
Exploring Olive Genetic Diversity in the Maltese Islands
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
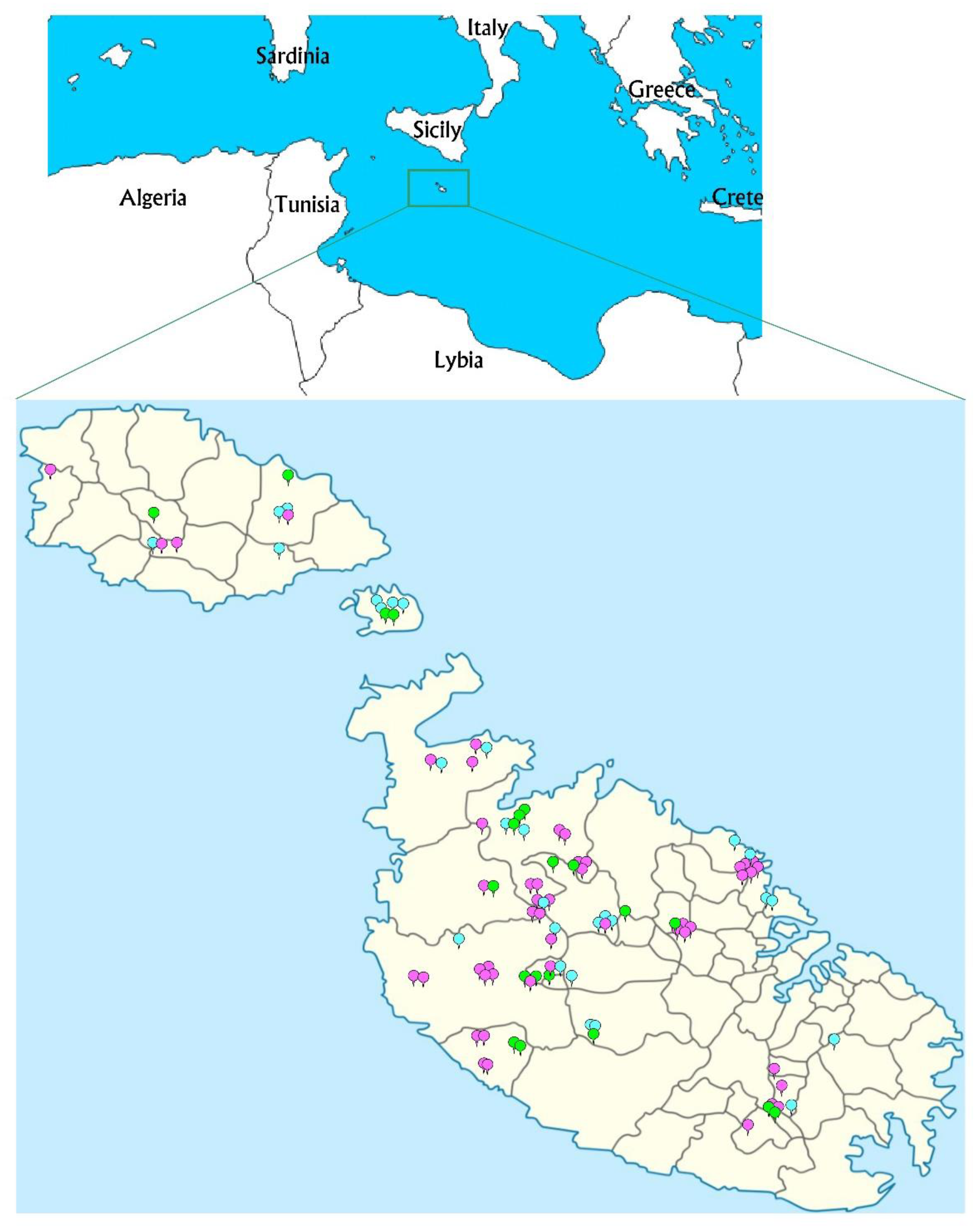
2.1. Plant Material and DNA Extraction
2.2. Chloroplast and Nuclear Marker Applied to Maltese Olive Genotypes
2.3. Frequency Analysis and Genetic Differentiation
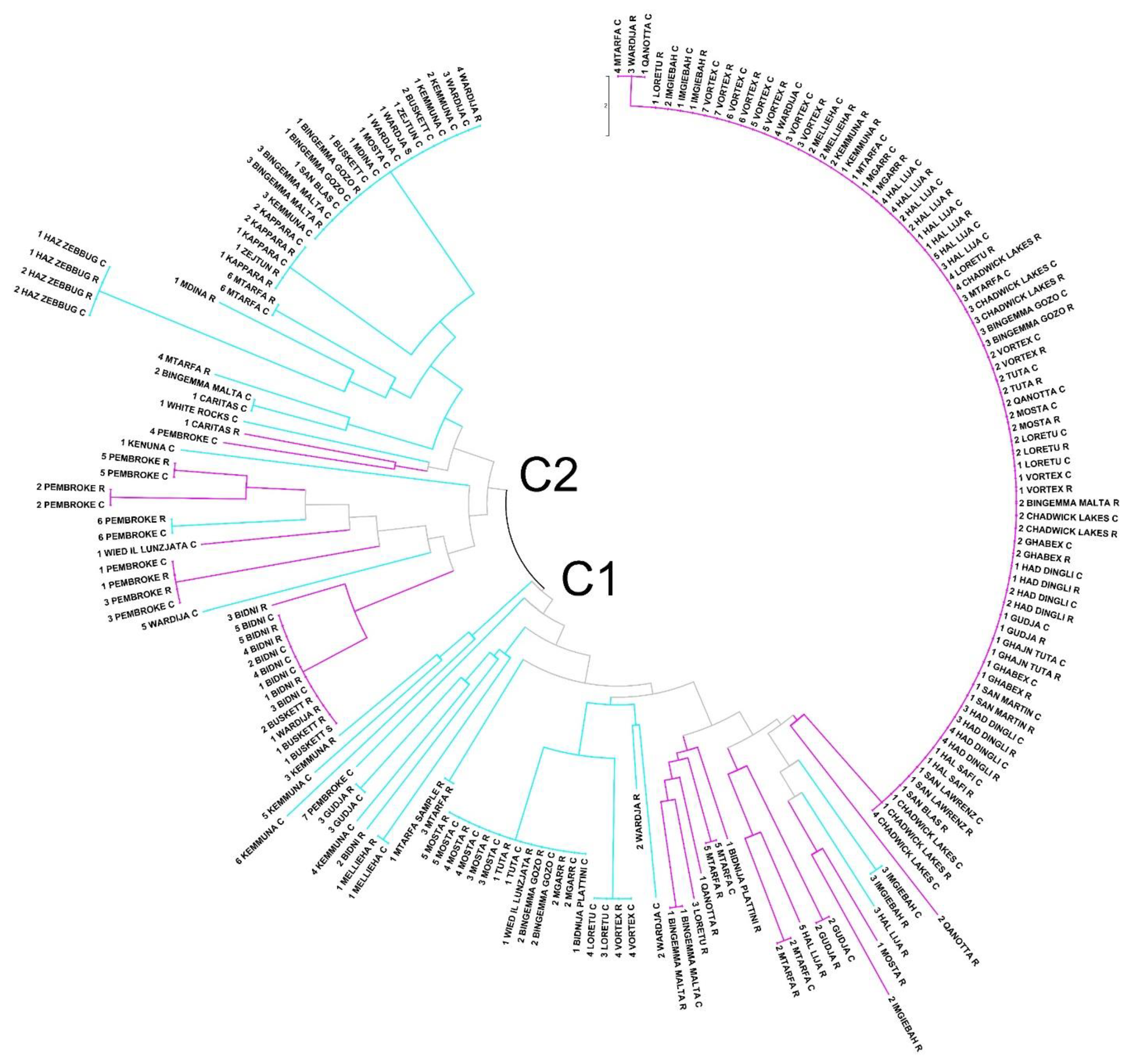
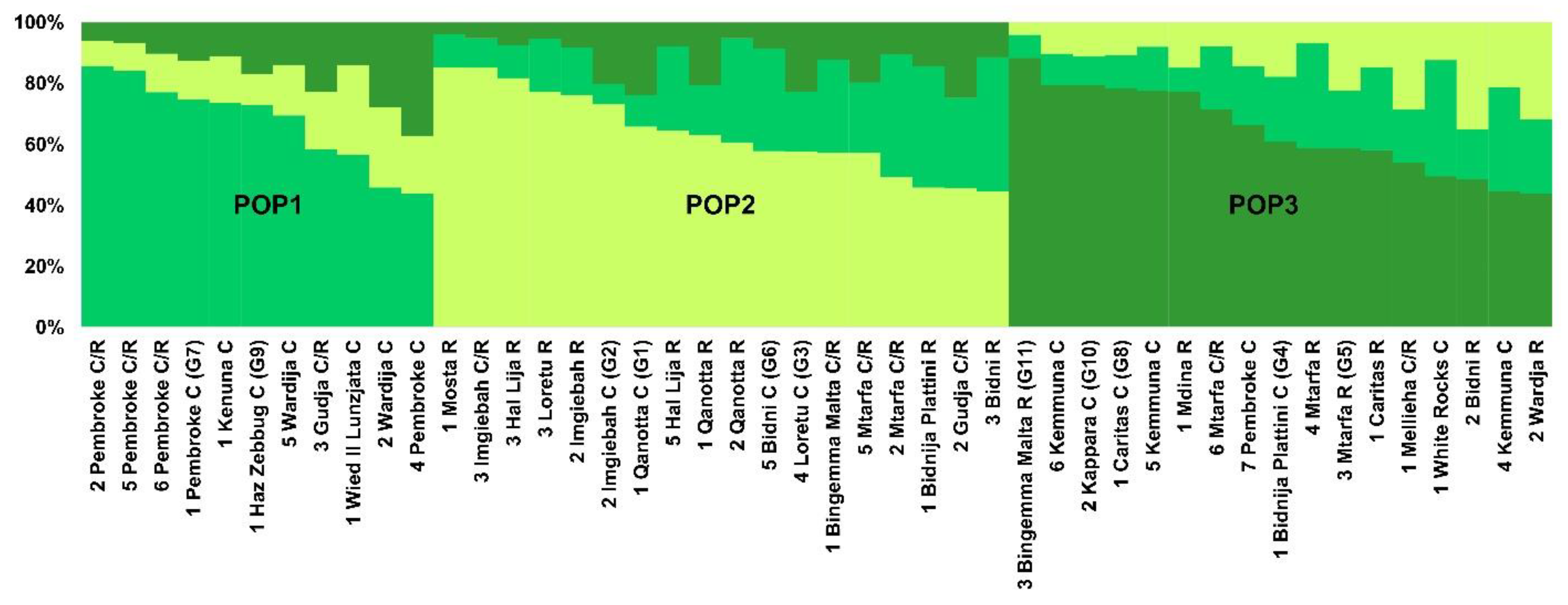
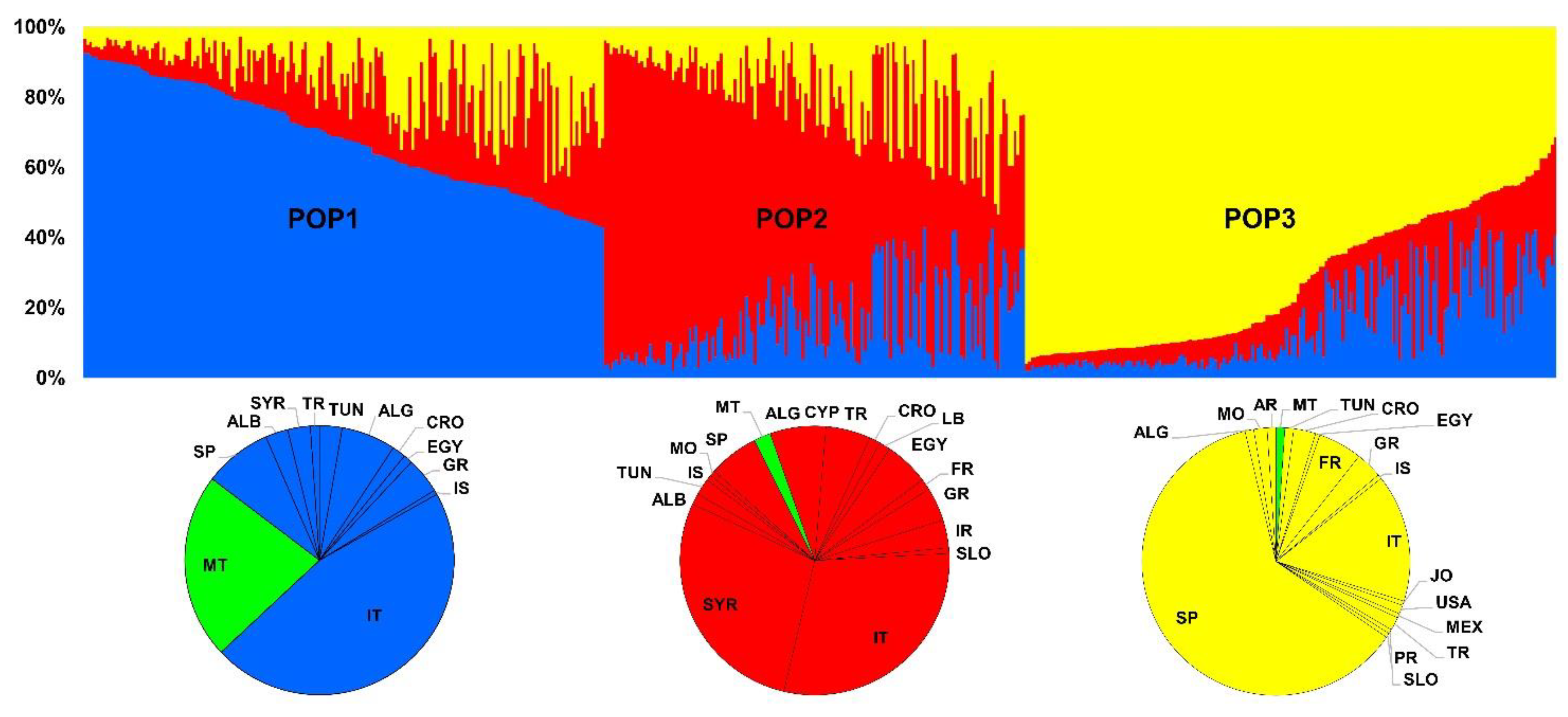
3. Results
3.1. Genetic Diversity Characterized according to SSR and cp-SSR Markers
3.2. Genetic Differentiation of Maltese Genotypes
3.3. Genetic Diversity and Differentiation between Maltese and Worldwide Genotypes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Díez, C.M.; Trujillo, I.; Martinez-Urdiroz, N.; Barranco, D.; Rallo, L.; Marfil, P.; Gaut, B.S. Olive domestication and diversification in the Mediterranean Basin. New Phytol. 2015, 206, 436–447. [Google Scholar] [CrossRef] [PubMed]
- Besnard, G.; Khadari, B.; Navascués, M.; Fernández-Mazuecos, M.; El Bakkali, A.; Arrigo, N.; Baali-Cherif, D.; Brunini-Bronzini de Caraffa, V.; Santoni, S.; Vargas, P.; et al. The complex history of the olive tree: From Late Quaternary diversification of Mediterranean lineages to primary domestication in the northern Levant. Proc. Natl. Acad. Sci. USA 2013, 280, 20122833. [Google Scholar] [CrossRef] [PubMed]
