1. Introduction
Given the context of a circular economy and the growing environmental protection concerns, all industries must move towards the minimal generation of waste flows that have the potential to harm the environment. The dairy industry is not an exception to this trend, as it produces large amounts of wastes consisting of whey, sludge and wastewaters from dairy product production that are a challenge to dispose of properly. These waste flows consist mainly of water with various amounts of organic and inorganic components, such as suspended and dissolved solids, lactose, nutrients, fats, sulphates and chlorides, with high biological and chemical oxygen demand [
1]. One of the main waste streams from cheese production is whey, representing 85–95% of the milk volume and retaining up to 55% of its nutrients (lactose—up to 5%, soluble proteins, lipids). This waste can be acidic or sweet, depending on the type of dairy product that is being processed [
2,
3]. Discharging this waste stream into natural water bodies can harm the environment, mainly due to its high organic load that can cause a rapid depletion of the dissolved oxygen that is essential to aquatic life, but also due to the other contaminants that it contains [
4].
Although whey is considered a waste stream, by taking into account the principles of a circular economy, a simple analysis of this stream highlights its potential to serve as feedstock for the further valorization of its components. One possibility for whey wastewater management and the simultaneous valorization of the components that are present in its composition is the cultivation of microalgae on this side-stream that is rich in nutrients. Microalgae are a complex group of microorganisms that fix CO
2 from the environment in their development process and they require only the minimal addition of nutrients in the growth medium [
5]. Microalgae biomass presents great potential for commercial valorization, being suitable for various sectors including nutritional, nutraceutical, animal feed, biofuels or the dye industry. If the aim is to produce vast amounts of microalgae biomass for these industries, then it is necessary to make the process more cost effective as it involves large volumes of water and the costly addition of nutrients. Growing microalgae on industrial waste streams achieves a double benefit, as this process produces valuable microalgae biomass and, at the same time, reduces the organic load of the waste stream that is used as the growth medium. One drawback of growing microalgae biomass on waste streams is their potential contamination with heavy metals or toxic compounds, especially when treating municipal or industrial waste streams. This limits the application range of the biomass to non-food or feed products. However, when using waste streams from the food industry, this is no longer an issue as these compounds are not present and the potential uses of the biomass that is obtained increase in value [
6].
As is the case for most waste streams, in order to use microalgae for their treatment it is required to dilute these streams a number of times so that the microalgae strain can adapt to these new and harsh conditions. For reducing the dilution degree, Marazzi et al. [
7] tested the potential of spontaneous cyanobacteria—microalgae consortia for treating milk whey wastewater, with high decreases in contaminant values, until the possibility of the effluent being discharged into surface waters was realized. Numerous studies have reported that biomass productivity increases with the use of whey wastewaters, for various sweet- and saltwater microalgae strains, at the same time reducing considerably the chemical oxygen demand (COD) and nitrogen values [
8]. Hemalatha et al. [
9] studied the growth of microalgae on dairy wastewaters in an open pond, achieving a 90% reduction efficiency for COD and a nitrate and phosphate reduction efficiency of 65.5/73%. Pereira et al. [
10] evaluated
Spirulina platensis biomass production, biochemical composition and antioxidant capacity when it was grown in autotrophic and mixotrophic conditions in a growth medium with various amounts of cheese whey. The maximum biomass concentration (2.98 g∙L
−1) was obtained with 5% whey, with an increase of 70% and 76% in the production of biomass and carbohydrates, respectively, compared to the control sample. Pandey et al. [
11] studied
Chlorella pyrenoidosa development on a medium containing whey and they obtained a biomass and lipid daily productivity of 230–270 mg∙L
−1 and 58.84–77.41 mg∙L
−1, respectively, corresponding to a 2.95–3.52-fold and 3.63–4.77-fold increase over the same strain of microalgae when it was grown in its specific medium. A study that was carried out by Patel et al. [
12] concluded that pretreated whey was a viable substrate for the growth of
Chlorella protothecoides, with organic and inorganic pollutants being removed almost entirely after 9 days of microalgae cultivation. The algae was also shown to be able to improve the carbon footprint and carbon and energy recoveries for the dairy industry.
Porphyridium purpureum is a red marine unicellular microalga that is widely studied due to its ability to accumulate high-value compounds (phycobiliproteins, polyunsaturated fatty acids and intracellular- and exopolysaccharides) and its adaptability to stress factors in the growth medium. A characteristic that is specific to
Porphyridium purpureum is the lack of a rigid cellulose microfibrillar component in the cell wall, it is instead encapsulated in a sulphated polysaccharide mucilage, which is constantly dissolved in the growth medium. While these exopolysaccharides play a protective role during cell growth, their production rates and, in some cases, their composition change in response to variations in the environmental factors, such as the nutrient composition, light intensity, temperature, etc.
Porphyridium-derived exopolysaccharides have been shown to have immunostimulatory effects, finding applications in cosmetics, growth promoters in agriculture and the health food market [
13]. Phycoerythrin, the main photosynthetic pigment that is present in this species of microalgae, has found a commercial application as a fluorescent dye in pigments and health care products, exhibiting activity as an anti-oxidant and for immunity enhancement. While currently derived from red macroalgae, extracting this pigment from
Porphyridium purpureum would increase the added value of growing this strain of microalgae and offset some of the costs [
14].
Porphyridium purpureum has already proven effective in wastewater treatment more recently in a study by Arashiro et al. (2020) that was carried out with wastewater from a plant-derived food company [
15]; coupling this information with the high-value compounds that are accumulated during its growth, it is clear that this strain of microalgae is a promising candidate for water reclamation from wastewaters that are derived from the dairy industry.
