Effects of Zeolite and Biochar Addition on Ammonia-Oxidizing Bacteria and Ammonia-Oxidizing Archaea Communities during Agricultural Waste Composting

Abstract
:1. Introduction
2. Materials and Methods
2.1. Raw Materials and Composting Setup
2.2. Samples Collection and Parameters Determination
2.3. AMO Enzyme Activity Measurement
2.4. Quantitative PCR
2.5. Data Analysis
3. Results
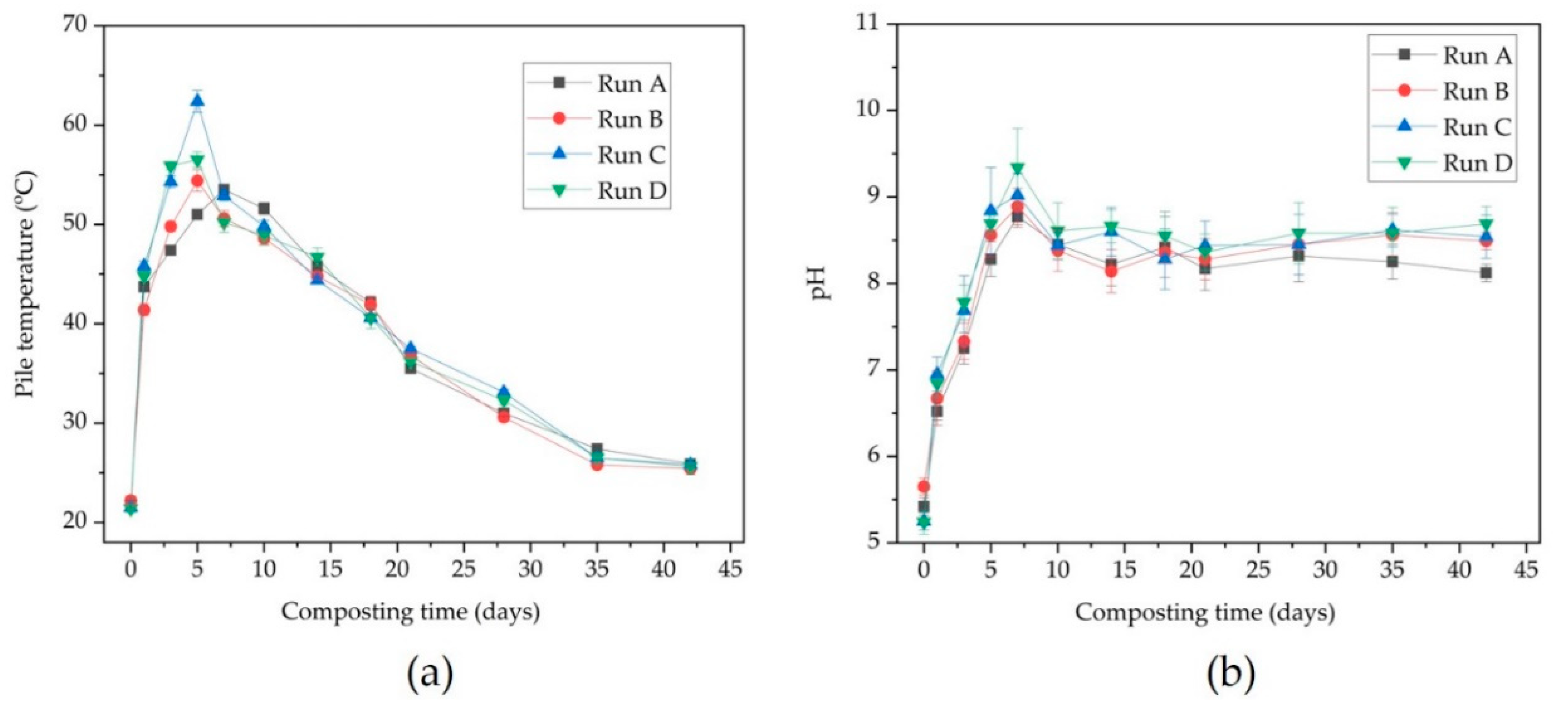
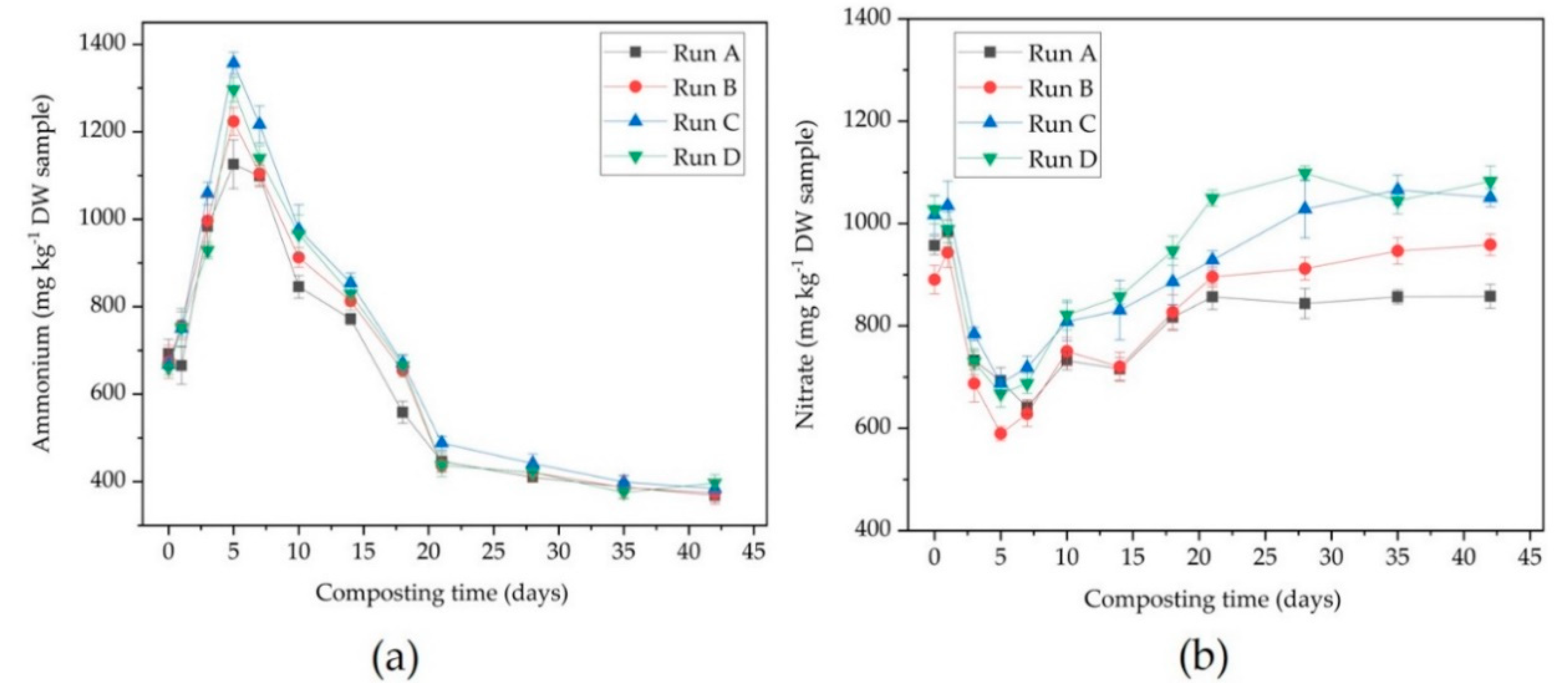
3.1. Physico-Chemical Parameters
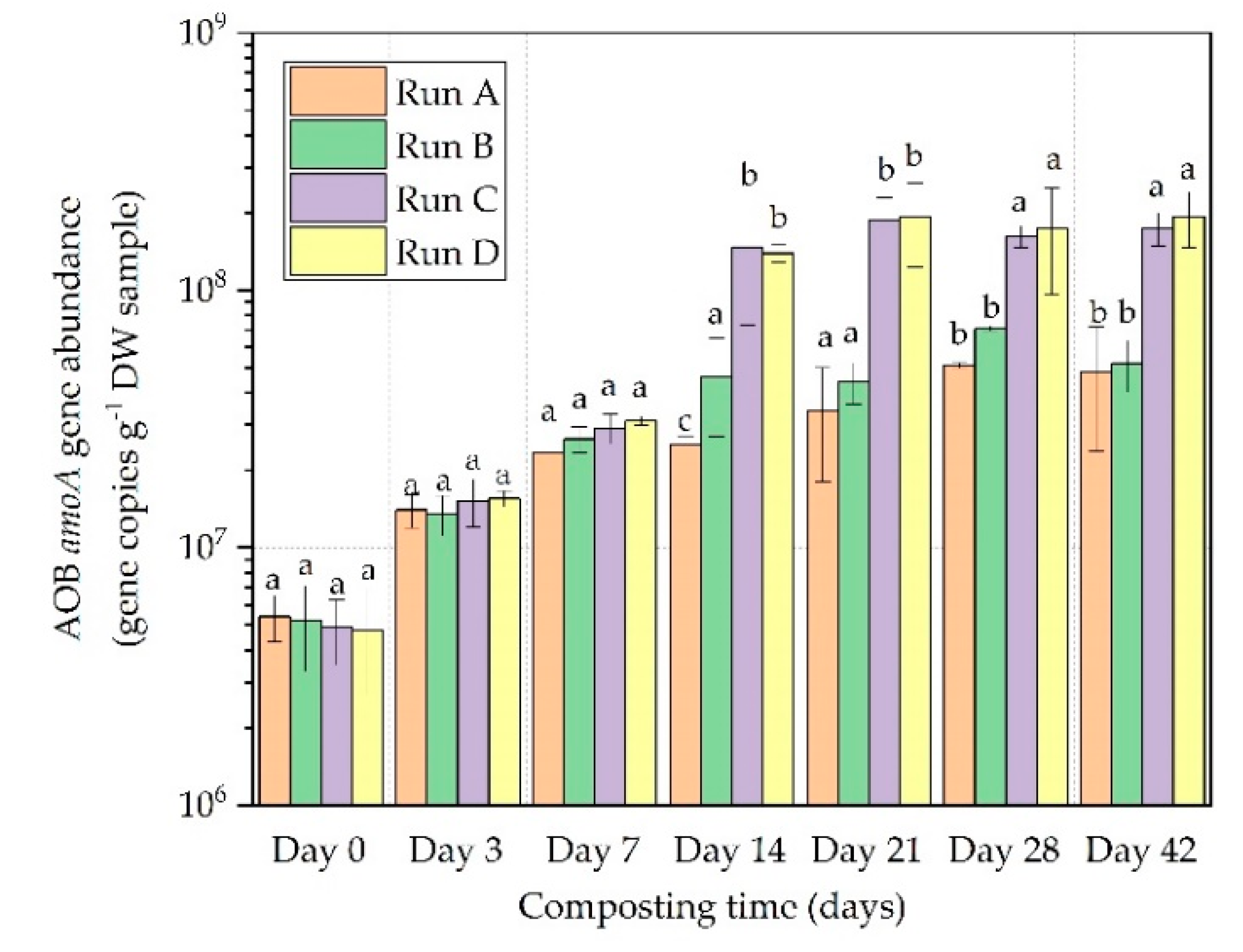
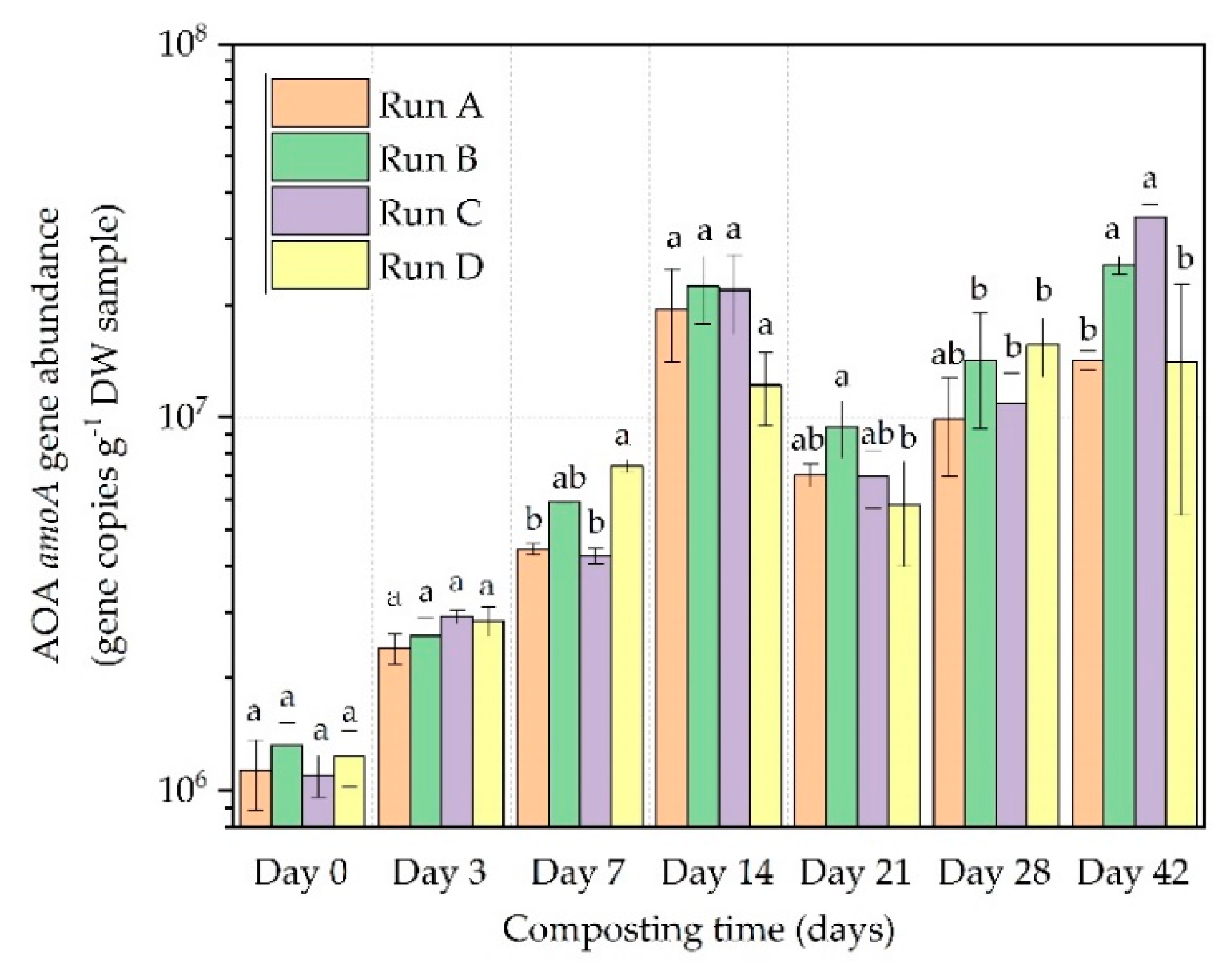
3.2. AOB and AOA amoA Gene Abundance
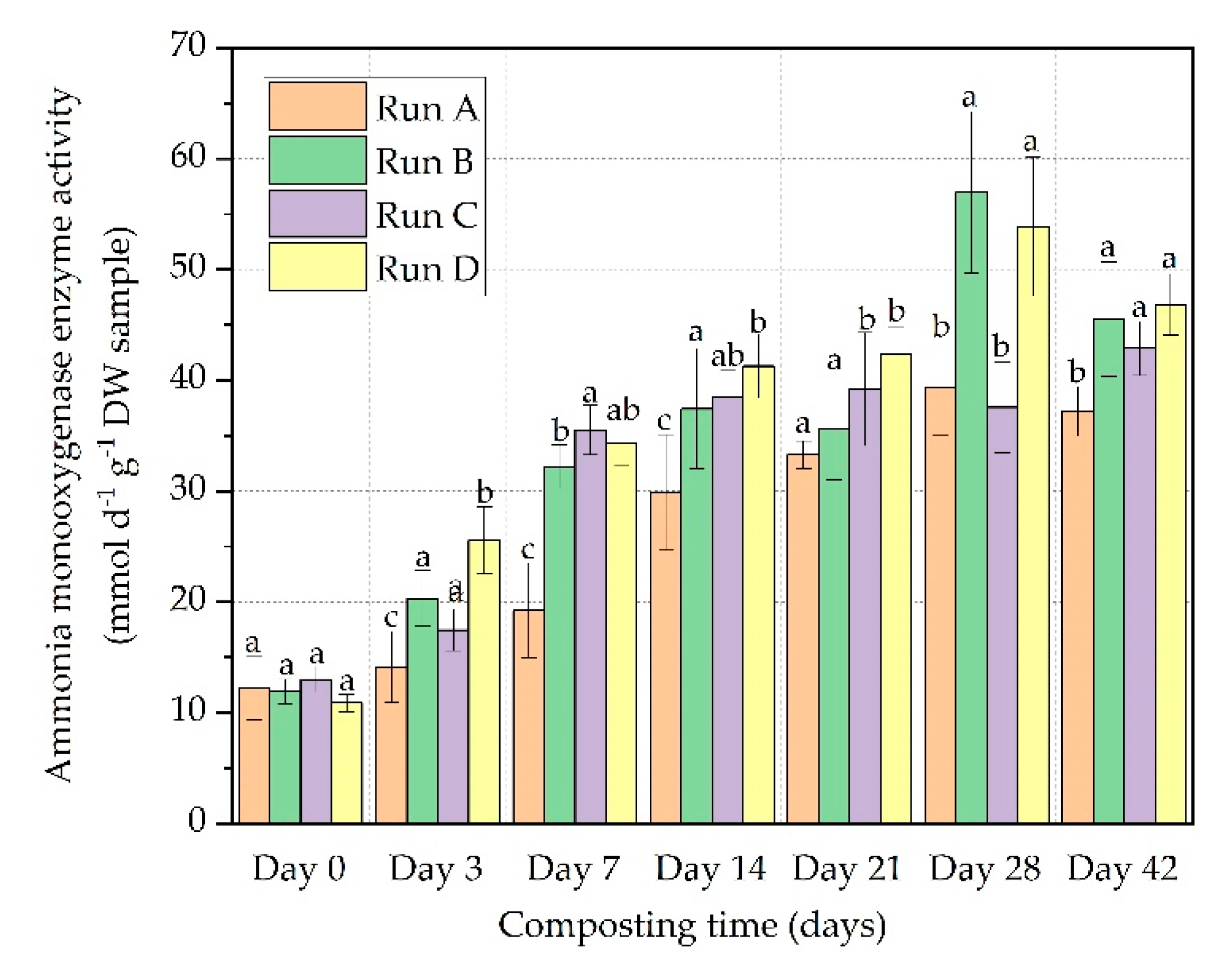
3.3. AMO Enzyme Activity
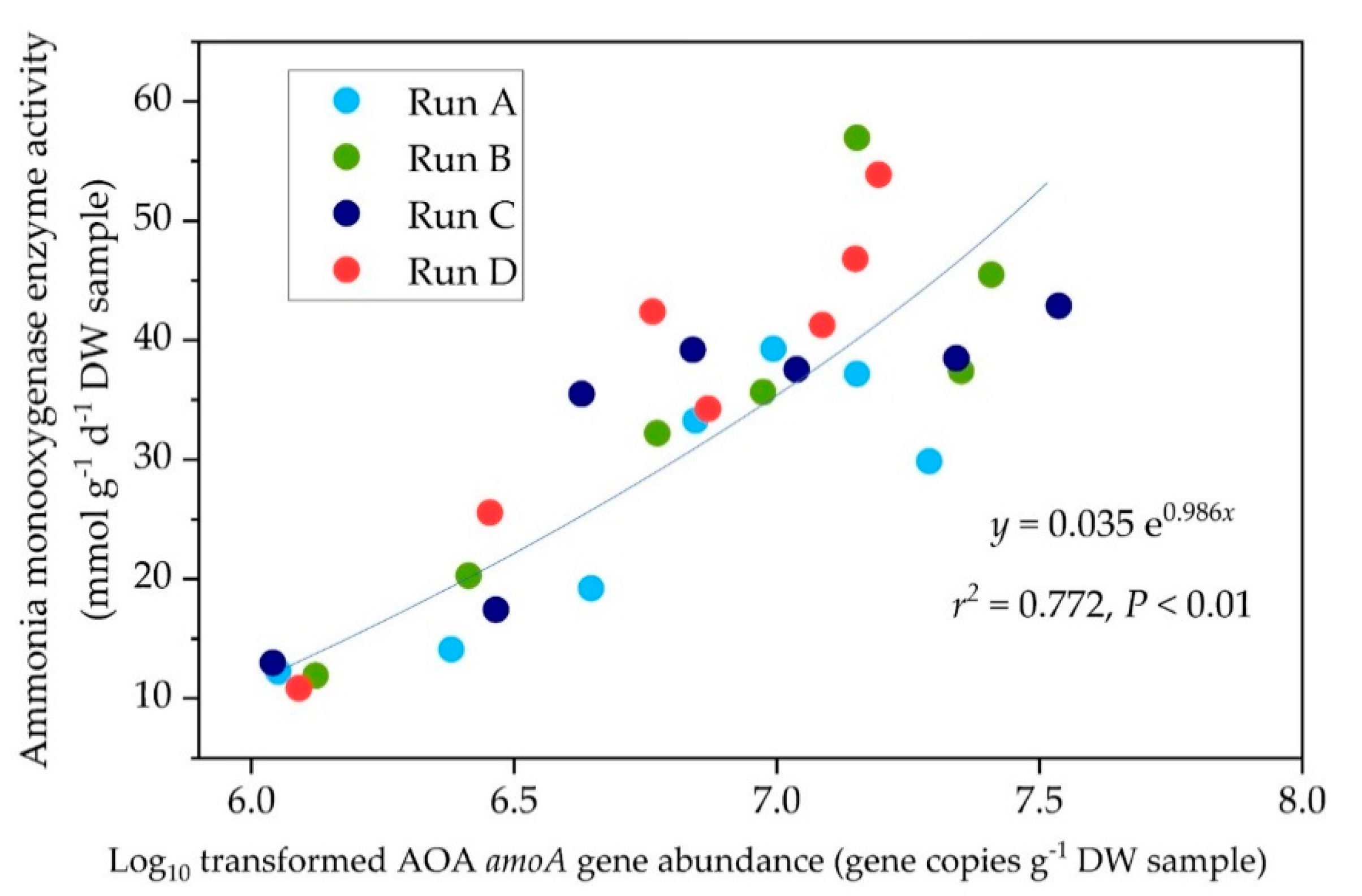
3.4. Relationship Between AMO Enzyme Activity and amoA Gene Abundance
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ren, L.; Cai, C.; Zhang, J.; Yang, Y.; Wu, G.; Luo, L.; Huang, H.; Zhou, Y.; Qin, P.; Yu, M. Key environmental factors to variation of ammonia-oxidizing archaea community and potential ammonia oxidation rate during agricultural waste composting. Bioresour. Technol. 2018, 270, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Zeng, G.; Cheng, M.; Huang, D.; Lai, C.; Xu, P.; Wei, Z.; Li, N.; Zhang, C.; He, X.; He, Y. Study of the degradation of methylene blue by semi-solid-state fermentation of agricultural residues with Phanerochaete chrysosporium and reutilization of fermented residues. Waste Manag. 2015, 38, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Bernal, M.; Alburquerque, J.; Moral, R. Composting of animal manures and chemical criteria for compost maturity assessment. A review. Bioresour. Technol. 2009, 100, 5444–5453. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, A.; Artola, A.; Font, X.; Gea, T.; Barrena, R.; Gabriel, D.; Sánchez-Monedero, M.Á.; Roig, A.; Cayuela, M.L.; Mondini, C. Greenhouse gas emissions from organic waste composting. Environ. Chem. Lett. 2015, 13, 223–238. [Google Scholar] [CrossRef] [Green Version]
- Guillermo, P.; Raúl, M.; Eduardo, A.; Agustín, D.P. Gaseous emissions from management of solid waste: A systematic review. Glob. Chang. Biol. 2015, 21, 1313–1327. [Google Scholar]
