1. Introduction
Inflammatory bowel diseases (IBDs) constitute a continuum of refractory autoimmune inflammatory disorders ranging from ulcerative colitis (UC) to Crohn’s disease (CD) [
1]. Inflammation is the basic underlying mechanism of these disorders, yet the exact etiology of IBDs has not been fully elucidated [
2]. Studies have documented that IBDs are the result of a plethora of interactions of genetic and non-genetic parameters involving the microbiome, the environment, and the immune system, which plays a crucial role in initiating an excessive inflammatory response in genetically susceptible individuals [
3,
4]. Consequently, in this process, a panoply of pro-inflammatory and anti-inflammatory secretions interact through various signaling pathways [
5].
Moreover, cell adhesion molecules (CAMs) have been implicated in the pathogenesis of IBDs, as a reduction in epithelial matrix adhesion is likely to cause decreased healing [
6]. For example, lower levels of syndecan-1 were found in the colon epithelium of IBD patients [
7]. In addition, the microbiome, whether dysbiotic or eubiotic, is an essential player in such a pathogenic process, as demonstrated in multiple human and animal studies [
8].
Accordingly, the treatment targets and modalities of IBDs have profoundly changed over recent years following discoveries of the multiple pathogenic pathways involved; currently, they are stressing healing rather than symptom control [
9]. On this basis, various management protocols for IBD included immunomodulators like cyclosporine and anti-cytokine antibodies like anti-TNF-α and anti-IL23 [
9]. In addition, the approach of using probiotics has been adopted in multiple instances with encouraging results both in animal and human studies but never in the presence of cyclosporine [
10]. Cyclosporine A (CyA), an immunosuppressant that interacts with cytoplasmic cyclophilin A (CypA) [
11,
12], was introduced in the mid-1980s. In addition, probiotics have been used in the last two decades to correct dysbiosis [
13].
IL-6 has a significant role in the etiology of IBDs through a pro-inflammatory effect [
14,
15]. Similarly, CD3 and CD147 are upregulated in IBDs, playing essential roles in modulating inflammation [
16,
17]. The activation of the PI3K/Akt/mTOR pathway in lymphocytes also leads to their activation and, hence, leads to IBD [
18]. Furthermore, many of the key cell–cell and cell–matrix interactions are regulated by beta integrins, whose deficiency may significantly enhance both the pathogenesis and development of IBDs [
19].
The data from the literature report that each of the adopted management protocols has not been without limitations and drawbacks. For instance, CyA exhibited several complications in terms of toxicity to various body organs [
20]. Accordingly, it was believed that the administration of the CypA compound that interacts with CyA would enhance its immunosuppressant effect and probably decrease its side effects [
21].
Knowing that the absence of syndecan-1 affects the gut microbiota and disrupts the homeostasis of microbes internally, increasing the likelihood of IBD development, and supported by data regarding polymorphisms in IBD susceptibility genes, this study explored and compared the IBD pathogenic process in syndecan-1 KO mice compared to wild-type Balb/c mice [
22,
23,
24]. Such an approach to IBDs will possibly pave the way to understanding the many mechanistic pathways involved in maintaining intestinal homeostasis. It will also show the potential difference in DSS-IBD induction between various mice strains and the possible role of syndecan-1 in such a process. Furthermore, this study assessed the effectiveness of CyA in the management of IBDs and the potential role of CypA in enhancing the healing process of an inflamed gut, particularly in the presence of probiotics. In brief, it highlights the potential role of probiotics in IBDs in syndecan-1 KO mice compared to wild type.
2. Materials and Methods
2.1. Animals
A total of 112, 6–8-week-old male mice were divided into 2 main groups: Wild-type Balb/c mice and KO Syndecan-1 null mice (56 mice of each strain). All the animals were housed in the Animal Care Facility of the American University of Beirut (AUB). All animal experiments and procedures strictly followed the guidelines of the Institutional Animal Care and Use Committee (IACUC) at the American University of Beirut for the care and use of laboratory animals.
2.2. Experimental Design
A total of 16 control mice—8 from each strain—were only provided with normal drinking water and intraperitoneal (IP) saline injections. IBD was induced in 48 animals from each strain (96 mice) by 2% DSS in drinking water. Each DSS cycle consisted of 7 days of DSS followed by 2 weeks of normal drinking water. At week 3, the animals were sacrificed.
