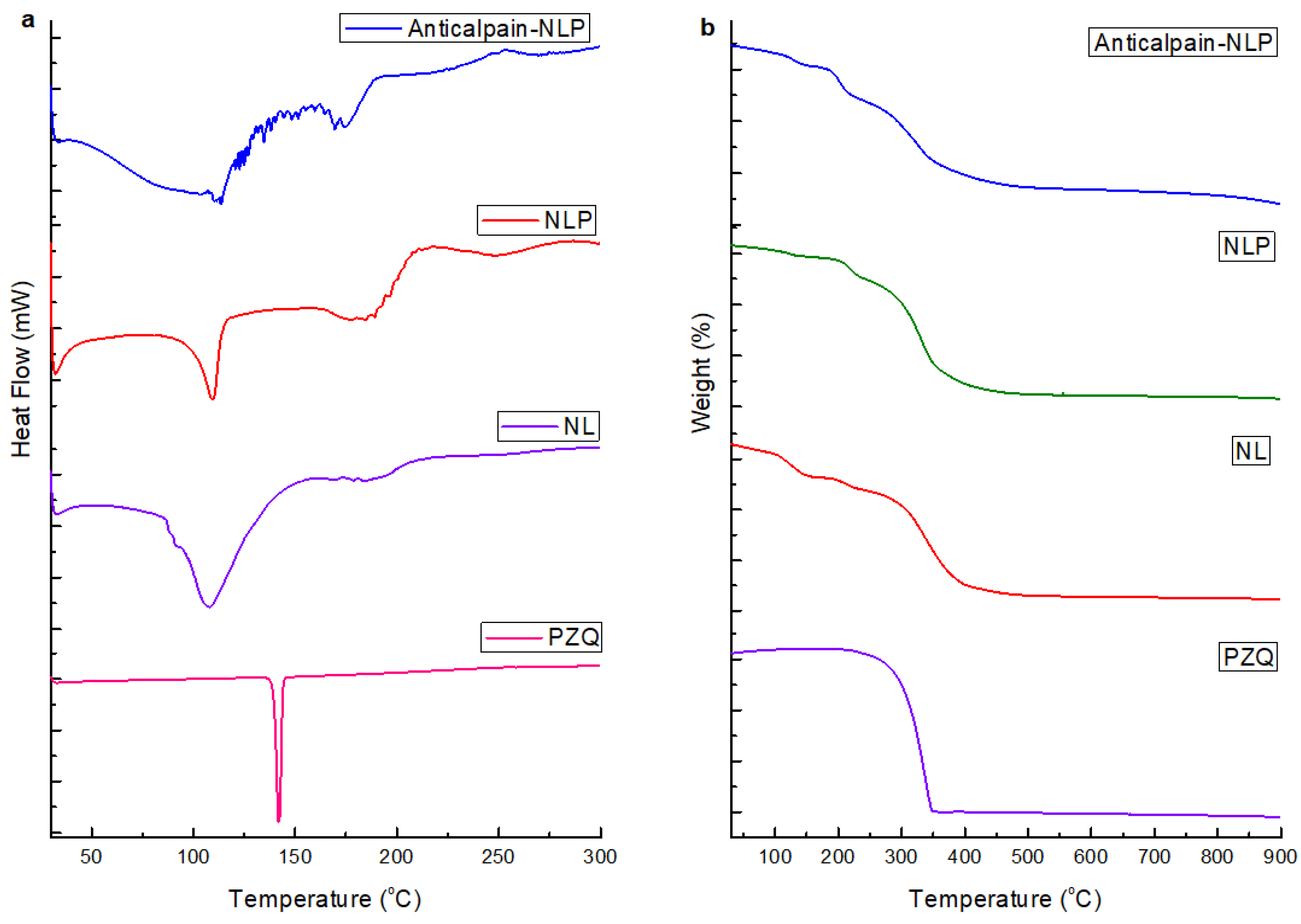
3.1.4. Thermal Behavior of the Formulated Nanoliposomes
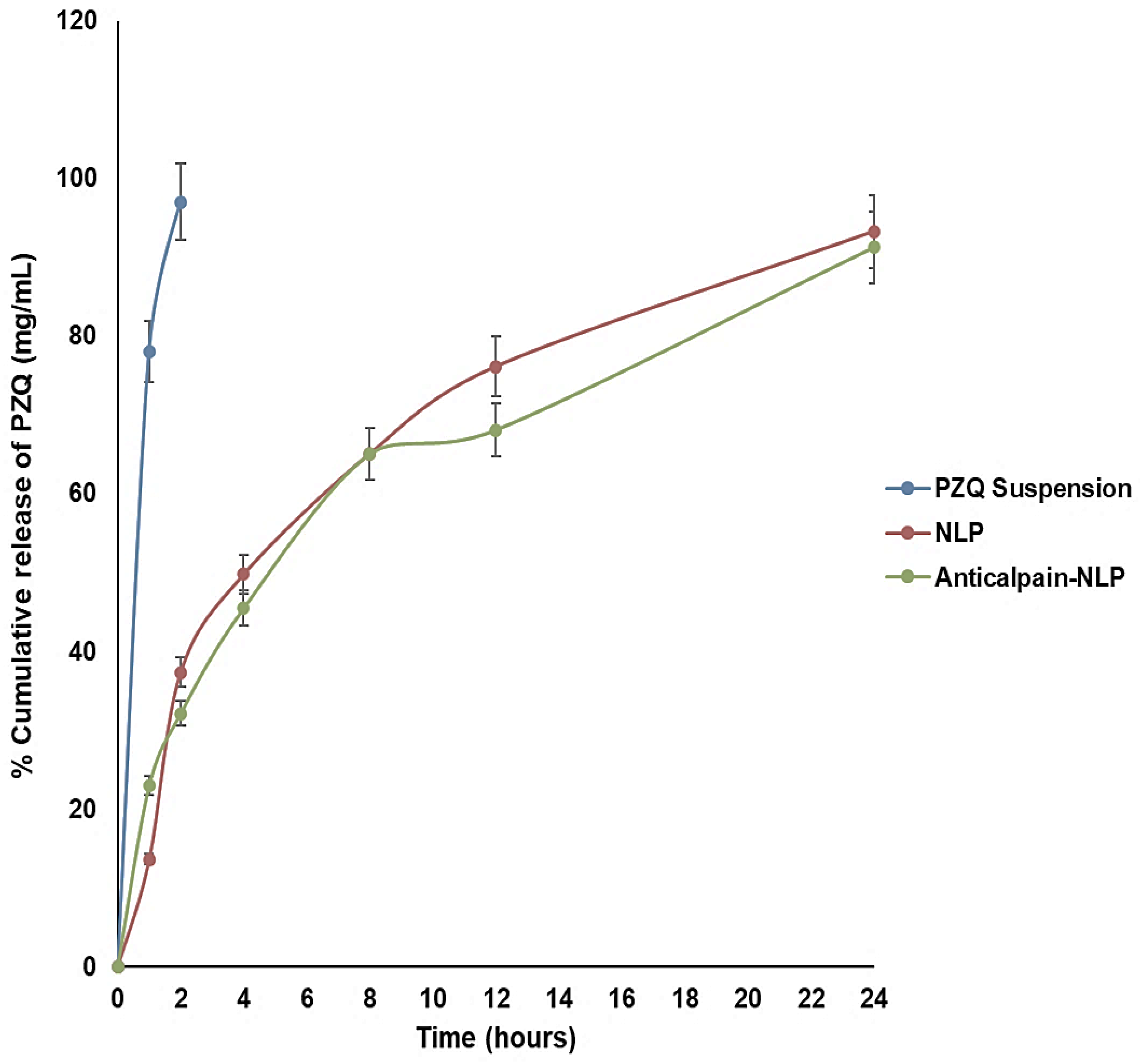
3.2. In Vitro Release Behavior of PZQ Analysis
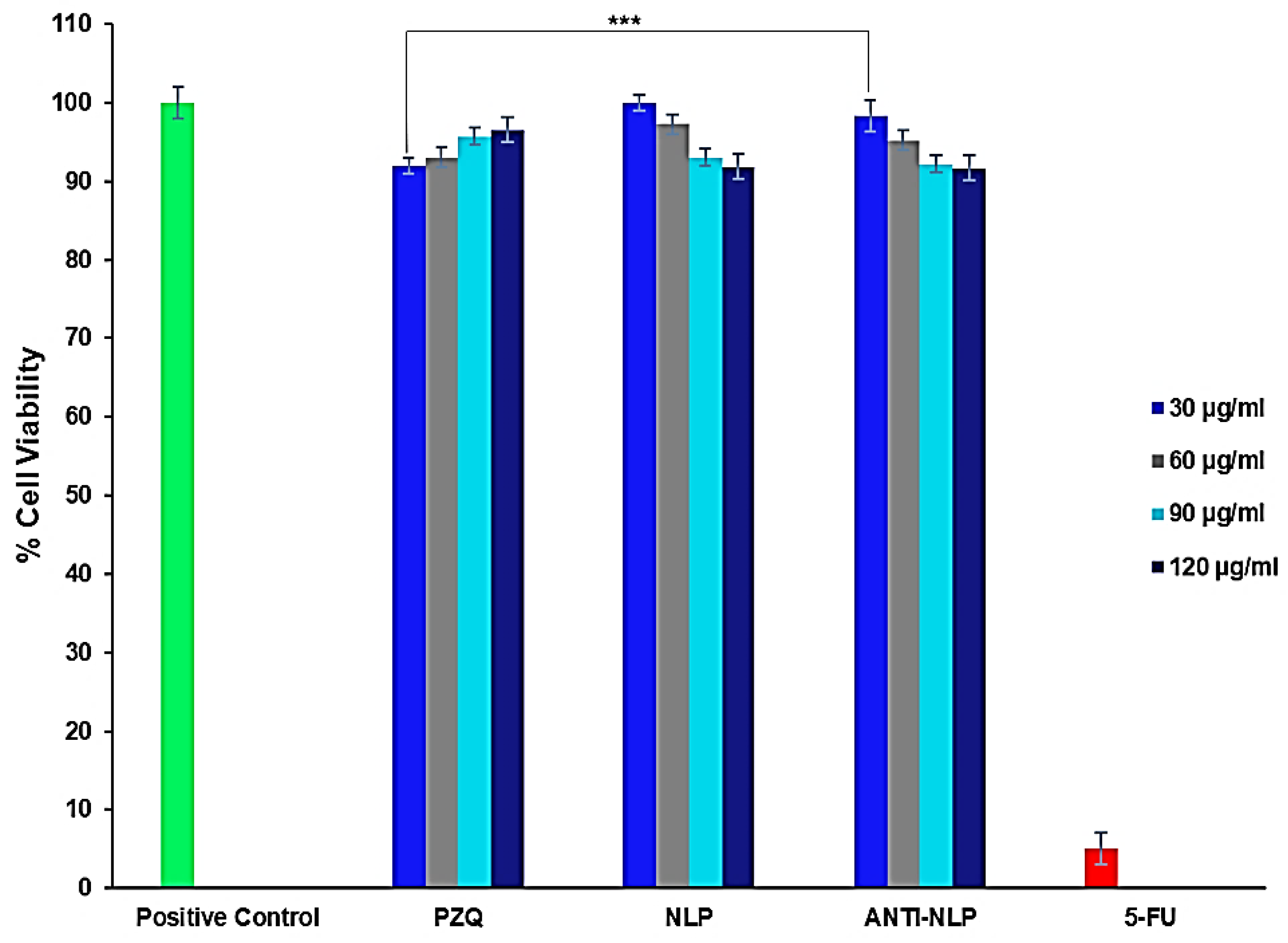
3.3. In Vitro Toxicity Analysis