- Chen, R.; Wang, Y.; Wang, W.; Wei, S.; Jing, Z.; Lin, X. N2O emissions and nitrogen transformation during windrow composting of dairy manure. J. Environ. Manag. 2015, 160, 121–127. [Google Scholar] [CrossRef]
- Du, J.; Zhang, Y.; Qu, M.; Yin, Y.; Fan, K.; Hu, B.; Zhang, H.; Wei, M.; Ma, C. Effects of biochar on the microbial activity and community structure during sewage sludge composting. Bioresour. Technol. 2019, 272, 171–179. [Google Scholar] [CrossRef]
- Zhang, J.; Luo, L.; Gao, J.; Peng, Q.; Huang, H.; Chen, A.; Lu, L.; Yan, B.; Wong, J.W. Ammonia-oxidizing bacterial communities and shaping factors with different Phanerochaete chrysosporium inoculation regimes during agricultural waste composting. RSC Adv. 2016, 6, 61473–61481. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, Y.; Chen, Y.; Lu, Q.; Li, M.; Wang, X.; Wei, Y.; Xie, X.; Wei, Z. A regulating method for reducing nitrogen loss based on enriched ammonia-oxidizing bacteria during composting. Bioresour. Technol. 2016, 221, 276–283. [Google Scholar] [CrossRef]
- Awasthi, M.K.; Wang, M.; Pandey, A.; Chen, H.; Awasthi, S.K.; Wang, Q.; Ren, X.; Lahori, A.H.; Li, D.-S.; Li, R. Heterogeneity of zeolite combined with biochar properties as a function of sewage sludge composting and production of nutrient-rich compost. Waste Manag. 2017, 68, 760–773. [Google Scholar] [CrossRef]
- Jiang, T.; Ma, X.; Tang, Q.; Yang, J.; Li, G.; Schuchardt, F. Combined use of nitrification inhibitor and struvite crystallization to reduce the NH3 and N2O emissions during composting. Bioresour. Technol. 2016, 217, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.; Bustamante, M.; Tortosa, G.; Moral, R.; Bernal, M. Gaseous emissions and process development during composting of pig slurry: The influence of the proportion of cotton gin waste. J. Clean. Prod. 2016, 112, 81–90. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, Y.; Wang, H.; Zhao, X.; Cui, H.; Wei, Z. Reducing nitrogen loss and phytotoxicity during beer vinasse composting with biochar addition. Waste Manag. 