- Besnard, G.; Terral, J.F.; Cornille, A. On the origins and domestication of the olive: A review and perspectives. Ann. Bot. 2018, 121, 385–403. [Google Scholar] [CrossRef] [PubMed]
- Kassa, A.; Konrad, H.; Geburek, T. Molecular diversity and gene flow within and among different subspecies of the wild olive (Olea europaea L.): A review. Flora 2019, 250, 18–26. [Google Scholar] [CrossRef]
- Erre, P.; Chessa, I.; Muñoz-Diez, C.; Belaj, A.; Rallo, L.; Trujillo, I. Genetic diversity and relationships between wild and cultivated olives (Olea europaea L.) in Sardinia as assessed by SSR markers. Genet. Resour. Crop Evol. 2010, 57, 41–54. [Google Scholar] [CrossRef]
- Marra, F.P.; Caruso, T.; Costa, F.; Di Vaio, C.; Mafrica, R.; Marchese, A. Genetic relationships, structure and parentage simulation among the olive tree (Olea europaea L. subsp. europaea) cultivated in Southern Italy revealed by SSR markers. Tree Genet. Genomes 2013, 9, 961–973. [Google Scholar] [CrossRef]
- Mousavi, S.; Stanzione, V.; Mencuccini, M.; Baldoni, L.; Bufacchi, M.; Mariotti, R. Biochemical and molecular profiling of unknown olive genotypes from central Italy: Determination of major and minor components. Eur. Food Res. Technol. 2019, 245, 83–94. [Google Scholar] [CrossRef]
