Improving ‘Lipid Productivity’ in Microalgae by Bilateral Enhancement of Biomass and Lipid Contents: A Review

 ,
,  , ,
, ,  ,
,  , and
, and
Abstract
:1. Introduction
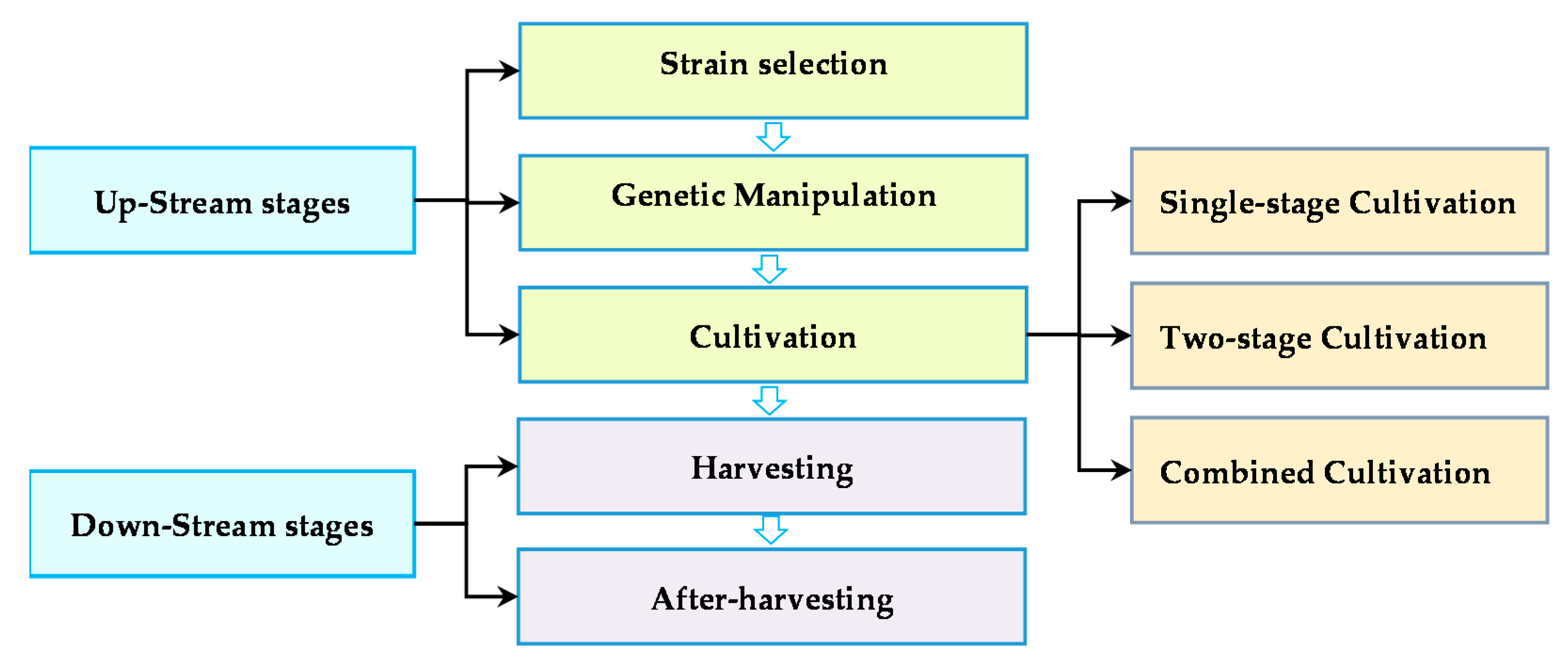
2. Bilateral Improvement in Biomass and Lipid Productivities
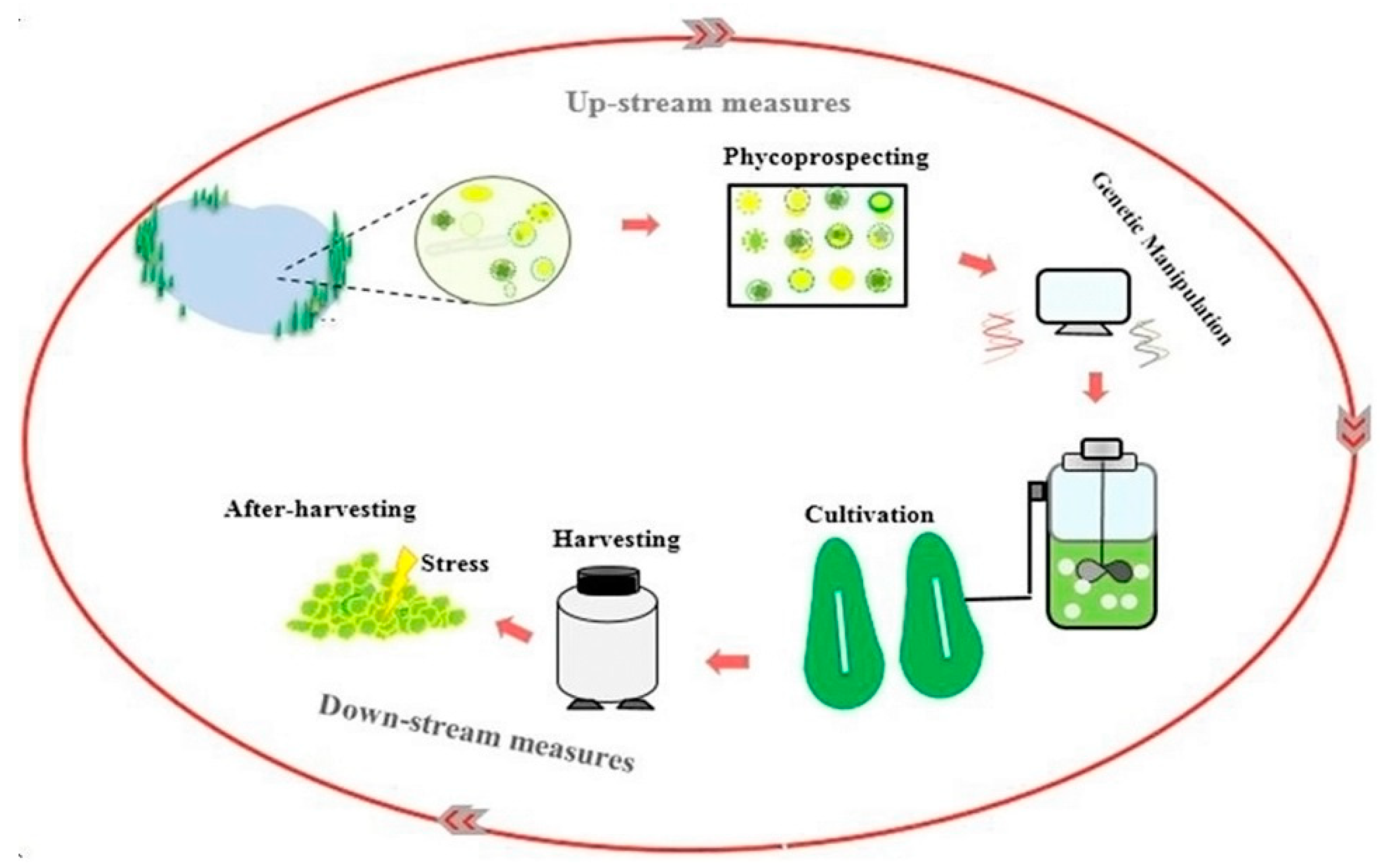
3. Upstream Measure
3.1. Phycoprospecting
3.2. Molecular Approaches
3.2.1. Lipid Biosynthesis Pathway
3.2.2. Molecular Approaches for Modulation the Stress-Related Mechanisms
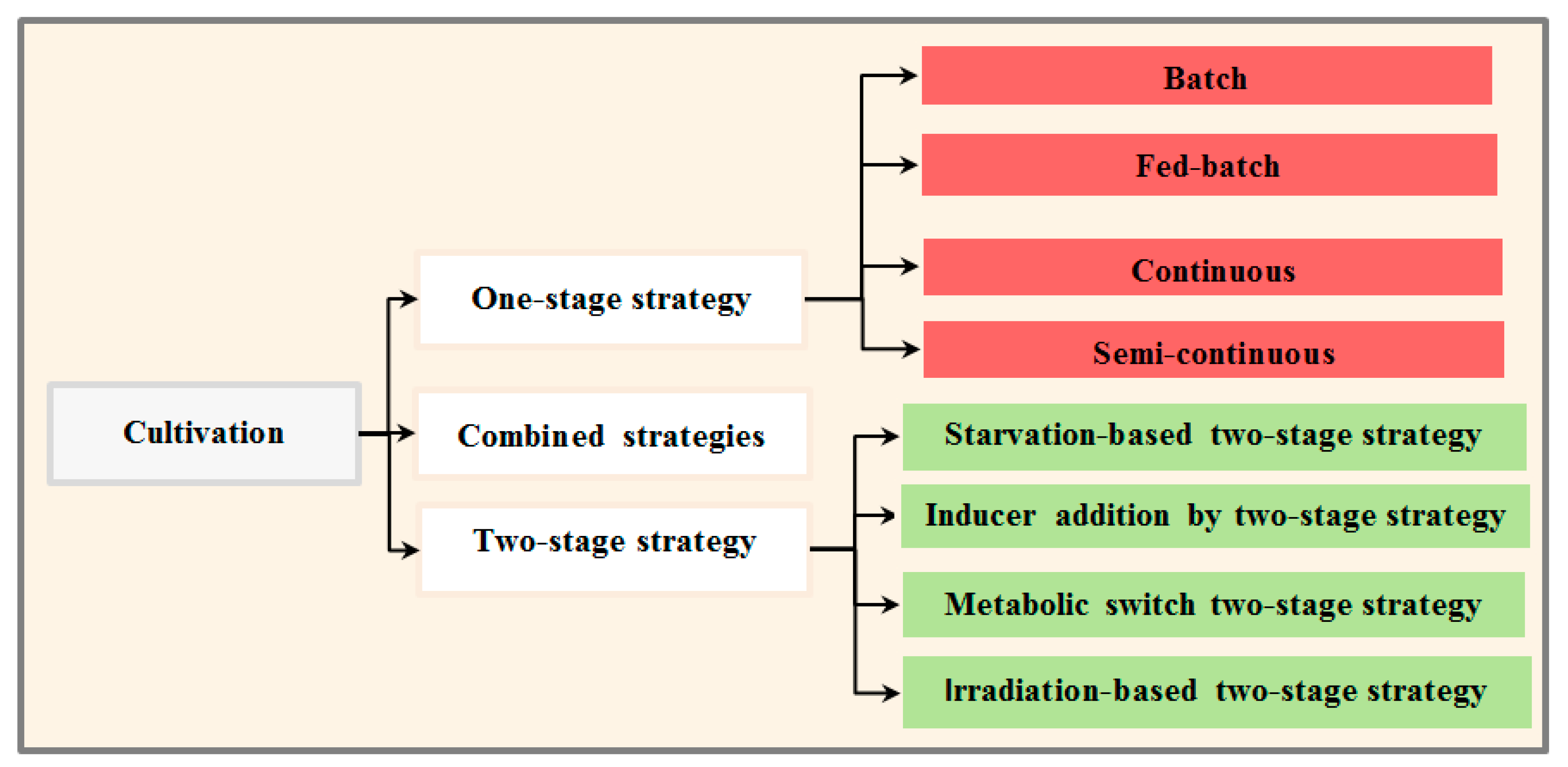
3.3. Cultivation Stage
3.3.1. One-Stage Cultivation Strategy
3.3.2. Two-Stage Strategy
Nutrient Starvation
Inducer Addition
Metabolic Switch
Irradiation
3.3.3. Combined Cultivation Strategies
4. Downstream Measure
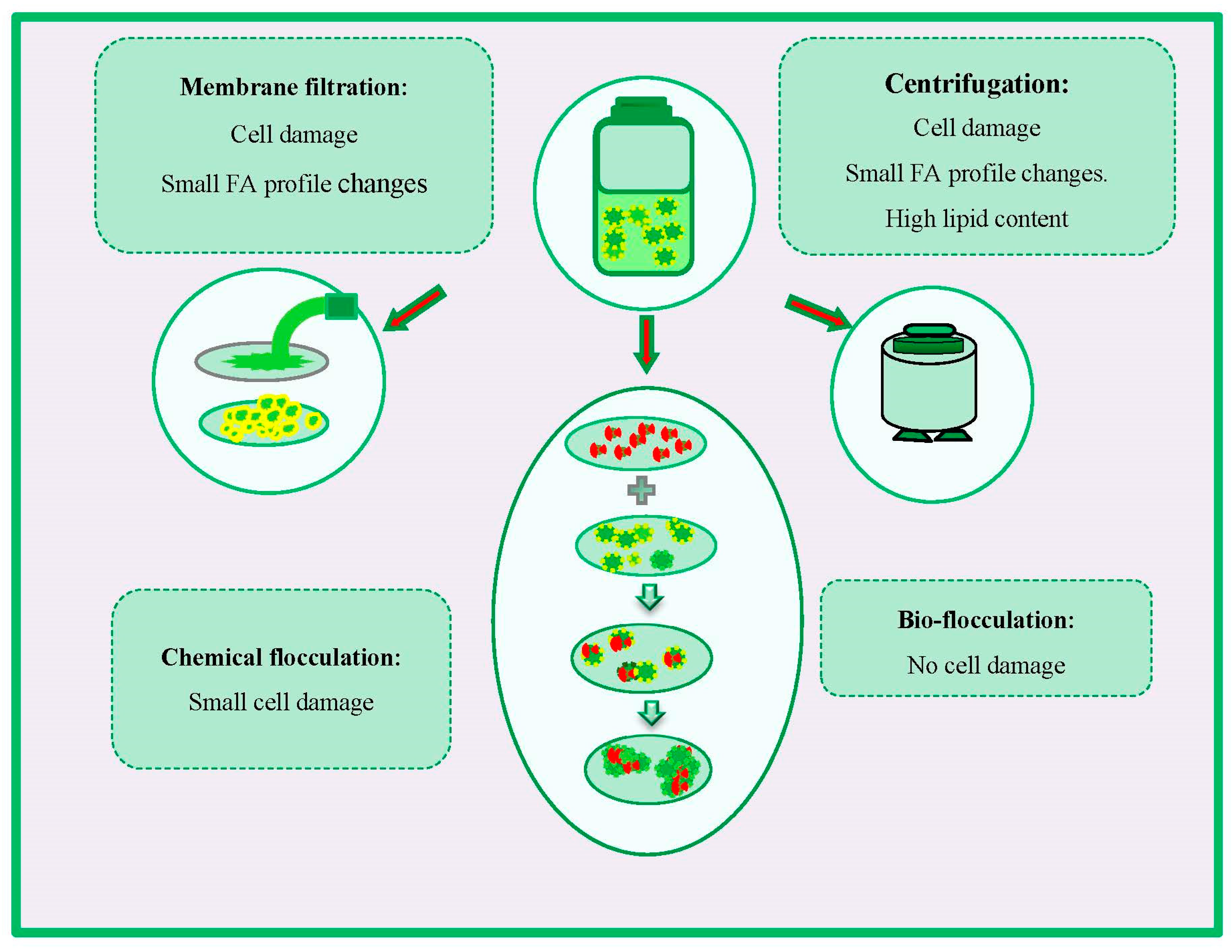
4.1. Harvesting Stage
4.2. Post-Harvesting Stage
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Singh, D.; Sharma, D.; Soni, S.; Sharma, S.; Sharma, P.K.; Jhalani, A. A review on feedstocks, production processes, and yield for different generations of biodiesel. Fuel 2020, 262, 116553. [Google Scholar] [CrossRef]
- Günay, M.E.; Türker, L.; Tapan, N.A. Significant parameters and technological advancements in biodiesel production systems. Fuel 2019, 250, 27–41. [Google Scholar] [CrossRef]
- Milano, J.; Ong, H.C.; Masjuki, H.; Chong, W.; Lam, M.K.; Loh, P.K.; Vellayan, V. Microalgae biofuels as an alternative to fossil fuel for power generation. Renew. Sustain. Energy Rev. 2016, 58, 180–197. [Google Scholar] [CrossRef]
- Ullah, K.; Ahmad, M.; Sharma, V.K.; Lu, P.; Harvey, A.; Zafar, M.; Sultana, S. Assessing the potential of algal biomass opportunities for bioenergy industry: A review. Fuel 2015, 143, 414–423. [Google Scholar] [CrossRef]
- Shanmugam, S.; Hari, A.; Pandey, A.; Mathimani, T.; Felix, L.; Pugazhendhi, A. Comprehensive review on the application of inorganic and organic nanoparticles for enhancing biohydrogen production. Fuel 2020, 270, 117453. [Google Scholar] [CrossRef]
- Sharma, Y.; Singh, B.; Upadhyay, S. Advancements in development and characterization of biodiesel: A review. Fuel 2008, 87, 2355–2373. [Google Scholar] [CrossRef]
- Stephen, J.L.; Periyasamy, B. Innovative developments in biofuels production from organic waste materials: A review. Fuel 2018, 214, 623–633. [Google Scholar] [CrossRef]
- Vassilev, S.V.; Vassileva, C.G. Composition, properties and challenges of algae biomass for biofuel application: An overview. Fuel 2016, 181, 1–33. [Google Scholar] [CrossRef]
- Khalid, A.A.H.; Yaakob, Z.; Abdullah, S.R.S.; Takriff, M.S. Growth improvement and metabolic profiling of native and commercial Chlorella sorokiniana strains acclimatized in recycled agricultural wastewater. Bioresour. Technol. 2018, 247, 930–939. [Google Scholar] [CrossRef]
- Deng, X.; Li, Y.; Fei, X. Microalgae: A promising feedstock for biodiesel. Afr. J. Microbiol. Res. 2009, 3, 1008–1014. [Google Scholar]
- Nayak, M.; Suh, W.I.; Chang, Y.K.; Lee, B. Exploration of two-stage cultivation strategies using nitrogen starvation to maximize the lipid productivity in Chlorella sp. HS2. Bioresour. Technol. 2019, 276, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Shokravi, H.; Shokravi, Z.; Heidarrezaei, M.; Lau, W.J.; Koloor, S.S.R.; Petrů, M.; Chyuan, O.H.; Ismail, A.F. Fourth Generation Biofuel: A Review on Challenges and Future Directions. Fuel 2020. under review. [Google Scholar]
- Adams, C.; Godfrey, V.; Wahlen, B.; Seefeldt, L.; Bugbee, B. Understanding precision nitrogen stress to optimize the growth and lipid content tradeoff in oleaginous green microalgae. Bioresour. Technol. 2013, 131, 188–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.; Wan, C.; Mehmood, M.A.; Chang, J.-S.; Bai, F.; Zhao, X. Manipulating environmental stresses and stress tolerance of microalgae for enhanced production of lipids and value-added products—A review. Bioresour. Technol. 2017, 244, 1198–1206. [Google Scholar] [CrossRef]
- Ghosh, A.; Khanra, S.; Mondal, M.; Halder, G.; Tiwari, O.; Saini, S.; Bhowmick, T.K.; Gayen, K. Progress toward isolation of strains and genetically engineered strains of microalgae for production of biofuel and other value added chemicals: A review. Energy Convers. Manag. 2016, 113, 104–118. [Google Scholar] [CrossRef]
- Sajjadi, B.; Chen, W.-Y.; Raman, A.A.A.; Ibrahim, S. Microalgae lipid and biomass for biofuel production: A comprehensive review on lipid enhancement strategies and their effects on fatty acid composition. Renew. Sustain. Energy Rev. 2018, 97, 200–232. [Google Scholar] [CrossRef]
- Shokravi, H.; Shokravi, Z.; Aziz, M.A.; Shokravi, H. 11 Algal Biofuel: A Promising. In Fossil Free Fuels: Trends in Renewable Energy; Taylor & Francis Group: Oxford, UK, 2019; pp. 187–211. [Google Scholar]
- Shokravi, H.; Shokravi, Z.; Aziz, M.A.; Shokravi, H. 12 The Fourth-Generation. In Fossil Free Fuels: Trends in Renewable Energy; Taylor & Francis Group: Oxford, UK, 2019; pp. 213–251. [Google Scholar]
- Ho, S.-H.; Ye, X.; Hasunuma, T.; Chang, J.-S.; Kondo, A. Perspectives on engineering strategies for improving biofuel production from microalgae—A critical review. Biotechnol. Adv. 2014, 32, 1448–1459. [Google Scholar] [CrossRef]
- Peng, L.; Fu, D.; Chu, H.; Wang, Z.; Qi, H. Biofuel production from microalgae: A review. Environ. Chem. Lett. 2019, 18, 285–297. [Google Scholar] [CrossRef]
- Lim, D.K.Y.; Schenk, P.M. Microalgae selection and improvement as oil crops: GM vs non-GM strain engineering. AIMS Environ. Sci. 2017, 4, 151–161. [Google Scholar] [CrossRef]
- He, Q.; Yang, H.; Xu, L.; Xia, L.; Hu, C. Sufficient utilization of natural fluctuating light intensity is an effective approach of promoting lipid productivity in oleaginous microalgal cultivation outdoors. Bioresour. Technol. 2015, 180, 79–87. [Google Scholar] [CrossRef]
- Přibyl, P.; Cepák, V.; Zachleder, V. Production of lipids in 10 strains of Chlorella and Parachlorella, and enhanced lipid productivity in Chlorella vulgaris. Appl. Microbiol. Biotechnol. 2012, 94, 549–561. [Google Scholar] [CrossRef]
- Kwak, H.S.; Kim, J.Y.H.; Woo, H.M.; Jin, E.; Min, B.K.; Sim, S.J. Synergistic effect of multiple stress conditions for improving microalgal lipid production. Algal Res. 2016, 19, 215–224. [Google Scholar] [CrossRef]
- Kim, H.S.; Hsu, S.-C.; Han, S.-I.; Thapa, H.R.; Guzman, A.R.; Browne, D.R.; Tatli, M.; Devarenne, T.P.; Stern, D.B.; Han, A. High-throughput droplet microfluidics screening platform for selecting fast-growing and high lipid-producing microalgae from a mutant library. Plant Direct 2017, 1, e00011. [Google Scholar] [CrossRef] [Green Version]
- Challagulla, V.; Nayar, S.; Walsh, K.; Fabbro, L. Advances in techniques for assessment of microalgal lipids. Crit. Rev. Biotechnol. 2016, 37, 566–578. [Google Scholar] [CrossRef]