2017, 61, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Awasthi, M.K.; Ren, X.; Zhao, J.; Li, R.; Wang, Z.; Wang, M.; Chen, H.; Zhang, Z. Combining biochar, zeolite and wood vinegar for composting of pig manure: The effect on greenhouse gas emission and nitrogen conservation. Waste Manag. 2018, 74, 221–230. [Google Scholar] [CrossRef]
- Sanchez-Garcia, M.; Alburquerque, J.A.; Sanchez-Monedero, M.A.; Roig, A.; Cayuela, M.L. Biochar accelerates organic matter degradation and enhances N mineralisation during composting of poultry manure without a relevant impact on gas emissions. Bioresour. Technol. 2015, 192, 272–279. [Google Scholar] [CrossRef]
- Chan, M.T.; Selvam, A.; Wong, J.W. Reducing nitrogen loss and salinity during ‘struvite’ food waste composting by zeolite amendment. Bioresour. Technol. 2016, 200, 838–844. [Google Scholar] [CrossRef]
- Awasthi, M.K.; Wang, Q.; Huang, H.; Ren, X.; Lahori, A.H.; Mahar, A.; Ali, A.; Shen, F.; Li, R.; Zhang, Z. Influence of zeolite and lime as additives on greenhouse gas emissions and maturity evolution during sewage sludge composting. Bioresour. Technol. 2016, 216, 172–181. [Google Scholar] [CrossRef]
- Awasthi, M.K.; Wang, Q.; Ren, X.; Zhao, J.; Huang, H.; Awasthi, S.K.; Lahori, A.H.; Li, R.; Zhou, L.; Zhang, Z. Role of biochar amendment in mitigation of nitrogen loss and greenhouse gas emission during sewage sludge composting. Bioresour. Technol. 2016, 219, 270–280. [Google Scholar] [CrossRef]
- Czekała, W.; Malińska, K.; Cáceres, R.; Janczak, D.; Dach, J.; Lewicki, A. Co-composting of poultry manure mixtures amended with biochar–The effect of biochar on temperature and C-CO2 emission. Bioresour. Technol. 2016, 200, 921–927. [Google Scholar] [CrossRef]
- Jindo, K.; Suto, K.; Matsumoto, K.; García, C.; Sonoki, T.; Sanchez-Monedero, M. Chemical and biochemical characterisation of biochar-blended composts prepared from poultry manure. Bioresour. Technol. 2012, 110, 396–404. [Google Scholar] [CrossRef]