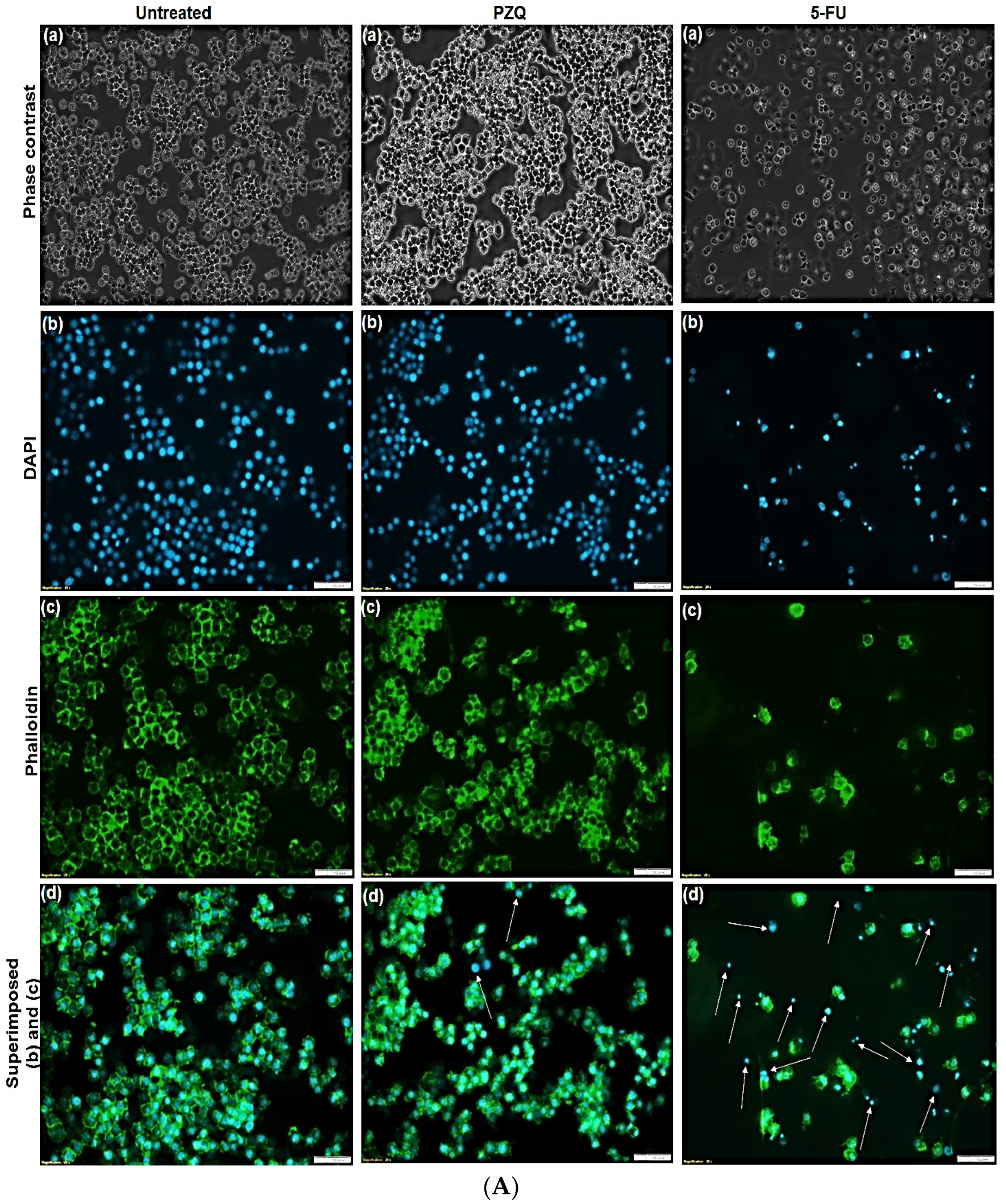
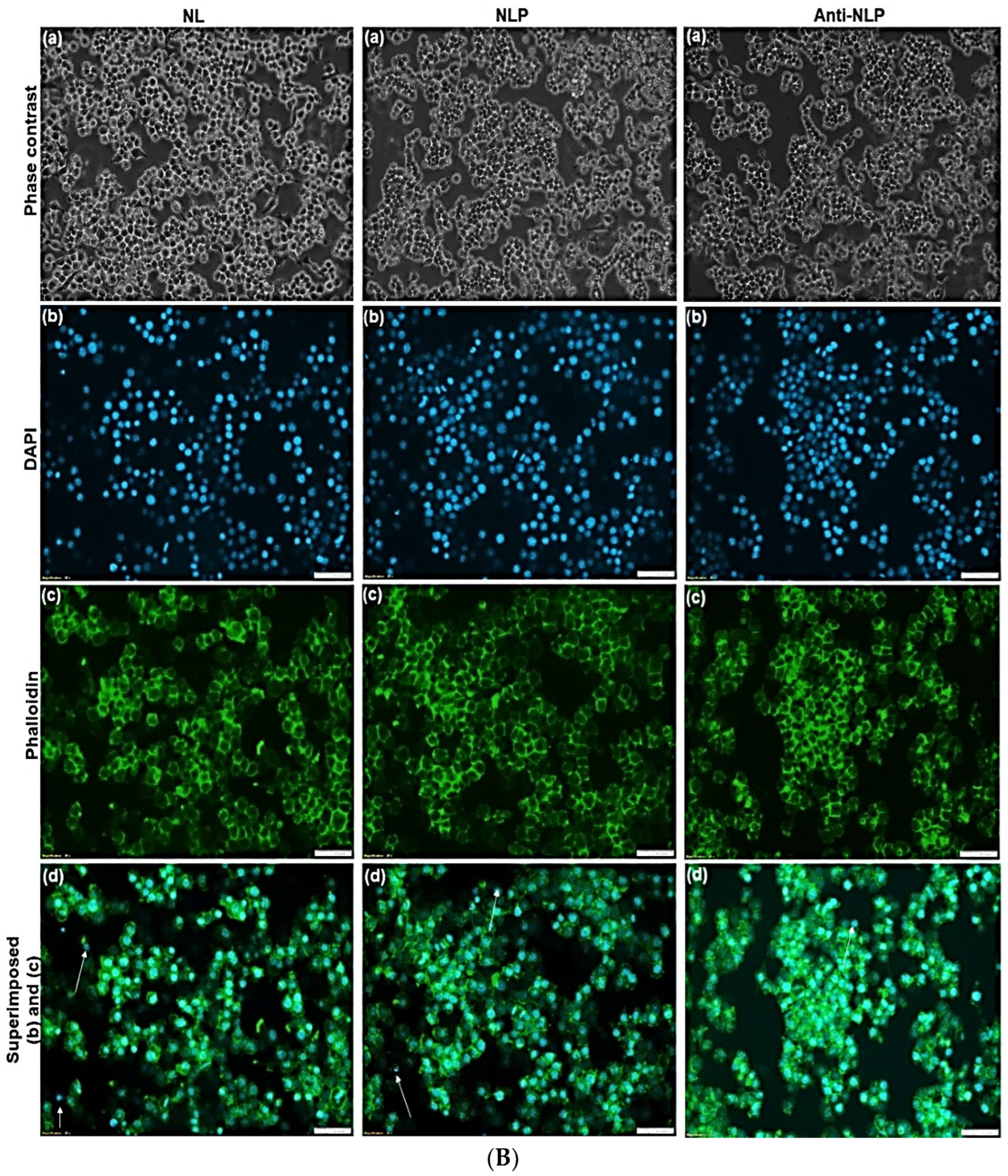
3.4. Morphological Studies on RAW 264.7
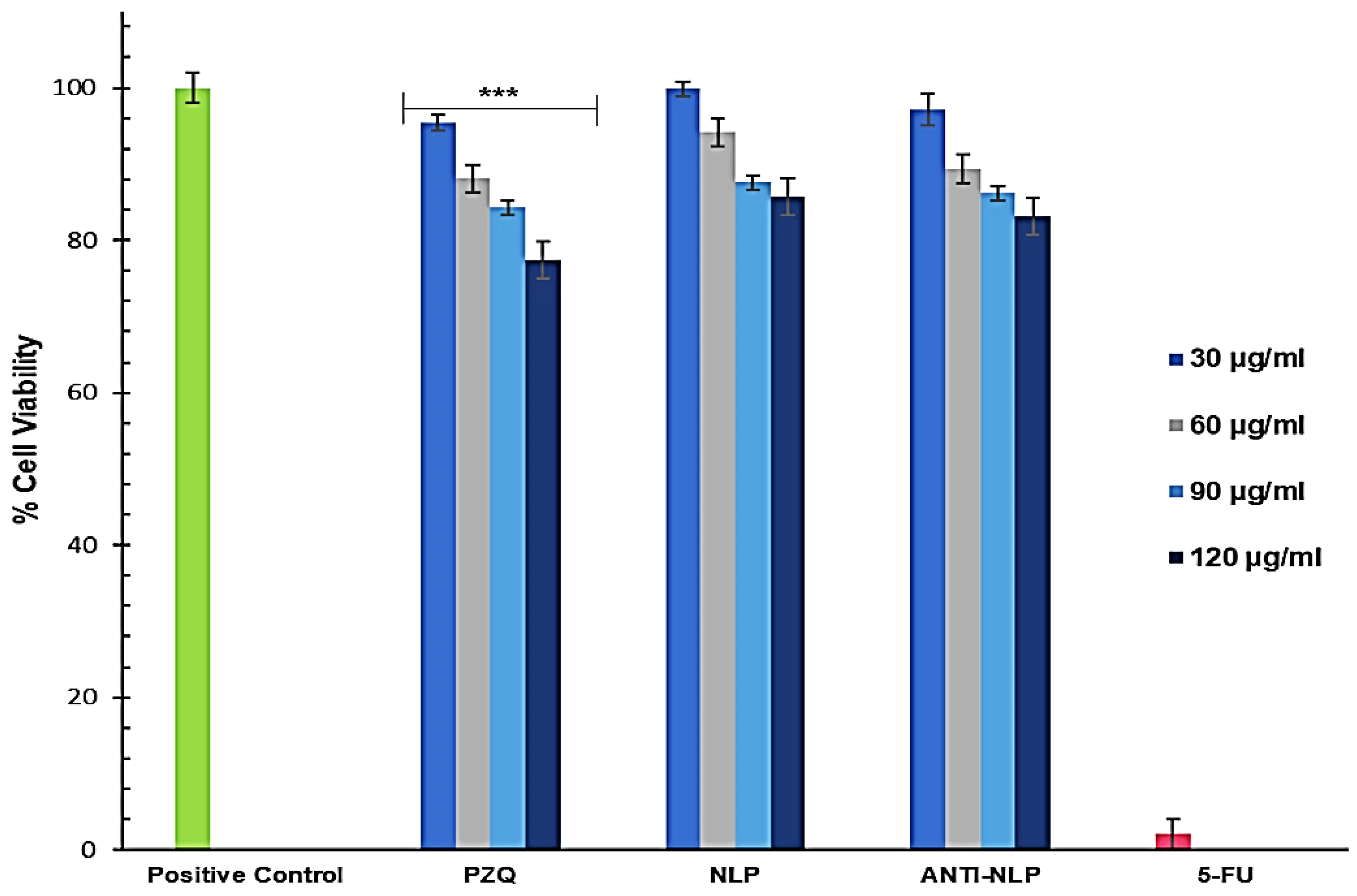
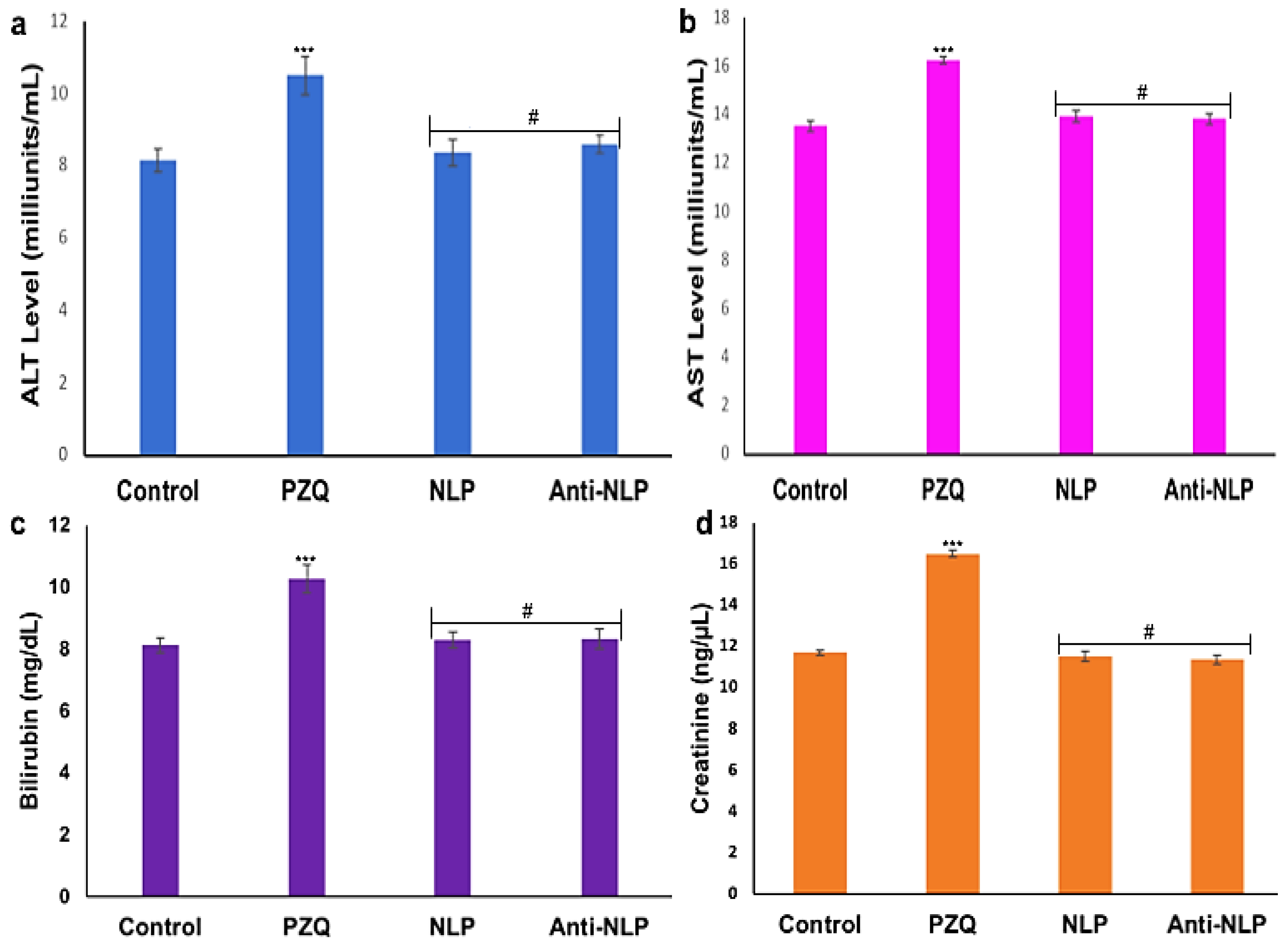
3.5. In Vivo Toxicity Analysis
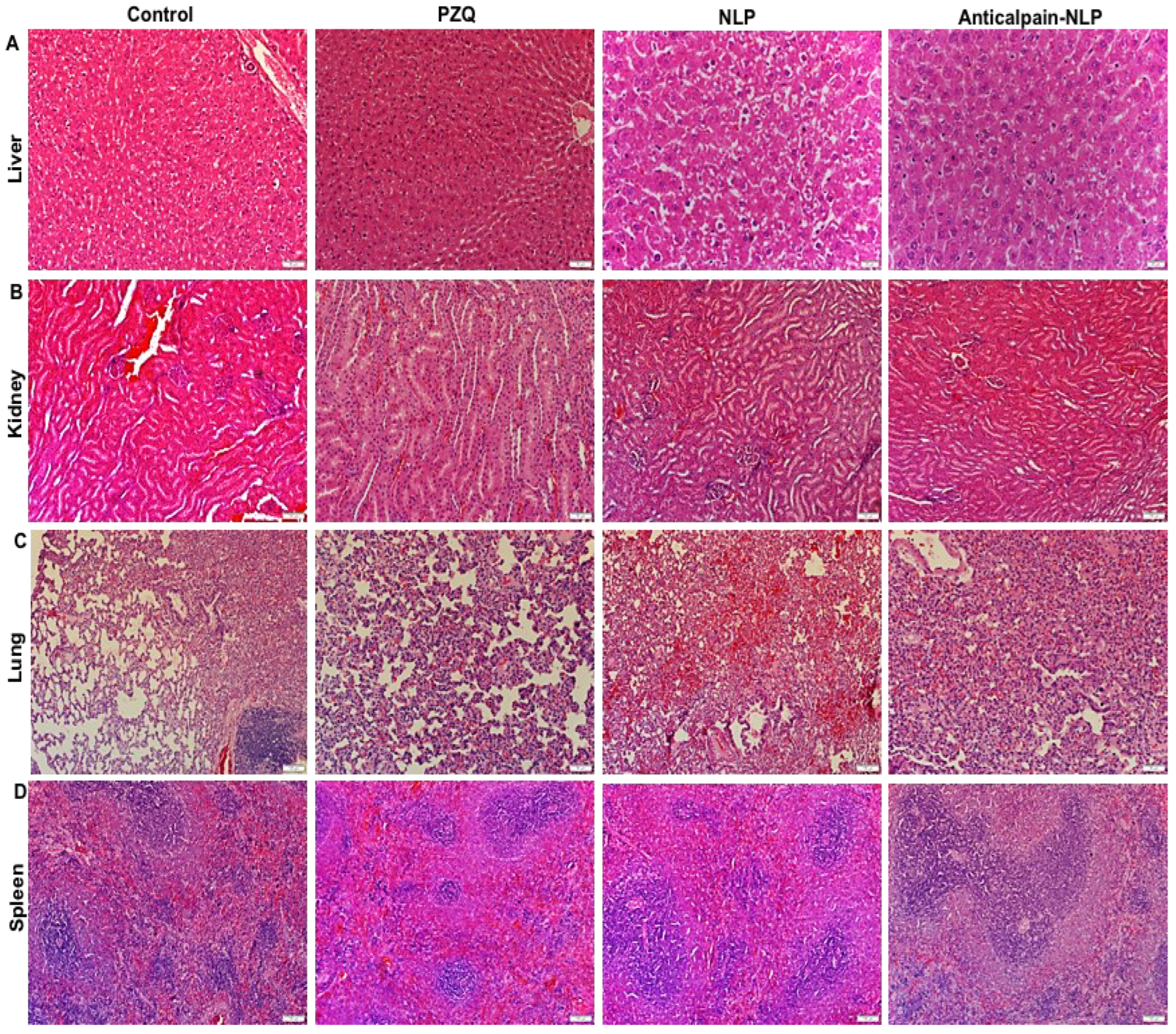
3.6. Histopathological Analysis