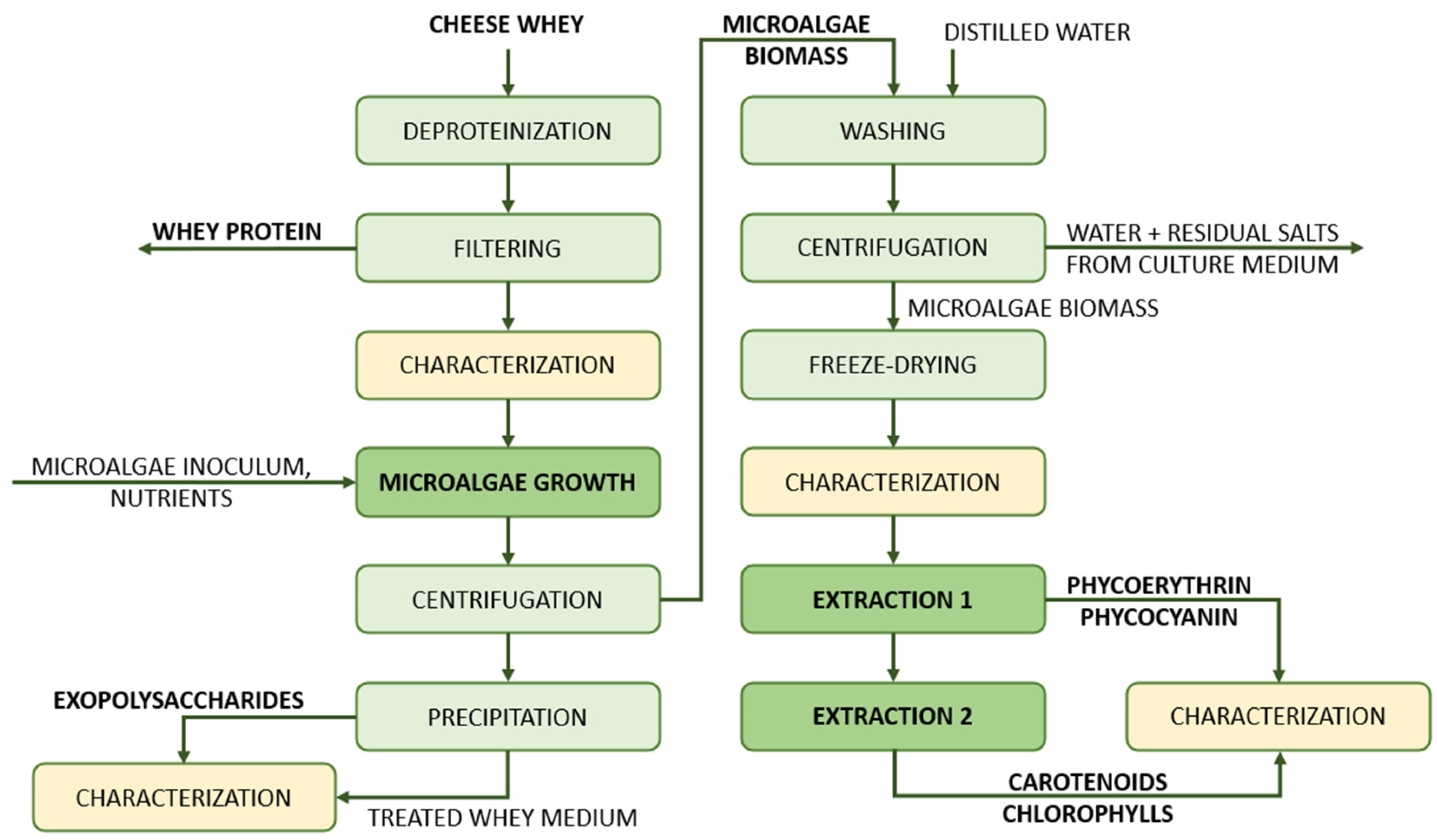
The purpose of this work is to demonstrate the efficient reduction of both the organic and inorganic pollutants in cheese whey by means of the cultivation of the microalgae strain
Porphyridium purpureum. From our knowledge, this is the first study describing the possibility of using the microalgae
Porphyridium purpureum to treat dairy wastewater. Another innovative aspect is represented by the use of the two illumination scenarios—continuous illumination and a day–night cycle—in the treatment process of the dairy wastewater. Also, we aim to prove that, simultaneous to the purification of this wastewater by its lactose content reduction, the microalgae strain consumes and uses the lactose and other nutrients as building blocks for the accumulation of valuable bioactive compounds such as chlorophylls, carotenoids and phycobiliproteins. This process adds value to an otherwise waste stream that requires pricy disposal processes. The commercial viability of such a process is possible, but several factors must be taken into consideration for the scaling-up, such as the optimization of the inoculum preparation, uniformity of the distribution of the nutrients and inoculum, bioreactor configuration and the biomass and lipid yields’ optimization. The flow scheme of the experimental process that was employed in the present study is presented in
Figure 1. This describes the step-by-step procedures that were carried out, from the cheese whey pretreatment to the characterization of the compounds that were extracted from microalgal biomass.
2. Materials and Methods
2.1. Microalgae Strain
The strain of microalgae Porphyridium purpureum was chosen for these experiments. This strain is available as part of INCDCP-ICECHIM’s private microalgae strain collection, it is preserved and replicated constantly to maintain its integrity.
2.2. Cheese Whey Pretreatment and Characterization
The cheese whey was obtained from a dairy farm in Romania. The cheese whey that was used for the sample preparation was initially deproteinized, by precipitation, so as not to inhibit the microalgal growth process. This process was performed by boiling the cheese whey solution for 15 min, followed by cooling it in order to facilitate precipitation, and then the centrifugation and filtration of the suspended solids. The determination of the lactose in the diluted whey samples, both initially and after the microalgae’s growth process, was determined using a UV-spectrophotometer (Ultra 3600 RIGOL, Romspectra, Bucharest, Romania)-based dinitrosalicylic acid (DNSA) assay for reducing the sugars, performed at 540 nm [
16].
For determining the initial lactose concentration, 0.2 mL of the sample solution was diluted up to 2 mL, followed by addition of 1 mL of DNSA reagent. For determining the final lactose concentration, 2 mL of the sample solution was taken, followed by the addition of 1 mL of DNSA reagent. The mixture was heated in hot water bath at 95 °C for 5 min and then immediately cooled using cold water. The solution was analyzed using UV spectrophotometry that was performed at 540 nm.
The pH was measured by a Consort C931 (Consort, Turnhout, Belgium) instrument. The total phosphorus (TP) content was determined using an adaptation of the standard methods for the examination of water and wastewater, edition 20, 4500-P C, vanado-molybdophosphoric acid method (total phosphorus determination kit from Hanna Instruments, Woonsocket, RI, USA) and the total nitrogen (TN) content was determined using a chromotropic acid method (Hanna Instruments total nitrogen determination kit). The aluminum content in the supernatant was determined using an adaptation of the method with aluminum (total nitrogen determination kit from Hanna Instruments). The chemical oxygen demand (COD) was determined using an adaptation of the methods that were developed by the US Environmental Protection Agency (EPA 410.4/1993) and approved for water and surface waste water COD determination (Hanna Instruments chemical oxygen demand determination kit).
2.3. Microalgae Growth Medium and Inoculation
The experiments were carried out using a modified medium that was based on the optimum growth medium of artificial sea water (ASW), containing the following components dissolved in distilled water: 1 g∙L−1 KNO3, 0.076 g∙L−1 KH2PO4, 0.54 g∙L−1 NaHCO3, 15 g∙L−1 NaCl, 3.055 g∙L−1 MgSO4 × 7 H2O, 2.8 g∙L−1 MgCl2 × 6 H2O and 0.75 g∙L−1 CaCl2 × 2 H2O. This was then submitted to sterilization at 121 °C for 15 min. After sterilization, 1 mL∙L−1 of Microelement solution was added (containing 610 mg H3BO3, 380 mg (NH4)6Mo7O24·4 H2O, 60 mg CuSO4·5 H2O, CoCl2·6 H2O, 51 mg; ZnCl2, 4.1 mg; MnCl2·4 H2O and 4.1 mg per 1 L:), as was 1 mL∙L−1 of Fe chelate solution (containing Na2EDTA, 189 mg; FeCl3·6 H2O and 244 mg per 1 L).
The growth medium samples were prepared from the stock deproteinized whey solution by dilution in the ASW optimum growth medium, with the addition of increasing concentrations of cheese whey, calculated as the amount of lactose that was being added (0, 2.5, 5, 7.5, 10 g∙L−1). The culture growth parameters were as follows: high-light illumination (240 μmol photons m−2∙s−1)—continuous lighting vs. a day–night cycle, for a duration of 7 days, at a temperature of 25 °C, with 100 rpm agitation in an Innova 42R incubator (New Brunswick, Eppendorf, Germany) with a home-made illumination system incorporated.
The inoculation was made using 10% (v/v) stationary phase microalgae culture in the growth medium. The total volume of each sample was 200 mL, 10% of which was represented by the inoculum. The microalgae inoculum suspension was prepared and stored refrigerated so as to have a uniform batch for all of the experiments. The samples were prepared in Erlenmeyer flasks that were equipped with sterile cotton caps so as to prevent contamination but to allow air to penetrate inside the flasks in order to promote the growth of the microalgae biomass. The growth medium and the inoculum volume were added to the growth medium, taking into account that we needed to shake the microalgae suspension flask so as to collect a uniform sample. The experiments were performed in triplicate.
2.4. Biomass Productivity
In order to determine the biomass’ productivity, the samples were centrifuged at 7000 rpm for 15 min and the supernatant was separated and collected for polysaccharide precipitation while the biomass was washed with distilled water in order to remove the remaining salts from the growth medium. After removing the water, again via centrifugation, the wet biomass samples were frozen and lyophilized in order to remove any existing moisture. The dry samples were weighed and the biomass’ productivity was calculated by referring to the initial volume of the growth medium. Samples from the supernatant were also taken and analyzed via spectrophotometry in order to determine the concentration of the lactose after cultivation.