The 96 mice with DSS induced IBD were divided according to the different treatment modalities into a total of 12 subgroups (8 animals per group). Treatments were administered as illustrated in
Table 1.
2.3. Induction of IBD and Treatments
Optimized concentration of the pro-inflammatory agent dextran sodium sulfate (DSS; Sigma-Aldrich, St. Louis, MO, USA, 42867-100G) 2% was prepared in autoclaved water and administered to animals in their drinking water. Each DSS cycle consisted of 7 days of DSS followed by 2 weeks normal drinking water and treatments.
Cyclophilin A (human recombinant expressed in E-coli) from Sigma-Aldrich (C3805-1MG) was injected intraperitoneally (IP) at a dose of 25 µg/kg/day for two weeks starting day 7 of DSS administration. Similarly, cyclosporine A (Novartis, Basel, Switzerland, SPE31) was administered by IP injections at a concentration of 200 µg every other day for 2 weeks starting day 7 of DSS treatment. In addition, the probiotics (P) used was a mixture of 7 strains of lactic acid-producing bacteria: Lactobacillus rhamnosus, Saccharomyces boulardii, Bifidobacterium breve, Bifidobacterium lactis, Lactobacillus acidophilus, Lactobacillus plantarum and Lactobacillus reuteri. One capsule of (P) was dissolved in 1.75 L of autoclaved tap water to reach a daily dose of 108 CFU per animal and given for 2 weeks starting day 7 of DSS treatment.
2.4. Clinical Course Assessment
During the experimental period, the animals were monitored daily for clinical symptoms and signs including body weight, stool aspect, and rectal bleeding. Scores were recorded and calculated throughout the experiment based on the parameters: gross bleeding (0 = absence; 2 = blood stained; 4 = clear presence of blood), stool consistency and watery diarrhea (0 = normal, 2 = loose, 4 = diarrhea), weight loss (0 = normal; 1 = 1–5%; 2 = 5–10%; 3 = 10–20%; 4 more than 20%), and a previously validated clinical disease activity index, (DAI) which assesses weight loss, diarrhea, fur quality, anal bleeding and posture with a range of 0 to 4 calculated [
25].
2.5. Measurement of Fecal Occult Blood
Collection of feces was completed by placing a single mouse in an empty cage without bedding material for a few minutes; feces were collected, and occult blood was measured using HemoCue America Beckman Coulter™ Hemoccult™ Fecal Occult Blood Slide Test System (Beckman Coulter, Brea, CA, USA), as per the manufacturer’s instructions [
26].
2.6. Dissection, Colon Length Measurement and Biopsy Removal
After 3 weeks, at the experiment endpoint, animals were sacrificed by isofurane overdose and cervical dislocation; then, they were dissected in order to remove their colon. We measured and recorded the length of each isolated colon from the ileocecal valve to the rectum using a ruler; then, it was quickly flushed on ice with phosphate-buffered saline (PBS) to clean it. A portion of this clean colon, the sigmoid, was fixed in 10% buffered formalin for routine histological processing. The other portion of the sigmoid and the descending colon were frozen and kept in liquid nitrogen for further molecular studies.
