
Influence of Light Intensity and Photoperiod on the Photoautotrophic Growth and Lipid Content of the Microalgae Verrucodesmus verrucosus in a Photobioreactor

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental
2.2. Algal Sample
2.3. Culture Medium
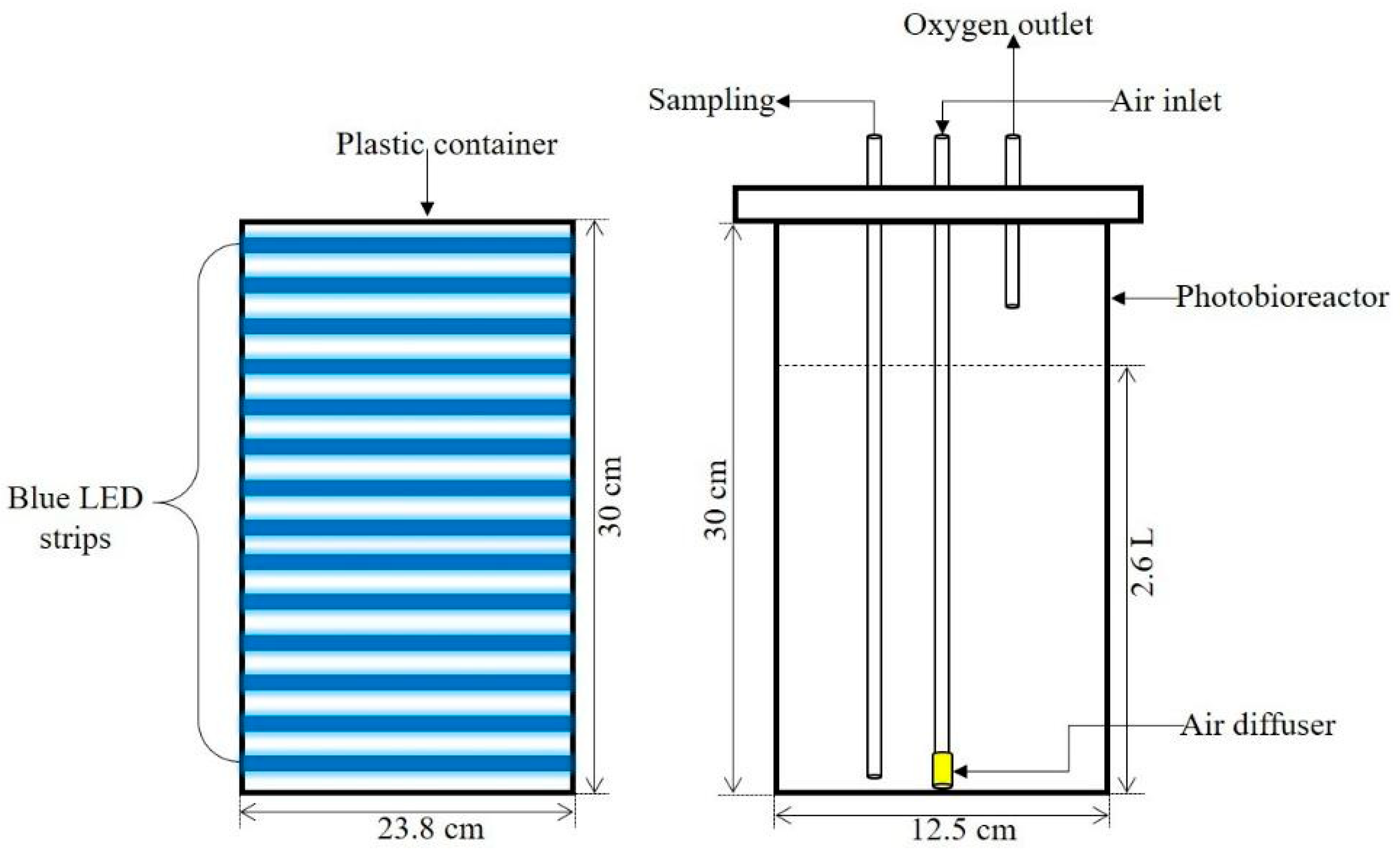
2.4. Photobioreactor Design and Culture Conditions
2.5. Growth Kinetics and Biomass Estimation
- Specific growth rate (μ) was calculated from the equation:
- Where, N2 and N1 were the concentration of the number of cells at times t2 and t1 (t2 was considered the final time and t1 the beginning).
