Effects on Carbon Sources and Sinks from Conversion of Over-Mature Forest to Major Secondary Forests and Korean Pine Plantation in Northeast China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Plot Layout
2.2. Study Measurements
2.2.1. Carbon Sequestration and NPP
2.2.2. Soil Gas Fluxes
2.2.3. Soil Properties Analysis
2.2.4. Carbon Source and Sink Analysis
2.3. Calculation and Statistical Analysis
3. Results
3.1. Soil Properties
3.2. Emission of GHGs
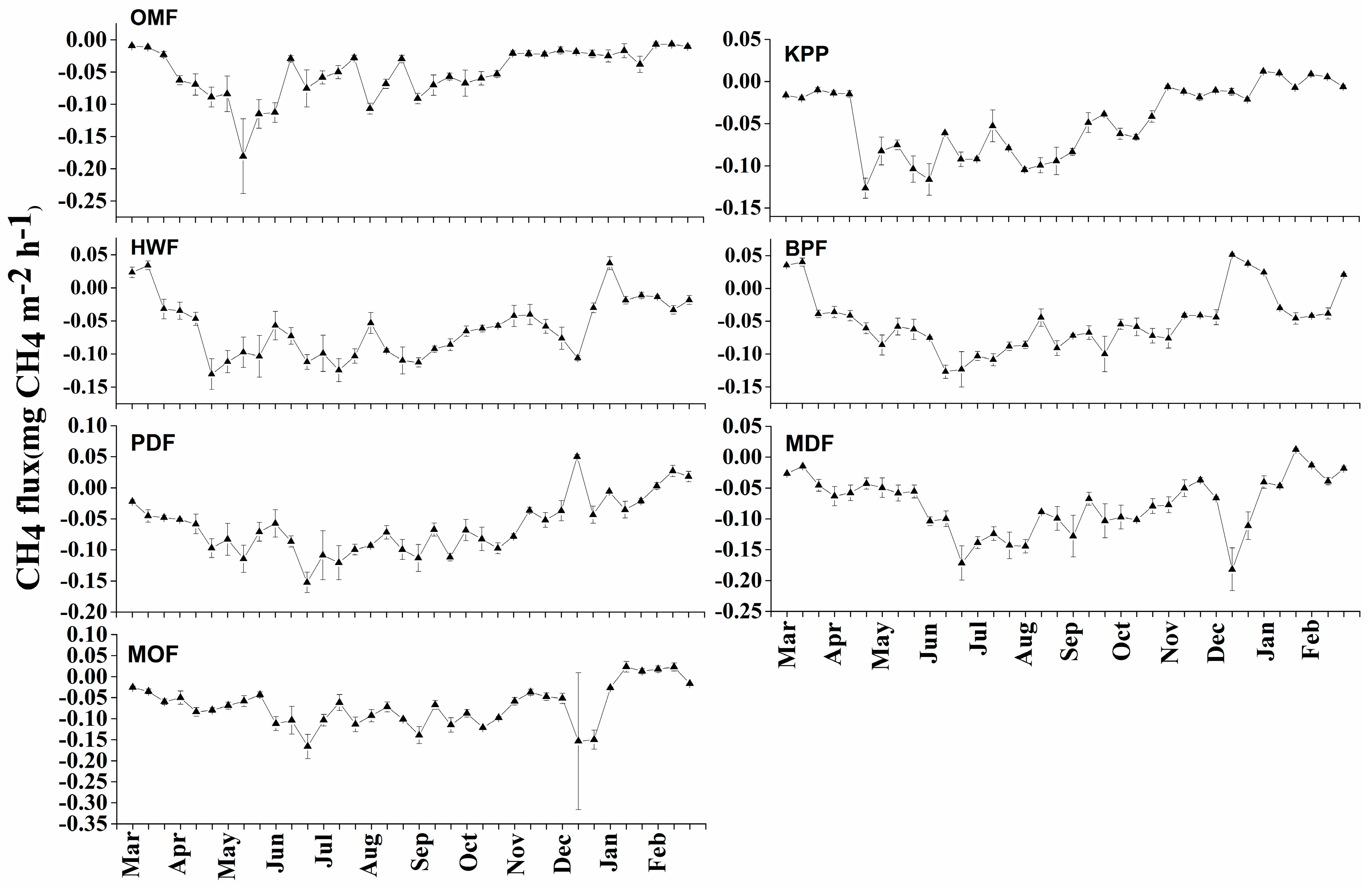
3.2.1. Methane Emission
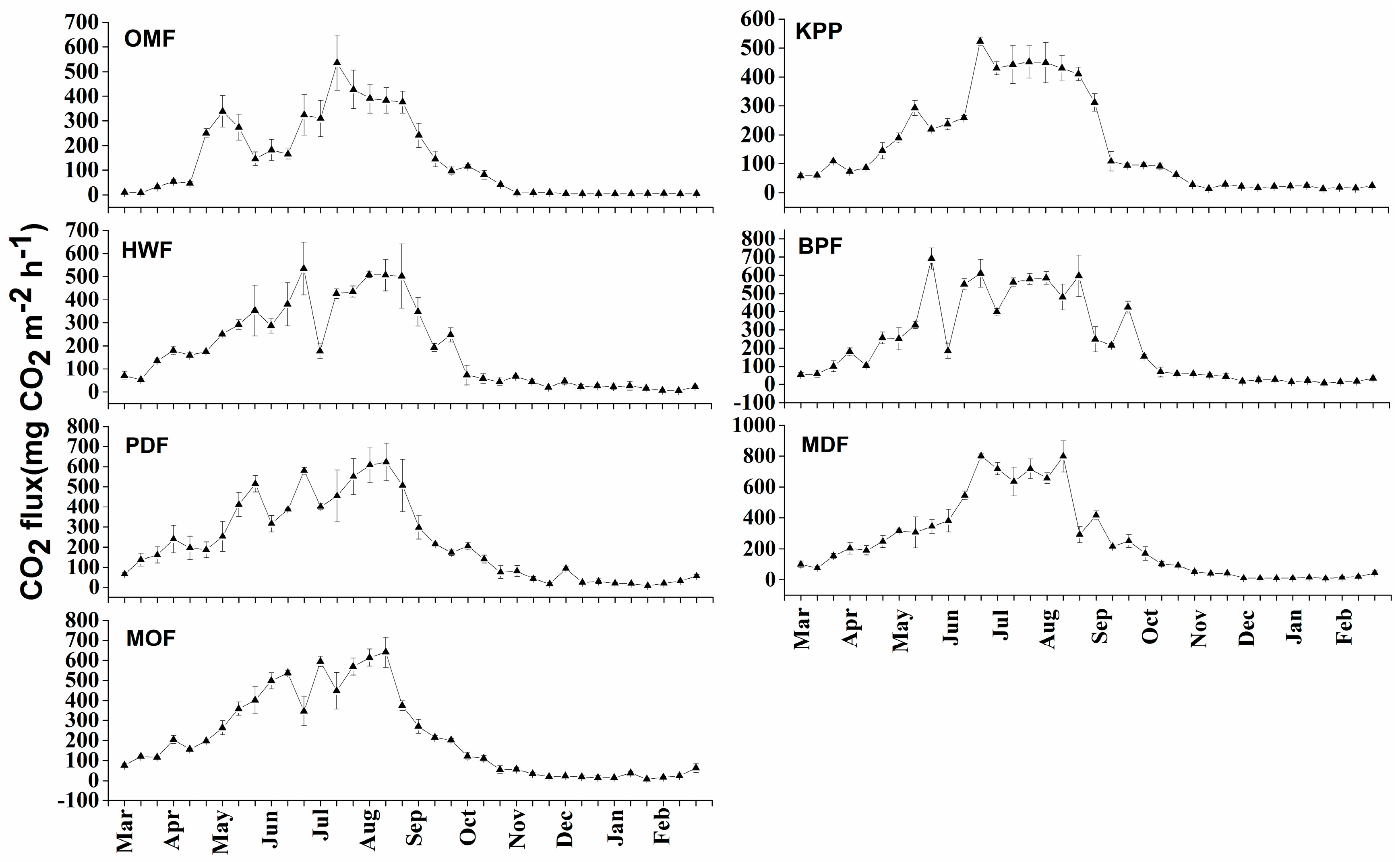
3.2.2. Carbon Dioxide Emission
3.2.3. Key Soil Factors Affecting CH4 and CO2 Fluxes
3.3. Net Primary Productivity and Annual Net C Sequestration
3.4. Carbon Sources and Sinks
4. Discussion
4.1. Carbon Fluxes
4.1.1. Methane Emission and Seasonal Dynamics
4.1.2. Carbon Dioxide Emission and Seasonal Dynamics
4.1.3. Emission Factors
4.2. Net Primary Productivity and Net Carbon Sequestration
4.3. Carbon Balance
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schimel, D.S.; House, J.I.; Hibbard, K.A.; Bousquet, P.; Ciais, P.; Peylin, P.; Braswell, B.H.; Apps, M.J.; Baker, D.; Bondeau, A. Recent patterns and mechanisms of carbon recent patterns and mechanisms of carbon exchange by terrestrial ecosystems. Nature 2001, 414, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Shiogama, H.; Ogura, T. Climate science: Clouds of uncertainty. Nature 2014, 515, 34. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Melillo, J.M.; Kicklighter, D.W.; McGuire, A.D.; Helfrich, J. The sensitivity of terrestrial carbon storage to historical climate variability and atmospheric CO2 in the United States. Tellus B Chem. Phys. Meteorol. 1999, 51, 414–452. [Google Scholar] [CrossRef]
- Gundersen, P.; Christiansen, J.R.; Alberti, G.; Brüggemann, N.; Castaldi, S.; Gasche, R.; Kitzler, B.; Klemedtsson, L.; Lobo–do–Vale, R.; Moldan, F.; et al. The response of methane and nitrous oxide fluxes to forest change in Europe. Biogeosciences 2012, 9, 6129–6168. [Google Scholar] [CrossRef]
- Dixon, R.K.; Solomon, A.M.; Brown, S.; Houghton, R.A.; Trexier, M.C.; Wiśniewski, J. Carbon Pools and Flux of Global Forest Ecosystems. Science 1994, 263, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.Q.; Wang, S.L.; Zhang, C.M.; Gong, C.; Hu, Q. Carbon storage in evergreen broad–leaf forests in mid–subtropical re–gion of China at four succession stages. J. For. Res. 2013, 24, 677–682. [Google Scholar] [CrossRef]
- Callewaert, P.; Lenders, S.; Gryze, S.D.; Morris, S.J.; Gregorich, E.G.; Paul, E.A.; Paustian, K. Measuring and understanding carbon storage in afforested soils by physical fractionation. Soil Sci. Soc. Am. J. 2007, 66, 1981–1987. [Google Scholar]
- Vogt, K. Carbon budgets of temperate forest ecosystems. Tree Physiol. 1991, 9, 69–86. [Google Scholar] [CrossRef]
- Luyssaert, S.; Inglima, I.; Jung, M.; Richardson, A.D.; Reichsteins, M.; Papale, D.; Piao, S.l.; Schulzes, E.D.; Wingate, L.; Matteucci, G.; et al. CO2 balance of boreal, temperate, and tropical forests derived from a global database. Glob. Chang. Biol. 2007, 13, 2509–2537. [Google Scholar] [CrossRef]
- Veiko, U.; Mai, K.; Jürgen, A.; Mats, V.; Hardo, B.; Gunnar, M.; Kalle, K. Ecosystems carbon budgets of differently aged downy birch stands growing on well–drained peatlands. For. Ecol. Manag. 2017, 399, 82–93. [Google Scholar]
- Valentini, R.; Matteucci, G.; Dolman, A.J.; Schulze, E.-D.; Rebmann, C.; Moors, E.J.; Granier, A.; Gross, P.; Jensen, N.O.; Pilegaard, K.; et al. Respiration as the main determinant of carbon balance in European forests. Nature 2000, 404, 861–865. [Google Scholar] [CrossRef]
- Liski, J.; Nissinen, A.; Erhard, M.; Taskinen, O. Climatic effects on litter decomposition from Arctic tundra to tropical rainforest. Glob. Chang. Biol. 2010, 9, 575–584. [Google Scholar] [CrossRef]
- Kaipainen, T.; Liski, J.; Pussinen, A.; Karjalainen, T. Managing carbon sinks by changing rotation length in European forests. Environ. Sci. Policy 2004, 7, 205–219. [Google Scholar] [CrossRef]
- Pacala, S.W.; Hurtt, G.C.; Baker, D.; Peylin, P.; Houghton, R.A.; Birdsey, R.A.; Heath, L.; Sundquist, E.T.; Stallard, R.; Ciais, P.; et al. Consistent Land- and Atmosphere-Based U.S. Carbon Sink Estimates. Science 2001, 292, 2316–2320. [Google Scholar] [CrossRef] [Green Version]