- Zhou, G.; Xu, X.; Qiu, X.; Zhang, J. Biochar influences the succession of microbial communities and the metabolic functions during rice straw composting with pig manure. Bioresour. Technol. 2019, 272, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Cáceres, R.; Malińska, K.; Marfà, O. Nitrification within composting: A review. Waste Manag. 2018, 72, 119–137. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zeng, G.; Chen, Y.; Liang, J.; Zhang, C.; Huang, B.; Sun, W.; Chen, M.; Yu, M.; Huang, H. Phanerochaete chrysosporium inoculation shapes the indigenous fungal communities during agricultural waste composting. Biodegradation 2014, 25, 669–680. [Google Scholar] [CrossRef]
- Zhang, J.; Zeng, G.; Chen, Y.; Yu, M.; Huang, H.; Fan, C.; Zhu, Y.; Li, H.; Liu, Z.; Chen, M. Impact of Phanerochaete chrysosporium inoculation on indigenous bacterial communities during agricultural waste composting. Appl. Microbiol. Biotechnol. 2013, 97, 3159–3169. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.E.; Giguere, A.T.; Zoebelein, C.M.; Myrold, D.D.; Bottomley, P.J. Modeling of soil nitrification responses to temperature reveals thermodynamic differences between ammonia-oxidizing activity of archaea and bacteria. ISME J. 2017, 11, 896–908. [Google Scholar] [CrossRef]
- Zeng, G.; Zhang, J.; Chen, Y.; Yu, Z.; Yu, M.; Li, H.; Liu, Z.; Chen, M.; Lu, L.; Hu, C. Relative contributions of archaea and bacteria to microbial ammonia oxidation differ under different conditions during agricultural waste composting. Bioresour. Technol. 2011, 102, 9026–9032. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, Z.; Awasthi, M.K.; Jiang, Y.; Li, R.; Ren, X.; Zhao, J.; Shen, F.; Wang, M.; Zhang, Z. Evaluation of medical stone amendment for the reduction of nitrogen loss and bioavailability of heavy metals during pig manure composting. Bioresour. Technol. 2016, 220, 297–304. [Google Scholar] [CrossRef]
- Wang, C.; Lu, H.; Dong, D.; Deng, H.; Strong, P.; Wang, H.; Wu, W. Insight into the effects of biochar on manure composting: Evidence supporting the relationship between N2O emission and denitrifying community. Environ. Sci. Technol. 2013, 47, 7341–7349. [Google Scholar] [CrossRef]