- Baldoni, L.; Tosti, N.; Ricciolini, C.; Belaj, A.; Arcioni, S.; Pannelli, G.; Germana, M.A.; Mulas, M.; Porceddu, A. Genetic structure of wild and cultivated olives in the central Mediterranean basin. Ann. Bot. 2006, 98, 935–942. [Google Scholar] [CrossRef]
- Díez, C.M.; Trujillo, I.; Barrio, E.; Belaj, A.; Barranco, D.; Rallo, L. Centennial olive trees as a reservoir of genetic diversity. Ann. Bot. 2011, 108, 797–807. [Google Scholar] [CrossRef]
- El Bakkali, A.; Essalouh, L.; Tollon, C.; Rivallan, R.; Mournet, P.; Moukhli, A.; Hadidou, A.; Sikaoui, L.; Khadari, B. Characterization of Worldwide Olive Germplasm Banks of Marrakech (Morocco) and Córdoba (Spain): Towards management and use of olive germplasm in breeding programs. PLoS ONE. 2019, 14, e0223716. [Google Scholar] [CrossRef] [Green Version]
- Barazani, O.; Westberg, E.; Hanin, N.; Dag, A.; Kerem, Z.; Tugendhaft, Y.; Hmidat, M.; Hijawi, T.; Kadereit, J.W. A comparative analysis of genetic variation in rootstocks and scions of old olive trees–a window into the history of olive cultivation practices and past genetic variation. BMC Plant Biol. 2014, 14, 146. [Google Scholar] [CrossRef] [PubMed]