- Cabanelas, I.T.D.; Van Der Zwart, M.; Kleinegris, D.M.M.; Barbosa, M.J.; Wijffels, R.H. Rapid method to screen and sort lipid accumulating microalgae. Bioresour. Technol. 2015, 184, 47–52. [Google Scholar] [CrossRef]
- Erickson, R.A.; Jimenez, R. Microfluidic cytometer for high-throughput measurement of photosynthetic characteristics and lipid accumulation in individual algal cells. Lab Chip 2013, 13, 2893–2901. [Google Scholar] [CrossRef]
- Liu, J.; Mukherjee, J.; Hawkes, J.J.; Wilkinson, S.J. Optimization of lipid production for algal biodiesel in nitrogen stressed cells of Dunaliella salina using FTIR analysis. J. Chem. Technol. Biotechnol. 2013, 88, 1807–1814. [Google Scholar] [CrossRef]
- Shokravi, Z.; Mehrad, L.; Ramazani, A. Detecting the frequency of aminoglycoside modifying enzyme encoding genes among clinical isolates of methicillin-resistant Staphylococcus aureus. BioImpacts 2015, 5, 87–91. [Google Scholar] [CrossRef] [Green Version]
- Shokravi, Z.; Haseli, M.; Mehrad, L.; Ramazani, A. Distribution of Staphylococcal cassette chromosome mecA (SCCmec) types among coagulase-negative Staphylococci isolates from healthcare workers in the North-West of Iran. Iran. J. Basic Med. Sci. 2020, 23, 1489–1493. [Google Scholar] [CrossRef]
- Ramazani, A.; Shokravi, Z.; Mehrad, L.; Sorouri, R.; Amirmoghaddami, H. Study the Genes Encoding Aminoglycoside-Modifying Enzymes among Clinical Isolates of Methicillin Resistance Staphylococcus Aureus. 2013. Available online: https://www.sid.ir/en/Seminar/ViewPaper.aspx?ID=32537 (accessed on 29 October 2020).
- Heidarrezaei, M.; Shokravi, H.; Huyop, F.; Koloor, S.R.; Petru, M. Isolation and Characterization of a Novel Bacterium from the Marine Environment for Trichloroacetic Acid Bioremediation. Appl. Sci. 2020, 10, 4593. [Google Scholar] [CrossRef]
- Sharma, P.K.; Saharia, M.; Srivstava, R.; Kumar, S.; Sahoo, L. Tailoring Microalgae for Efficient Biofuel Production. Front. Mar. Sci. 2018, 5, 382. [Google Scholar] [CrossRef]
- Trentacoste, E.M.; Shrestha, R.P.; Smith, S.R.; Glé, C.; Hartmann, A.C.; Hildebrand, M.; Gerwick, W.H. Metabolic engineering of lipid catabolism increases microalgal lipid accumulation without compromising growth. Proc. Natl. Acad. Sci. USA 2013, 110, 19748–19753. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.H.T.; Park, S.; Jeong, J.; Shin, Y.S.; Sim, S.J.; Jin, E. Enhancing lipid productivity by modulating lipid catabolism using the CRISPR-Cas9 system in Chlamydomonas. Environ. Biol. Fishes 2020, 32, 2829–2840. [Google Scholar] [CrossRef]
- Park, S.; Nguyen, T.H.T.; Jin, E. Improving lipid production by strain development in microalgae: Strategies, challenges and perspectives. Bioresour. Technol. 2019, 292, 121953. [Google Scholar] [CrossRef]
- Xia, L.; Ge, H.; Zhou, X.; Zhang, D.; Hu, C. Photoautotrophic outdoor two-stage cultivation for oleaginous microalgae Scenedesmus obtusus XJ-15. Bioresour. Technol. 2013, 144, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.-H.; Cho, D.-H.; Lee, S.; Heo, J.; Tran, Q.-G.; Chang, Y.K.; Kim, H.-S. Hybrid operation of photobioreactor and wastewater-fed open raceway ponds enhances the dominance of target algal species and algal biomass production. Algal Res. 2018, 29, 319–329. [Google Scholar] [CrossRef]
- Nagappan, S.; Devendran, S.; Tsai, P.-C.; Dahms, H.-U.; Ponnusamy, V.K. Potential of two-stage cultivation in microalgae biofuel production. Fuel 2019, 252, 339–349. [Google Scholar] [CrossRef]
- Aziz, M.A.; Kassim, K.A.; Shokravi, Z.; Jakarni, F.M.; Liu, H.Y.; Zaini, N.; Tan, L.S.; Islam, A.S.; Shokravi, H.; Lieu, H.Y. Two-stage cultivation strategy for simultaneous increases in growth rate and lipid content of microalgae: A review. Renew. Sustain. Energy Rev. 2020, 119, 109621. [Google Scholar] [CrossRef]
- Bhatia, S.; Mehariya, S.; Bhatia, R.K.; Kumar, M.; Pugazhendhi, A.; Awasthi, M.K.; Atabani, A.; Kumar, G.; Kim, W.; Seo, S.-O.; et al. Wastewater based microalgal biorefinery for bioenergy production: Progress and challenges. Sci. Total. Environ. 2021, 751, 141599. [Google Scholar] [CrossRef]
- Borowitzka, M.A. Species and strain selection. In Algae for Biofuels and Energy; Springer: Berlin, Germany, 2013; pp. 77–89. [Google Scholar]
- Singh, P.; Kumari, S.; Guldhe, A.; Misra, R.; Rawat, I.; Bux, F. Trends and novel strategies for enhancing lipid accumulation and quality in microalgae. Renew. Sustain. Energy Rev. 2016, 55, 1–16. [Google Scholar] [CrossRef]
- Chu, W.-L. Strategies to enhance production of microalgal biomass and lipids for biofuel feedstock. Eur. J. Phycol. 2017, 52, 419–437. [Google Scholar] [CrossRef]
- Chung, Y.-S.; Lee, J.-W.; Chung, C.-H. Molecular challenges in microalgae towards cost-effective production of quality biodiesel. Renew. Sustain. Energy Rev. 2017, 74, 139–144. [Google Scholar] [CrossRef]
- Shin, Y.S.; Choi, H.I.; Choi, J.W.; Lee, J.S.; Sung, Y.J.; Sim, S.J. Multilateral approach on enhancing economic viability of lipid production from microalgae: A review. Bioresour. Technol. 2018, 258, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Piligaev, A.V.; Sorokina, K.N.; Samoylova, Y.V.; Parmon, V.N. Production of Microalgal Biomass with High Lipid Content and Their Catalytic Processing Into Biodiesel: A Review. Catal. Ind. 2019, 11, 349–359. [Google Scholar] [CrossRef]
- Menegazzo, M.L.; Fonseca, G.G. Biomass recovery and lipid extraction processes for microalgae biofuels production: A review. Renew. Sustain. Energy Rev. 2019, 107, 87–107. [Google Scholar] [CrossRef]
- Poh, Z.L.; Kadir, W.N.A.; Lam, M.-K.; Uemura, Y.; Suparmaniam, U.; Lim, J.W.; Show, P.L.; Lee, K.T. The effect of stress environment towards lipid accumulation in microalgae after harvesting. Renew. Energy 2020, 154, 1083–1091. [Google Scholar] [CrossRef]
- Vandamme, D.; Gheysen, L.; Muylaert, K.; Foubert, I. Impact of harvesting method on total lipid content and extraction efficiency for Phaeodactylum tricornutum. Sep. Purif. Technol. 2018, 194, 362–367. [Google Scholar] [CrossRef]
- Liu, Y.; Jin, W.; Zhou, X.; Han, S.-F.; Tu, R.; Feng, X.; Jensen, P.D.; Wang, Q. Efficient harvesting of Chlorella pyrenoidosa and Scenedesmus obliquus cultivated in urban sewage by magnetic flocculation using nano-Fe3O4 coated with polyethyleneimine. Bioresour. Technol. 2019, 290, 121771. [Google Scholar] [CrossRef]
- Nwokoagbara, E.; Olaleye, A.K.; Wang, M. Biodiesel from microalgae: The use of multi-criteria decision analysis for strain selection. Fuel 2015, 159, 241–249. [Google Scholar] [CrossRef]
- Sibi, G.; Shetty, V.; Mokashi, K. Enhanced lipid productivity approaches in microalgae as an alternate for fossil fuels—A review. J. Energy Inst. 2016, 89, 330–334. [Google Scholar] [CrossRef]