- Villasenor, J.; Rodriguez, L.; Fernandez, F. Composting domestic sewage sludge with natural zeolites in a rotary drum reactor. Bioresour. Technol. 2011, 102, 1447–1454. [Google Scholar] [CrossRef]
- Zhang, J.; Sui, Q.; Li, K.; Chen, M.; Tong, J.; Qi, L.; Wei, Y. Influence of natural zeolite and nitrification inhibitor on organics degradation and nitrogen transformation during sludge composting. Environ. Sci. Pollut. Res. 2015, 23, 1324–1334. [Google Scholar] [CrossRef]
- Li, R.; Wang, Q.; Zhang, Z.; Zhang, G.; Li, Z.; Wang, L.; Zheng, J. Nutrient transformation during aerobic composting of pig manure with biochar prepared at different temperatures. Environ. Technol. 2015, 36, 815–826. [Google Scholar] [CrossRef] [PubMed]
- Venglovsky, J.; Sasakova, N.; Vargova, M.; Pacajova, Z.; Placha, I.; Petrovsky, M.; Harichova, D. Evolution of temperature and chemical parameters during composting of the pig slurry solid fraction amended with natural zeolite. Bioresour. Technol. 2005, 96, 181–189. [Google Scholar] [CrossRef]
- Kithome, M.; Paul, J.; Bomke, A. Reducing nitrogen losses during simulated composting of poultry manure using adsorbents or chemical amendments. J. Environ. Qual. 1999, 28, 194–201. [Google Scholar] [CrossRef]
- Steiner, C.; Das, K.C.; Melear, N.; Lakly, D. Reducing nitrogen loss during poultry litter composting using biochar. J. Environ. Qual. 2010, 39, 1236–1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malińska, K.; Zabochnicka-Świątek, M.; Dach, J. Effects of biochar amendment on ammonia emission during composting of sewage sludge. Ecol. Eng. 2014, 71, 474–478. [Google Scholar] [CrossRef]
- Enwall, K.; Nyberg, K.; Bertilsson, S.; Cederlund, H.; Stenström, J.; Hallin, S. Long-term impact of fertilization on activity and composition of bacterial communities and metabolic guilds in agricultural soil. Soil Biol. Biochem. 2007, 39, 106–115. [Google Scholar] [CrossRef]
- Mendum, T.; Hirsch, P. Changes in the population structure of β-group autotrophic ammonia oxidising bacteria in arable soils in response to agricultural practice. Soil Biol. Biochem. 2002, 34, 1479–1485. [Google Scholar] [CrossRef]