- Bernabei, M. The age of the olive trees in the Garden of Gethsemane. J. Archaeol. Sci. 2015, 53, 43–48. [Google Scholar] [CrossRef]
- Beghè, D.; Piotti, A.; Satovic, Z.; De La Rosa, R.; Belaj, A. Pollen-mediated gene flow and fine-scale spatial genetic structure in Olea europaea subsp. europaea var. sylvestris. Ann. Bot. 2017, 119, 671–679. [Google Scholar]
- Ninot, A.; Howad, W.; Aranzana, M.J.; Senar, R.; Romero, A.; Mariotti, R.; Baldoni, L.; Belaj, A. Survey of over 4, 500 monumental olive trees preserved on-farm in the northeast Iberian Peninsula, their genotyping and characterization. Sci. Hortic. 2018, 231, 253–264. [Google Scholar] [CrossRef]
- Mousavi, S.; Mariotti, R.; Bagnoli, F.; Costantini, L.; Cultrera, N.G.; Arzani, K.; Pandolfi, S.; Torkzaban, B.; Hosseini-Mazinani, M.; Baldoni, L. The eastern part of the Fertile Crescent concealed an unexpected route of olive (Olea europaea L.) differentiation. Ann. Bot. 2017, 119, 1305–1318. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Rueda, P.; Franco-Navarro, J.D.; Messora, R.; Espartero, J.; Rivero-Núñez, C.M.; Aleza, P.; Capote, N.; Cantos, M.; García-Fernández, J.L.; de Cires, A.; et al. SILVOLIVE, a germplasm collection of wild subspecies with high genetic variability as a source of rootstocks and resistance genes for olive breeding. Front. Plant Sci. 2020, 11, 629. [Google Scholar] [CrossRef]