2.7. Histology
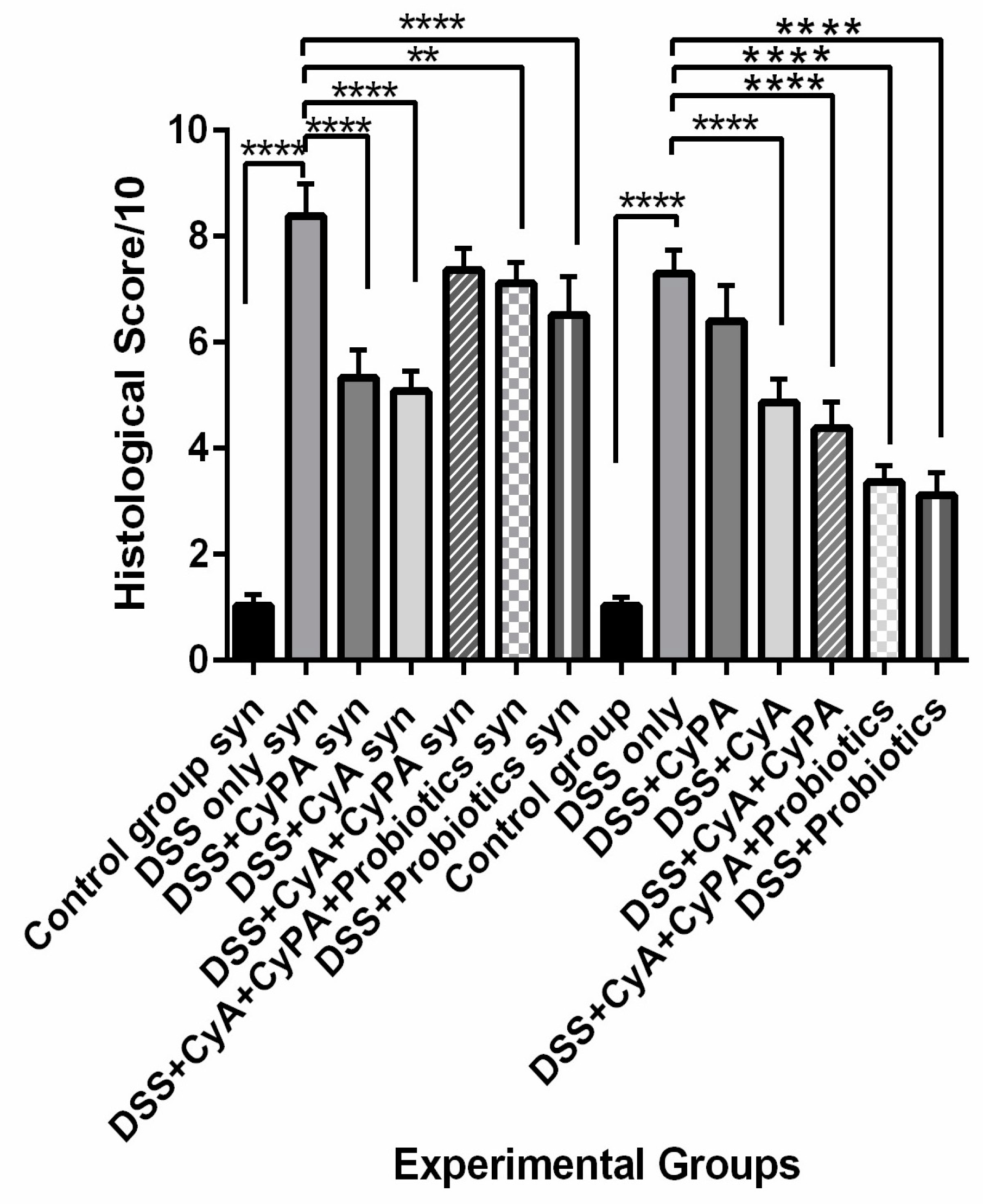
Biopsies were embedded in paraffin and seven slides of each biopsy, each having 2 sections, were stained either with hematoxylin and eosin (H&E) or periodic acid–Schiff (PAS). The slides were finally scanned using a light microscope. The histological score of each tissue section of each animal within each group was assessed and calculated based on a histological scoring system shown in
Table 2 [
27]. The different sections were photographed using an Olympus CX-41 microscope (Olympus, Tokyo, Japan).
2.8. Western Blot
Western blotting analysis was performed using 100 mg of the frozen colon tissue according to standard protocols. Protein concentration was determined by the Lowry method using the DC™ Protein Assay Kit (#5000111, BIO-RAD, Hercules, CA, USA). Antibodies against IL-6 (anti-mouse SC-57315), CD147 (anti-mouse SC-46700), Actin (anti-mouse SC-47778), CD3 (anti-mouse 20047), and pAKT (anti-mouse SC57315) were detected using a secondary antibody (horseradish peroxidase-conjugated anti-mouse; Abcam 97,046 at a dilution of 3:40,000 (Abcam, Boston, MA, USA)). The immunoprecipitated protein bands were detected with ChemiDoc MP Imaging System-Biorad (BioRad, Hercules, CA, USA).