- Law, B.E.; Thornton, P.E.; Irvine, J.; Anthoni, P.M.; Tuyl, S.V. Carbon storage and fluxes in ponderosa pine forests at different developmental stages. Glob. Chang. Biol. 2010, 7, 755–777. [Google Scholar] [CrossRef]
- Schulze, E.-D.; Lloyd, J.; Kelliher, F.M.; Wirth, C.; Rebmann, C.; Lühker, B.; Mund, M.; Knohl, A.; Milyukova, I.M.; Schulze, W.; et al. Productivity of forests in the Eurosiberian boreal region and their potential to act as a carbon sink—A synthesis. Glob. Chang. Boil. 1999, 5, 703–722. [Google Scholar] [CrossRef]
- Machimura, T.; Kobayashi, Y.; Lwahana, G.; Hirano, T.; Lopez, L.; Fukuda, M.; Fedorov, A.N. Change of Carbon Dioxide Budget during Three Years after Deforestation in Eastern Siberian Larch Forest. J. Agric. Meteorol. 2005, 60, 653–656. [Google Scholar] [CrossRef]
- Zaehle, S.; Bondeau, A.; Carter, T.R.; Cramer, W.; Erhard, M.; Prentice, I.C.; Reginster, I.; Rounsevell, M.D.A.; Sitch, S.; Smith, B.; et al. Projected Changes in Terrestrial Carbon Storage in Europe under Climate and Land-use Change, 1990–2100. Ecosystems 2007, 10, 380–401. [Google Scholar] [CrossRef]
- Singh, J.S.; Singh, D.P. Reforestation: A potential approach to mitigate excess atmospheric CH4 build-up. Ecol. Manag. Restor. 2012, 13, 245–248. [Google Scholar] [CrossRef]
- Poeplau, C.; Don, A. Sensitivity of soil organic carbon stocks and fractions to different land-use changes across Europe. Geoderma 2013, 192, 189–201. [Google Scholar] [CrossRef]
- Geider, R.J.; Delucia, E.H.; Falkowski, P.G.; Finzi, A.C.; Grime, J.P.; Grace, J.; Kana, T.M.; Roche, J.L.; Long, S.P.; Osborne, B.A.; et al. Primary productivity of planet earth: Biological determinants and physical constraints in terrestrial and aquatic habitats. Glob. Chang. Biol. 2002, 7, 849–882. [Google Scholar] [CrossRef]
- Battle, M.; Bender, M.L.; Tans, P.P.; White, J.W.C.; Ellis, J.T.; Conway, T.; Francey, R.J. Global carbon sinks and their variability inferred from atmospheric O2 and δ13 C. Science 2000, 287, 2467–2470. [Google Scholar] [CrossRef]
- Schimel, D. Contribution of Increasing CO2 and Climate to Carbon Storage by Ecosystems in the United States. Science 2000, 287, 2004–2006. [Google Scholar] [CrossRef]
- Clark, K.L.; Gholz, H.L.; Castro, M.S. Carbon Dynamics Along a Chronosequence of Slash Pine Plantations in North Florida. Ecol. Appl. 2004, 14, 1154–1171. [Google Scholar] [CrossRef]
- Kowalski, S.; Sartore, M.; Burlett, R.; Berbigier, P.; Loustau, D. The annual carbon budget of a French pine forest (Pinus pinaster) following harvest. Glob. Chang. Biol. 2010, 9, 1051–1065. [Google Scholar] [CrossRef]
- chulze, E.D.; Wirth, C.; Heimann, M. Managing forests after Kyoto. Science 2000, 289, 2058–2059. [Google Scholar] [CrossRef]
- Cohen, W.B.; Harmon, M.E.; Wallin, D.O.; Fiorella, M. Two Decades of Carbon Flux from Forests of the Pacific Northwest. Bioscience 1996, 46, 836–844. [Google Scholar] [CrossRef] [Green Version]
- Laporte, M.; Duchesne, L.; Morrison, I. Effect of clearcutting, selection cutting, shelterwood cutting and microsites on soil surface CO2 efflux in a tolerant hardwood ecosystem of northern Ontario. Ecol. Manag. 2003, 174, 565–575. [Google Scholar] [CrossRef]
- Coutinho, R.P.; Urquiaga, S.; Boddey, R.M.; Alves, B.J.R.; Torres, A.Q.A.; Jantalia, C.P. Estoque de carbono e nitrogênio e emissão de N2O em diferentes usos do solo na Mata Atlântica. Pesqui. Agropecuária Bras. 2010, 45, 195–203. [Google Scholar] [CrossRef]
- Shi, B.; Gao, W.; Jin, G. Effects on rhizospheric and heterotrophic respiration of conversion from primary forest to secondary forest and plantations in northeast China. Eur. J. Soil Boil. 2015, 66, 11–18. [Google Scholar] [CrossRef]
- Sheng, H.; Yang, Y.; Yang, Z.; Chen, G.; Xie, J.; Guo, J.; Zou, S. The dynamic response of soil respiration to land-use changes in subtropical China. Glob. Chang. Boil. 2010, 16, 1107–1121. [Google Scholar] [CrossRef]
- Shi, Z.; Li, Y.; Wang, S.; Wang, G.; Ruan, H.; He, R.; Tang, Y.; Zhang, Z. Accelerated soil CO2 efflux after conversion from secondary oak forest to pine plantation in southeastern China. Ecol. Res. 2009, 24, 1257–1265. [Google Scholar] [CrossRef]
- Prescott, C. Effects of clearcutting and alternative silvicultural systems on rates of decomposition and nitrogen mineralization in a coastal montane coniferous forest. Ecol. Manag. 1997, 95, 253–260. [Google Scholar] [CrossRef]