- Doubling time: (Td), the time required to double the number of cells was determined according to the equation:
- For the evaluation percentage of nitrogen consumption by microalgal biomass the total nitrogen (N) was analyzed by Kjeldahl method [23], considering the initial and final concentration.
2.6. Oil Extraction
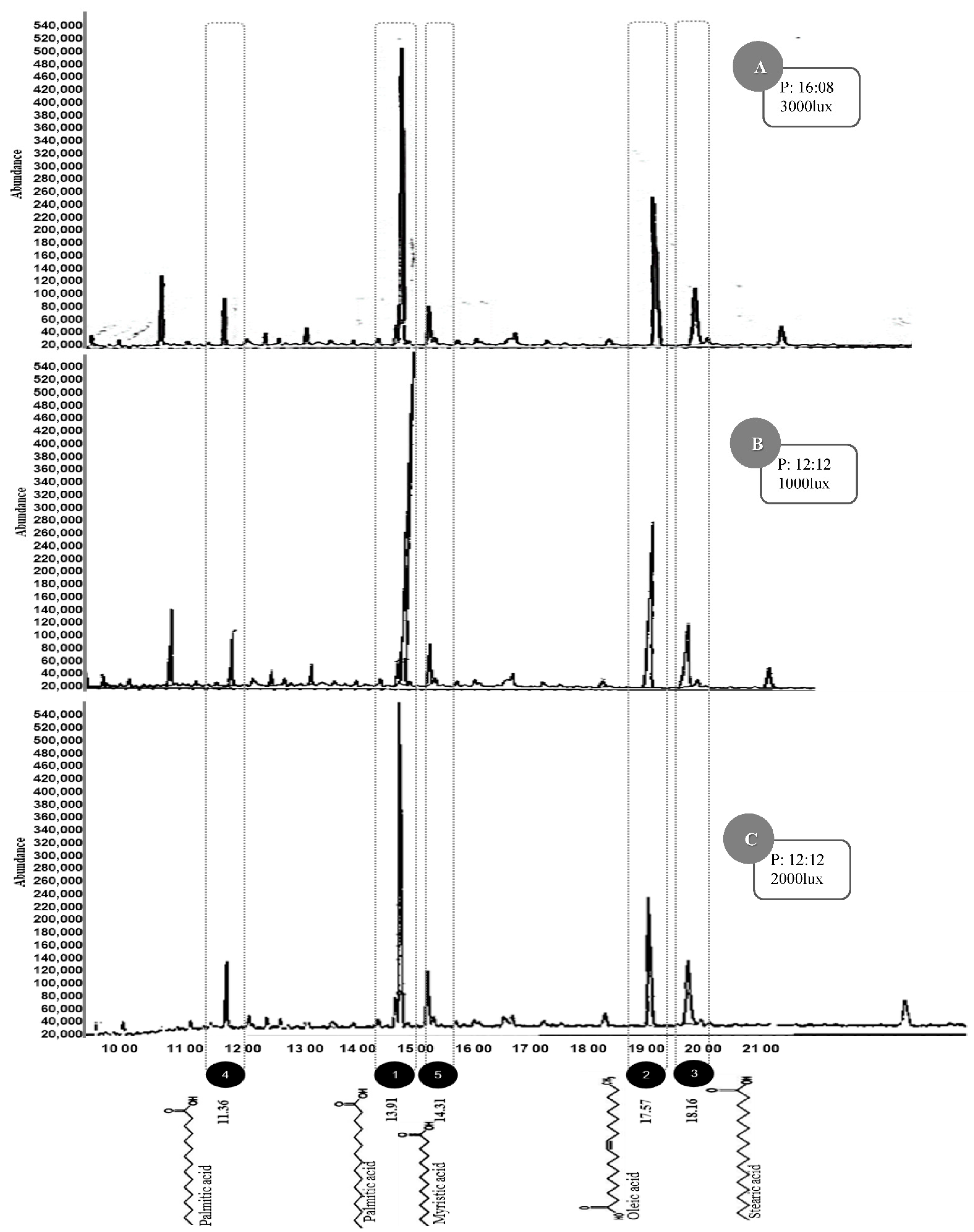
2.7. Fatty Acid Profile of Total Lipids of Verrucodesmus verrucosus
2.8. Statistical Analysis
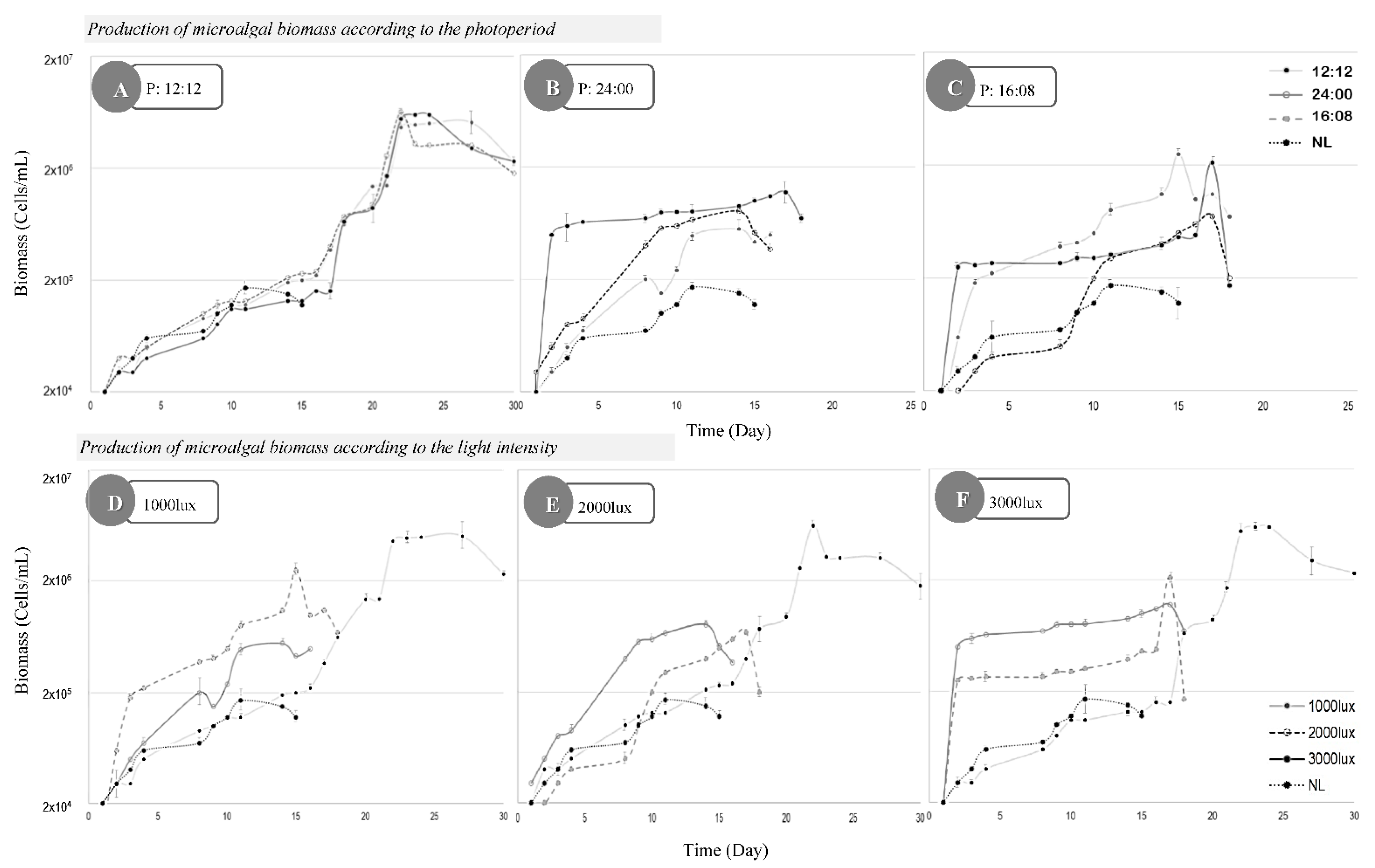
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hossain, N.; Hasan, M.H.; Mahlia, T.M.I.; Shamsuddin, A.H.; Silitonga, A.S. Feasibility of microalgae as feedstock for alternative fuel in Malaysia: A review. Energy Strategy Rev. 2020, 32, 100536. [Google Scholar] [CrossRef]
- Giwa, A.; Adeyemi, I.; Dindi, A.; Lopez, C.G.B.; Lopresto, C.G.; Curcio, S.; Chakraborty, S. Techno-economic assessment of the sustainability of an integrated biorefinery from microalgae and Jatropha: A review and case study. Renew. Sustain. Energy Rev. 2018, 88, 239–257. [Google Scholar] [CrossRef]
- Silitonga, A.S.; Masjuki, H.H.; Ong, H.C.; Mahlia, T.M.I.; Kusumo, F. Optimization of extraction of lipid from Isochrysis galbana microalgae species for biodiesel synthesis. Energy Sources Part A Recovery Util. Environ. Eff. 2017, 39, 1167–1175. [Google Scholar] [CrossRef]
- Knothe, G.; Razon, L.F. Biodiesel fuels. Prog. Energy Combust. Sci. 2017, 58, 36–59. [Google Scholar] [CrossRef]
- Batista, F.R.M.; Lucchesi, K.W.; Carareto, N.D.D.; Costa, M.C.D.; Meirelles, A.J.A. Properties of microalgae oil from the species Chlorella protothecoides and its ethylic biodiesel. Braz. J. Chem. Eng. 2018, 35, 1383–1394. [Google Scholar] [CrossRef] [Green Version]
- Vasistha, S.; Khanra, A.; Clifford, M.; Rai, M.P. Current advances in microalgae harvesting and lipid extraction processes for improved biodiesel production: A review. Renew. Sustain. Energy Rev. 2020, 110498. [Google Scholar]
- Patel, A.; Gami, B.; Patel, P.; Patel, B. Microalgae: Antiquity to era of integrated technology. Renew. Sustain. Energy Rev. 2017, 71, 535–547. [Google Scholar] [CrossRef]