- Gao, Y.; Yang, M.; Wang, C. Nutrient deprivation enhances lipid content in marine microalgae. Bioresour. Technol. 2013, 147, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Wan, L.; Li, A.; Zhang, C. Responses in growth, lipid accumulation, and fatty acid composition of four oleaginous microalgae to different nitrogen sources and concentrations. Chin. J. Oceanol. Limnol. 2013, 31, 1306–1314. [Google Scholar] [CrossRef]
- Yu, N.; Dieu, L.T.J.; Harvey, S.; Lee, D.-Y. Optimization of process configuration and strain selection for microalgae-based biodiesel production. Bioresour. Technol. 2015, 193, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Ren, Y.; Mao, X.; Li, X.; Zhang, H.; Lao, Y.; Chen, F. Harnessing C/N balance of Chromochloris zofingiensis to overcome the potential conflict in microalgal production. Commun. Biol. 2020, 3, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, H.-J.; Su, C.-H.; Chien, L.-J. Accumulation of lipid production in Chlorella minutissima by triacylglycerol biosynthesis-related genes cloned from Saccharomyces cerevisiae and Yarrowia lipolytica. J. Microbiol. 2012, 50, 526–534. [Google Scholar] [CrossRef]
- Moussa, I.D.-B.; Chtourou, H.; Hassairi, I.; Sayadi, S.; Dhouib, A. The effect of switching environmental conditions on content and structure of lipid produced by a wild strain Picochlorum sp. Renew. Energy 2019, 134, 406–415. [Google Scholar] [CrossRef]
- Pedro, A.S.; González-López, C.; Acién, F.; Molina-Grima, E. Marine microalgae selection and culture conditions optimization for biodiesel production. Bioresour. Technol. 2013, 134, 353–361. [Google Scholar] [CrossRef]
- Pal, P.; Chew, K.W.; Yen, H.-W.; Lim, J.W.; Lam, M.K.; Show, P.L. Cultivation of oily microalgae for the production of third-generation biofuels. Sustainability 2019, 11, 5424. [Google Scholar] [CrossRef] [Green Version]
- De Bhowmick, G.; Koduru, L.; Sen, R. Metabolic pathway engineering towards enhancing microalgal lipid biosynthesis for biofuel application—A review. Renew. Sustain. Energy Rev. 2015, 50, 1239–1253. [Google Scholar] [CrossRef]
- El Arroussi, H.; Benhima, R.; El Mernissi, N.; Bouhfid, R.; Tilsaghani, C.; Bennis, I.; Wahby, I. Screening of marine microalgae strains from Moroccan coasts for biodiesel production. Renew. Energy 2017, 113, 1515–1522. [Google Scholar] [CrossRef]
- Huang, S.T.; Goh, J.L.; Ahmadzadeh, H.; Murry, M.A. A rapid sampling technique for isolating highly productive lipid-rich algae strains from environmental samples. Biofuel Res. J. 2019, 6, 920–926. [Google Scholar] [CrossRef] [Green Version]
- Neofotis, P.; Huang, A.; Sury, K.; Chang, W.; Joseph, F.; Gabr, A.; Twary, S.; Qiu, W.; Holguin, O.; Polle, J.E. Characterization and classification of highly productive microalgae strains discovered for biofuel and bioproduct generation. Algal Res. 2016, 15, 164–178. [Google Scholar] [CrossRef] [Green Version]
- Andersen, R.A.; Kawachi, M. Traditional Microalgae Isolation Techniques. In Algal Culturing Techniques; Elsevier Academic Press: Burlington, MA, USA, 2005; pp. 83–100. [Google Scholar]
- Mutanda, T.; Ramesh, D.; Karthikeyan, S.; Kumari, S.; Anandraj, A.; Bux, F. Bioprospecting for hyper-lipid producing microalgal strains for sustainable biofuel production. Bioresour. Technol. 2011, 102, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Cabanelas, I.T.D.; Van Der Zwart, M.; Kleinegris, D.M.M.; Wijffels, R.H.; Barbosa, M.J. Sorting cells of the microalga Chlorococcum littorale with increased triacylglycerol productivity. Biotechnol. Biofuels 2016, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Pereira, H.; Barreira, L.; Mozes, A.; Florindo, C.; Polo, C.; Duarte, C.V.; Custodio, L.; Varela, J. Microplate-based high throughput screening procedure for the isolation of lipid-rich marine microalgae. Biotechnol. Biofuels 2011, 4, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.S.; Guzman, A.R.; Thapa, H.R.; Devarenne, T.P.; Han, A. A droplet microfluidics platform for rapid microalgal growth and oil production analysis. Biotechnol. Bioeng. 2016, 113, 1691–1701. [Google Scholar] [CrossRef]
- Del Río, E.; Acién, F.G.; Guerrero, M.G.; Sanchez, E.D.R. Photoautotrophic production of astaxanthin by the microalga Haematococcus pluvialis. In Sustainable Biotechnology; Springer: Dordrecht, The Netherlands, 2010; pp. 247–258. [Google Scholar]
- Sung, M.-G.; Lee, B.; Kim, C.W.; Nam, K.; Chang, Y.K. Enhancement of lipid productivity by adopting multi-stage continuous cultivation strategy in Nannochloropsis gaditana. Bioresour. Technol. 2017, 229, 20–25. [Google Scholar] [CrossRef]
- Abdullah, B.; Muhammad, S.A.F.S.; Shokravi, Z.; Ismail, S.; Kassim, K.A.; Mahmood, A.N.; Aziz, M.A. Fourth generation biofuel: A review on risks and mitigation strategies. Renew. Sustain. Energy Rev. 2019, 107, 37–50. [Google Scholar] [CrossRef]
- Davis, R.; Aden, A.; Pienkos, P.T. Techno-economic analysis of autotrophic microalgae for fuel production. Appl. Energy 2011, 88, 3524–3531. [Google Scholar] [CrossRef]
- Eguiheneuf, F.; Ekhan, A.; Tran, L.-S.P. Genetic Engineering: A Promising Tool to Engender Physiological, Biochemical, and Molecular Stress Resilience in Green Microalgae. Front. Plant Sci. 2016, 7, 400. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.W.M.; Lee, Y.-K. The dilemma for lipid productivity in green microalgae: Importance of substrate provision in improving oil yield without sacrificing growth. Biotechnol. Biofuels 2016, 9, 255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talebi, A.F.; Tohidfar, M.; Bagheri, A.; Lyon, S.R.; Salehi-Ashtiani, K.; Tabatabaei, M. Manipulation of carbon flux into fatty acid biosynthesis pathway in Dunaliella salina using AccD and ME genes to enhance lipid content and to improve produced biodiesel quality. Biofuel Res. J. 2014, 1, 91–97. [Google Scholar] [CrossRef]
- Xue, J.; Niu, Y.-F.; Huang, T.; Yang, W.-D.; Liu, J.-S.; Li, H.-Y. Genetic improvement of the microalga Phaeodactylum tricornutum for boosting neutral lipid accumulation. Metab. Eng. 2015, 27, 1–9. [Google Scholar] [CrossRef]
- Ma, Y.-H.; Wang, X.; Niu, Y.-F.; Yang, Z.-K.; Zhang, M.-H.; Wang, Z.; Yang, W.-D.; Liu, J.-S.; Li, H.-Y. Antisense knockdown of pyruvate dehydrogenase kinase promotes the neutral lipid accumulation in the diatom Phaeodactylum tricornutum. Microb. Cell Factories 2014, 13, 100. [Google Scholar] [CrossRef] [Green Version]
- Muto, M.; Tanaka, M.; Liang, Y.; Yoshino, T.; Matsumoto, M.; Tanaka, T. Enhancement of glycerol metabolism in the oleaginous marine diatom Fistulifera solaris JPCC DA0580 to improve triacylglycerol productivity. Biotechnol. Biofuels 2015, 8, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, J.; Ning, K.; Zeng, X.; Luo, Y.; Wang, D.; Hu, J.; Li, J.; Xu, H.; Huang, J.; Wan, M.; et al. Genomic foundation of starch-to-lipid switch in Oleaginous Chlorella spp. Plant Physiol. 2015, 169, 2444–2461. [Google Scholar]