- Ewel, K.C.; Cropper, W.P., Jr.; Gholz, H.L. Soil CO2 evolution in Florida slash pine plantations. I. Changes through time. Can. J. Res. 1987, 17, 325–329. [Google Scholar] [CrossRef]
- Kolari, P.; Pumpanen, J.; Rannik, Ü.; Ilvesniemi, H.; Hari, P.; Berninger, F. Carbon balance of different aged Scots pine forests in Southern Finland. Glob. Chang. Biol. 2010, 10, 1106–1119. [Google Scholar] [CrossRef]
- Litton, C.M.; Ryan, M.G.; Knight, D.H.; Stahl, P.D. Soil–Surface carbon dioxide efflux and microbial biomass in relation to tree density 13 years after a stand replacing fire in a lodgepole pine ecosystem. Glob. Chang. Biol. 2010, 9, 680–696. [Google Scholar] [CrossRef]
- Misson, L.; Tang, J.; Xu, M.; McKay, M.; Goldstein, A. Influences of recovery from clear-cut, climate variability, and thinning on the carbon balance of a young ponderosa pine plantation. Agric. Meteorol. 2005, 130, 207–222. [Google Scholar] [CrossRef]
- Veldkamp, E.; Purbopuspito, J.; Corre, M.D.; Brumme, R.; Murdiyarso, D. Land use change effects on trace gas fluxes in the forest margins of Central Sulawesi, Indonesia. J. Geophys. Res. Space Phys. 2008, 113. [Google Scholar] [CrossRef] [Green Version]
- Keller, M.; Reiners, W.A. Soil–atmosphere exchange in nitrous oxide, nitric oxide, and methane under secondary succession of pasture to forest in the Atlantic lowlands of Costa Rica. Glob. Biogeochem. Cycles 1994, 8, 399–409. [Google Scholar] [CrossRef]
- Hiltbrunner, D.; Zimmermann, S.; Karbin, S.; Hagedorn, F.; Niklaus, P.A. Increasing soil methane sink along a 120-year afforestation chronosequence is driven by soil moisture. Glob. Chang. Boil. 2012, 18, 3664–3671. [Google Scholar] [CrossRef]
- Priemé, A.; Christensen, S. Seasonal and spatial variation of methane oxidation in a Danish spruce forest. Soil Boil. Biochem. 1997, 29, 1165–1172. [Google Scholar] [CrossRef]
- Benanti, G.; Saunders, M.; Tobin, B.; Osborne, B. Contrasting impacts of afforestation on nitrous oxide and methane emissions. Agric. Meteorol. 2014, 198, 82–93. [Google Scholar] [CrossRef]
- Houghton, R.A. The annual net flux of carbon to the atmosphere from changes in land use 1850–1990. Tellus B Chem. Phys. Meteorol. 1999, 51, 298–313. [Google Scholar] [CrossRef]
- Wang, C.K.; Yang, J.Y.; Zhang, Q.Z. Soil respiration in six temperate forests in China. Glob. Chang. Biol. 2006, 12, 2103–2114. [Google Scholar] [CrossRef]
- Zhang, H.D.; You, W.Z.; Xing, Z.K.; Wei, W.J.; Zhao, G.; Yan, T.W. Soil respiration of natural secondary forest and larch plantation in Montane region of eastern Liaoning province. J. Soil Water Conserv. 2011, 25, 198–201. [Google Scholar]
- Li, Y.; Dong, S.; Liu, S.; Zhou, H.; Gao, Q.; Cao, G.; Wang, X.; Su, X.; Zhang, Y.; Tang, L.; et al. Seasonal changes of CO2, CH4 and N2O fluxes in different types of alpine grassland in the Qinghai-Tibetan Plateau of China. Soil Boil. Biochem. 2015, 80, 306–314. [Google Scholar] [CrossRef]
- Wang, X.; Wang, C.; Guo, Q.; Wang, J. Improving the CO2 storage measurements with a single profile system in a tall-dense-canopy temperate forest. Agric. Meteorol. 2016, 228, 327–338. [Google Scholar] [CrossRef]
- Wang, C.K. Biomass allometric equations for 10 co–occurring tree species in Chinese temperate forests. Ecol. Manag. 2006, 222, 9–16. [Google Scholar] [CrossRef]
- Chen, D.K.; Zhou, X.F.; Zhao, H.X.; Wang, Y.H.; Jing, Y.Y. Study on the Structure, Function and Succession of the Four Types in Natural Secondary Forest; Northeast Forestry University Press: Harbin, China, 1982. [Google Scholar]
- Dou, X.; Zhou, W.; Zhang, Q.; Cheng, X. Greenhouse gas (CO2, CH4, N2O) emissions from soils following afforestation in central China. Atmos. Environ. 2015, 126, 98–106. [Google Scholar] [CrossRef]
- Van Miegroet, H.; Nicholas, N.S.; Creed, I.F.; Barker, M. Variation in overstory nitrogen uptake in a small, high-elevation southern Appalachian spruce-fir watershed. Can. J. Res. 2002, 32, 1741–1752. [Google Scholar]
- Yarie, J. The Role of Understory Vegetation in the Nutrient Cycle of Forested Ecosystems in the Mountain Hemlock Biogeoclimatic Zone. Ecology 1980, 61, 1498–1514. [Google Scholar] [CrossRef] [Green Version]
- Van Miegroet, H.; Moore, P.; Tewksbury, C.; Nicholas, N. Carbon sources and sinks in high-elevation spruce–fir forests of the Southeastern US. Ecol. Manag. 2007, 238, 249–260. [Google Scholar] [CrossRef]