2.5. Exopolysaccharide Quantification and Characterization
The exopolysaccharides (EPS) were separated from the growth medium after microalgae separation via precipitation with 96% ethanol on an ice bath. The samples were initially concentrated via boiling until the volume was halved. The purpose of concentrating the samples was to reduce the required ethanol as well as to help to induce precipitation. The exopolysaccharides were isolated via centrifugation, purified by washing with ethanol and oven dried at 105 °C.
Thermogravimetry (TGA) of the EPS was carried out on a Q 600 analyzer (TA Instruments) using nitrogen as the purge gas at a flow rate of 50 mL min−1. The samples (6–7.5 mg) were packed in aluminum pans and tested from the ambient temperature to 750 °C at a heating rate of 10 °C min−1.
2.6. Bioactive Compounds Extraction, Identification and Quantification
Biomass samples were collected and subjected to 3 successive extraction steps, the first 2 with a sodium phosphate buffer solution with a pH value of 6.8 and the third with methanol. For each extraction, the dry biomass to solvent ratio that was used was 0.03 g to 2 mL. The extractions were carried out overnight, at 4 °C, under stirring at 150 rpm, and this was followed by the removal of the supernatant via centrifugation and pipetting.
The extracts were analyzed through spectrophotometry; the absorption spectra were measured between 400 and 800 nm. The phycoerythrin (PE) and phycocyanin (PC) concentrations were determined from the first two extracts in the buffer solution, while the carotenoids and chlorophyll a and b were determined from the last extract in methanol. The quantification of the phycoerythrin and phycocyanin was done using Equations (1) and (2), determined by Beer and Eshel [
17,
18], which remain the preferred methods as can be seen in the more recent work of Pereira et al. [
17,
18].
The maximum absorbances of the phycoerythrin and phycocyanin were found at 564 nm and 618 nm, respectively, while the absorbances for baseline correction were found at 592 nm, 455 nm and 645 nm. The absorption coefficients for each compound are 0.12 and 0.15, while the correction constants are 0.2 and 0.5.
The concentrations of chlorophyll a and b, as well as that of carotenoids, were determined using Equations (3)–(5), published in the work of Xiong et al. [
19], based on the spectrophotometric analysis of the methanol extracts which were obtained using a similar extraction procedure as that which was described for phycoerythrin and phycocyanin.
3. Results and Discussion
3.1. Microalgae Growth Medium Preparation and Characterization
The analysis of the cheese whey, after the deproteinization process, is presented in
Table 1 and it indicated a lactose concentration of 48.9 g∙L
−1.
The samples were prepared as presented in
Table 2. The amount of whey that was added is calculated as the lactose concentration in the final volume of the sample.
The samples that were prepared with different concentrations of lactose are presented in
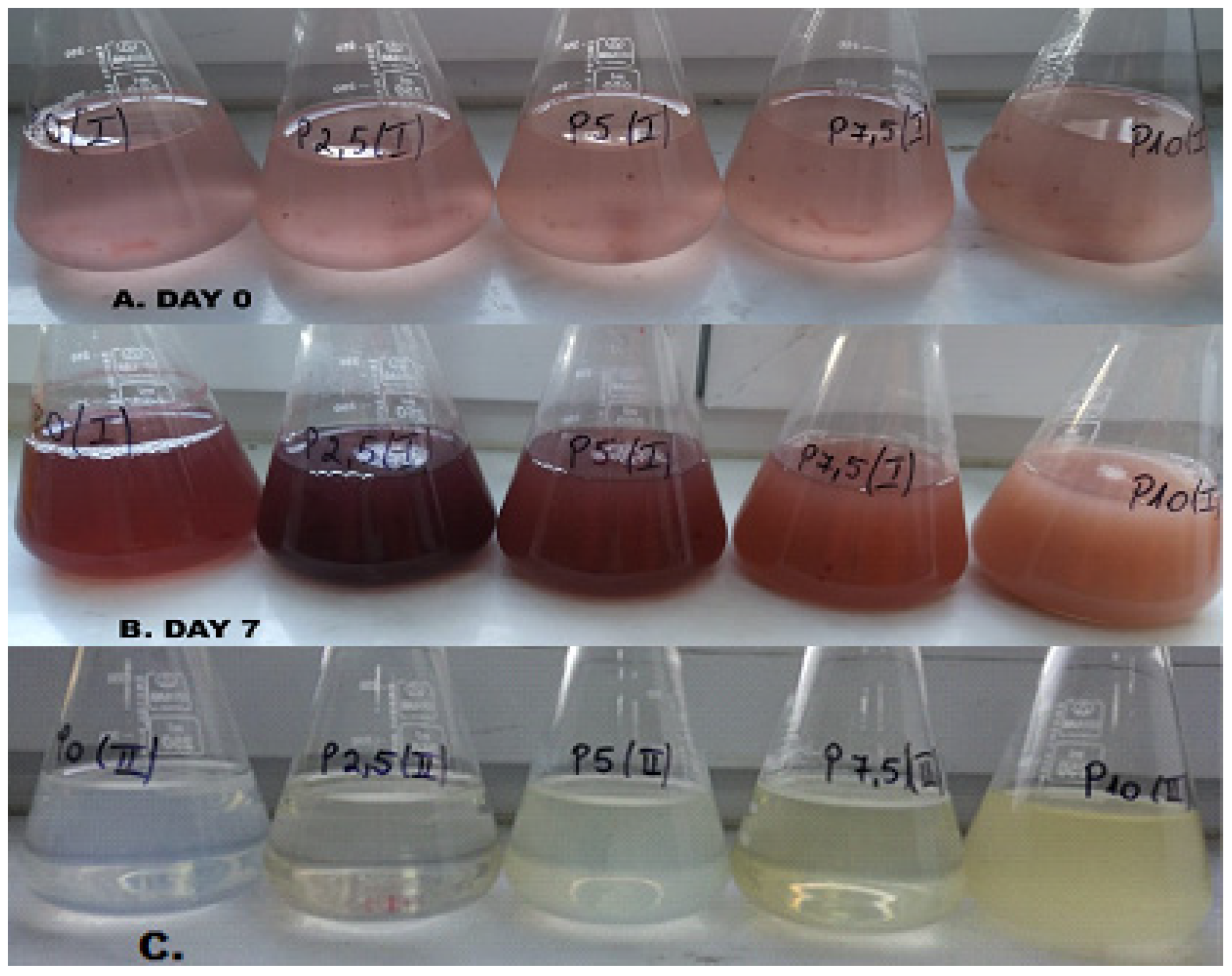
Figure 2A. A blank sample was also prepared, containing no lactose—only the ASW specific medium, so as to be able to highlight the effects of lactose being present in the growth medium. After 7 days of incubation at 25 °C, 100 rpm and continuous illumination (light stress), the microalgae suspension is presented in
Figure 2B. The samples that were grown under a day–night cycle present a very similar appearance. A primary visual analysis of the sample indicates that the microalgae strain has produced considerable amounts of exopolysaccharides as a result of the light stress that was imposed by continuous illumination. This excretion of polysaccharides under stress conditions by
Porphyridium purpureum was also noted by Li et al. [
20] who underlined a rapid secretion of EPS with limited nitrogen and strong light intensity. The presence of polysaccharides in the microalgae suspension can be observed by the high opacity and turbidity of the suspension volume and was also determined analytically.
3.2. Nutrient Removal Efficiency
The nutrient removal efficiency of the microalgae strain
Porphyridium purpureum was monitored on a daily basis and the results are presented in
Supplementary Materials—Figures S1–S6, for the COD, nitrogen and phosphorus in both of the illumination scenarios.