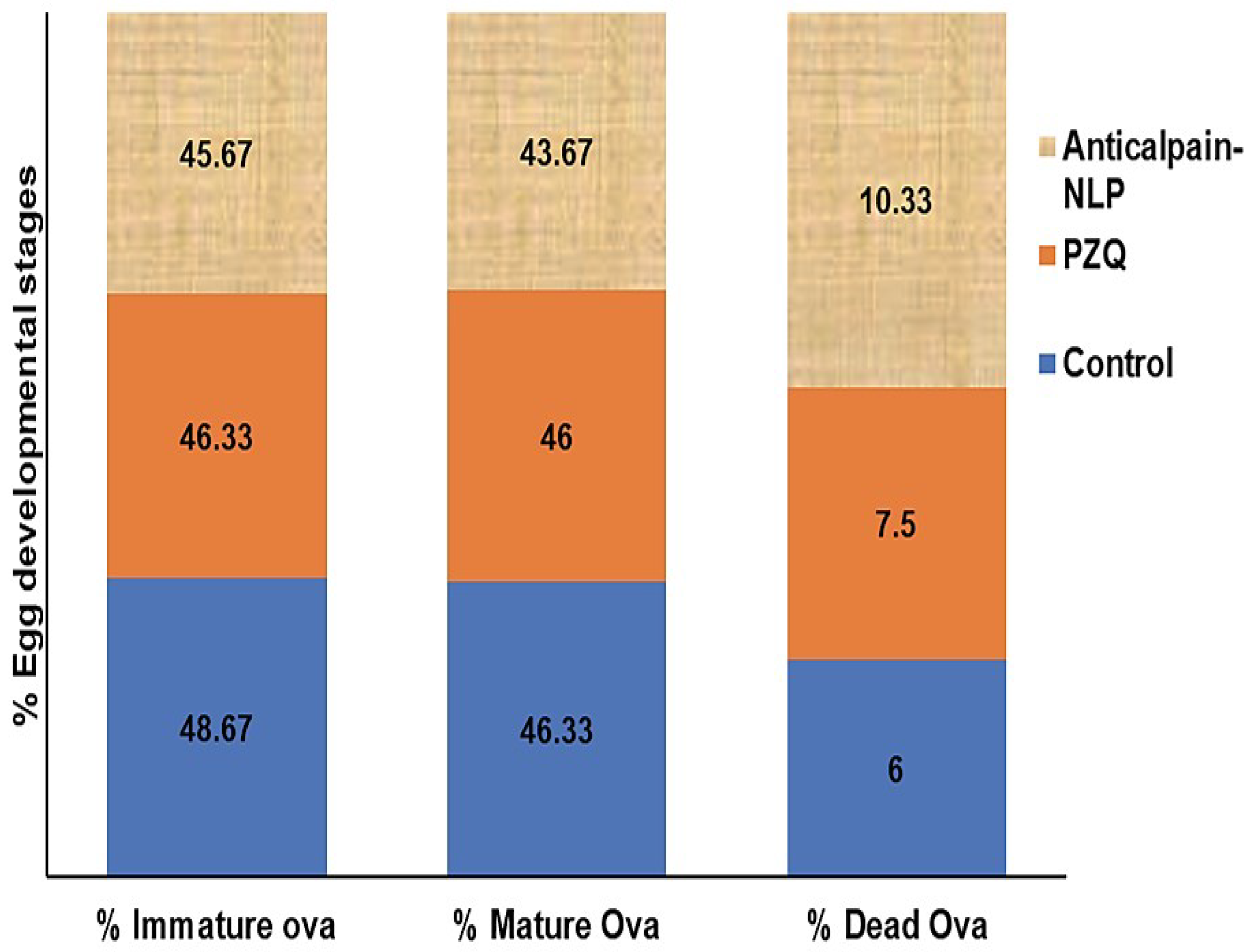
3.7. Assessment of Parasitological Cure Rate
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Adekiya, T.A.; Aruleba, R.T.; Oyinloye, B.E.; Okosun, K.O.; Kappo, A.P. The effect of climate change and the snail-schistosome cycle in transmission and bio-control of schistosomiasis in Sub-Saharan Africa. Int. J. Environ. Res. Public Health 2020, 17, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adekiya, T.A.; Kappo, A.P.; Okosun, K.O. Temperature and rainfall impact on schistosomiasis. Glob. J. Pure Appl. Math. 2017, 13, 8453–8469. [Google Scholar]
- Elflein, J. Statista Dossier on Neglected Tropical Diseases. 28 January 2021. Available online: https://0-www-statista-com.brum.beds.ac.uk/study/55475/neglected-tropical-diseases/ (accessed on 19 July 2021).