- Bhatia, A.; Sasmal, S.; Jain, N.; Pathak, H.; Kumar, R.; Singh, A. Mitigating nitrous oxide emission from soil under conventional and no-tillage in wheat using nitrification inhibitors. Agric. Ecosyst. Environ. 2010, 136, 247–253. [Google Scholar] [CrossRef]
- Jain, N.; Arora, P.; Tomer, R.; Mishra, S.V.; Bhatia, A.; Pathak, H.; Chakraborty, D.; Kumar, V.; Dubey, D.; Harit, R.; et al. Greenhouse gases emission from soils under major crops in Northwest India. Sci. Total Environ. 2016, 542, 551–561. [Google Scholar] [CrossRef]
- Mu, C.C.; Cheng, W.; Sun, X.X.; Wu, Y.X. Seasonal Variation of Emission Fluxes of CO2, N2O and CH4 from Larix gemlinii Swamps Soils in Xiaoxing’An Mountains of China. Sci. Silvae Sin. 2010, 46, 7–15. [Google Scholar]
- Carmo, J.B.D.; Neto, E.R.D.S.; Duarte-Neto, P.J.; Ometto, J.P.H.B.; Martinelli, L.A. Conversion of the coastal Atlantic forest to pasture: Consequences for the nitrogen cycle and soil greenhouse gas emissions. Agric. Ecosyst. Environ. 2012, 148, 37–43. [Google Scholar] [CrossRef]
- Mu, C.C.; Zhang, B.W.; Han, L.D.; Yu, L.L.; Gu, H. Short–term effects of fire disturbance on greanhouse gases emission from Betula platyphylla–forested wetland in Xiaoxing’an Mountains, Northeast China. Chin. J. Appl. Ecol. 2011, 22, 857–865. [Google Scholar]
- Tang, X.; Liu, S.; Zhou, G.; Zhang, D.; Zhou, C. Soil–atmospheric exchange of CO₂, CH₄, and N₂O in three subtropical forest ecosystems in southern China. Glob. Chang. Biol. 2006, 12, 546–560. [Google Scholar] [CrossRef]
- Geng, S.C.; Chen, Z.J.; Zhang, J.H.; Lou, X.; Wang, X.X.; Dai, G.H.; Han, S.J.; Yu, D.D. Soil methane fluxes of three forest types in Changbai Mountain of Northeast China. Chin. J. Ecol. 2013, 32, 1091–1096. [Google Scholar]
- Smith, K.A.; Ball, T.; Conen, F.; Dobbie, K.E.; Massheder, J.; Rey, A. Exchange of greenhouse gases between soil and atmosphere: Interactions of soil physical factors and biological processes. Eur. J. Soil Sci. 2003, 54, 779–791. [Google Scholar] [CrossRef]
- Tate, K.; Ross, D.; Saggar, S.; Hedley, C.; Dando, J.; Singh, B.; Lambie, S. Methane uptake in soils from Pinus radiata plantations, a reverting shrubland and adjacent pastures: Effects of land-use change, and soil texture, water and mineral nitrogen. Soil Boil. Biochem. 2007, 39, 1437–1449. [Google Scholar] [CrossRef]
- Kolb, S. The quest for atmospheric methane oxidizers in forest soils. Environ. Microbiol. Rep. 2009, 1, 336–346. [Google Scholar] [CrossRef]
- Maurer, D.; Kolb, S.; Haumaier, L.; Borken, W. Inhibition of atmospheric methane oxidation by monoterpenes in Norway spruce and European beech soils. Soil Boil. Biochem. 2008, 40, 3014–3020. [Google Scholar] [CrossRef]
- Konda, R.; Ohta, S.; Ishizuka, S.; Heriyanto, J.; Wicaksono, A. Seasonal changes in the spatial structures of N2O, CO2, and CH4 fluxes from Acacia mangium plantation soils in Indonesia. Soil Boil. Biochem. 2010, 42, 1512–1522. [Google Scholar] [CrossRef]
- Tate, K.R. Soil methane oxidation and land-use change—From process to mitigation. Soil Boil. Biochem. 2015, 80, 260–272. [Google Scholar] [CrossRef]
- Wu, X.; Yao, Z.; Brüggemann, N.; Shen, Z.; Wolf, B.; Dannenmann, M.; Zheng, X.; Butterbach-Bahl, K.; Shen, Z. Effects of soil moisture and temperature on CO2 and CH4 soil–atmosphere exchange of various land use/cover types in a semi-arid grassland in Inner Mongolia, China. Soil Boil. Biochem. 2010, 42, 773–787. [Google Scholar] [CrossRef]
- Nesbit, S.; Breitenbeck, G. A laboratory study of factors influencing methane uptake by soils. Agric. Ecosyst. Environ. 1992, 41, 39–54. [Google Scholar] [CrossRef]
- Melillo, J.M.; Aber, J.D.; Castro, M.S.; Steudler, P.A.; Bowden, R.D. Factors controlling atmospheric methane consumption by temperate forest soils. Glob. Biogeochem. Cycles 1995, 9, 1–10. [Google Scholar]
- Wang, X.; Jiang, Y.; Jia, B.; Wang, F.; Zhou, G. Comparison of soil respiration among three temperate forests in Changbai Mountains, China. Can. J. Res. 2010, 40, 788–795. [Google Scholar] [CrossRef]
- Zhang, P.; Tian, X.; He, X.; Song, F.; Ren, L.; Jiang, P. Effect of litter quality on its decomposition in broadleaf and coniferous forest. Eur. J. Soil Boil. 2008, 44, 392–399. [Google Scholar] [CrossRef]
- Imaji, A.; Seiwa, K. Carbon allocation to defense, storage, and growth in seedlings of two temperate broad–leaved tree species. Oecologia 2010, 162, 273–281. [Google Scholar] [CrossRef]