- Deng, X.; Li, Y.; Fei, X. The mRNA abundance of pepc2 gene is negatively correlated with oil content in Chlamydomonas reinhardtii. Biomass Bioenergy 2011, 35, 1811–1817. [Google Scholar] [CrossRef]
- Muñoz, C.F.; Weusthuis, R.A.; D’Adamo, S.; Wijffels, R.H. Effect of Single and Combined Expression of Lysophosphatidic Acid Acyltransferase, Glycerol-3-Phosphate Acyltransferase, and Diacylglycerol Acyltransferase on Lipid Accumulation and Composition in Neochloris oleoabundans. Front. Plant Sci. 2019, 10, 1573. [Google Scholar] [CrossRef]
- Shahid, A.; Rehman, A.U.; Usman, M.; Ashraf, M.U.F.; Javed, M.R.; Khan, A.Z.; Gill, S.S.; Mehmood, M.A. Engineering the metabolic pathways of lipid biosynthesis to develop robust microalgal strains for biodiesel production. Biotechnol. Appl. Biochem. 2020, 67, 41–51. [Google Scholar] [CrossRef]
- Ran, W.; Wang, H.; Liu, Y.; Qi, M.; Xiang, Q.; Yao, C.; Zhang, Y.; Lan, X. Storage of starch and lipids in microalgae: Biosynthesis and manipulation by nutrients. Bioresour. Technol. 2019, 291, 121894. [Google Scholar] [CrossRef]
- Zhu, B.-H.; Shi, H.-P.; Yang, G.; Lv, N.-N.; Yang, M.; Pan, K.-H. Silencing UDP-glucose pyrophosphorylase gene in Phaeodactylum tricornutum affects carbon allocation. New Biotechnol. 2016, 33, 237–244. [Google Scholar] [CrossRef]
- Wan, M.; Jin, X.; Xia, J.; Rosenberg, J.N.; Yu, G.; Nie, Z.; Oyler, G.A.; Betenbaugh, M. The effect of iron on growth, lipid accumulation, and gene expression profile of the freshwater microalga Chlorella sorokiniana. Appl. Microbiol. Biotechnol. 2014, 98, 9473–9481. [Google Scholar] [CrossRef]
- Chauton, M.S.; Winge, P.; Brembu, T.; Vadstein, O.; Bones, A.M. Gene Regulation of Carbon Fixation, Storage, and Utilization in the Diatom Phaeodactylum tricornutum Acclimated to Light/Dark Cycles. Plant Physiol. 2012, 161, 1034–1048. [Google Scholar] [CrossRef] [Green Version]
- Solovchenko, A. Physiological role of neutral lipid accumulation in eukaryotic microalgae under stresses. Russ. J. Plant Physiol. 2012, 59, 167–176. [Google Scholar] [CrossRef]
- Iwai, M.; Ikeda, K.; Shimojima, M.; Ohta, H. Enhancement of extraplastidic oil synthesis in C hlamydomonas reinhardtii using a type-2 diacylglycerol acyltransferase with a phosphorus starvation–inducible promoter. Plant Biotechnol. J. 2014, 12, 808–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eiwai, M.; Ehori, K.; Esasaki-Sekimoto, Y.; Eshimojima, M.; Ohta, H. Manipulation of oil synthesis in Nannochloropsis strain NIES-2145 with a phosphorus starvation–inducible promoter from Chlamydomonas reinhardtii. Front. Microbiol. 2015, 6, 912. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Hao, Q.; Bai, L.; Xu, J.; Yin, W.-B.; Song, L.; Xu, L.; Guo, X.; Fan, C.; Chen, Y.; et al. Overexpression of the soybean transcription factor GmDof4 significantly enhances the lipid content of Chlorella ellipsoidea. Biotechnol. Biofuels 2014, 7, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, C.-H.; Warakanont, J.; Takeuchi, T.; Sears, B.B.; Moellering, E.R.; Benning, C. The protein Compromised Hydrolysis of Triacylglycerols 7 (CHT7) acts as a repressor of cellular quiescence in Chlamydomonas. Proc. Natl. Acad. Sci. USA 2014, 111, 15833–15838. [Google Scholar] [CrossRef] [Green Version]
- Ngan, C.Y.; Wong, C.-H.; Choi, C.; Yoshinaga, Y.; Louie, K.; Jia, J.; Chen, C.; Bowen, B.; Cheng, H.; Leonelli, L.; et al. Lineage-specific chromatin signatures reveal a regulator of lipid metabolism in microalgae. Nat. Plants 2015, 1, 15107. [Google Scholar] [CrossRef]
- Ajjawi, I.; Verruto, J.; Aqui, M.; Soriaga, L.B.; Coppersmith, J.; Kwok, K.; Peach, L.; Orchard, E.; Kalb, R.; Xu, W.; et al. Lipid production in Nannochloropsis gaditana is doubled by decreasing expression of a single transcriptional regulator. Nat. Biotechnol. 2017, 35, 647–652. [Google Scholar] [CrossRef] [PubMed]
- Kang, N.K.; Jeon, S.; Kwon, S.; Koh, H.G.; Shin, S.-E.; Lee, B.; Choi, G.-G.; Yang, J.-W.; Jeong, B.-R.; Chang, Y.K. Effects of overexpression of a bHLH transcription factor on biomass and lipid production in Nannochloropsis salina. Biotechnol. Biofuels 2015, 8, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, S.; Kang, N.K.; Koh, H.G.; Shin, S.-E.; Lee, B.; Jeong, B.-R.; Chang, Y.K. Enhancement of biomass and lipid productivity by overexpression of a bZIP transcription factor in Nannochloropsis salina. Biotechnol. Bioeng. 2017, 115, 331–340. [Google Scholar] [CrossRef] [Green Version]
- Liang, M.-H.; Jiang, J.-G. Analysis of carotenogenic genes promoters and WRKY transcription factors in response to salt stress in Dunaliella bardawil. Sci. Rep. 2017, 7, 37025. [Google Scholar] [CrossRef]
- Matthijs, M.; Fabris, M.; Broos, S.; Vyverman, W.; Goossens, A. Profiling of the Early Nitrogen Stress Response in the Diatom Phaeodactylum tricornutum Reveals a Novel Family of RING-Domain Transcription Factors. Plant Physiol. 2015, 170, 489–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Z.; Li, Y.; Wu, G.; Li, G.; Sun, H.; Deng, S.; Shen, Y.; Chen, G.; Zhang, R.; Meng, C.; et al. Transcriptome Analysis in Haematococcus pluvialis: Astaxanthin Induction by Salicylic Acid (SA) and Jasmonic Acid (JA). PLoS ONE 2015, 10, e0140609. [Google Scholar] [CrossRef]
- Tevatia, R.; Allen, J.; Blum, P.; Demirel, Y.; Black, P. Modeling of rhythmic behavior in neutral lipid production due to continuous supply of limited nitrogen: Mutual growth and lipid accumulation in microalgae. Bioresour. Technol. 2014, 170, 152–159. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Muthuraj, M.; Palabhanvi, B.; Das, D. Synchronized growth and neutral lipid accumulation in Chlorella sorokiniana FC6 IITG under continuous mode of operation. Bioresour. Technol. 2016, 200, 770–779. [Google Scholar] [CrossRef]
- Wen, X.; Geng, Y.; Li, Y. Enhanced lipid production in Chlorella pyrenoidosa by continuous culture. Bioresour. Technol. 2014, 161, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Del Río, E.; García-Gómez, E.; Moreno, J.; Guerrero, M.G.; Garcia-González, M.; Sanchez, E.D.R. Microalgae for oil. Assessment of fatty acid productivity in continuous culture by two high-yield strains, Chlorococcum oleofaciens and Pseudokirchneriella subcapitata. Algal Res. 2017, 23, 37–42. [Google Scholar] [CrossRef] [Green Version]
- Sobczuk, T.M.; Chisti, Y. Potential fuel oils from the microalga Choricystis minor. J. Chem. Technol. Biotechnol. 2010, 85, 100–108. [Google Scholar] [CrossRef]
- Tang, H.; Chen, M.; Ng, K.S.; Salley, S.O. Continuous microalgae cultivation in a photobioreactor. Biotechnol. Bioeng. 2012, 109, 2468–2474. [Google Scholar] [CrossRef] [PubMed]