- Zou, X.; Xu, K.; Chang, W.; Qu, Y.; Li, Y. A Novel Microalgal Biofilm Reactor Using Walnut Shell as Substratum for Microalgae Biofilm Cultivation and Lipid Accumulation. Renew. Energy 2021. [Google Scholar] [CrossRef]
- Matos, Â.P.; Cavanholi, M.G.; Moecke, E.H.S.; Sant’Anna, E.S. Effects of different photoperiod and trophic conditions on biomass, protein and lipid production by the marine alga Nannochloropsis gaditana at optimal concentration of desalination concentrate. Bioresour. Technol. 2017, 224, 490–497. [Google Scholar] [CrossRef]
- Ananthi, V.; Brindhadevi, K.; Pugazhendhi, A.; Arun, A. Impact of abiotic factors on biodiesel production by microalgae. Fuel 2021, 284, 118962. [Google Scholar] [CrossRef]
- Kumar, V.; Sharma, N.; Jaiswal, K.K.; Vlaskin, M.S.; Nanda, M.; Tripathi, M.K.; Kumar, S. Microalgae with a truncated light-harvesting antenna to maximize photosynthetic efficiency and biomass productivity: Recent advances and current challenges. Process. Biochem. 2021. [Google Scholar] [CrossRef]
- Wahidin, S.; Idris, A.; Shaleh, S.R.M. The influence of light intensity and photoperiod on the growth and lipid content of microalgae Nannochloropsis sp. Bioresour. Technol. 2013, 129, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Yu, L.; Zhang, Z.; Wang, Y.; Wang, W.; Yang, H.; Zhu, D. Intrinsic kinetic model of photoautotrophic microalgae based on chlorophyll fluorescence analysis. Math. Biosci. 2019, 315, 108234. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, G.; Alam, M.A.; Mofijur, M.; Jahirul, M.I.; Lv, Y.; Xiong, W.; Xu, J. Modern developmental aspects in the field of economical harvesting and biodiesel production from microalgae biomass. Renew. Sustain. Energy Rev. 2021, 135, 110209. [Google Scholar] [CrossRef]
- Pattison, M.; Hansen, M.; Bardsley, N.; Elliott, C.; Lee, K.; Pattison, L.; Tsao, J. Lighting R&D Opportunities (No. DOE/EE-2008); Solid State Lighting Solutions (SSLS) Inc.: Santa Barbara, CA, USA, 2019. [Google Scholar]
- Schulze, P.S.; Guerra, R.; Pereira, H.; Schüler, L.M.; Varela, J.C. Flashing LEDs for microalgal production. Trends Biotechnol. 2017, 35, 1088–1101. [Google Scholar] [CrossRef]