- Vesterdal, L.; Elberling, B.; Christiansen, J.R.; Callesen, I.; Schmidt, I.K. Soil respiration and rates of soil carbon turnover differ among six common European tree species. Ecol. Manag. 2012, 264, 185–196. [Google Scholar] [CrossRef]
- Christiansen, J.R.; Vesterdal, L.; Callesen, I.; Elberling, B.; Schmidt, I.K.; Gundersen, P. Role of six European tree species and land–use legacy for nitrogen and water budgets in forests. Glob. Chang. Biol. 2010, 16, 2224–2240. [Google Scholar] [CrossRef]
- Liu, H.; Zhao, P.; Lu, P.; Wang, Y.-S.; Lin, Y.-B.; Rao, X.-Q. Greenhouse gas fluxes from soils of different land-use types in a hilly area of South China. Agric. Ecosyst. Environ. 2008, 124, 125–135. [Google Scholar] [CrossRef]
- Sun, X. A Study on Forest Soil CO2, N2O and CH4 Emission Flux and Dynamics in Xishan Area, Beijing; Beijing Forestry University Press: Beijing, China, 1999. [Google Scholar]
- Dong, Y.; Zhang, S.; Qi, Y.; Chen, Z.; Geng, Y. Fluxes of CO2, N2O and CH4 from a typical temperate grassland in Inner Mongolia and its daily variation. Chin. Sci. Bull. 2000, 45, 1590–1594. [Google Scholar] [CrossRef]
- Fest, B.J.; Hinko-Najera, N.; Wardlaw, T.; Griffith, D.W.T.; Livesley, S.J.; Arndt, S.K. Soil methane oxidation in both dry and wet temperate eucalypt forests shows a near-identical relationship with soil air-filled porosity. Biogeosciences 2017, 14, 467–479. [Google Scholar] [CrossRef]
- Davidson, E.A.; Verchot, L.V.; Cattânio, J.H.; Ackerman, I.L.; Carvalho, J. Effects of soil water content on soil respiration in forests and cattle pastures of eastern Amazonia. Biogeochemistry 2000, 48, 53–69. [Google Scholar] [CrossRef]
- Whalen, S.; Reeburgh, W. Effect of nitrogen fertilization on atmospheric methane oxidation in boreal forest soils. Chemosphere—Glob. Chang. Sci. 2000, 2, 151–155. [Google Scholar] [CrossRef] [Green Version]
- Reay, D.S.; Nedwell, D.B. Methane oxidation in temperate soils: Effects of inorganic N. Soil Boil. Biochem. 2004, 36, 2059–2065. [Google Scholar] [CrossRef]
- Visscher, A.D.; Cleemput, O.V. Induction of enhanced CH4 oxidation in soils: NH4+ inhibition patterns. Soil Biol. Biochem. 2003, 35, 907–913. [Google Scholar] [CrossRef]
- Bradford, M.; Wookey, P.; Ineson, P.; Lappin-Scott, H.; Bradford, M. Controlling factors and effects of chronic nitrogen and sulphur deposition on methane oxidation in a temperate forest soil. Soil Boil. Biochem. 2001, 33, 93–102. [Google Scholar] [CrossRef]
- Sun, X.; Mu, C.; Shi, L.; Cheng, W.; Liu, X.; Wu, Y.; Feng, D. Methane emission from forested swamps in Xiaoxing’an Mountains, Northeastern China. Chin. J. Plant Ecol. 2009, 33, 535–545. [Google Scholar]
- Amundson, R.; Niu, X.-F.; Wang, Y.; Niu, X. Seasonal and altitudinal variation in decomposition of soil organic matter inferred from radiocarbon measurements of soil CO2 flux. Glob. Biogeochem. Cycles 2000, 14, 199–211. [Google Scholar]
- Andrews, J.A.; Matamala, R.; Westover, K.M.; Schlesinger, W.H. Temperature effects on the diversity of soil heterotrophs and the δ13C of soil-respired CO2. Soil Boil. Biochem. 2000, 32, 699–706. [Google Scholar] [CrossRef]
- Dong, Y.J.; Cai, M.; Zhou, J.B. Effects of moisture and carbonate additions on CO2 emission from calcareous soil during closed–jar incubation. J. Arid Land 2014, 6, 37–43. [Google Scholar] [CrossRef]
- Oertel, C.; Matschullat, J.; Zurba, K.; Zimmermann, F.; Erasmi, S. Greenhouse gas emissions from soils—A review. Geochemistry 2016, 76, 327–352. [Google Scholar] [CrossRef]
- Tian, Y.; Haibara, K.; Toda, H.; Ding, F.; Liu, Y.; Choi, D. Microbial biomass and activity along a natural pH gradient in forest soils in a karst region of the upper Yangtze River, China. J. Res. 2008, 13, 205–214. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, S.; He, T.; Liu, L.; Wu, J. Response of organic carbon mineralization and microbial community to leaf litter and nutrient additions in subtropical forest soils. Soil Boil. Biochem. 2014, 71, 13–20. [Google Scholar] [CrossRef]
- Weintraub, M.N.; Schimel, J.P. Interactions between Carbon and Nitrogen Mineralization and Soil Organic Matter Chemistry in Arctic Tundra Soils. Ecosystems 2003, 6, 129–143. [Google Scholar] [CrossRef]
- Cai, H.; Shi, B.; Geng, P.; Di, X.; Chang, S.X.; Wang, C.; Jin, G. Carbon storage, net primary production, and net ecosystem production in four major temperate forest types in northeastern China. Can. J. Res. 2016, 46, 143–151. [Google Scholar] [CrossRef]