The microalgae strain Porphyridium purpureum presented very good results for nutrient reduction from dairy waste waters. The reduction of lactose from the growth medium was up to 100%, with a minimum reduction of 87% for the most concentrated sample. Regarding the COD values, the minimum reduction was 68% and it reached up to 92% in only 7 days of cultivation. The nitrogen and phosphorus reductions were as high as 100%, with the best results rendered from the continuous illumination strategy. The reduction of these parameters is proportional to the production of the microalgal biomass. The microalgae strain uses the nutrients that are present in the growth medium to grow and multiply and this can be seen from the experimental results.
There is a European regulation that provides the limitations for water streams that are released into the environment (European Directive 91/271/EEC) and it states the following requirements: a reduction of minimum 70–80% for total nitrogen, a reduction of minimum 80% for phosphorus and a reduction of at least 75% for COD. The present research results are well within these limitations that are imposed by the EU.
Regarding the mechanism of organic load removal by the
Porphyridium purpureum microalgae strain from cheese whey, literature sources present that, in order to assimilate lactose, living organisms have to synthesize a β-galactosidase enzyme in order to hydrolyze the lactose into glucose and galactose and to internalize the molecules through the adequate transmembrane proteins. In
Escherichia coli, the lac operon is responsible for the expression and regulation of these enzymes as a function of the lactose and glucose availability in its environment. Certain plant and bacterial β-galactosidases can even be secreted outside the cell, resulting in a necessary internalization of the glucose and galactose molecules instead. In the microalgae
Chlorella kessleri, transmembrane transporters allow for this internalization to occur, but with different affinities for glucose and galactose. The ability of
S. obliquus to grow on glucose suggests that a similar sugar transport mechanism is present in its genus [
21].
3.3. Biomass Productivity and Polysaccharide Concentration
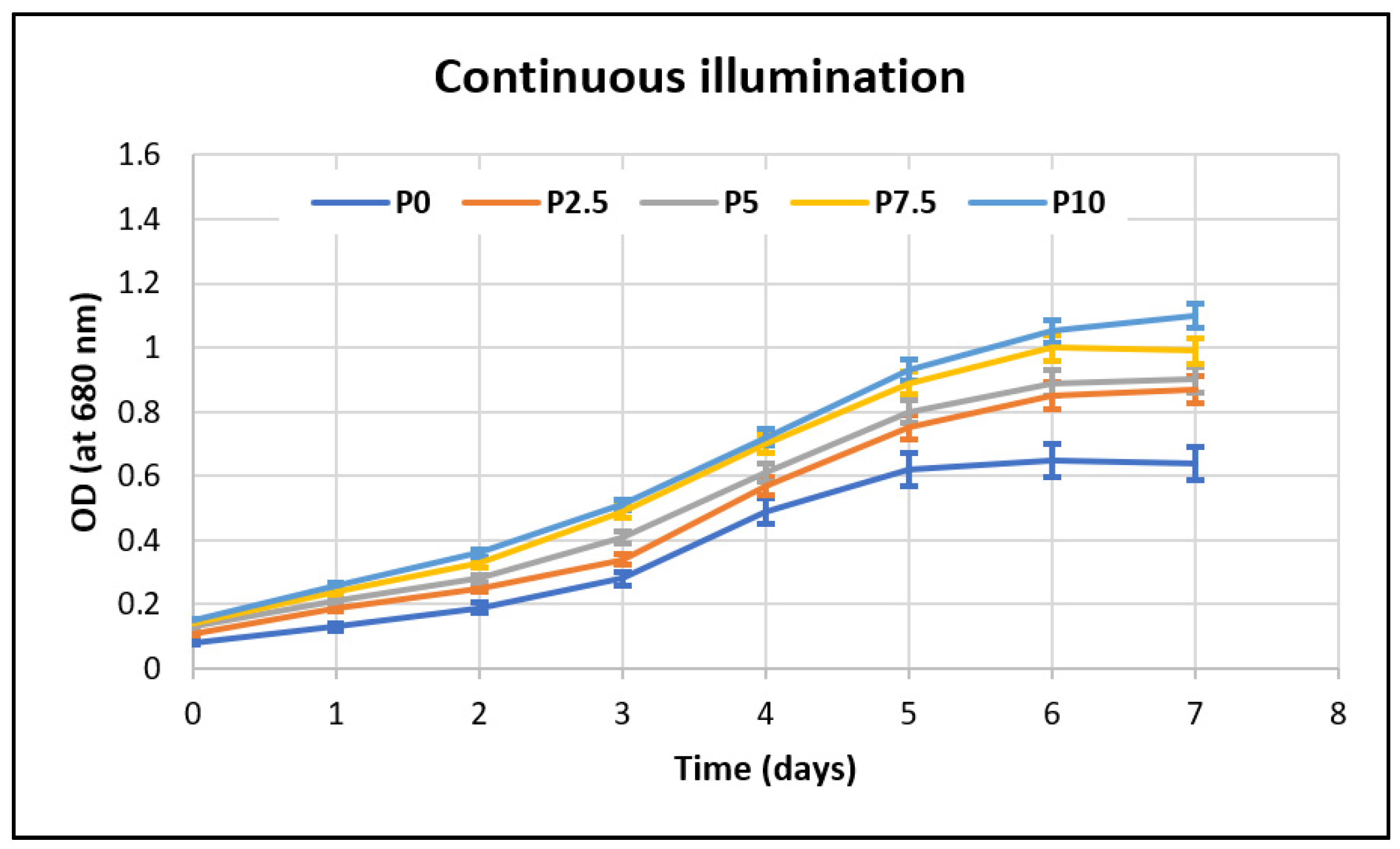
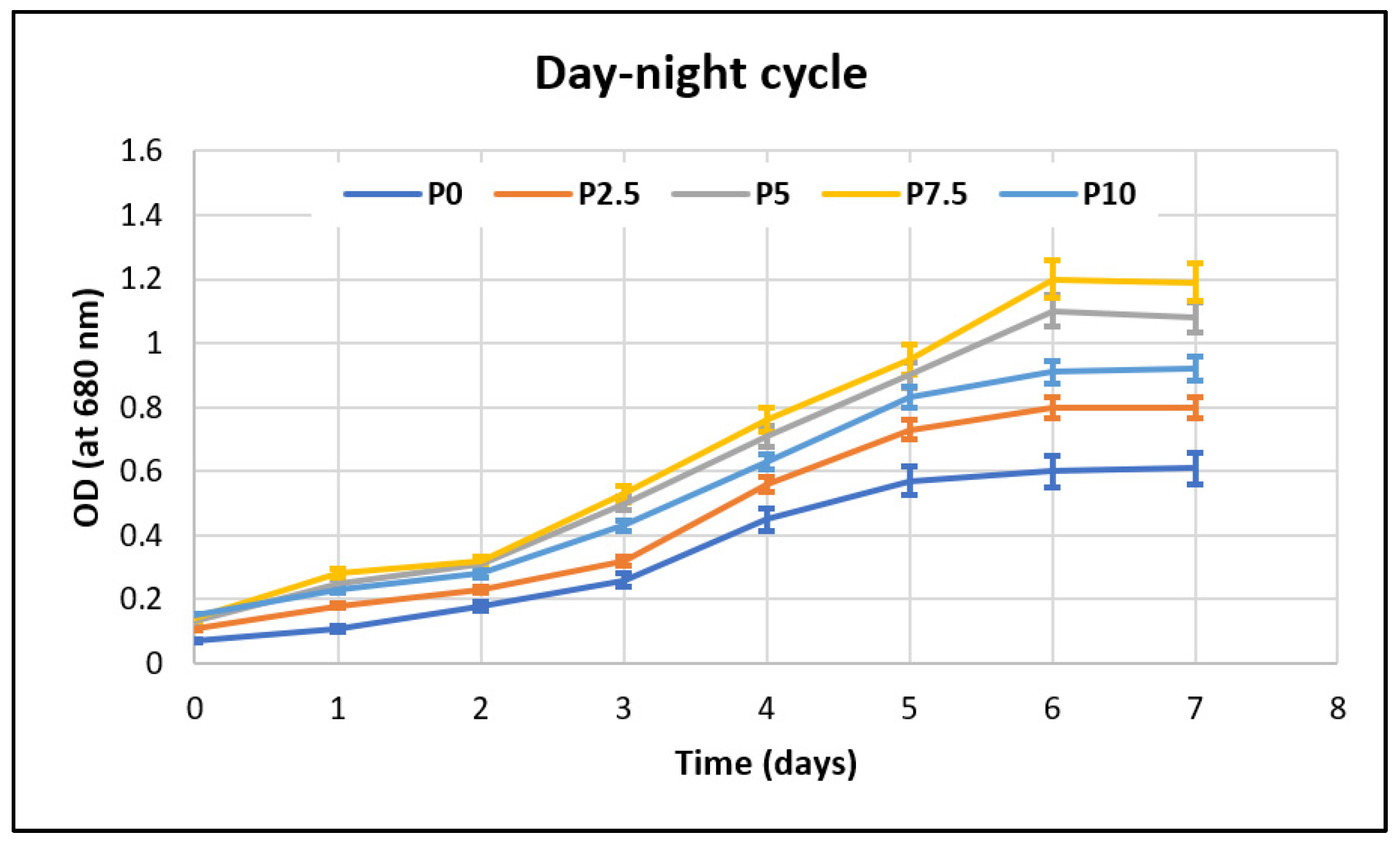
Throughout the growth period, samples were taken daily and analyzed spectrophotometrically, at 680 nm, in order to monitor the effect that the stress factors had on the microalgae’s development. The biomass growth curves of
Porphyridium purpureum are presented in
Figure 3, for continuous illumination, and
Figure 4, for the day–night cycle.