- Klok, A.J.; Martens, D.E.; Wijffels, R.H.; Lamers, P.P. Simultaneous growth and neutral lipid accumulation in microalgae. Bioresour. Technol. 2013, 134, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.R.; Glé, C.; Abbriano, R.M.; Traller, J.C.; Davis, A.; Trentacoste, E.; Vernet, M.; Allen, A.E.; Hildebrand, M. Transcript level coordination of carbon pathways during silicon starvation-induced lipid accumulation in the diatom Thalassiosira pseudonana. New Phytol. 2016, 210, 890–904. [Google Scholar] [CrossRef] [PubMed]
- Zaparoli, M.; Ziemniczak, F.G.; Mantovani, L.; Costa, J.A.V.; Colla, L.M. Cellular Stress Conditions as a Strategy to Increase Carbohydrate Productivity in Spirulina platensis. Bioenergy Res. 2020, 1–14. [Google Scholar] [CrossRef]
- Jiang, X.-M.; Han, Q.; Gao, X.; Gao, G. Conditions optimising on the yield of biomass, total lipid, and valuable fatty acids in two strains of Skeletonema menzelii. Food Chem. 2016, 194, 723–732. [Google Scholar] [CrossRef]
- Brennan, L.; Owende, P. Biofuels from microalgae—A review of technologies for production, processing, and extractions of biofuels and co-products. Renew. Sustain. Energy Rev. 2010, 14, 557–577. [Google Scholar] [CrossRef]
- Rezania, S.; Oryani, B.; Cho, J.; Sabbagh, F.; Rupani, P.F.; Talaiekhozani, A.; Rahimi, N.; Ghahroud, M.L. Technical Aspects of Biofuel Production from Different Sources in Malaysia—A Review. Processes 2020, 8, 993. [Google Scholar] [CrossRef]
- Hu, X.; Liu, B.; Deng, Y.; Bao, X.; Yang, A.; Zhou, J. A novel two-stage culture strategy used to cultivate Chlorella vulgaris for increasing the lipid productivity. Sep. Purif. Technol. 2019, 211, 816–822. [Google Scholar] [CrossRef]
- Rezania, S.; Mohamad, S.E. Response Surface Methodology for Optimization of Ethanol Production from Cocoa Waste. J. Energy Environ. Pollut. 2020, 1, 7–12. [Google Scholar]
- Cheng, D.; Li, D.; Yuan, Y.; Zhou, L.; Li, X.; Wu, T.; Wang, L.; Zhao, Q.; Wei, W.; Sun, Y. Improving carbohydrate and starch accumulation in Chlorella sp. AE10 by a novel two-stage process with cell dilution. Biotechnol. Biofuels 2017, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, M.J.; Harrison, S.T.L. Lipid productivity as a key characteristic for choosing algal species for biodiesel production. Environ. Biol. Fishes 2009, 21, 493–507. [Google Scholar] [CrossRef]
- Gao, G.; Wu, M.; Fu, Q.; Li, X.; Xu, J. A two-stage model with nitrogen and silicon limitation enhances lipid productivity and biodiesel features of the marine bloom-forming diatom Skeletonema costatum. Bioresour. Technol. 2019, 289, 121717. [Google Scholar] [CrossRef]
- Álvarez-Díaz, P.D.; Ruiz, J.; Arbib, Z.; Barragán, J.; Garrido-Pérez, C.; Perales, J.A. Lipid Production of Microalga Ankistrodesmus falcatus Increased by Nutrient and Light Starvation in a Two-Stage Cultivation Process. Appl. Biochem. Biotechnol. 2014, 174, 1471–1483. [Google Scholar] [CrossRef] [PubMed]
- Anne-Marie, K.; Yee, W.; Loh, S.H.; Aziz, A.; Cha, T.S. Effects of Excess and Limited Phosphate on Biomass, Lipid and Fatty Acid Contents and the Expression of Four Fatty Acid Desaturase Genes in the Tropical Selenastraceaen Messastrum gracile SE-MC4. Appl. Biochem. Biotechnol. 2019, 190, 1438–1456. [Google Scholar] [CrossRef] [PubMed]
- Shokravi, H.; Shaghaghi, T.M. Experimental Study on After-Mixing Temperature Control of Concrete in Batching Plant Production. IOP Mater. Sci. Eng. 2020, 713. [Google Scholar] [CrossRef]
- Ahmad, A.; Osman, S.M.; Cha, T.C.; Loh, S.H. Phosphate-induced changes in fatty acid biosynthesis in Chlorella sp. KS-MA2 Strain. Bio Technologia 2016, 97, 295–304. [Google Scholar] [CrossRef]
- Xia, L.; Rong, J.; Yang, H.; He, Q.; Zhang, D.; Hu, C. NaCl as an effective inducer for lipid accumulation in freshwater microalgae Desmodesmus abundans. Bioresour. Technol. 2014, 161, 402–409. [Google Scholar] [CrossRef]
- Goh, B.H.H.; Ong, H.C.; Cheah, M.Y.; Chen, W.-H.; Yu, K.L.; Mahlia, T.M.I. Sustainability of direct biodiesel synthesis from microalgae biomass: A critical review. Renew. Sustain. Energy Rev. 2019, 107, 59–74. [Google Scholar] [CrossRef]
- Park, H.; Jung, D.; Lee, J.; Kim, P.; Cho, Y.; Jung, I.; Kim, Z.-H.; Lim, S.-M.; Lee, C.-G. Improvement of biomass and fatty acid productivity in ocean cultivation of Tetraselmis sp. using hypersaline medium. Environ. Biol. Fishes 2018, 30, 2725–2735. [Google Scholar] [CrossRef]
- Ra, C.-H.; Kang, C.-H.; Jung, J.-H.; Jeong, G.-T.; Kim, S.-K. Effects of light-emitting diodes (LEDs) on the accumulation of lipid content using a two-phase culture process with three microalgae. Bioresour. Technol. 2016, 212, 254–261. [Google Scholar] [CrossRef]
- Yang, H.; He, Q.; Rong, J.; Xia, L.; Hu, C. Rapid neutral lipid accumulation of the alkali-resistant oleaginous Monoraphidium dybowskii LB50 by NaCl induction. Bioresour. Technol. 2014, 172, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Pancha, I.; Chokshi, K.; Maurya, R.; Trivedi, K.; Patidar, S.K.; Ghosh, A.; Mishra, S. Salinity induced oxidative stress enhanced biofuel production potential of microalgae Scenedesmus sp. CCNM 1077. Bioresour. Technol. 2015, 189, 341–348. [Google Scholar] [CrossRef]
- Wensel, P.; Helms, G.; Hiscox, B.; Davis, W.C.; Kirchhoff, H.; Bule, M.; Yu, L.; Chen, S. Isolation, characterization, and validation of oleaginous, multi-trophic, and haloalkaline-tolerant microalgae for two-stage cultivation. Algal Res. 2014, 4, 2–11. [Google Scholar] [CrossRef]
- Srinivasan, R.; Mageswari, A.; Subramanian, P.; Suganthi, C.; Chaitanyakumar, A.; Aswini, V.; Gothandam, K.M. Bicarbonate supplementation enhances growth and biochemical composition of Dunaliella salina V-101 by reducing oxidative stress induced during macronutrient deficit conditions. Sci. Rep. 2018, 8, 6972. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Wang, H.-P.; Han, B.; Yu, X. Coupling of abiotic stresses and phytohormones for the production of lipids and high-value by-products by microalgae: A review. Bioresour. Technol. 2019, 274, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Guldhe, A.; Renuka, N.; Singh, P.; Bux, F. Effect of phytohormones from different classes on gene expression of Chlorella sorokiniana under nitrogen limitation for enhanced biomass and lipid production. Algal Res. 2019, 40, 101518. [Google Scholar] [CrossRef]
- Sivaramakrishnan, R.; Incharoensakdi, A. Enhancement of total lipid yield by nitrogen, carbon, and iron supplementation in isolated microalgae. J. Phycol. 2017, 53, 855–868. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, D.; Xu, J.-W.; Zhao, P.; Li, T.; Ma, H.; Yu, X. Melatonin enhances lipid production in Monoraphidium sp. QLY-1 under nitrogen deficiency conditions via a multi-level mechanism. Bioresour. Technol. 2018, 259, 46–53. [Google Scholar] [CrossRef]
- Che, R.; Ding, K.; Huang, L.; Zhao, P.; Xu, J.-W.; Li, T.; Ma, H.; Yu, X. Enhancing biomass and oil accumulation of Monoraphidium sp. FXY-10 by combined fulvic acid and two-step cultivation. J. Taiwan Inst. Chem. Eng. 2016, 67, 161–165. [Google Scholar] [CrossRef]