- Lima, S.; Schulze, P.S.; Schüler, L.M.; Rautenberger, R.; Morales-Sánchez, D.; Santos, T.F.; Kiron, V. Flashing light emitting diodes (LEDs) induce proteins, polyunsaturated fatty acids and pigments in three microalgae. J. Biotechnol. 2021, 325, 15–24. [Google Scholar] [CrossRef]
- Hegewald, E.; Bock, C.; Krienitz, L. A phylogenetic study on Scenedesmaceae with the description of a new species of Pectinodesmus and the new genera Verrucodesmus and Chodatodesmus (Chlorophyta, Chlorophyceae). Fottea 2013, 13, 14. [Google Scholar] [CrossRef]
- Prescott, G.W. Algae of the Western Great Lakes Area; WC Brown Company: Dubuque, IA, USA, 1962; Volume 31. [Google Scholar]
- Wehr, J.D. Freshwater habitats of algae. Freshw. Algae N. Am. Ecol. Classif. 2003, 11–57. [Google Scholar] [CrossRef]
- Rippka, R.; Deruelles, J.; Waterbury, J.B.; Herdman, M.; Stanier, R.Y. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. Microbiology 1979, 111, 1–61. [Google Scholar] [CrossRef] [Green Version]
- Contreras-Pool, P.Y.; Peraza-Echeverria, S.; Ku-González, Á.F.; Herrera-Valencia, V.A. The phytohormone abscisic acid increases triacylglycerol content in the green microalga Chlorella saccharophila (Chlorophyta). Algae 2016, 31, 267–276. [Google Scholar] [CrossRef]
- Xu, Y.; Ibrahim, I.M.; Harvey, P.J. The influence of photoperiod and light intensity on the growth and photosynthesis of Dunaliella salina (chlorophyta) CCAP 19/30. Plant Physiol. Biochem. 2016, 106, 305–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soto-León, S.; Zazueta-Patrón, I.E.; Piña-Valdez, P.; Nieves-Soto, M.; Reyes-Moreno, C.; Contreras-Andrade, I. Extracción de lípidos de Tetraselmis suecica: Proceso asistido por ultrasonido y solventes. Rev. Mex. Ing. Quími. 2014, 13, 723–737. [Google Scholar]
- Härtig, C. Rapid identification of fatty acid methyl esters using a multidimensional gas chromatography–mass spectrometry database. J. Chromatogr. A 2008, 1177, 159–169. [Google Scholar] [CrossRef]
- Sun, H.; Zhao, W.; Mao, X.; Li, Y.; Wu, T.; Chen, F. High-value biomass from microalgae production platforms: Strategies and progress based on carbon metabolism and energy conversion. Biotechnol. Biofuels 2018, 11, 1–23. [Google Scholar]