After the removal of the microalgal biomass from the suspension, the exopolysaccharides were separated from the growth medium via precipitation with 96% ethanol on an ice bath. A visual analysis of the growth medium samples after biomass removal, as presented in
Figure 2C, highlights the increase in the exopolysaccharide concentration with the increase in the lactose content of the samples.
The samples containing exopolysaccharides were subjected to an initial concentration step, via boiling, until the volume was halved. The purpose of concentrating the samples was to facilitate their precipitation with ethanol. The exopolysaccharides were isolated via centrifugation, washed with ethanol and dried. The results regarding the biomass and exopolysaccharides’ concentration as well as the remaining lactose concentration after the microalgae growth are presented in
Table 3, for the microalgae cultivation under both the continuous lighting and the day–night cycle.
It can be observed from
Table 4 and also from
Figure 3 and
Figure 4 that the biomass production increases with the amount of lactose that is present in the growth medium for both of the tested illumination options. The microalgal biomass’ growth benefits from the presence of the extra carbon source, as can be seen from the minimum 4-fold increase in the biomass concentration compared to that of the control sample without lactose. This result highlights that dairy wastewaters that are rich in lactose can be used efficiently as a carbon source for the growth of
Porphyridium purpureum without affecting the growth process and while, moreover, increasing the biomass’ concentration and achieving a high reduction in the lactose from this waste stream.
Regarding the exopolysaccharide concentration in the growth medium, this amount also increased with the lactose availability, which indicates that the microalgal biomass consumed lactose for cell growth, but also by transforming it and excreting polysaccharides as a mechanism for coping with the stress that was caused by the excessive presence of this carbon source [
22].
3.4. Pigment Extraction and Quantification
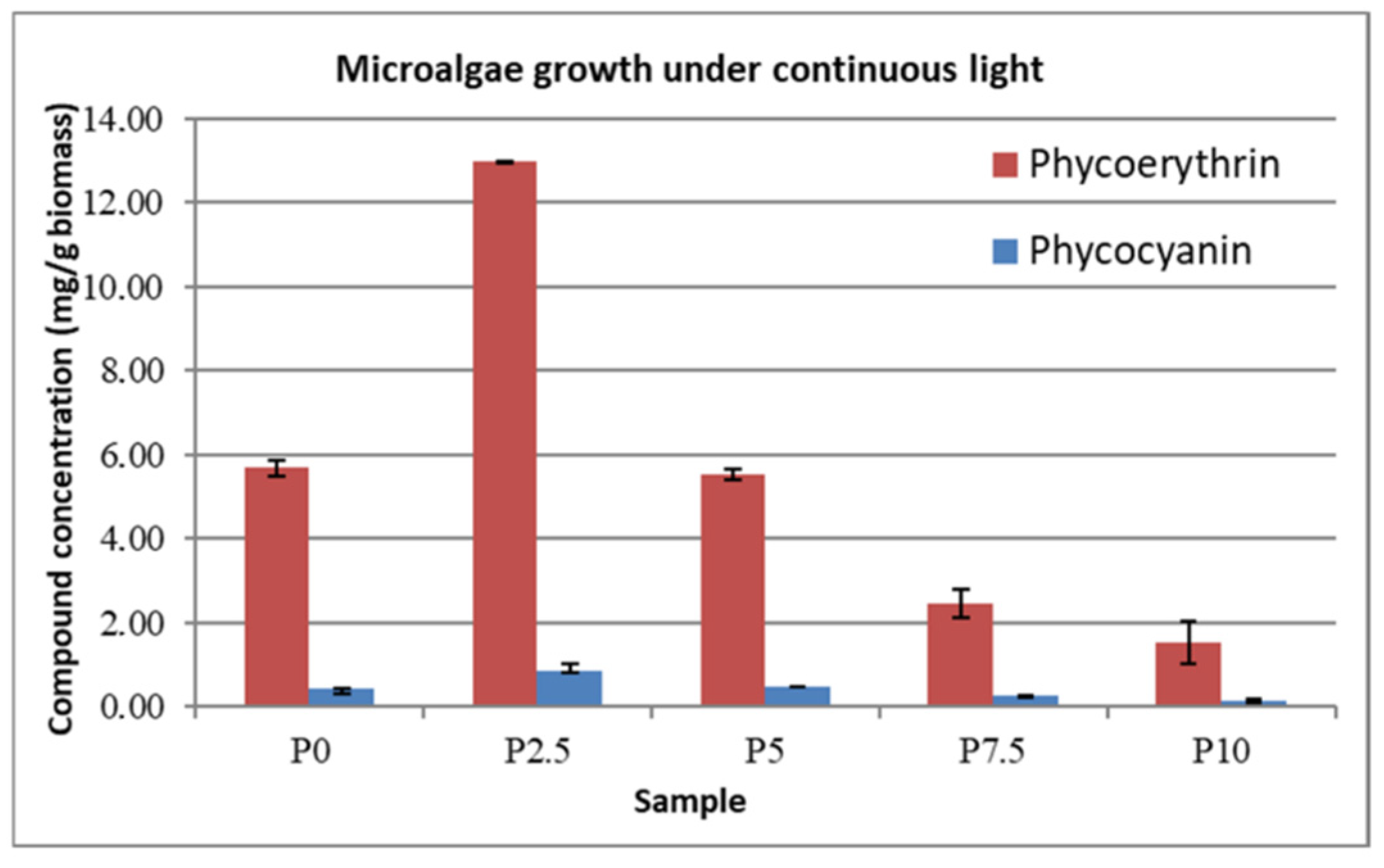
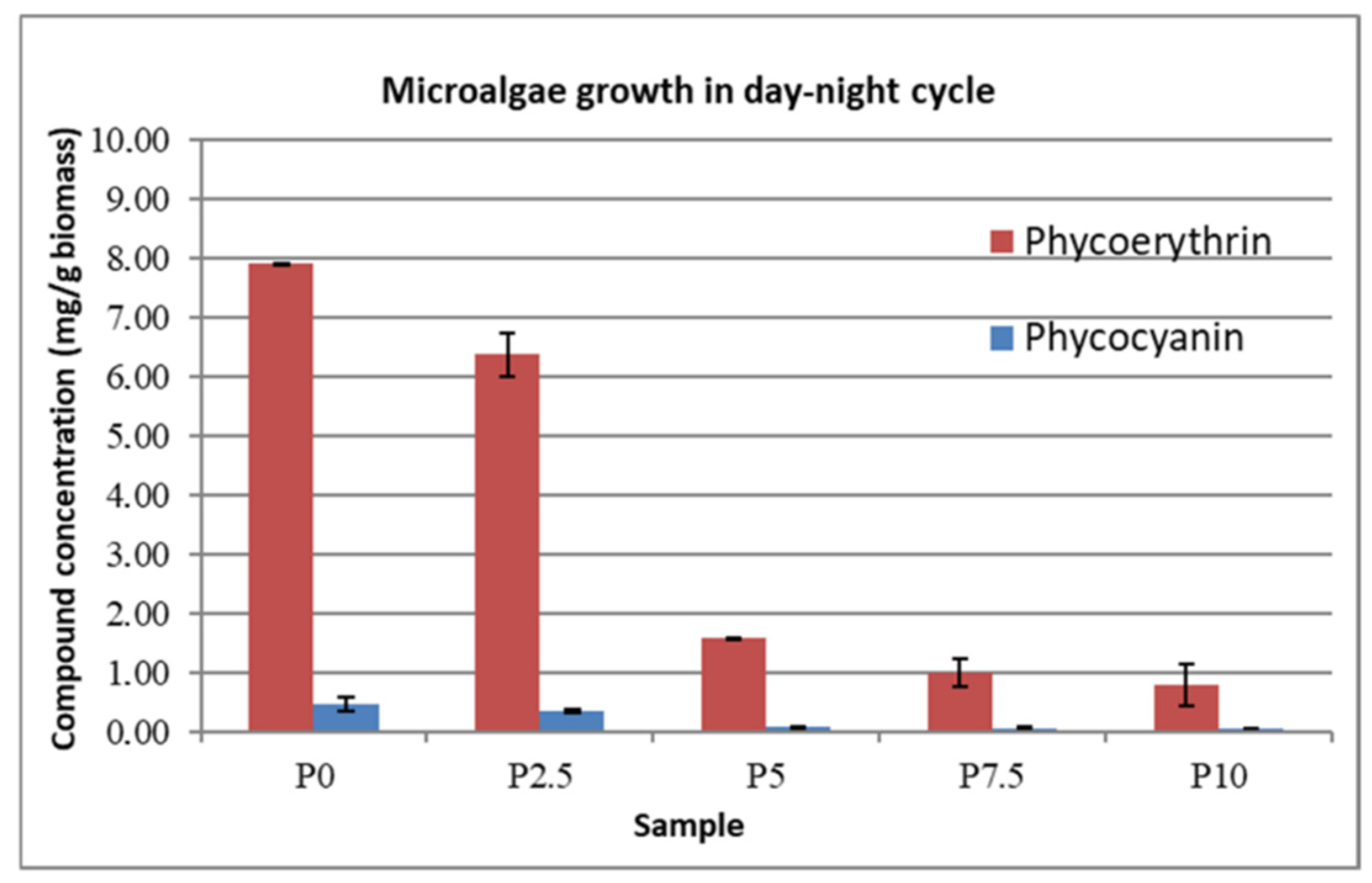
In
Figure 5 and
Figure 6, the concentrations of phycoerythrin and phycocyanin that were obtained for each sample are represented, with the highest concentrations being obtained under the continuous light condition in the case of the ASW that was modified with 2.5 g∙L
−1 lactose. While a small variation can be observed for the control samples between the two types of cultivation, more significant growth can be observed for every whey-modified sample under continuous illumination over the growth of the equivalent samples that were grown in day–night cycles.
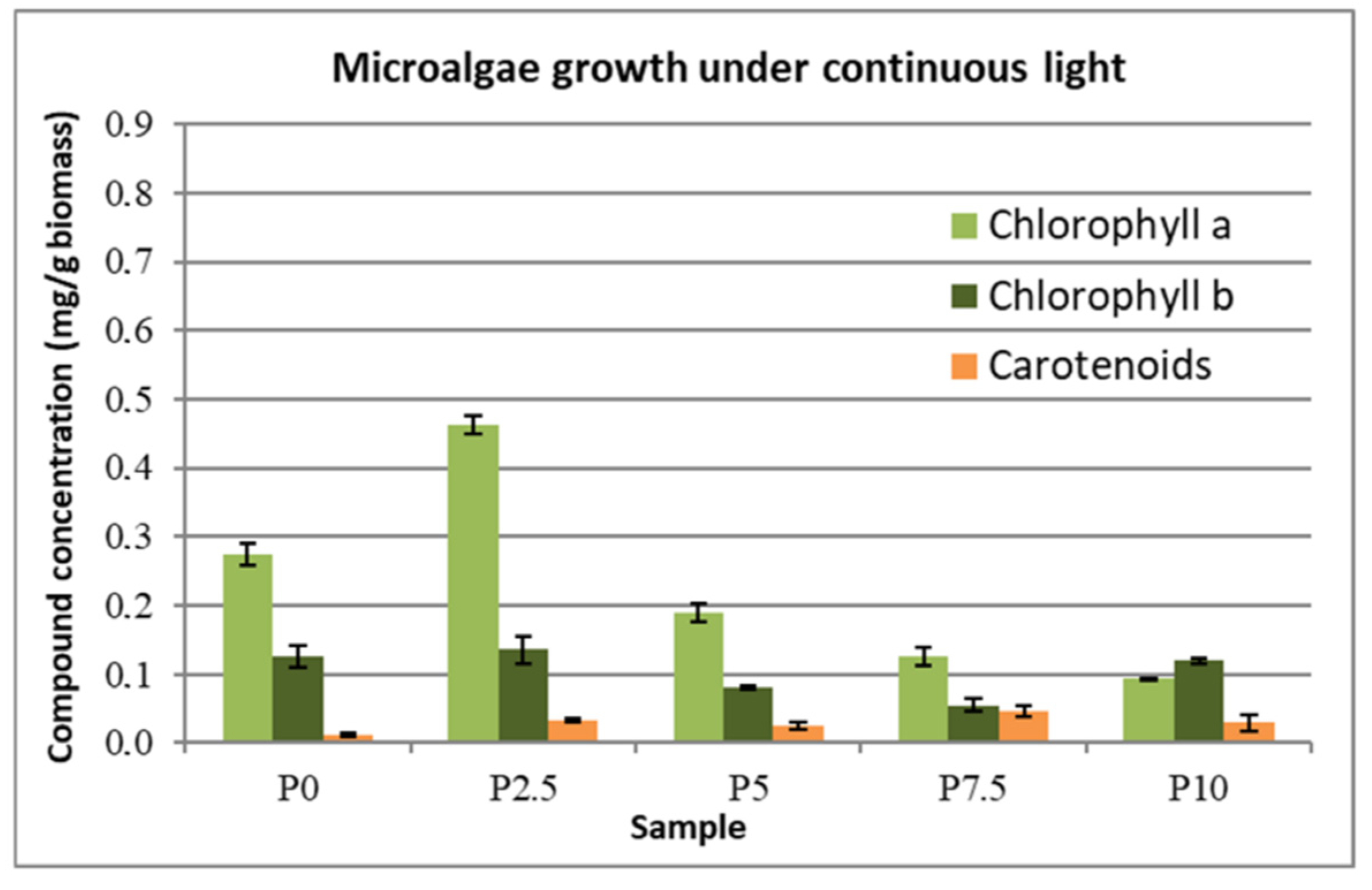
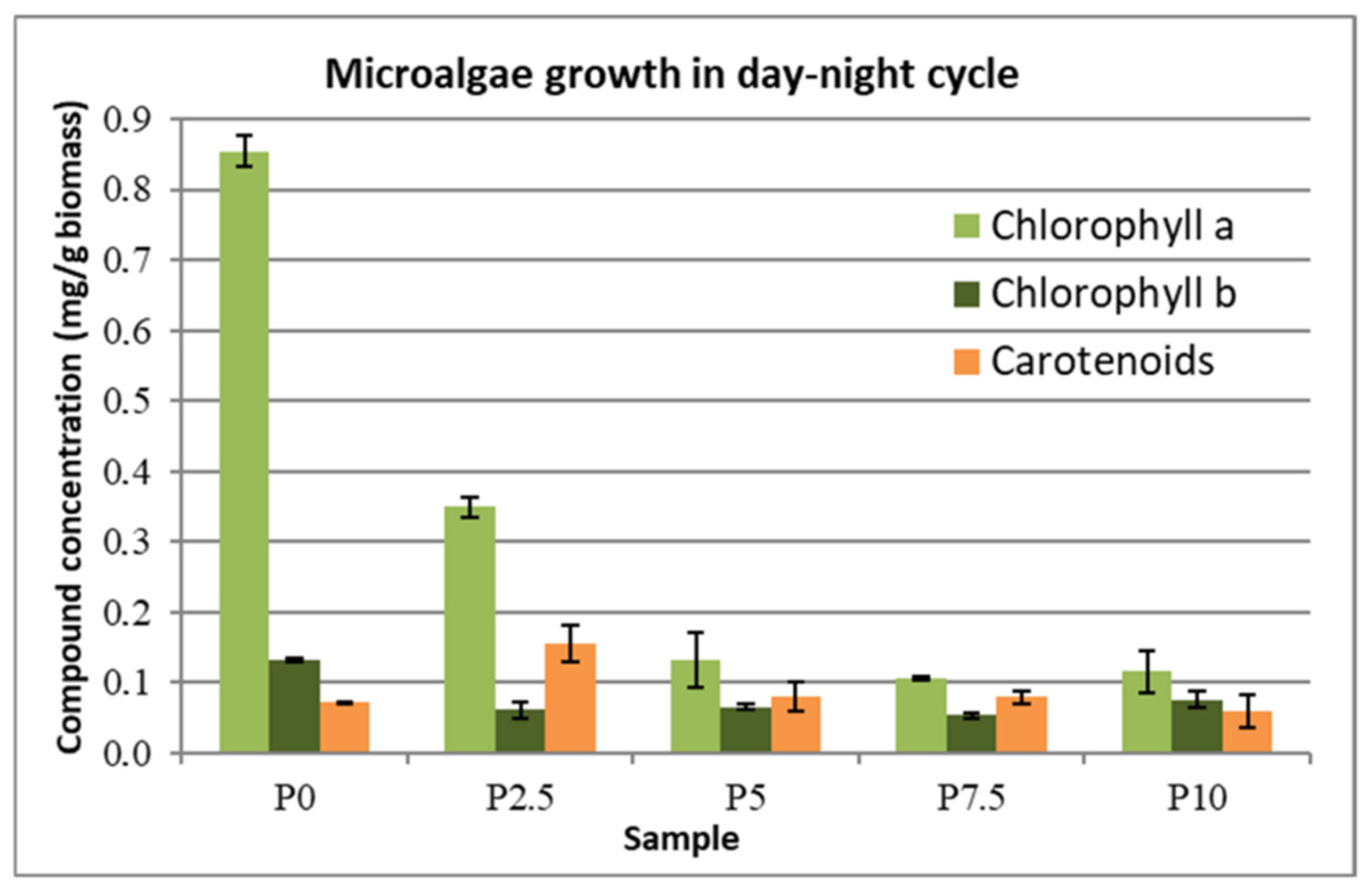
The concentrations of the chlorophylls and carotenoids that were produced by the microalgal biomass under continuous illumination and the day–night illumination cycle are presented in
Figure 7 and