- Babu, A.G.; Wu, X.; Kabra, A.N.; Kim, D.-P. Cultivation of an indigenous Chlorella sorokiniana with phytohormones for biomass and lipid production under N-limitation. Algal Res. 2017, 23, 178–185. [Google Scholar] [CrossRef]
- Su, C.-H.; Chien, L.-J.; Gomes, J.; Lin, Y.-S.; Yu, Y.-K.; Liou, J.-S.; Syu, R.-J. Factors affecting lipid accumulation by Nannochloropsis oculata in a two-stage cultivation process. Environ. Biol. Fishes 2010, 23, 903–908. [Google Scholar] [CrossRef]
- Abomohra, A.E.-F.; El-Naggar, A.H.; Alaswad, S.O.; Elsayed, M.; Li, M.; Li, W. Enhancement of biodiesel yield from a halophilic green microalga isolated under extreme hypersaline conditions through stepwise salinity adaptation strategy. Bioresour. Technol. 2020, 310, 123462. [Google Scholar] [CrossRef]
- Hang, L.T.; Mori, K.; Tanaka, Y.; Morikawa, M.; Toyama, T. Enhanced lipid productivity of Chlamydomonas reinhardtii with combination of NaCl and CaCl2 stresses. Bioprocess Biosyst. Eng. 2020, 43, 971–980. [Google Scholar] [CrossRef] [PubMed]
- Mirizadeh, S.; Nosrati, M.; Shojaosadati, S.A. Synergistic Effect of Nutrient and Salt Stress on Lipid Productivity of Chlorella vulgaris through Two-Stage Cultivation. BioEnergy Res. 2019, 13, 507–517. [Google Scholar] [CrossRef]
- Acién, F.; Molina, E.; Reis, A.; Torzillo, G.; Zittelli, G.; Sepúlveda, C.; Masojídek, J. Photobioreactors for the production of microalgae. In Microalgae-Based Biofuels and Bioproducts; Elsevier: Cambridge, UK, 2017; pp. 1–44. [Google Scholar]
- Yen, H.-W.; Chang, J.-T. A two-stage cultivation process for the growth enhancement of Chlorella vulgaris. Bioprocess Biosyst. Eng. 2013, 36, 1797–1801. [Google Scholar] [CrossRef]
- Cheah, W.Y.; Show, P.-L.; Juan, J.C.; Chang, J.-S.; Ling, T.C. Enhancing biomass and lipid productions of microalgae in palm oil mill effluent using carbon and nutrient supplementation. Energy Convers. Manag. 2018, 164, 188–197. [Google Scholar] [CrossRef]
- Caprio, F.D.; Andrea, V.; Altimari, P.; Toro, L.; Masciocchi, B.; Iaquaniello, G.; Pagnanelli, F. Two stage process of microalgae cultivation for starch and carotenoid production. Chem. Eng. Trans. 2016, 49, 415–420. [Google Scholar]
- Ra, C.H.; Kang, C.-H.; Kim, N.K.; Lee, C.-G.; Kim, S.-K. Cultivation of four microalgae for biomass and oil production using a two-stage culture strategy with salt stress. Renew. Energy 2015, 80, 117–122. [Google Scholar] [CrossRef]
- Chen, G.; Zhao, L.; Qi, Y. Enhancing the productivity of microalgae cultivated in wastewater toward biofuel production: A critical review. Appl. Energy 2015, 137, 282–291. [Google Scholar] [CrossRef]
- Bora, A.P.; Gupta, D.P.; Durbha, K.S. Sewage sludge to bio-fuel: A review on the sustainable approach of transforming sewage waste to alternative fuel. Fuel 2020, 259, 116262. [Google Scholar] [CrossRef]
- Xiong, W.; Gao, C.; Yan, N.; Wu, C.; Wu, Q. Double CO2 fixation in photosynthesis–fermentation model enhances algal lipid synthesis for biodiesel production. Bioresour. Technol. 2010, 101, 2287–2293. [Google Scholar] [CrossRef]
- Zhou, W.; Hu, B.; Li, Y.; Min, M.; Mohr, M.; Du, Z.; Chen, P.; Ruan, R. Mass Cultivation of Microalgae on Animal Wastewater: A Sequential Two-Stage Cultivation Process for Energy Crop and Omega-3-Rich Animal Feed Production. Appl. Biochem. Biotechnol. 2012, 168, 348–363. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Min, M.; Li, Y.; Hu, B.; Ma, X.; Cheng, Y.; Liu, Y.; Chen, P.; Ruan, R. A hetero-photoautotrophic two-stage cultivation process to improve wastewater nutrient removal and enhance algal lipid accumulation. Bioresour. Technol. 2012, 110, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Hong, Y.; Liu, P.; Zhan, J.; Yan, R. Effects of cultivation strategies on the cultivation of Chlorella sp. HQ in photoreactors. Front. Environ. Sci. Eng. 2019, 13, 78. [Google Scholar] [CrossRef]
- Jagadevan, S.; Banerjee, C.; Banerjee, C.; Guria, C.; Tiwari, R.; Baweja, M.; Shukla, P. Recent developments in synthetic biology and metabolic engineering in microalgae towards biofuel production. Biotechnol. Biofuels 2018, 11, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.-W.; Huang, L.; Ji, P.-Y.; Chen, C.-P.; Li, X.-S.; Gao, Y.-H.; Liang, J.-R. Using a mixture of wastewater and seawater as the growth medium for wastewater treatment and lipid production by the marine diatom Phaeodactylum tricornutum. Bioresour. Technol. 2019, 289, 121681. [Google Scholar] [CrossRef]
- Farooq, W.; Lee, Y.-C.; Ryu, B.-G.; Kim, B.-H.; Kim, H.-S.; Choi, Y.-E.; Yang, J.-W. Two-stage cultivation of two Chlorella sp. strains by simultaneous treatment of brewery wastewater and maximizing lipid productivity. Bioresour. Technol. 2013, 132, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Hena, S.; Fatihah, N.; Tabassum, S.; Ismail, N. Three stage cultivation process of facultative strain of Chlorella sorokiniana for treating dairy farm effluent and lipid enhancement. Water Res. 2015, 80, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Zheng, H.; Addy, M.; Anderson, E.; Liu, Y.; Chen, P.; Ruan, R. Cultivation of Chlorella vulgaris in wastewater with waste glycerol: Strategies for improving nutrients removal and enhancing lipid production. Bioresour. Technol. 2016, 207, 252–261. [Google Scholar] [CrossRef]
- Ge, S.; Qiu, S.; Tremblay, D.; Viner, K.; Champagne, P.; Jessop, P.G. Centrate wastewater treatment with Chlorella vulgaris: Simultaneous enhancement of nutrient removal, biomass and lipid production. Chem. Eng. J. 2018, 342, 310–320. [Google Scholar] [CrossRef]
- Johnson, T.J.; Katuwal, S.; Anderson, G.A.; Gu, L.; Zhou, R.; Gibbons, W.R. Photobioreactor cultivation strategies for microalgae and cyanobacteria. Biotechnol. Prog. 2018, 34, 811–827. [Google Scholar] [CrossRef] [PubMed]
- Sirisuk, P.; Ra, C.-H.; Jeong, G.-T.; Kim, S.-K. Effects of wavelength mixing ratio and photoperiod on microalgal biomass and lipid production in a two-phase culture system using LED illumination. Bioresour. Technol. 2018, 253, 175–181. [Google Scholar] [CrossRef]
- Ramanna, L.; Rawat, I.; Bux, F. Light enhancement strategies improve microalgal biomass productivity. Renew. Sustain. Energy Rev. 2017, 80, 765–773. [Google Scholar] [CrossRef]
- Ra, C.H.; Kang, C.-H.; Jung, J.-H.; Jeong, G.-T.; Kim, S.-K. Enhanced biomass production and lipid accumulation of Picochlorum atomus using light-emitting diodes (LEDs). Bioresour. Technol. 2016, 218, 1279–1283. [Google Scholar] [CrossRef]
- Huerlimann, R.; Denys, R.; Heimann, K. Growth, lipid content, productivity, and fatty acid composition of tropical microalgae for scale-up production. Biotechnol. Bioeng. 2010, 107, 245–257. [Google Scholar] [CrossRef]