- Yao, H.; Gao, Y.; Nicol, G.W.; Campbell, C.D.; Prosser, J.I.; Zhang, L.; Han, W.; Singh, B.K. Links between ammonia oxidizer community structure, abundance, and nitrification potential in acidic soils. Appl. Environ. Microbiol. 2011, 77, 4618–4625. [Google Scholar] [CrossRef] [Green Version]
- Clough, T.; Condron, L.; Kammann, C.; Müller, C. A review of biochar and soil nitrogen dynamics. Agronomy 2013, 3, 275–293. [Google Scholar] [CrossRef] [Green Version]
- Agyarko-Mintah, E.; Cowie, A.; Singh, B.P.; Joseph, S.; Van Zwieten, L.; Cowie, A.; Harden, S.; Smillie, R. Biochar increases nitrogen retention and lowers greenhouse gas emissions when added to composting poultry litter. Waste Manag. 2017, 61, 138–149. [Google Scholar] [CrossRef]
- Sun, D.; Lan, Y.; Xu, E.G.; Meng, J.; Chen, W. Biochar as a novel niche for culturing microbial communities in composting. Waste Manag. 2016, 54, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, N.; Otawa, K.; Nakai, Y. Diversity and abundance of ammonia-oxidizing bacteria and ammonia-oxidizing archaea during cattle manure composting. Microb. Ecol. 2010, 60, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Nozomi, Y.; Ryu, O.; Yoshihisa, S.; Chika, T.; Yutaka, N. Ammonia-oxidizing bacteria rather than ammonia-oxidizing archaea were widely distributed in animal manure composts from field-scale facilities. Microbes Environ. 2012, 27, 519–524. [Google Scholar]
- Tang, J.; Zhang, J.; Ren, L.; Zhou, Y.; Gao, J.; Luo, L.; Yang, Y.; Peng, Q.; Huang, H.; Chen, A. Diagnosis of soil contamination using microbiological indices: A review on heavy metal pollution. J. Environ. Manag. 2019, 242, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Feike, J.; Jurgens, K.; Hollibaugh, J.T.; Kruger, S.; Jost, G.; Labrenz, M. Measuring unbiased metatranscriptomics in suboxic waters of the central Baltic Sea using a new in situ fixation system. ISME J. 2012, 6, 461–470. [Google Scholar] [CrossRef]
- Kuypers, M.M.M.; Marchant, H.K.; Kartal, B. The microbial nitrogen-cycling network. Nat. Rev. Microbiol. 2018, 16, 263–276. [Google Scholar] [CrossRef]