- Kato, Y.; Fujihara, Y.; Vavricka, C.J.; Chang, J.S.; Hasunuma, T.; Kondo, A. Light/dark cycling causes delayed lipid accumulation and increased photoperiod-based biomass yield by altering metabolic flux in oleaginous Chlamydomonas sp. Biotechnol. Biofuels 2019, 12, 1–11. [Google Scholar] [CrossRef]
- Fathurrahman, L.; Hajar, A.H.; Sakinah, D.W.; Nurhazwani, Z.; Ahmad, J. Flashing light as growth stimulant in cultivation of green microalgae, Chlorella sp. utilizing airlift photobioreactor. Pak. J. Biol. Sci. 2013, 16, 1517–1523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Gao, L.; Zhou, W.; Liu, T. Growth and palmitoleic acid accumulation of filamentous oleaginous microalgae Tribonema minus at varying temperatures and light regimes. Bioproc. Biosyst. Eng. 2016, 39, 1589–1595. [Google Scholar] [CrossRef]
- Sirisuk, P.; Ra, C.H.; Jeong, G.T.; Kim, S.K. Effects of wavelength mixing ratio and photoperiod on microalgal biomass and lipid production in a two-phase culture system using LED illumination. Bioresour. Technol. 2018, 253, 175–181. [Google Scholar] [CrossRef]
- Elango, R.K.; Sathiasivan, K.; Muthukumaran, C.; Thangavelu, V.; Rajesh, M.; Tamilarasan, K. Transesterification of castor oil for biodiesel production: Process optimization and characterization. Microchem. J. 2019, 145, 1162–1168. [Google Scholar] [CrossRef]
- Lehmuskero, A.; Chauton, M.S.; Boström, T. Light and photosynthetic microalgae: A review of cellular-and molecular-scale optical processes. Prog. Oceanogr. 2018, 168, 43–56. [Google Scholar] [CrossRef] [Green Version]
- Keil, T.; Dittrich, B.; Rührer, J.; Morschett, H.; Lattermann, C.; Möller, M.; Büchs, J. Polymer-based ammonium-limited fed-batch cultivation in shake flasks improves lipid productivity of the microalga Chlorella vulgaris. Bioresour. Technol. 2019, 291, 121821. [Google Scholar] [CrossRef]