- Wang, T.; Tian, X.; Liu, T.; Wang, Z.-J.; Guan, W.; Guo, M.; Chu, J.; Zhuang, Y. A two-stage fed-batch heterotrophic culture of Chlorella protothecoides that combined nitrogen depletion with hyperosmotic stress strategy enhanced lipid yield and productivity. Process. Biochem. 2017, 60, 74–83. [Google Scholar] [CrossRef]
- Yoon, S.Y.; Hong, M.E.; Chang, W.S.; Sim, S.J. Enhanced biodiesel production in Neochloris oleoabundans by a semi-continuous process in two stage photobioreactors. Bioprocess Biosyst. Eng. 2015, 38, 1415–1421. [Google Scholar] [CrossRef]
- Nayak, M.; Suh, W.I.; Cho, J.M.; Kim, H.S.; Lee, B.; Chang, Y.K. Strategic implementation of phosphorus repletion strategy in continuous two-stage cultivation of Chlorella sp. HS2: Evaluation for biofuel applications. J. Environ. Manag. 2020, 271, 111041. [Google Scholar] [CrossRef]
- Ghidossi, T.; Marison, I.; Devery, R.; Gaffney, D.; Forde, C. Characterization and Optimization of a Fermentation Process for the Production of High Cell Densities and Lipids Using Heterotrophic Cultivation of Chlorella protothecoides. Ind. Biotechnol. 2017, 13, 253–259. [Google Scholar] [CrossRef]
- Palabhanvi, B.; Muthuraj, M.; Mukherjee, M.; Kumar, V.; Das, D. Process engineering strategy for high cell density-lipid rich cultivation of Chlorella sp. FC2 IITG via model guided feeding recipe and substrate driven pH control. Algal Res. 2016, 16, 317–329. [Google Scholar] [CrossRef]
- Zheng, Y.; Li, T.; Yu, X.; Bates, P.D.; Dong, T.; Chen, S. High-density fed-batch culture of a thermotolerant microalga Chlorella sorokiniana for biofuel production. Appl. Energy 2013, 108, 281–287. [Google Scholar] [CrossRef]
- Muylaert, K.; Bastiaens, L.; Vandamme, D.; Gouveia, L. Harvesting of microalgae: Overview of process options and their strengths and drawbacks. In Microalgae-Based Biofuels and Bioproducts; Elsevier: Cambridge, UK, 2017; pp. 113–132. [Google Scholar]
- Zhang, B.; Chen, S. Effect of different organic matters on flocculation of Chlorella sorokiniana and optimization of flocculation conditions in swine manure wastewater. Bioresour. Technol. 2015, 192, 774–780. [Google Scholar] [CrossRef] [PubMed]
- Chatsungnoen, T.; Chisti, Y. Oil production by six microalgae: Impact of flocculants and drying on oil recovery from the biomass. Environ. Biol. Fishes 2016, 28, 2697–2705. [Google Scholar] [CrossRef]
- Vandamme, D.; Foubert, I.; Fraeye, I.; Muylaert, K. Influence of organic matter generated by Chlorella vulgaris on five different modes of flocculation. Bioresour. Technol. 2012, 124, 508–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, A.; Shah, R.; Yadav, P.; Verma, R.; Srivastava, S. Harvesting of freshwater microalgae Scenedesmus sp. by electro–coagulation–flocculation for biofuel production: Effects on spent medium recycling and lipid extraction. Environ. Sci. Pollut. Res. 2019, 27, 3497–3507. [Google Scholar] [CrossRef]
- Fayad, N.; Yehya, T.; Audonnet, F.; Vial, C. Harvesting of microalgae Chlorella vulgaris using electro-coagulation-flocculation in the batch mode. Algal Res. 2017, 25, 1–11. [Google Scholar] [CrossRef]
- Mathimani, T.; Mallick, N. A comprehensive review on harvesting of microalgae for biodiesel—Key challenges and future directions. Renew. Sustain. Energy Rev. 2018, 91, 1103–1120. [Google Scholar] [CrossRef]
- Vashist, V.; Chauhan, D.; Bhattacharya, A.; Rai, M.P. Role of silica coated magnetic nanoparticle on cell flocculation, lipid extraction and linoleic acid production from Chlorella pyrenoidosa. Nat. Prod. Res. 2019, 34, 2852–2856. [Google Scholar] [CrossRef]
- Ahmad, A.L.; Yasin, N.M.; Derek, C.; Lim, J. Comparison of harvesting methods for microalgae Chlorella sp. and its potential use as a biodiesel feedstock. Environ. Technol. 2014, 35, 2244–2253. [Google Scholar] [CrossRef]
- Borges, L.; Caldas, S.; D’Oca, M.G.M.; Abreu, P.C. Effect of harvesting processes on the lipid yield and fatty acid profile of the marine microalga Nannochloropsis oculata. Aquac. Rep. 2016, 4, 164–168. [Google Scholar] [CrossRef] [Green Version]
- Yasin, N.H.M.; Shafei, N.I.; Rushan, N.H.; Abu Sepian, N.R.; Said, F.M. The Effect of Microalgae Harvesting on Lipid for Biodiesel Production. Mater. Today Proc. 2019, 19, 1582–1590. [Google Scholar] [CrossRef]
- Coward, T.; Lee, J.; Caldwell, G.S. Harvesting microalgae by CTAB-aided foam flotation increases lipid recovery and improves fatty acid methyl ester characteristics. Biomass Bioenergy 2014, 67, 354–362. [Google Scholar] [CrossRef] [Green Version]
- Magnusson, M.; Mata, L.; Denys, R.; Paul, N. Biomass, Lipid and Fatty Acid Production in Large-Scale Cultures of the Marine Macroalga Derbesia tenuissima (Chlorophyta). Mar. Biotechnol. 2014, 16, 456–464. [Google Scholar] [CrossRef]
- Sekar, M.; Mathimani, T.; Alagumalai, A.; Chi, N.T.L.; Duc, P.A.; Bhatia, S.K.; Brindhadevi, K.; Pugazhendhi, A. A review on the pyrolysis of algal biomass for biochar and bio-oil-Bottlenecks and scope. Fuel 2021, 283, 119190. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | References | Genus and Species | Upstream Approaches | Downstream Approaches | |||
|---|---|---|---|---|---|---|---|
| Strain Selection | Genetic Approach | Cultivation | Harvesting | After Harvesting | |||
| 2012 | Přibyl et al. [23] | C. vulgaris | ✓ | ✕ | ✕ | ✕ | ✕ |
| 2013 | Borowitzk et al. [43] | * | ✓ | ✕ | ✓ | ✕ | ✕ |
| 2013 | Xia et al. [38] | S. obtusus | ✕ | ✓ | ✕ | ✕ | |
| 2013 | Trentacoste et al. [35] | T. pseudonana | ✕ | ✓ | ✕ | ✕ | ✕ |
| 2014 | Ho et al. [19] | * | ✕ | ✓ | ✓ | ✕ | ✕ |
| 2015 | Singh et al. [44] | * | ✕ | ✓ | ✕ | ✕ | ✕ |
| 2016 | Challagulla et al. [26] | * | ✓ | ✕ | ✕ | ✕ | ✕ |
| 2017 | Chen et al. [14] | * | ✕ | ✓ | ✓ | ✕ | ✕ |
| 2017 | Chu [45] | * | ✓ | ✓ | ✕ | ✕ | ✕ |
| 2017 | Kim et al. [25] | * | ✓ | ✕ | ✕ | ✕ | ✕ |
| 2017 | Chung et al. [46] | * | ✕ | ✓ | ✕ | ✕ | ✕ |
| 2018 | Sharma et al. [34] | * | ✕ | ✓ | ✕ | ✕ | ✕ |
| 2018 | Yun et al. [39] | N. oleoabundans | ✕ | ✕ | ✓ | ✕ | ✕ |
| 2018 | Shin et al. [47] | * | ✓ | ✕ | ✓ | ✕ | ✕ |
| 2018 | Sajadi et al. [16] | * | ✓ | ✕ | ✕ | ✕ | ✕ |
| 2019 | Nagappan et al. [40] | * | ✕ | ✕ | ✓ | ✕ | ✕ |
| 2019 | Piligaev et al. [48] | * | ✕ | ✕ | ✓ | ✕ | ✕ |
| 2019 | Park et al. [37] | * | ✕ | ✓ | ✕ | ✕ | ✕ |
| 2019 | Menegazzo et al. [49] | * | ✕ | ✕ | ✕ | ✓ | ✕ |
| 2020 | Nguyen et al. [36] | Chlamydomonas sp. | ✕ | ✓ | ✕ | ✕ | ✕ |
| 2020 | Poh et al. [50] | Chlorella sp. | ✕ | ✕ | ✕ | ✕ | ✓ |
| 2020 | Aziz et al. [41] | * | ✕ | ✕ | ✓ | ✕ | ✕ |
| 2020 | Bhatia et al. [42] | * | ✕ | ✕ | ✓ | ✓ | ✕ |
| Genus and Species | Approach | Comments | Ref. |
|---|---|---|---|
| Several microalgae strains | Inverted fluorescence microscope | Analyzes growth and lipid content | [65] |
| Chlorococcum littorale | Fluorescence assisted cell sorting | detect microalgae cells with high lipid content | [69] |