- Urakawa, H.; Rajan, S.; Feeney, M.E.; Sobecky, P.A.; Mortazavi, B. Ecological response of nitrification to oil spills and its impact on the nitrogen cycle. Environ. Microbiol. 2019, 21, 18–33. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Xu, Y.; He, T.; Jia, H.; Feng, M.; Xiang, S.; Wang, S.; Ni, J.; Xie, D.; Li, Z. Ammonium transformed into nitrous oxide via nitric oxide by Pseudomonas putida Y-9 under aerobic conditions without hydroxylamine as intermediate. Bioresour. Technol. 2019, 277, 87–93. [Google Scholar] [CrossRef]
- Madrini, B.; Shibusawa, S.; Kojima, Y.; Hosaka, S. Effect of natural zeolite (clinoptilolite) on ammonia emissions of leftover food-rice hulls composting at the initial stage of the thermophilic process. J. Agric. Meteorol. 2016, 72, 12–19. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.-S.; Lee, D.-S.; Kwak, J.-H.; Park, H.-J.; Kim, H.-Y.; Choi, W.-J. Fly ash and zeolite amendments increase soil nutrient retention but decrease paddy rice growth in a low fertility soil. J. Soils Sediment 2016, 16, 756–766. [Google Scholar] [CrossRef]
- Koenig, R.T.; Palmer, M.D.; Miner, F.D., Jr.; Miller, B.E.; Harrison, J.D. Chemical amendments and process controls to reduce ammonia volatilization during in-house composting. Compost Sci. Util. 2005, 13, 141–149. [Google Scholar] [CrossRef]
- Martens-Habbena, W.; Stahl, D.A. Nitrogen metabolism and kinetics of ammonia-oxidizing archaea. Methods Enzymol. 2011, 496, 465–487. [Google Scholar] [PubMed]
- Verhamme, D.T.; Prosser, J.I.; Nicol, G.W. Ammonia concentration determines differential growth of ammonia-oxidising archaea and bacteria in soil microcosms. ISME J. 2011, 5, 1067–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di, H.J.; Cameron, K.C.; Shen, J.P.; Winefield, C.S.; O’Callaghan, M.; Bowatte, S.; He, J.Z. Ammonia-oxidizing bacteria and archaea grow under contrasting soil nitrogen conditions. FEMS Microbiol. Ecol. 2010, 72, 386–394. [Google Scholar] [CrossRef] [Green Version]
- Hatzenpichler, R.; Lebedeva, E.V.; Spieck, E.; Stoecker, K.; Richter, A.; Daims, H.; Wagner, M. A moderately thermophilic ammonia-oxidizing crenarchaeote from a hot spring. Proc. Natl. Acad. Sci. USA 2008, 105, 2134–2139. [Google Scholar] [CrossRef] [Green Version]