- McManus, D.P. Defeating schistosomiasis. N. Engl. J. Med. 2019, 381, 2567–2568. [Google Scholar] [CrossRef] [PubMed]
- Deol, A.K.; Fleming, F.M.; Calvo-Urbano, B.; Walker, M.; Bucumi, V.; Gnandou, I.; Tukahebwa, E.M.; Jemu, S.; Mwingira, U.J.; Alkohlani, A.; et al. Schistosomiasis—Assessing progress toward the 2020 and 2025 global goals. N. Engl. J. Med. 2019, 381, 2519–2528. [Google Scholar] [CrossRef]
- Hotez, P.J.; Engels, D.; Gyapong, M.; Ducker, C.; Malecela, M.N. Female genital schistosomiasis. N. Engl. J. Med. 2019, 381, 2493–2495. [Google Scholar] [CrossRef]
- Roberts, M.; Cross, J.; Pohl, U.; Lucas, S.; Dean, A. Cerebral schistosomiasis. Lancet Infect. Dis. 2006, 6, 820. [Google Scholar] [CrossRef]
- Liu, H.; Lim, C.T.; Feng, X.; Yao, Z.; Chen, Y.; Sun, H. MRI in cerebral schistosomiasis: Characteristic nodular enhancement in 33 patients. Am. J. Roentgenol. 2008, 191, 582–588. [Google Scholar] [CrossRef]
- Wan, H.; Lei, D.; Mao, Q. Cerebellar schistosomiasis: A case report with clinical analysis. Korean J. Parasitol. 2009, 47, 53–56. [Google Scholar] [CrossRef] [Green Version]
- Imai, K.; Koibuchi, T.; Kumagai, T.; Maeda, T.; Osada, Y.; Ohta. Cerebral schistosomiasis due to Schistosoma haematobium confirmed by PCR analysis of brain specimen. J. Clin. Microbiol. 2011, 49, 3703–3706. [Google Scholar] [CrossRef] [Green Version]
- Ravi, N.; Yi, W.; Yu, L.; Ping, H.; Hao, C. Cerebral schistosomiasis. SA J. Radiol. 2013, 17, 143–144. [Google Scholar] [CrossRef] [Green Version]
- Rose, M.F.; Zimmerman, E.E.; Hsu, L.; Golby, A.J.; Saleh, E.; Folkerth, R.D. Atypical presentation of cerebral schistosomiasis four years after exposure to Schistosoma mansoni. Epilepsy Behav. Case Rep. 2014, 2, 80–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaqout, A.; Abid, F.B.; Murshed, K.; Al-Bozom, I.; Al-Rumaihi, G.; Al Soub, H. Cerebral schistosomiasis: Case series from Qatar. Int. J. Infect. Dis. 2019, 86, 167–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aruleba, R.T.; Adekiya, T.A.; Oyinloye, B.E.; Masamba, P.; Mbatha, L.S.; Pretorius, A.; Kappo, A.P. PZQ therapy: How close are we in the development of effective alternative anti-schistosomal drugs? Infect. Disord. Drug Targets. 2019, 19, 337–349. [Google Scholar] [CrossRef] [PubMed]
- Vale, N.; Gouveia, M.J.; Rinaldi, G.; Brindley, P.J.; Gärtner, F.; Correia da Costa, J.M. Praziquantel for schistosomiasis: Single-drug metabolism revisited, mode of action, and resistance. Antimicrob. Agents Chemother. 2017, 61, e02582-16. [Google Scholar] [CrossRef] [Green Version]
- Xiao, S.H.; Sun, J.; Chen, M.G. Pharmacological and immunological effects of praziquantel against Schistosoma japonicum: A scoping review of experimental studies. Infect. Dis. Poverty 2018, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Doenhoff, M.J.; Cioli, D.; Utzinger, J. Praziquantel: Mechanisms of action, resistance and new derivatives for schistosomiasis. Curr. Opin. Infect. Dis. 2008, 21, 659–667. [Google Scholar] [CrossRef]
- Adekiya, T.A.; Kondiah, P.P.; Choonara, Y.E.; Kumar, P.; Pillay, V. A review of nanotechnology for targeted anti-schistosomal therapy. Front. Bioeng. Biotechnol. 2020, 8, 32. [Google Scholar] [CrossRef]
- Adekiya, T.A.; Kumar, P.; Kondiah, P.P.; Pillay, V.; Choonara, Y.E. Synthesis and therapeutic delivery approaches for praziquantel: A patent review (2010-present). Expert Opin. Ther. Pat. 2021, 31, 851–865. [Google Scholar] [CrossRef]
- Mufamadi, M.S.; Choonara, Y.E.; Kumar, P.; Modi, G.; Naidoo, D.; van Vuuren, S. Ligand-functionalized nanoliposomes for targeted delivery of galantamine. Int. J. Pharm. 2013, 448, 267–281. [Google Scholar] [CrossRef]
- Cheng, W.; Li, X.; Zhang, C.; Chen, W.; Yuan, H.; Xu, S. Preparation and In Vivo-In Vitro evaluation of polydatin-phospholipid complex with improved dissolution and bioavailability. Int. J. Drug Dev. Res. 2017, 9, 39–43. [Google Scholar]
- Li, S.; Bouchy, S.; Penninckx, S.; Marega, R.; Fichera, O.; Gallez, B. Antibody-functionalized gold nanoparticles as tumor-targeting radiosensitizers for proton therapy. Nanomedicine 2019, 14, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Dai, Q.; Yan, Y.; Ang, C.S.; Kempe, K.; Kamphuis, M.M.; Dodds, S.J. Monoclonal antibody-functionalized multilayered particles: Targeting cancer cells in the presence of protein coronas. ACS Nano 2015, 9, 2876–2885. [Google Scholar] [CrossRef] [Green Version]