- Gower, S.T.; Gholz, H.L.; Nakane, K.; Galdwin, V.C. Production and carbon allocation patterns of pine forests. Ecol. Bull. 1994, 43, 115–135. [Google Scholar]
- Gough, C.M.; Vogel, C.S.; Hardiman, B.; Curtis, P.S. Wood net primary production resilience in an unmanaged forest transitioning from early to middle succession. Ecol. Manag. 2010, 260, 36–41. [Google Scholar] [CrossRef]
- He, H.; Pan, Y.; Zhu, W.; Liu, X.; Zhang, Q.; Zhu, X. Measurement of terrestrial ecosystem service value in China. J. Appl. Ecol. 2005, 16, 16. [Google Scholar]
- Li, Y.P.; Ji, J.J. Simulations of carbon exchange between global terrestrial ecosystem and the atmosphere. Acta Geogr. Sin. 2001, 56, 379–389. [Google Scholar]
- Pregitzer, K.S.; Euskirchen, E.S. Carbon cycling and storage in world forests: Biome patterns related to forest age. Glob. Chang. Boil. 2004, 10, 2052–2077. [Google Scholar] [CrossRef]
- Wang, C.K.; Yang, J.Y. Rhizospheric and heterotrophic components of soil respiration in six Chinese temperate forests. Glob. Chang. Biol. 2010, 13, 123–131. [Google Scholar] [CrossRef]
- Hardiman, B.S.; Bohrer, G.; Gough, C.M.; Vogel, C.S.; Curtis, P.S. The role of canopy structural complexity in wood net primary production of a maturing northern deciduous forest. Ecology 2011, 92, 1818–1827. [Google Scholar] [CrossRef] [Green Version]
- Hynynen, J.; Niemistö, P.; Viheräaarnio, A.; Brunner, A.; Hein, S.; Velling, P. Silviculture of birch (Betula pendula Roth and Betula pubescens Ehrh.) in northern Europe. Forestry 2010, 83, 103–119. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Forest Type | Age (Years) | Main Species Composition b (Dominant Species) | Land-use History | Density (Trees ha−1) | BA (m2 ha−1) | DBH (cm) | |
|---|---|---|---|---|---|---|---|
| Mean | Range | ||||||
| OMF | ˃150 | (8), 9, 2, 3, 10, 11, 12, 13, 14 | Natural forest | 789 | 54.6 | 25.3 | 4.0–69.1 |
| KPP | 51 | (8), 6, 3, 9, 5, 7, 6 | Reforestation in 1965 | 2111 | 42.1 | 15.0 | 3.9–34.4 |
| HWF | 60 | (4), (3), 2, (7) | Natural regeneration after clearcutting in 1955 | 1578 | 26.2 | 10.1 | 1.6–50.3 |
| BPF | 65 | (6), 15, 12, 17, 18 | Natural regeneration after clearcutting in 1950 | 3022 | 25.4 | 7.4 | 1.4–34.8 |
| 65 | (6), (5), 7, 4, 3, 2 | Natural regeneration after clearcutting in 1950 | 1833 | 34.1 | 12.1 | 1.6–49.4 | |
| MDF | 65 | 2, 7, 4, 3, 5, 6 | Natural regeneration after clearcutting in 1950 | 2022 | 34.8 | 11.3 | 1.9–49.6 |
| MOF | 66 | (1), 2, 3, 4, 5, 6 | Natural regeneration after clearcutting in 1949 | 1511 | 36.6 | 13.7 | 2.0–55.4 |
| Properties | OMF | KPP | HWF | BPF | MDF | MOF | |
|---|---|---|---|---|---|---|---|
| Air T (°C) | 5.92 ± 0.01B | 5.63 ± 0.01B | 3.44 ± 0.16E | 4.79 ± 0.02C | 4.05 ± 0.21D | 4.93 ± 0.05C | 6.93 ± 0.05A |
| Soil T (°C) | 7.20 ± 0.01A | 4.42 ± 0.01E | 4.17 ± 0.06F | 5.24 ± 0.07D | 5.63 ± 0.04C | 5.17 ± 0.15D | 6.90 ± 0.11B |
| Soil moisture (%) | 35.45 ± 0.49A | 27.90 ± 0.12C | 34.42 ± 0.33A | 26.58 ± 0.32D | 30.84 ± 0.41B | 27.35 ± 0.56D | 23.96 ± 0.47E |
| Soil pH | 5.51 ± 0.04C | 6.02 ± 0.03A | 5.66 ± 0.16B | 5.43 ± 0.05BC | 5.47 ± 0.24BC | 5.53 ± 0.06B | 5.72 ± 0.05B |
| NO3-N (mg kg−1) | 5.42 ± 0.35B | 9.76 ± 0.19A | 2.58 ± 0.26C | 1.09 ± 0.13D | 2.22 ± 0.42C | 2.18 ± 0.35C | 1.98 ± 0.20C |
| NH4+-N (mg kg−1) | 6.46 ± 0.48B | 24.12 ± 2.32A | 4.97 ± 0.07B | 4.95 ± 0.66B | 4.60 ± 1.69B | 4.38 ± 0.75B | 4.32 ± 0.19B |
| Soil organic C (g kg−1) | 72.02 ± 7.25B | 55.26 ± 1.08CD | 96.37 ± 8.15A | 51.96 ± 3.32D | 61.77 ± 2.07BCD | 69.92 ± 4.52BC | 48.42 ± 2.37D |
| Observe Period | Vegetation Types | ||||||
|---|---|---|---|---|---|---|---|
| OMF | KPP | HWF | BPF | MDF | MOF | ||
| Spring | −0.072 ± 0.010Db | −0.051 ± 0.005BCb | −0.053 ± 0.003BCb | −0.033 ± 0.003Ab | −0.067 ± 0.003CDb | −0.043 ± 0.003ABa | −0.053 ± 0.003BCab |
| Summer | −0.062 ± 0.003Ab | −0.088 ± 0.008ABc | −0.093 ± 0.012ABCc | −0.093 ± 0.003ABCd | −0.100 ± 0.015BCc | −0.123 ± 0.009Cc | −0.100 ± 0.015BCc |
| Autumn | −0.052 ± 0.006ABb | −0.042 ± 0.003Ab | −0.070 ± 0.006BCb | −0.063 ± 0.003BCc | −0.080 ± 0.010Cbc | −0.080 ± 0.006Cb | −0.083 ± 0.007Cbc |
| Winter | −0.018 ± 0.003ABCa | −0.002 ± 0.001Aa | −0.030 ± 0.001BCa | −0.007 ± 0.003ABa | −0.007 ± 0.003ABa | −0.057 ± 0.009Da | −0.037 ± 0.019CDa |