2.9. Immunohistochemistry
Immunohistochemistry was performed on 5-micron thick paraffin embedded sections. Slides were stained with a primary antibody (anti-integrin ß1-JB 1B sc-59,829 mouse IgG from Santa Cruz Biotechnology, Dallas, TX, USA). Quantification was performed by obtaining the measurement of mean fluorescence intensity (MFI) of integrin ß1 in a region of interest (ROI) and calculating the integrated density using Zen 2.3 Software (Carl Zeiss Microscopy GmbH, Oberkochen, Germany) [
28].
2.10. Statistical Analysis
Statistical analysis was performed using GraphPad Prism 8.0.1. Data were expressed as a mean ± standard deviation. Significant differences were evaluated using the one way ANOVA by the Tukey–Kramer multiple comparisons test. A value of p < 0.05 was considered significant.
4. Discussion
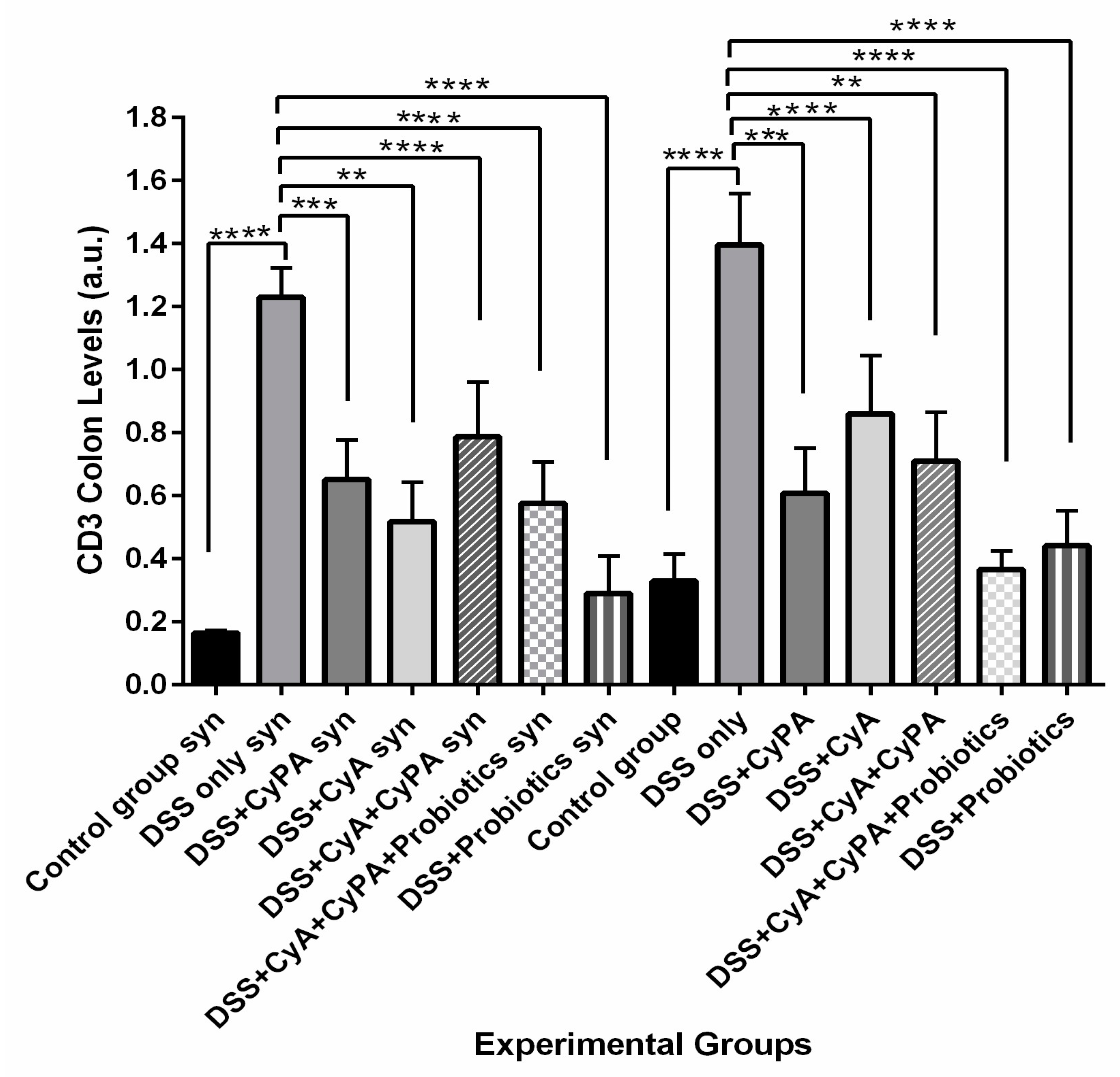
Data from this study highlighted the role of syndecan-1 in the pathogenesis and management of IBD in combination with the CypA, CyA complex and probiotics. In comparison to the wild type, the inflammation was more severe in the KO mice, and treatment with CyA led to a greater reduction in inflammation and a greater suppression of expression of immune cell markers. However, the CypA-CyA complex was less effective in controlling the disease and caused a greater exacerbation of inflammation in IL-6, CD147 and CD3 in the KO than in the wild type. On the other hand, when probiotics were added to the CypA-CyA complex, they had less effect in controlling IBD inflammatory markers in the KO compared to more effect in the wild type. Probiotics used alone significantly ameliorated the health status in both strains but more so in the wild type.
In the absence of syndecan-1, the mice were more sensitive to DSS inflammatory stimulation. Actually, the loss of syndecan-1 contributes to epithelial permeability and defect in the barrier, leading to microbiota access to lamina propria and causing a larger chronic and persistent inflammatory reaction and accumulation of inflammatory cells [
29]. In its modulation of different proteolytic activities, syndecan-1 plays a crucial role in a plethora of biological processes including cell proliferation, differentiation and redifferentiation [
30]. These functions are achieved through its role in integrin activity and migration as well as enhancing the motility of macrophages. Syndecan-1 is also associated with an anti-inflammatory M2 macrophage polarization involved in maintaining the function of the mucosa barrier and the restoration of tight junctions to maintain epithelial cell integrity [