- Dou, X.Q.; Wang, H.; Zhang, J.; Wang, F.; Xu, G.L.; Xu, C.C. Aptamer–drug conjugate: Targeted delivery of doxorubicin in a HER3 aptamer-functionalized liposomal delivery system reduces cardiotoxicity. Int. J. Nanomed. 2018, 13, 763–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, J.M.; Zupancic, E.; Vandermeulen, G.; Oliveira, V.G.; Salgado, A.; Videira, M. In Vivo delivery of peptides and Toll-like receptor ligands by mannose-functionalized polymeric nanoparticles induces prophylactic and therapeutic anti-tumor immune responses in a melanoma model. J. Control Release 2015, 198, 91–103. [Google Scholar] [CrossRef]
- Siddiqui, A.A.; Zhou, Y.; Podesta, R.B.; Karcz, S.R.; Tognon, C.E.; Strejan, G.H. Characterization of Ca2+-dependent neutral protease (calpain) from human blood flukes, Schistosoma mansoni. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 1993, 1181, 37–44. [Google Scholar] [CrossRef]
- Wang, Q.; Da’dara, A.A.; Skelly, P.J. The human blood parasite Schistosoma mansoni expresses extracellular tegumental calpains that cleave the blood clotting protein fibronectin. Sci. Rep. 2017, 7, 12912. [Google Scholar] [CrossRef] [PubMed]
- Skelly, P.J.; Kim, J.W.; Cunningham, J.; Shoemaker, C.B. Cloning, characterization, and functional expression of cDNAs encoding glucose transporter proteins from the human parasite Schistosoma mansoni. J. Biol. Chem. 1994, 269, 4247–4253. [Google Scholar] [CrossRef]
- Skelly, P.J.; Tielens, A.G.M.; Shoemaker, C.B. Glucose transport and metabolism in mammalian-stage schistosomes. Parasitol. Today 1998, 14, 402–406. [Google Scholar] [CrossRef]
- MacDonald, K.; Buxton, S.; Kimber, M.J.; Day, T.A.; Robertson, A.P.; Ribeiro, P. Functional characterization of a novel family of acetylcholine-gated chloride channels in Schistosoma mansoni. PLoS Pathog. 2014, 10, e1004181. [Google Scholar] [CrossRef] [Green Version]
- Mansour, T.; Mansour, J. Targets in the Tegument of Flatworms. In Chemotherapeutic Targets in Parasites: Contemporary Strategies; Cambridge University Press: Cambridge, UK, 2002; pp. 189–214. [Google Scholar]
- Song, S.; Liu, D.; Peng, J.; Sun, Y.; Li, Z.; Gu, J.R. Peptide ligand-mediated liposome distribution and targeting to EGFR expressing tumor in vivo. Int. J. Pharm. 2008, 363, 155–161. [Google Scholar] [CrossRef]
- Mossallam, S.F.; Abou-El-Naga, I.F.; Abdel Bary, A.; Elmorsy, E.A.; Diab, R.G. Schistosoma mansoni egg-derived extracellular vesicles: A promising vaccine candidate against murine schistosomiasis. PLoS Negl. Trop. Dis. 2021, 15, e0009866. [Google Scholar] [CrossRef] [PubMed]
- Abd El Wahab, W.M.; El-Badry, A.A.; Mahmoud, S.S.; El-Badry, Y.A.; El-Badry, M.A.; Hamdy, D.A. Ginger (Zingiber Officinale)-derived nanoparticles in Schistosoma mansoni infected mice: Hepatoprotective and enhancer of etiological treatment. PLoS Negl. Trop. Dis. 2021, 15, e0009423. [Google Scholar] [CrossRef] [PubMed]
- Kloetzel, K.U.R.T. A suggestion for the prevention of severe clinical forms of schistosomiasis mansoni. Bull World Health Organ. 1967, 37, 686–687. [Google Scholar]
- Mati, V.L.T.; Melo, A.L. Current applications of oogram methodology in experimental schistosomiasis; fecundity of female Schistosoma mansoni and egg release in the intestine of AKR/J mice following immunomodulatory treatment with pentoxifylline. J. Helminthol. 2013, 87, 115–124. [Google Scholar] [CrossRef]
- Aguilar-Pérez, K.M.; Avilés-Castrillo, J.I.; Medina, D.I.; Parra-Saldivar, R.; Iqbal, H. Insight into nanoliposomes as smart nanocarriers for greening the twenty-first century biomedical settings. Front. Bioeng. Biotechnol. 2020, 8, 1441. [Google Scholar] [CrossRef] [PubMed]
- Bozzuto, G.; Molinari, A. Liposomes as nanomedical devices. Int. J. Nanomed. 2015, 10, 579536. [Google Scholar] [CrossRef] [Green Version]
- Amara, R.O.; Ramadan, A.A.; El-Moslemany, R.M.; Eissa, M.M.; El-Azzouni, M.Z.; El-Khordagui, L.K. Praziquantel–lipid nanocapsules: An oral nanotherapeutic with potential Schistosoma mansoni tegumental targeting. Int. J. Nanomed. 2018, 13, 4493–4505. [Google Scholar] [CrossRef] [Green Version]
- Frezza, T.F.; Gremião, M.P.D.; Zanotti-Magalhães, E.M.; Magalhães, L.A.; de Souza, A.L.R.; Allegretti, S.M. Liposomal-praziquantel: Efficacy against Schistosoma mansoni in a preclinical assay. Acta Trop. 2013, 128, 70–75. [Google Scholar] [CrossRef]
- Daeihamed, M.; Dadashzadeh, S.; Haeri, A.; Faghih Akhlaghi, M. Potential of liposomes for enhancement of oral drug absorption. Curr. Drug Deliv. 2017, 14, 289–303. [Google Scholar] [CrossRef]
- Clayton, K.N.; Salameh, J.W.; Wereley, S.T.; Kinzer-Ursem, T.L. Physical characterization of nanoparticle size and surface modification using particle scattering diffusometry. Biomicrofluidics 2016, 10, 054107. [Google Scholar] [CrossRef] [Green Version]