| Gs | −0.079 ± 0.006Ab | −0.088 ± 0.003Ab | −0.098 ± 0.007Ab | −0.084 ± 0.004Ab | −0.096 ± 0.011Ab | −0.101 ± 0.006Ab | −0.092 ± 0.011Ab |
| Ngs | −0.031 ± 0.004Ba | −0.016 ± 0.001Aa | −0.035 ± 0.003Ba | −0.026 ± 0.001ABa | −0.038 ± 0.006Ba | −0.060 ± 0.005Ca | −0.054 ± 0.005Ca |
| Aa | −0.051 ± 0.003AB | −0.046 ± 0.002A | −0.061 ± 0.004BC | −0.050 ± 0.002AB | −0.062 ± 0.006BC | −0.077 ± 0.004D | −0.070 ± 0.006CD |
| Observe Period | Vegetation Types | ||||||
|---|---|---|---|---|---|---|---|
| OMF | KPP | HWF | BPF | MDF | MOF | ||
| Spring | 129.295 ± 14.521Bb | 137.162 ± 5.692Bb | 185.272 ± 17.789ABb | 224.517 ± 12.656Ab | 240.538 ± 33.223Ab | 215.354 ± 29.795Ab | 210.625 ± 14.015Ab |
| Summer | 344.022 ± 58.402Ea | 403.728 ± 8.646DEa | 417.525 ± 3.152CDEa | 505.714 ± 21.556BCa | 491.544 ± 22.390BCDa | 615.834 ± 23.961Aa | 513.435 ± 31.230Ba |
| Autumn | 83.378 ± 11.224Cbc | 92.808 ± 2.466BCc | 121.463 ± 5.368ABc | 146.722 ± 10.923Ac | 138.262 ± 16.974Ac | 153.552 ± 10.633Ab | 120.554 ± 4.170ABc |
| Winter | 5.003 ± 0.418Cc | 20.063 ± 0.816Bd | 21.297 ± 6.254Bd | 19.283 ± 2.307Bd | 32.750 ± 1.515Ad | 15.347 ± 2.192Bc | 24.763 ± 3.564ABd |
| Gs | 299.579 ± 47.196Ca | 326.703 ± 6.949Ca | 357.814 ± 6.843BCa | 435.952 ± 8.252ABa | 420.000 ± 21.039ABa | 492.280 ± 23.938Aa | 421.820 ± 26.689ABa |
| Ngs | 26.743 ± 2.048Db | 46.825 ± 1.261Cb | 63.944 ± 6.477BCb | 72.707 ± 2.705ABb | 87.042 ± 12.142Ab | 76.979 ± 8.340ABb | 71.291 ± 3.665ABb |
| Aa | 140.425 ± 20.629D | 163.441 ± 3.610CD | 186.387 ± 5.151BC | 224.057 ± 2.277AB | 225.773 ± 15.610A | 250.023 ± 14.472A | 217.343 ± 9.122AB |
| Soil Layer/cm | Sample Site | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| OMF | KPP | HWF | BPF | MDF | MOF | ||||
| CH4 | temperature | Air T | −0.632 ** | −0.791 ** | −0.525 ** | −0.629 ** | −0.722 ** | −0.375 * | −0.419 * |
| 0 | −0.612 ** | −0.855 ** | −0.692 ** | −0.732 ** | −0.777 ** | −0.590 ** | −0.465 ** | ||
| 5 | −0.588 ** | −0.840 ** | −0.688 ** | −0.725 ** | −0.772 ** | −0.592 ** | −0.512 ** | ||
| 10 | −0.564 ** | −0.822 ** | −0.685 ** | −0.730 ** | −0.780 ** | −0.611 ** | −0.543 ** | ||
| 15 | −0.557 ** | −0.817 ** | −0.707 ** | −0.727 ** | −0.779 ** | −0.630 ** | −0.561 ** | ||
| 20 | −0.550 ** | −0.803 ** | −0.705 ** | −0.727 ** | −0.777 ** | −0.651 ** | −0.582 ** | ||
| 25 | −0.497 ** | −0.789 ** | −0.691 ** | −0.725 ** | −0.774 ** | −0.662 ** | −0.584 ** | ||
| 30 | −0.532 ** | −0.777 ** | −0.705 ** | −0.726 ** | −0.778 ** | −0.667 ** | −0.592 ** | ||
| 40 | −0.496 ** | −0.747 ** | −0.284 ** | −0.723 ** | −0.774 ** | −0.682 ** | −0.616 ** | ||
| soil volume water content | 0 | 0.084 ns | 0.769 ** | −0.284 ns | −0.308 ns | −0.387 * | 0.077 ns | −0.349 ns | |
| 5 | 0.007 ns | 0.750 ** | −0.404 * | −0.456 ** | −0.470 ** | 0.100 ns | −0414 * | ||
| 10 | 0.056 ns | 0.766 ** | −0.402 * | −0.503 ** | −0.612 ** | 0.030 ns | −0.452 * | ||
| 15 | 0.048 ns | 0.831 ** | −0.598 ** | −0.679 ** | −0.738 ** | −0.544 ** | −0.513 ** | ||
| 20 | −0.191 ns | 0.789 ** | −0.601 ** | −0.706 ** | −0.763 ** | −0.548 ** | −0.521 ** | ||
| 25 | −0.284 ns | 0.782 ** | −0.618 ** | −0.705 ** | −0.766 ** | −0.558 ** | −0.529 ** | ||
| 30 | −0.200 ns | 0.689 ** | −0.616 ** | −0.712 ** | −0.776 ** | −0.549 ** | −0.515 ** | ||
| 40 | −0.167 ns | 0.622 ** | −0.596 ** | −0.705 ** | −0.762 ** | −0.536 ** | −0.502 ** | ||
| CO2 | temperature | Air T | 0.799 ** | 0.780 ** | 0.817 ** | 0.791 ** | 0.827 ** | 0.826 ** | 0.847 ** |
| 0 | 0.844 ** | 0.930 ** | 0.860 ** | 0.891 ** | 0.894 ** | 0.921 ** | 0.878 ** | ||
| 5 | 0.898 ** | 0.939 ** | 0.877 ** | 0.892 ** | 0.912 ** | 0.914 ** | 0.886 ** | ||
| 10 | 0.917 ** | 0.927 ** | 0.884 ** | 0.890 ** | 0.900 ** | 0.914 ** | 0.904 ** | ||
| 15 | 0.920 ** | 0.932 ** | 0.887 ** | 0.885 ** | 0.899 ** | 0.905 ** | 0.906 ** | ||
| 20 | 0.915 ** | 0.932 ** | 0.877** | 0.883 ** | 0.890 ** | 0.899 ** | 0.900 ** | ||
| 25 | 0.820 ** | 0.922 ** | 0.879 ** | 0.879 ** | 0.887 ** | 0.895 ** | 0.899 ** | ||
| 30 | 0.903 ** | 0.910 ** | 0.871 ** | 0.874 ** | 0.885 ** | 0.879 ** | 0.897 ** | ||
| 40 | 0.882 ** | 0.892 ** | 0.856 ** | 0.863 ** | 0.873 ** | 0.875 ** | 0.889 ** | ||
| soil volume water content | 0 | −0.269 ns | −0.321 ns | 0.318 ns | 0.395 * | 0.591 ** | 0.116ns | 0.724 ** | |
| 5 | −0.052 ns | −0.513 * | 0.175 ns | 0.445 * | 0.677 ** | 0.132 ns | 0.741 ** | ||
| 10 | −0.057 ns | −0.612 ** | 0.151 ns | 0.482 ** | 0.652 ** | 0.303 ns | 0.763 ** | ||