- Mutaf, T.; Oz, Y.; Kose, A.; Elibol, M.; Oncel, S.S. The effect of medium and light wavelength towards Stichococcus bacillaris fatty acid production and composition. Bioresour. Technol. 2019, 289, 121732. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Roque, Y.; Luna, Y.P.; Acosta, J.M.; Vázquez, N.F.; Sebastian, J.P.; Hernández, R.B. Optimization for the production of verrucodesmus verrucosus biomass through crops in autotrophic and mixotrophic conditions with potential for the production of biodiesel. Rev. Mex. Ing. Quím. 2020, 19, 133–147. [Google Scholar] [CrossRef] [Green Version]
- Xin, L.; Hong-Ying, H.; Ke, G.; Ying-Xue, S. Effects of different nitrogen and phosphorus concentrations on the growth, nutrient uptake, and lipid accumulation of a freshwater microalga Scenedesmus sp. Bioresour. Technol. 2010, 101, 5494–5500. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Sharma, A.K.; Daniell, H.; Kumar, S. Altered lipid composition and enhanced lipid production in green microalga by introduction of brassica diacylglycerol acyltransferase 2. Plant Biotechnol. J. 2015, 13, 540–550. [Google Scholar] [CrossRef] [Green Version]
- Sandouqa, A.; Al-Hamamre, Z. Energy analysis of biodiesel production from jojoba seed oil. Renew. Energ. 2019, 130, 831–842. [Google Scholar] [CrossRef]
- Chen, J.; Li, J.; Dong, W.; Zhang, X.; Tyagi, R.D.; Drogui, P.; Surampalli, R.Y. The potential of microalgae in biodiesel production. Renew. Sustain. Energy Rev. 2018, 90, 336–346. [Google Scholar] [CrossRef]
- Seffati, K.; Honarvar, B.; Esmaeili, H.; Esfandiari, N. Enhanced biodiesel production from chicken fat using CaO/CuFe2O4 nanocatalyst and its combination with diesel to improve fuel properties. Fuel 2019, 235, 1238–1244. [Google Scholar] [CrossRef]
- Xue, Z.; Yu, Y.; Yu, W.; Gao, X.; Zhang, Y.; Kou, X. Development prospect and preparation technology of edible oil from microalgae. Front. Mar. Sci. 2020, 7, 402. [Google Scholar] [CrossRef]
- Tan, X.B.; Yang, L.B.; Zhang, W.W.; Zhao, X.C. Lipids production and nutrients recycling by microalgae mixotrophic culture in anaerobic digestate of sludge using wasted organics as carbon source. Bioresour. Technol. 2020, 297, 122379. [Google Scholar] [CrossRef]
- Sadaf, S.; Iqbal, J.; Ullah, I.; Bhatti, H.N.; Nouren, S.; Nisar, J.; Iqbal, M. Biodiesel production from waste cooking oil: An efficient technique to convert waste into biodiesel. Sustain. Cities Soc. 2018, 41, 220–226. [Google Scholar]