Figure 8. For the samples without the addition of lactose (the control samples), the content of pigments was higher in the case of day–night cycle of illumination than in the case of the light stress that was caused by continuous illumination. This can be explained by the fact that the microalgae cannot cope with this stress, therefore it cannot produce the compounds that were previously mentioned. This explanation is supported by the fact that, in the presence of small amounts of lactose (2.5 g∙L
−1), the pigment concentration increased 2-fold, meaning that the presence of this extra carbon source aided the cells to cope with the light stress. However, by increasing the lactose concentration in the growth medium (5–10 g∙L
−1) this condition itself becomes a supplementary stress factor, thus leading the concentration of the pigments to decrease with the increase of the lactose concentration. Carotenoids seem to be an exception to this rule, as their concentration increases slightly in the case of light stress with the presence of lactose and, for the case of the day–night cycle, the concentration doubled at 2.5 g∙L
−1 lactose addition, while at higher concentrations of lactose it remained the same as the sample without lactose. The carotenoid concentration also decreased in the case of light stress, as was the case for the other compounds.
3.5. Exopolysaccharide Characterization
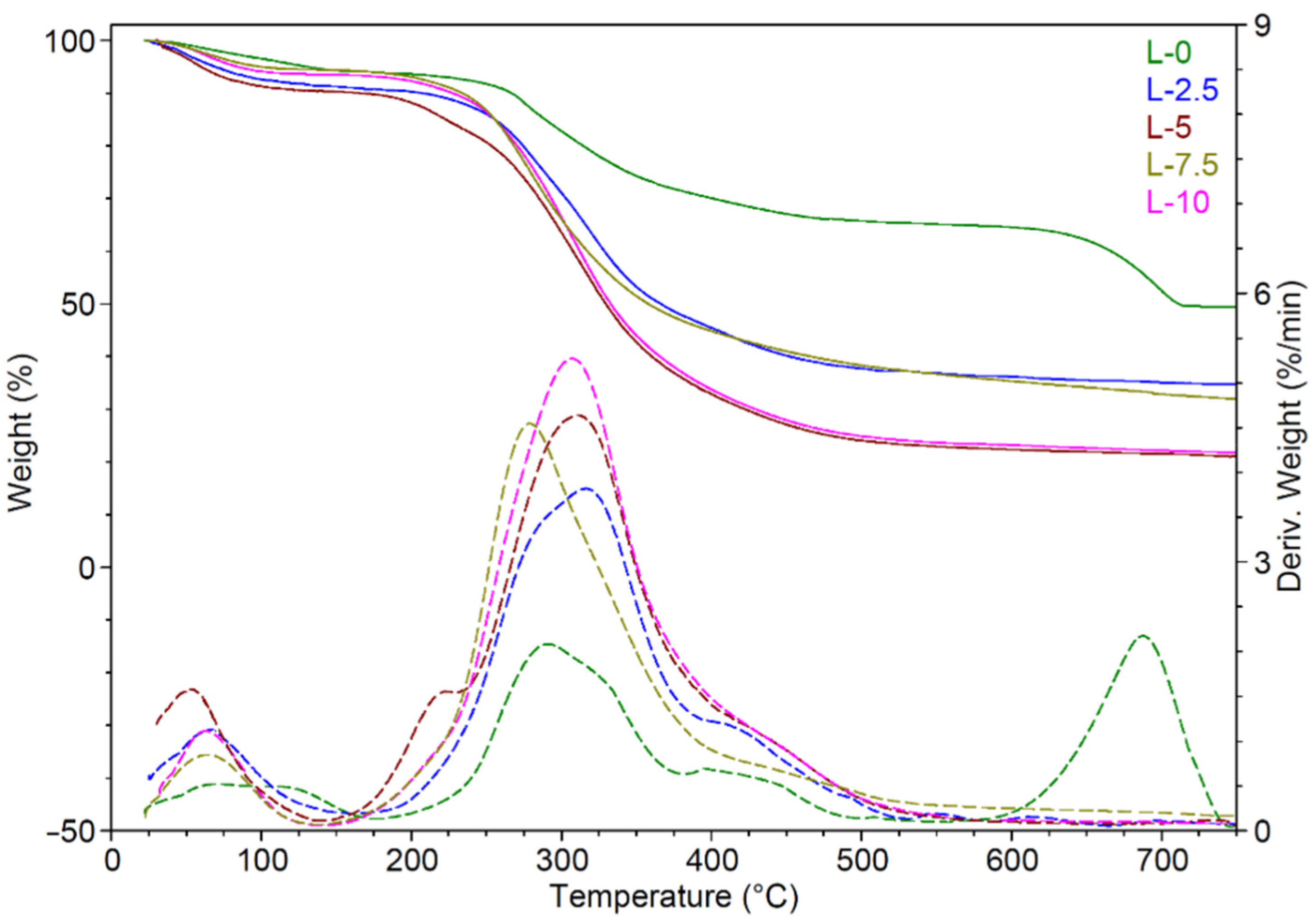
Thermogravimetric analysis (TGA) was carried out only for the EPS samples that were obtained under continuous illumination, considering that the results for both growth processes were similar.