| 15 | −0.088 ns | −0.475 ns | −0.434 * | 0.685 ** | 0.761 ** | 0.905 ** | 0.826 ** | ||
| 20 | 0.004 ns | −0.479 ns | −0.443 * | 0.689 ** | 0.748 ** | 0.912 ** | 0.814 ** | ||
| 25 | 0.009 ns | −0.364 ns | −0.449 * | 0.706 ** | 0.734 ** | 0.912 ** | 0.811 ** | ||
| 30 | −0.222 ns | −0.352 ns | −0.434 * | 0.722 ** | 0.727 ** | 0.918 ** | 0.826 ** | ||
| 40 | −0.257 ns | −0.495 * | −0.421 * | 0.742 ** | 0.753 ** | 0.913 ** | 0.784 ** | ||
| Plot | AT | ST | SVWC | pH | NO3−-N | NH4+-N | SOC | Intercept | R2 | p |
|---|---|---|---|---|---|---|---|---|---|---|
| Models for soil CH4 | ||||||||||
| OMF | −0.056 + | −0.008 * | −0.005 ** | 0.587 * | 0.663 | <0.05 | ||||
| KPP | −0.002 * | 0.002 + | 0.004 * | 0.001 + | −0.129 ** | 0.842 | <0.01 | |||
| HWF | 0.003 * | −0.007 * | −0.045 * | 0.612 | <0.05 | |||||
| BPF | −0.004 ** | 0.002 * | 0.198 *** | −0.003 + | −0.117 * | 0.927 | <0.001 | |||
| −0.003 ** | −0.047 *** | 0.626 | <0.01 | |||||||
| MDF | 0.004 * | −0.007 * | −0.057 ** | 0.441 | <0.1 | |||||
| MOF | −0.160 + | 0.836 + | 0.330 | <0.1 | ||||||
| Models for soil CO2 | ||||||||||
| OMF | 12.920 *** | 15.382 * | 14.930 + | −512.687 * | 0.891 | <0.001 | ||||
| KPP | 21.477 *** | −6.017 * | 122.783+ | −2.275 + | −476.729 | 0.973 | <0.001 | |||
| HWF | 16.692 *** | 114.102 *** | 0.833 | <0.001 | ||||||
| BPF | 23.067 *** | 57.177 * | 0.895 | <0.001 | ||||||
| 25.094 *** | 5.426 + | −485.257 + | −5.748 + | 2928.966 * | 0.944 | <0.001 | ||||
| MDF | −8.155 + | 33.999 *** | 11.265 + | −10.888 * | 454.856 * | 0.936 | <0.001 | |||
| MOF | −19.018 * | 49.746 *** | −112.542 * | −5.259 + | 388.194 ** | 0.943 | <0.001 | |||
| u | Layer | Vegetation Types | ||||||
|---|---|---|---|---|---|---|---|---|
| OMF | KPP | HWF | BPF | MDF | MOF | |||
| Net Primary Productivity (NPP)/(t ha−1 a−1) | Arborous layer | 7.06 ± 1.17B | 11.44 ± 1.40A | 8.55 ± 0.51B | 7.98 ± 0.56B | 6.99 ± 0.70B | 8.81 ± 0.72B | 7.36 ± 0.27B |
| Shrubby layer | 0.08 ± 0.02AB | 0.04 ± 0.01B | 0.10 ± 0.01A | 0.07 ± 0.02AB | 0.08 ± 0.01AB | 0.10 ± 0.03A | 0.06 ± 0.01AB | |
| Herbaceous layer | 1.71 ± 0.98A | 0.36 ± 0.02A | 1.34 ± 0.33A | 1.63 ± 0.71A | 0.80 ± 0.09A | 0.64 ± 0.05A | 0.71 ± 0.09A | |
| Vegetation | 8.85 ± 0.51B | 11.84 ± 1.42A | 9.99 ± 0.49AB | 9.68 ± 0.82AB | 7.87 ± 0.62B | 9.55 ± 0.80AB | 8.13 ± 0.33B | |
| Annual NetC Sequestration (ANCS)/(t ha−1 a−1) | Arborous layer | 3.52 ± 0.58BC | 6.08 ± 0.75A | 4.57 ± 0.27B | 3.68 ± 0.25BC | 3.06 ± 0.31C | 4.42 ± 0.36BC | 3.50 ± 0.13BC |
| Shrubby layer | 0.03 ± 0.01AB | 0.02 ± 0.01B | 0.05 ± 0.01A | 0.03 ± 0.01AB | 0.03 ± 0.01AB | 0.05 ± 0.01A | 0.03 ± 0.01AB | |
| Herbaceous layer | 0.75 ± 0.43A | 0.16 ± 0.01A | 0.59 ± 0.15A | 0.68 ± 0.29A | 0.32 ± 0.04A | 0.30 ± 0.02A | 0.32 ± 0.04A | |
| Vegetation | 4.30 ± 0.27BCD | 6.26 ± 0.75A | 5.21 ± 0.25AB | 4.38 ± 0.35BCD | 3.41 ± 0.27D | 4.76 ± 0.40BC | 3.85 ± 0.16CD | |
| Item | Vegetation Types | ||||||
|---|---|---|---|---|---|---|---|
| OMF | KPP | HWF | BPF | MDF | MOF | ||
| Annual Net C Sequestration (ANCS)/(t ha−1 y−1) | 4.30 ± 0.27BCD | 6.26 ± 0.75A | 5.21 ± 0.25AB | 4.38 ± 0.35BCD | 3.41 ± 0.27D | 4.76 ± 0.40BC | 3.85 ± 0.16CD |
| Annual C Emission (ACE)/(t ha−1 y−1) | 3.15 ± 0.47D | 3.72 ± 0.08CD | 4.39 ± 0.12BC | 5.28 ± 0.05A | 5.32 ± 0.37A | 5.89 ± 0.34A | 5.12 ± 0.21AB |
| Carbon Source/Sink (CSS)/(t ha−1 y−1) | 1.15 ± 0.65AB | 2.54 ± 0.80A | 0.82 ± 0.15B | −0.89 ± 0.30C | −1.90 ± 0.64C | −1.12 ± 0.25C | −1.27 ± 0.37C |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, B.; Mu, C.; Zhao, J.; Zhou, X.; Zhang, J. Effects on Carbon Sources and Sinks from Conversion of Over-Mature Forest to Major Secondary Forests and Korean Pine Plantation in Northeast China. Sustainability 2019, 11, 4232. https://0-doi-org.brum.beds.ac.uk/10.3390/su11154232
Wu B, Mu C, Zhao J, Zhou X, Zhang J. Effects on Carbon Sources and Sinks from Conversion of Over-Mature Forest to Major Secondary Forests and Korean Pine Plantation in Northeast China. Sustainability. 2019; 11(15):4232. https://0-doi-org.brum.beds.ac.uk/10.3390/su11154232
Chicago/Turabian StyleWu, Bin, Changcheng Mu, Jiaqi Zhao, Xuejiao Zhou, and Junhui Zhang. 2019. "Effects on Carbon Sources and Sinks from Conversion of Over-Mature Forest to Major Secondary Forests and Korean Pine Plantation in Northeast China" Sustainability 11, no. 15: 4232. https://0-doi-org.brum.beds.ac.uk/10.3390/su11154232