- Yoruk, B.; Taskin, V. Genetic diversity and relationships of wild and cultivated olives in Turkey. Plant Syst. Evol. 2014, 300, 1247–1258. [Google Scholar] [CrossRef]
- Breton, C.; Tersac, M.; Bervillé, A. Genetic diversity and gene flow between the wild olive (oleaster, Olea europaea L.) and the olive: Several Plio-Pleistocene refuge zones in the Mediterranean basin suggested by simple sequence repeats analysis. J. Biogeogr. 2006, 33, 1916–1928. [Google Scholar] [CrossRef]
- Zohary, D.; Hopf, M.; Weiss, E. The origin and spread of domesticated plants in Southwest Asia, Europe, and the Mediterranean Basin. In Domestication of Plants in the Old World, 4th ed.; Oxford University Press on Demand: Oxford, UK, 2012. [Google Scholar]
- Breton, C.; Terral, J.F.; Pinatel, C.; Médail, F.; Bonhomme, F.; Bervillé, A. The origins of the domestication of the olive tree. C. R. Biol. 2009, 332, 1059–1064. [Google Scholar] [CrossRef]
- Kaniewski, D.; Van Campo, E.; Boiy, T.; Terral, J.F.; Khadari, B.; Besnard, G. Primary domestication and early uses of the emblematic olive tree: Palaeobotanical, historical and molecular evidence from the Middle East. Biol. Rev. 2012, 87, 885–899. [Google Scholar] [CrossRef]