- Atmanli, A. Experimental comparison of biodiesel production performance of two different microalgae. Fuel 2020, 278, 118311. [Google Scholar] [CrossRef]
- Veljković, V.B.; Biberdžić, M.O.; Banković-Ilić, I.B.; Djalović, I.G.; Tasić, M.B.; Nježić, Z.B.; Stamenković, O.S. Biodiesel production from corn oil: A review. Renew. Sustain. Energ. Rev. 2018, 91, 531–548. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| BG-11 Medium for Blue Green Algae | Trace Metal Mix A5 | ||
|---|---|---|---|
| NaNO3 | 1.5 g | H3BO3 | 2.86 g |
| K2HPO4·3H2O | 4 g | MnCl2·4H2O | 1.81 g |
| K2HPO4 | 3.05 g | MoO4 | 0.018 g |
| MgSO4·7H2O | 7.5 g | ZnSO4·7H2O | 0.222 g |
| CaCl2·2H2O | 3.6 g | NaMoO4·2H2O | 0.39 g |
| HOC(COOH)(CH2COOH)2·H2O | 0.6 g | CuSO4·5H2O | 0.05 g |
| (NH4)5[Fe(C6H4O7)2] | 0.6 g | COCl2·6H2O | 49.4 mg |
| Na2Mg EDTA | 0.1 g | Distilled water | 1.0 L |
| Na2CO3 | 0.02 g | ||
| Trace metal mix A5 | 1.0 mL | ||
| Distilled water | 1.0 L | ||
| Light Intensity (Lux) | Photoperiod (Cycle L:D) | Initial Biomass (Cells · mL−1) | Final Biomass (Cells · mL−1) | µ (Day−1) | Td (Day−1) | Lipids (%) | Nitrogen Consumption (%) |
|---|---|---|---|---|---|---|---|
| 1000 | 16:08 | 2 × 104 ± 0.12 ª * | 2.50 × 106 ± 0.14 b | 1.2 ± 0.10 ª | 0.62 ± 0.06 b | 13.61 ± 0.16 c | 37 ± 0.08 c |
| 12:12 | 2 × 104 ± 0.10 ª | 5.10 × 106 ± 0.06 ª | 0.50 ± 0.09 b | 1.39 ± 0.20 ª | 42.41 ± 0.11 a | 55 ± 0.11 b | |
| 24:00 | 2 × 104 ± 0.21 ª | 5.60 × 106 ± 0.01 ª | 0.84 ± 0.10 b | 0.83 ± 0.10 ª | 38.21 ± 0.10 b | 57 ± 0.07 b | |
| 2000 | 16:08 | 2 × 104 ± 0.17 ª | 7.00 × 105 ± 0.02 b | 0.70 ± 0.00 a | 0.99 ± 0.00 b | 37.35 ± 0.09 c | 19 ± 0.02 d |
| 12:12 | 2 × 104 ± 0.14 ª | 6.30 × 106 ± 0.11 ª | 0.60 ± 0.20 b | 1.24 ± 0.40 ª | 50.42 ± 0.07 ª | 66 ± 0.13 a | |
| 24:00 | 2 × 104 ± 0.11 ª | 8.10 × 105 ± 0.13 b | 0.97 ± 0.00 a | 0.72 ± 0.00 c | 43.09 ± 0.6 b | 21 ± 0.21 d | |
| 3000 | 16:08 | 2 × 104 ± 0.07 ª | 4.80 × 105 ± 0.12 b | 0.82 ± 0.10 ª | 0.85 ± 0.10 b | 41.32 ± 0.12 ª | 17 ± 0.16 d |
| 12:12 | 2 × 104 ± 0.12 ª | 6.00 × 106 ± 0.09 ª | 0.58 ± 0.09 b | 1.21 ± 0.20 ª | 29.29 ± 0.06 c | 62 ± 0.12 a | |