- De Gannes, V.; Eudoxie, G.; Dyer, D.H.; Hickey, W.J. Diversity and abundance of ammonia oxidizing archaea in tropical compost systems. Front. Microbiol. 2012, 3, 224. [Google Scholar] [CrossRef] [Green Version]
- Ding, L.J.; An, X.L.; Li, S.; Zhang, G.L.; Zhu, Y.G. Nitrogen loss through anaerobic ammonium oxidation coupled to iron reduction from paddy soils in a chronosequence. Environ. Sci. Technol. 2014, 48, 10641–10647. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Raw materials or Different Piles | TOC (%) | TN (%) | C/N ratio | Moisture (%) | pH | Ammonium (mg/kg) | Nitrate (mg/kg) |
|---|---|---|---|---|---|---|---|
| Rice straw | 436.8 | 8.7 | 50.2 | 7.9 | - | - | - |
| Vegetable | 112.5 | 4.8 | 23.4 | 79.6 | 7.2 | - | - |
| Soil | 62.4 | 2.3 | 22.8 | 15.8 | 5.8 | - | - |
| Bran | 474.8 | 44.5 | 10.7 | 12.5 | - | - | - |
| Zeolite | - | - | - | 1.2 | - | - | - |
| Biochar | 50.0 | - | - | 7.0 | 9.08 | 38.2 | 7.0 |
| Run A | 272.5 | 8.3 | 31.7 | 62.3 | 5.42 | 692.8 | 957.2 |
| Run B | 251.6 | 8.5 | 29.6 | 59.8 | 5.65 | 673.5 | 890.5 |
| Run C | 275.5 | 8.4 | 32.8 | 58.6 | 5.25 | 668.2 | 1016.8 |
| Run D | 259.6 | 8.7 | 30.9 | 58.2 | 5.24 | 659.8 | 1027.2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, X.; Ren, L.; Zhang, J.; Peng, H. Effects of Zeolite and Biochar Addition on Ammonia-Oxidizing Bacteria and Ammonia-Oxidizing Archaea Communities during Agricultural Waste Composting. Sustainability 2020, 12, 6336. https://0-doi-org.brum.beds.ac.uk/10.3390/su12166336
Wu X, Ren L, Zhang J, Peng H. Effects of Zeolite and Biochar Addition on Ammonia-Oxidizing Bacteria and Ammonia-Oxidizing Archaea Communities during Agricultural Waste Composting. Sustainability. 2020; 12(16):6336. https://0-doi-org.brum.beds.ac.uk/10.3390/su12166336
Chicago/Turabian StyleWu, Xin, Liheng Ren, Jiachao Zhang, and Hui Peng. 2020. "Effects of Zeolite and Biochar Addition on Ammonia-Oxidizing Bacteria and Ammonia-Oxidizing Archaea Communities during Agricultural Waste Composting" Sustainability 12, no. 16: 6336. https://0-doi-org.brum.beds.ac.uk/10.3390/su12166336