| Phaeodactylum tricornutum | microfluidic cytometer | measures lipid accumulation and photosynthetic efficiency | [28] |
| Dunaliella salina | Fourier transform infrared spectroscopy | Examines the growth and lipid yield | [29] |
| Chlamydomonas reinhardtii mutants | Droplet microfluidics-based screening | Analyze growth and lipid content in populations | [25] |
| Chlorococcum littorale | Fluorescence assisted microalgae screening | Screen algal cells with high lipid content under nitrogen deficiency | [69] |
| Genus and Species | Approach | Gene | Phenotypic Changes | Ref. |
|---|---|---|---|---|
| Phaeodactylum tricornutum | Suppression | Pyruvate dehydrogenase kinase | Improves lipid (up to 82%)/slight decrease in growth | [80] |
| Fistulifera solaris | Overexpression | Glycerol kinase | Improves lipid/biomass productivities | [81] |
| Chlorella pyrenoidosa | Overexpression | NAD (H) kinase | Enhance in lipid accumulation by 110.4%/without decreasing growth rate | [82] |
| Chlamydomonas reinhardtii | Suppression | Phosphoenol pyruvate carboxylase | Improves lipid content (14–28%) | [83] |
| Phaeodactylum tricornutum | Overexpression | Malic enzyme | Improves lipid content (2.5-fold) but did not affect the growth rate | [79] |
| N. oleoabundans | Overexpression | lysophosphatidyl-acyltransferases | Increase in lipid content (52% and 42%, respectively) without decreasing growth rate | [84] |
| Glycerol-3-phosphate acyltransferase | ||||
| diacylglycerol acyltransferase | ||||
| Chlorella minutissima | Overexpression | Co-expression of five acyltransferases | Improves lipid content (up to 2-fold) but did not affect the growth rate | [59] |
| Species | Transcription Factors | Purpose of Modification | Ref. |
|---|---|---|---|
| C. reinhardtii | Compromised Hydrolysis of Triacylglycerols (CHT7) | Manipulation of CHT7 TF increased lipid productivity | [94] |
| C. reinhardtii | PSR1 | Manipulation of PSR1 TF increased growth rate (two-fold or more) and an increase in lipid content | [95] |
| N. salina | NsbHLH2 | Overexpression of NsbHLH2 increased biomass productivity (509.3 mg L−1)/lipid content (9.96% DW) | [97] |
| N. salina | Basic leucine zipper (bZIP) | Under the N-deprivation conditions, transformants showed an increase of up to 88% and 39% in lipid content and biomass productivity, respectively. | [98] |
| C. ellipsoidea | GmDof4 | Overexpression of GmDof4 increased the lipid content by 52.9% but did not affect the growth rate | [93] |
| D. bardawil | WRKY | WRKY up-regulates carotenogenic genes to increase carotenoid and is important in adaptation to abiotic stress | [99] |
| P. tricornutum | RING-GAF-Gln | RGQ1 is involved in early nitrogen starvation | [100] |
| N. gaditana | Zn(ii)2Cys6 | Doubled the strain’s lipid content without decreasing growth rate | [96] |
| H. pluvialis | Myb | TFs can affect other TFs to enhance astaxanthin/carotene biosynthesis | [101] |
| Microalgae Species | Strategy Adopted | Productivity (mg L−1 day−1) | Dilution Rate | Ref. |
|---|---|---|---|---|
| Choricystis minor | Continuous | 82 | 0.014 h−1 | [106] |
| Chlorella minutissima Dunaliella tertiolecta | Continuous | 1.37 | 0.33 day−1 | [107] |
| 0.91 | 0.42 day−1 | |||
| Chlorella pyrenoidosa | Fed-batch | 1.45 | 388.0 µg L−1 h−1 | [102] |
| Batch | 96.28 | |||
| Neochloris oleoabundans | Continuous | - | - | [108] |
| Chlorella vulgaris | Batch | 0.94 | - | [13] |
| Chlorella sorokiniana | 0.85 | |||
| Neochloris oleoabundans | 1.31 | |||
| Chlorococcum oleofaciens | 0.86 | |||
| Scenedesmus naegleii | 0.83 | |||
| Scenedesmus dimorphus | 1.11 |
| Genus and Species | Lipid Productivity (g L−1 day−1) | Type of Inducer | Ref. |
|---|---|---|---|
| Tetraselmis sp. | 0.285 | Salinity + Nitrogen starvation | [125] |
| Chlorella sorokiniana DPK | 0.690 | Diethyl aminoethyl hexonate(DA-6), Indole-3-acetic acid (IAA) and N starvation | [136] |
| Chlorella sorokiniana | 0.502 0.494 | IAA cytokinin- kinetin (K) | [132] |
| Scenedesmus obtusus XJ-15 | 0.607 | Salinity stress | [38] |
| Nannochloropsis oculata | 0.324 | Salinity stress | [137] |
| Dunaliella salina KSA-HS022 | 0.565 | Salinity stress | [138] |
| Chlamydomonas reinhardtii | 0.109 | Combination of NaCl/CaCl2 | [139] |
| Chlorella vulgaris | 0.800 | Salinity + Nitrogen starvation | [140] |
| Microalgae Strain | Biomass Productivity (g L−1) | Lipid Productivity (mg L−1 day−1) | Metabolism Mode/ Type of Wastewater | Refs. |
|---|---|---|---|---|
| Chlorella vulgaris | 4.9 | 80 | Salinity + Nitrogen starvation/wastewater | [140] |
| Phaeodactylum tricornutum | 54.76 | 54.76 | Mixed municipal wastewater and seawater | [153] |
| Chlorella vulgaris | 226.6 | 108 | two-stage photoautotrophic–photoheterotrophic/mixotrophic mode | [154] |
| Chlorella sorokiniana | >7 | 83.5% | Three-stage cultivation/Farm wastewater | [155] |
| Chlorella vulgaris | 2.92 | 163 | Wastewater + glycerol addition | [156] |
| Chlorella vulgaris | 1.89 ± 0.07 | 24.7 ± 1.2 | Photo-mixotrophic/ Centrate wastewater | [157] |
| Microalga Species | Harvesting Method | FAME Profile | Lipid Yield (%) | Ref. |
|---|---|---|---|---|
| Chlorella sp. | flotation + centrifugation | MUFA increase | 13.4 | [180] |
| Centrifugation | PUFA increase | 9.9 | ||
| Chlorella sp. | Centrifugation | No change | 27 | [178] |
| Microfiltration | No change | 26 | ||
| Coagulation | No change | 15 | ||
| Nannochloropsis oculata | Flocculation with NaOH | The balance between SFA, PUFA, and MUFA | 4.3 | [177] |
| Flocculation with Magnafloc | Lower C20:5 and higher C14:0 and | ~4.4 | ||
| Filtration | The balance between, SFA, PUFA, and MUFA | 3.6 | ||
| Thalassiosira weissflogi | Flocculation with Flopam | C18:0, C18:1n9c, and C16 0, increase | 4.12 | |
| The balance between, SFA, PUFA, and MUFA | ~3.1 | |||
| The balance between MUFA, PUFA, and SFA | 2.77 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shokravi, Z.; Shokravi, H.; Chyuan, O.H.; Lau, W.J.; Koloor, S.S.R.; Petrů, M.; Ismail, A.F. Improving ‘Lipid Productivity’ in Microalgae by Bilateral Enhancement of Biomass and Lipid Contents: A Review. Sustainability 2020, 12, 9083. https://0-doi-org.brum.beds.ac.uk/10.3390/su12219083
Shokravi Z, Shokravi H, Chyuan OH, Lau WJ, Koloor SSR, Petrů M, Ismail AF. Improving ‘Lipid Productivity’ in Microalgae by Bilateral Enhancement of Biomass and Lipid Contents: A Review. Sustainability. 2020; 12(21):9083. https://0-doi-org.brum.beds.ac.uk/10.3390/su12219083
Chicago/Turabian StyleShokravi, Zahra, Hoofar Shokravi, Ong Hwai Chyuan, Woei Jye Lau, Seyed Saeid Rahimian Koloor, Michal Petrů, and Ahmad Fauzi Ismail. 2020. "Improving ‘Lipid Productivity’ in Microalgae by Bilateral Enhancement of Biomass and Lipid Contents: A Review" Sustainability 12, no. 21: 9083. https://0-doi-org.brum.beds.ac.uk/10.3390/su12219083