The thermogravimetric curves for the blank EPS sample (the EPS that were obtained from cultivation of microalgae without added whey) and the ones containing lactose are shown in
Figure 9 and the most important characteristics are summarized in
Table 5: the weight loss (WL), the maximum degradation temperatures (T
d) for the decomposition steps and the residue at 700 °C.
A good level of thermal stability of the EPS is crucial for preserving their structure and properties in practical applications where thermostable polymers are needed. One can observe that all of the analyzed EPS samples were stable up to 160 °C; the weight loss being below 10%, except for that of sample L5.
The blank EPS sample showed a three-step degradation process (
Figure 9). An initial weight loss of 5.8% was observed between 25 and 160 °C, corresponding to the dehydration process. This first step of degradation may be due to the presence of a large number of carboxyl groups in the EPS structure which favors water and moisture retention [
23,
24]. The second degradation step, which occurred between 160 and 575 °C with a major loss of mass (29.5%), involved the depolymerization of the EPS accompanied by chemical bond breakage [
25]. The final decomposition stage between 575 and 700 °C (15.43%) may be associated with the degraded residual monosaccharide and presence of trace inorganic salts [
26]. A similar degradation profile was found in the case of the EPS that were extracted from different sources [
27,
28,
29].
The maximum degradation temperature (T
d1) of the blank EPS, determined from the DTG curve, was found to be 291 °C (
Figure 9). This value is higher than that of some early reported EPS, such as those that were derived from
Lactobacillus plantarum JNULCC001 grown on 12% cheese whey, which were found to have a Td that was 27 °C lower than those which were obtained in this work [
24]. Moreover, a broad shoulder was observed on the right side of the blank EPS’s main peak (430 °C) and this was related to the pyrolysis of the high molecular weight polysaccharides [
29].
Similar shaped TG/DTG curves were obtained for the EPS samples containing lactose but with different characteristic temperatures and residue at 700 °C (
Figure 9,
Table 5). Notably, in all of the EPS samples there was an increase in the weight loss in the interval of temperature between 160 and 575 °C with increasing lactose concentration (
Figure 9,
Table 5). Thus, an increase of ~138% in weight loss was observed in the case of the EPS samples containing the higher lactose concentration. Additionally, insignificant weight loss values were found in the interval of temperature between 575 and 700 °C for the EPS samples containing lactose as compared with the blank EPS sample (
Table 5). The maximum degradation temperature (T
d1) was higher for the EPS samples containing lactose as compared to the blank EPS sample; the addition of lactose in low concentrations led to the highest increase of the Td1 value, by ~26 °C (E2.5), where at higher concentrations the increase was only by ~16 °C (E10). The L7.5 sample displayed a different thermal behavior with a slight decrease of the T
d1 with about 12 °C as compared with the blank EPS. This distinct thermal behavior could be due to the inhomogeneity of this sample. Moreover, a significant reduction in residue was obtained after the degradation of the EPS samples containing lactose compared to the blank EPS (
Table 5).
All of these results revealed that the EPS that is derived from cheese whey can be used for food applications or for active principles encapsulation where they can ensure the protection of the active principle up to high temperatures without losing their thermal performance.
Due to their lack of toxicity, EPS are increasingly taken into account in food technology through their interesting functional properties, which can be exploited to impart attractive properties to foods [
30]. Considering that the EPS that can be derived from
Porphyridium purpureum microalgae contain polysaccharide sulphate which can provide antioxidant, antibacterial and antiviral properties they have great potential to be used, besides pharmaceutically, in the food industry; for example, as a natural preservative (assuring the storage stability of processed food products) or as a natural thickener [
14,
31,
32,
33,
34]. Considering that food processing also involves technological operations that take place at high temperatures, the thermal stability of EPS is a must. So, the thermal stability is an important demand when taking into account that the biological activity of the bioactive compounds from EPS may depend on their structural integrity. Therefore, it is important to know the effects of heat treatment in order to maintain the EPS’ integrity in processed foods.
Moreover, because they are biodegradable and sustainable, EPS are also promising for food packaging development. For example, EPS can be used as filler in other biopolymers for obtaining food packaging biomaterials with antibacterial properties. In this respect, EPS needs to have a good level of thermal stability in order to maintain its structure during the processing methods that are typical for the food packaging thermoplastic industry, such as extrusion, melt-blowing, injection and others.
4. Conclusions and Perspectives
This paper has approached the study of dairy wastewater treatment and simultaneous biocompound production by Porphyridium purpureum, under continuous light and under a day–night cycle. The main goals were to achieve a reduction in the lactose content of the cheese wastewater that was tested and, at the same time, to obtain value-adding compounds so as to increase the economic value of the process.
The results show that biomass production increases with the amount of lactose that is present in the growth medium, for both of the illumination options, until the lactose is almost completely consumed from the medium (87–100%). The higher the biomass productivity, the better the nitrogen and phosphorus consumption from the growth medium. The COD levels were reduced by a minimum of 68% and up to 100%. The present research results are well within the limitations that are imposed by European Directive 91/271/EEC, which highlights a reduction of minimum 70–80% for total nitrogen, a reduction of minimum 80% for phosphorus and a reduction of at least 75% for COD. Regarding the exopolysaccharide concentration in the growth medium, this amount also increases with lactose availability, which indicates that the microalgal biomass consumes lactose for cell growth, but also by transforming it and excreting polysaccharides as a mechanism for coping with the stress that is caused by the excess presence of this carbon source.
The concentrations of phycobiliproteins that were obtained for each sample were highest in the samples which had small amounts of lactose in the medium (2.5 g∙L−1). A more significant increase in phycobiliproteins was observed for the samples that were under continuous illumination, over the equivalent samples that were grown in day–night cycles. The content of pigments was higher in the case of the day–night cycle of illumination than in the case of the light stress that was caused by continuous illumination. This can be explained by the fact that the microalgae cannot cope with this stress, therefore it cannot produce the compounds that have been mentioned. However, by increasing the lactose concentration in the growth medium (5–10 g∙L−1), this condition itself becomes a supplementary stress factor, thus leading the concentration of the pigments to decrease with the increase in the lactose concentration. In the case of light stress, carotenoid concentrations have increased slightly with the presence of lactose, but generally present a decrease over time, as is the case for other compounds.
The amount of exopolysaccharides that was excreted in the growth medium increased with the level of lactose availability, thus indicating that the microalgae biomass not only consumes lactose for cell growth, but also transforms it and excretes polysaccharides as a mechanism for coping with the stress that is caused by the excess presence of this carbon source. The TGA curves of the EPS samples demonstrate their very high thermal stability, a characteristic that is important for structure- and properties-preservation in practical applications where thermostable polymers are needed.
This result highlights that dairy wastewaters that are rich in lactose can be used efficiently as a carbon source for the growth of Porphyridium purpureum without affecting the growth process but also, moreover, increasing the biomass concentration and achieving a high reduction in the lactose from this waste stream while obtaining a microalgae biomass that is rich in valuable compounds. Further studies will focus on an extensive characterization of these compounds.


 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}