29]. The presence of syndecan-1 could also inhibit the secretion of pro-inflammatory cytokines due to its suppressed ectodomain shedding, thus causing an amelioration of intestinal inflammation and neutrophil transmigration [
31]. In this study, CyA was very effective in controlling the symptoms, improving and limiting histological alterations, and decreasing pro-inflammatory cytokines expressions. Such effects were more significant in the KO mice. Immunoregulators, including the classical immunosuppressive drug CyA, have been used in IBD therapy, in particular in ulcerative colitis [
32,
33,
34,
35]. CyA downregulates the activation of T-lymphocytes by blocking the production of IL-2, inhibits the production of pro-inflammatory cytokines and stimulates apoptosis [
32,
33,
34,
35]. Its mechanism of action starts when it binds to CypA, an intracellular binding protein for CyA, forming a Cyp-CypA complex, thus affording anti-inflammatory activity (more so in KO mice) [
36].
Moreover, the CypA treatment in the syndecan-1 KO mice also showed a decrease in the inflammation originally caused by the DSS, which is contrary to what is being reported in the literature [
37,
38,
39,
40,
41]. Furthermore, CypA, which is the primary cytosolic binding protein of CyA, was provided intraperitoneally, thus creating a high concentration of extracellular CypA, which can potentially bind to and block CD147 instead of stimulating their chemotactic activity.
Previous work has proven that pAkt is implicated in IBD pathogenesis where a significant increase in its expression has been reported in a DSS model of colitis [
42]. The pAkt, by going back to its normal control levels, deactivated by CypA, showed that CypA could interplay with the Akt pathway and somehow downregulates its activity in both strains. Such an effect is expressed in both strains and slightly more in the KO mice with respect to better tissue repair, more IL-6 suppression, more CD147 suppression and less beta 1 integrins expression, which favors better tissue repair.
Concerning the use of the combination of CyA + CypA, the data of this study showed no added effect in reducing further the inflammatory reaction in the intestines. On the contrary, the inflammation continued in the KO mice with little effect versus a more significant added effect in the wild type as expressed in the lowest pAkt, lowest beta 1 integrins levels, lower CD147 levels and lower IL-6 levels.