- Danaei, M.; Dehghankhold, M.; Ataei, S.; Hasanzadeh Davarani, F.; Javanmard, R.; Dokhani, A. Impact of particle size and polydispersity index on the clinical applications of lipidic nanocarrier systems. Pharmaceutics 2018, 10, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahari, L.A.S.; Hamishehkar, H. The impact of variables on particle size of solid lipid nanoparticles and nanostructured lipid carriers; a comparative literature review. Adv. Pharm. Bull. 2016, 6, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.K.; Harde, H.; Thanki, K.; Jain, S. Improved stability and antidiabetic potential of insulin containing folic acid functionalized polymer stabilized multilayered liposomes following oral administration. Biomacromolecules 2014, 15, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Patra, J.K.; Das, G.; Fraceto, L.F.; Campos, E.V.R.; del Pilar Rodriguez-Torres, M.; Acosta-Torres, L.S. Nano based drug delivery systems: Recent developments and future prospects. J. Nanobiotechnol. 2018, 16, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lombardo, D.; Kiselev, M.A.; Caccamo, M.T. Smart nanoparticles for drug delivery application: Development of versatile nanocarrier platforms in biotechnology and nanomedicine. J. Nanomater. 2019, 2019, 3702518. [Google Scholar] [CrossRef]
- Al-Khoury, H.; Espinosa-Cano, E.; Aguilar, M.R.; Román, J.S.; Syrowatka, F.; Schmidt, G. Anti-inflammatory surface coatings based on polyelectrolyte multilayers of heparin and polycationic nanoparticles of naproxen-bearing polymeric drugs. Biomacromolecules 2019, 20, 4015–4025. [Google Scholar] [CrossRef]
- Campani, V.; Giarra, S.; De Rosa, G. Lipid-based core-shell nanoparticles: Evolution and potentialities in drug delivery. OpenNano 2018, 3, 5–17. [Google Scholar] [CrossRef]
- Ghasemiyeh, P.; Mohammadi-Samani, S. Solid lipid nanoparticles and nanostructured lipid carriers as novel drug delivery systems: Applications, advantages and disadvantages. Res. Pharm. Sci. 2018, 13, 288–303. [Google Scholar]
- Lien, C.F.; Molnár, E.; Toman, P.; Tsibouklis, J.; Pilkington, G.J.; Górecki, D.C. In Vitro assessment of alkylglyceryl-functionalized chitosan nanoparticles as permeating vectors for the blood–brain barrier. Biomacromolecules 2012, 13, 1067–1073. [Google Scholar] [CrossRef]
- Elbaz, T.; Esmat, G. Hepatic and intestinal schistosomiasis. J. Adv. Res. 2013, 4, 445–452. [Google Scholar] [CrossRef] [Green Version]
- McManus, D.P.; Dunne, D.W.; Sacko, M.; Utzinger, J.; Vennervald, B.J.; Zhou, X.N. Schistosomiasis. Nat. Rev. Dis. Prim. 2018, 4, 13. [Google Scholar] [CrossRef] [PubMed]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
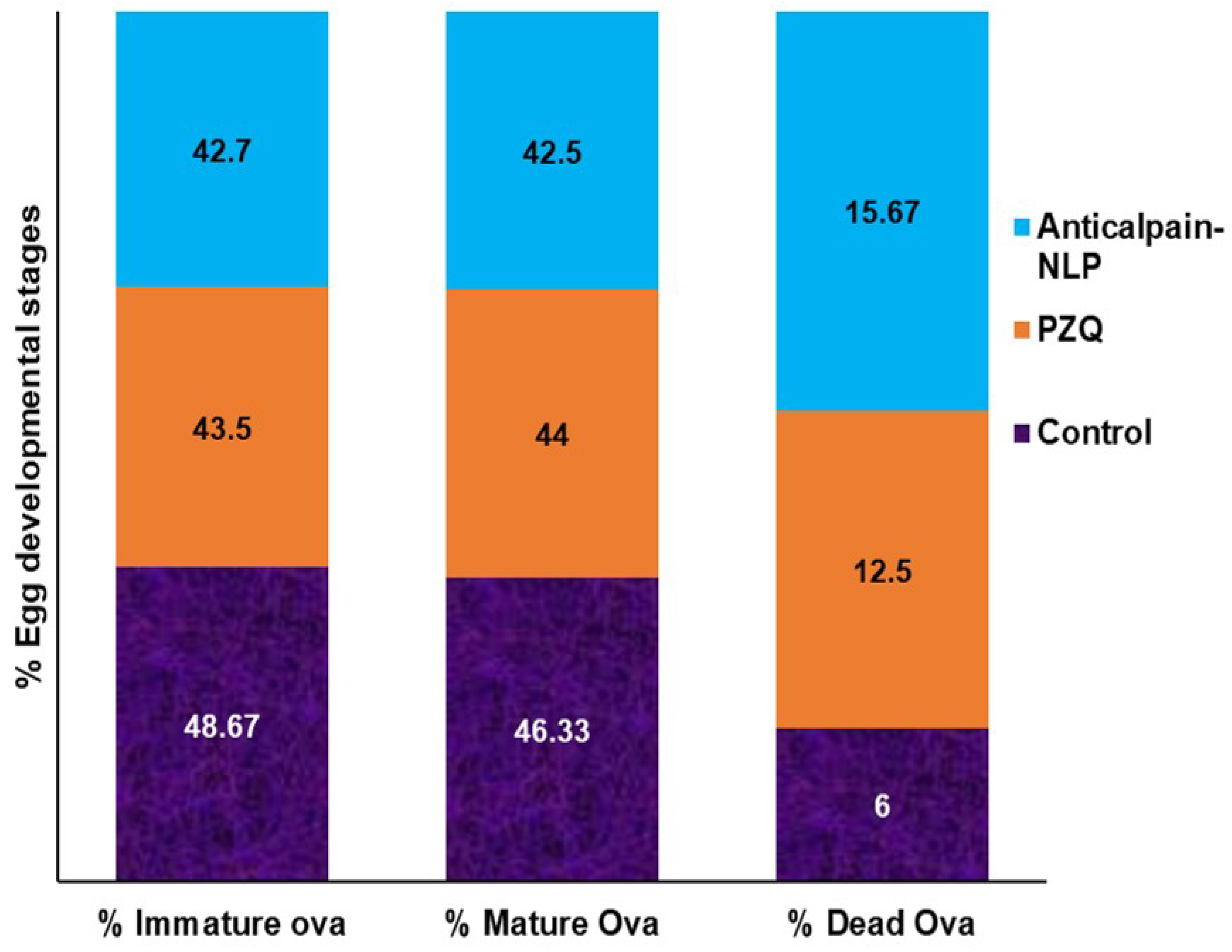{kind=link}
| Mean Worm Burden ±SD | % Reduction in Total Worm Burden | ||||
|---|---|---|---|---|---|
| (Liver and Porto-Mesenteric) | |||||
| Male | Female | Couples | Total | ||
| Control | 2.33 ± 0.81 | 0.33 ± 0.52 | 6.17 ± 0.75 | 15 ± 0.89 | |
| PZQ | 1.33 ± 1.21 | 0 | 4.33 ± 0.82 | 10.0 ± 1.41 | 33.3 |
| Antical-NLP | 1.10 ± 0.52 | 0.17 ± 0.41 | 3.67 ± 1.37 | 6.83 ± 2.4 | 54.5 |
| Mice Group | Liver | % Reduction in Ova Count in Liver | Intestine | % Reduction in Ova Count in the Intestine |
|---|---|---|---|---|
| Control | 28,202 ± 4372 | 31,902 ± 4342 | ||
| PZQ | 20,303 ± 2175 | 28.00 | 22,702 ± 5347 | 28.84 |
| Antical-NLP | 12,387 ± 2951 | 56.07 | 13,436 ± 2332 | 57.88 |
| Mean Worm Burden ±SD | % Reduction in Total Worm Burden | ||||
|---|---|---|---|---|---|
| (Liver and Porto-Mesenteric) | |||||
| Male | Female | Couples | Total | ||
| Control | 2.33 ± 0.81 | 0.33 ± 0.52 | 6.17 ± 0.75 | 15.00 ± 0.89 | |
| PZQ | 1.50 ± 1.40 | 0 | 2.00 ± 0.89 | 5.50 ± 2.60 | 63.3 |
| Antical-NLP | 1.00 ± 0.63 | 0 | 1.50 ± 0.83 | 4.17 ± 1.47 | 72.2 |
| Mice Group | Liver | % Reduction in Ova Count in Liver | Intestine | % Reduction in Ova Count in the Intestine |
|---|---|---|---|---|
| Control | 28,202 ± 4372 | 31,902 ± 4342 | ||
| PZQ | 13,626 ± 2936 | 51.68 | 14,658 ± 3699 | 54.05 |
| Antical-NLP | 10,112 ± 3745 | 64.14 | 9310 ± 3789 | 70.81 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adekiya, T.A.; Kumar, P.; Kondiah, P.P.D.; Choonara, Y.E. In Vivo Evaluation of an Antibody-Functionalized Lipoidal Nanosystem for Schistosomiasis Intervention. Pharmaceutics 2022, 14, 1531. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics14081531
Adekiya TA, Kumar P, Kondiah PPD, Choonara YE. In Vivo Evaluation of an Antibody-Functionalized Lipoidal Nanosystem for Schistosomiasis Intervention. Pharmaceutics. 2022; 14(8):1531. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics14081531
Chicago/Turabian StyleAdekiya, Tayo A., Pradeep Kumar, Pierre P. D. Kondiah, and Yahya E. Choonara. 2022. "In Vivo Evaluation of an Antibody-Functionalized Lipoidal Nanosystem for Schistosomiasis Intervention" Pharmaceutics 14, no. 8: 1531. https://0-doi-org.brum.beds.ac.uk/10.3390/pharmaceutics14081531