| 24:00 | 2 × 104 ± 0.18 ª | 1.20 × 106 ± 0.04 c | 0.60 ± 0.00 b | 1.15 ± 0.00 a | 37.48 ± 0.08 b | 33 ± 0.09 c | |
| Control | 2 × 104 ± 0.21 ª | 1.70 × 105 ± 0.05 c | 0.61 + 0.00 b | 1.14 + 0.02 a | 24.31 ± 0.13 c | 17.5 ± 0.03 d | |
| Compounds | Chemical Formula | Parent Ion (m/z) 2 | RT 1 (min) | Abundance Percentage (%) | Fatty Acid | ||
|---|---|---|---|---|---|---|---|
| p = 12:12 2000 Lux | p = 12:12 1000 Lux | p = 16:08 3000 Lux | |||||
 | C16H32O2 | 256.43 | 13.91 | 44.90 ± 0.05 b | 55.21 ± 0.12 a | 42.03 ± 0.01 b | Saturated |
 | C18H34O2 | 282.47 | 17.57 | 25.81 ± 0.11 b | 30.01 ± 0.10 b | 28.05 ± 0.05 b | Monounsaturated |
 | C18H36O2 | 284.48 | 18.16 | 9.81 ± 0.15 a | 9.55 ± 0.08 a | 9.76 ± 0.19 a | Saturated |
 | C16H30O2 | 254.41 | 11.36 | 7.87 ± 0.22 a | 7.00 ± 0.12 a | 5.50 ± 0.21 a | Monounsaturated |
 | C14H28O2 | 228.36 | 14.31 | 5.93 ± 0.07 a | 4.95 ± 0.02 a | 4.35 ± 017 a | Saturated |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vélez-Landa, L.; Hernández-De León, H.R.; Pérez-Luna, Y.D.C.; Velázquez-Trujillo, S.; Moreira-Acosta, J.; Berrones-Hernández, R.; Sánchez-Roque, Y. Influence of Light Intensity and Photoperiod on the Photoautotrophic Growth and Lipid Content of the Microalgae Verrucodesmus verrucosus in a Photobioreactor. Sustainability 2021, 13, 6606. https://0-doi-org.brum.beds.ac.uk/10.3390/su13126606
Vélez-Landa L, Hernández-De León HR, Pérez-Luna YDC, Velázquez-Trujillo S, Moreira-Acosta J, Berrones-Hernández R, Sánchez-Roque Y. Influence of Light Intensity and Photoperiod on the Photoautotrophic Growth and Lipid Content of the Microalgae Verrucodesmus verrucosus in a Photobioreactor. Sustainability. 2021; 13(12):6606. https://0-doi-org.brum.beds.ac.uk/10.3390/su13126606
Chicago/Turabian StyleVélez-Landa, Laura, Héctor Ricardo Hernández-De León, Yolanda Del Carmen Pérez-Luna, Sabino Velázquez-Trujillo, Joel Moreira-Acosta, Roberto Berrones-Hernández, and Yazmin Sánchez-Roque. 2021. "Influence of Light Intensity and Photoperiod on the Photoautotrophic Growth and Lipid Content of the Microalgae Verrucodesmus verrucosus in a Photobioreactor" Sustainability 13, no. 12: 6606. https://0-doi-org.brum.beds.ac.uk/10.3390/su13126606