These findings are not all in line with reports from the literature. The maintenance of inflammation in this combination group could probably be ascribed to the excess CypA administered along with the cyclosporine, thus forming a CypA + CyA complex that could probably, in the absence of syndecan-1, stimulate a significant increase in CD3, CD147 and even IL-6. Such changes in the KO group probably activated other signaling pathways, leading to a continuous exacerbation of the disease.
The addition of probiotics to the combination lead to an overall improvement—more so in the wild type than in the syndecan-1 KO mice. Despite the persistent inflammation in the combination with probiotics, the clinical signs and symptoms were improved, which reflects a better preserved intestinal barrier by the presence of probiotics. More investigations are needed to unveil the mechanism of action of the complex in the presence of probiotics.
On the other hand, the use of probiotics in the absence of the CypA-CyA complex revealed a marked reduction in inflammation in both strains but significantly more in the wild-type mice. At the same time, the histology of the sigmoid colon was almost normal and preserved the architecture in 75% of the section, yet very active lymphocyte aggregates existed with a relatively normal epithelial barrier. Regarding the molecular parameters, probiotics alone were more efficient than both drugs in reducing the inflammatory molecules in the colon. This result is in line with previous studies which have evidently shown that probiotics stimulate the differentiation of T-helper 1 cells, boost antibody production, promote the activity of both natural killer cells and phagocytic cells, and increase T-cell apoptosis by inhibiting the transcription of NF-ΚB. In addition, probiotics increase the production of anti-inflammatory cytokines and decrease pro-inflammatory cytokines [
43,
44,
45,
46]. Moreover, probiotics prevent the apoptosis of intestinal epithelial cells and stimulate the production of proteins essential for tight junction preservation, thus decreasing the paracellular permeability and restoring barrier function; and the effects of probiotics are in agreement with our results [
47,
48,
49,
50]. On the other hand, probiotics produce bacteriocins, thus creating an acidic medium detrimental to pathogenic bacteria yet favorable to the growth of beneficial microorganisms such as lactobacilli and bifidobacteria [
51,
52,
53,
54]. Furthermore, it has been shown that probiotics reduce the total T-cells and increase the number of Treg cells in the colonic tissue and blood in addition to enhancing the function of tight junctions [
55].
5. Conclusions
The potent role of cyclosporine in IBD therapy has been confirmed in both strains as demonstrated by the marked reduction in inflammation taking place particularly in the DSS + CyA group. Altogether, our findings suggest a therapeutic role for CypA in DSS-induced sdc-1 deficient mice. The presence of distinct receptors for extracellular CypA (other than CD147 and sdc-1) on its target cell merits further exploration. However, the low inhibitory effect of inflammation in the group treated with the complex group (CypA-CyA) needs further investigation, since it is recurring in both strains but shows relatively more inhibition in the wild type. Moreover, the effectiveness of probiotics has been clearly revealed when used alone in DSS-induced sdc-1 deficient mice as well as the Balb/c wild type. In contrast, this effectiveness has been partially inhibited in the presence of the CypA-CyA complex. The differential effects of CyA, CypA, probiotics and their combinations on the various inflammatory markers as well as the histological alterations and clinical signs and symptoms speak in favor of an anti-inflammatory role of syndecan-1. However, probiotics need to be considered after more exploration on the mechanisms involved in the presence of CypA and CyA, especially since pAkt was less active in their presence in both strains. Further and deeper investigations need to be carried out to answer the question of how the CypA-CyA complex decreases the anti-inflammatory effect of probiotics and ascertain the likely mechanisms involved.
Lastly, this study has a limitation with respect to the number of animals. Actually, the syndecan-1 KO mice were difficult to breed and achieved good numbers of progenies. This led to a decrease in the types of groups, whereby a group of CyA with probiotics and a group of CypA with probiotics could have been added in order to have a more complete picture. Another limitation included the study of other inflammatory markers like TGFβ and TNFα, which could have been useful in exploring other relevant signaling pathways that are probably involved in the pathogenic process.


 and
and 











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}