- Zohary, D.; Spiegel-Roy, P. Beginnings of Fruit Growing in the Old World: Olive, grape, date, and fig emerge as important Bronze Age additions to grain agriculture in the Near East. Science 1975, 187, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Aumeeruddy-Thomas, Y.; Moukhli, A.; Haouane, H.; Khadari, B. Ongoing domestication and diversification in grafted olive–oleaster agroecosystems in Northern Morocco. Reg. Environ. Chang. 2017, 17, 1315–1328. [Google Scholar] [CrossRef]
- Abbott, A. Olive tree gridlock eases: Court ruling paves way for containment of deadly plant disease, but risk of spread remains. Nature 2016, 533, 299–301. [Google Scholar] [CrossRef] [PubMed]
- Almeida, R.P. Can Apulia’s olive trees be saved? Science 2016, 353, 346–348. [Google Scholar] [CrossRef] [PubMed]
- Mousavi, S.; Mariotti, R.; Valeri, M.C.; Regni, L.; Lilli, E.; Albertini, E.; Proietti, P.; Businelli, D.; Baldoni, L. Characterization of differentially expressed genes under salt stress in olive. Int. J. Mol. Sci. 2021, 23, 154. [Google Scholar] [CrossRef] [PubMed]
- Lazović, B.; Adakalić, M.; Pucci, C.; Perović, T.; Bandelj, D.; Belaj, A.; Mariotti, R.; Baldoni, L. Characterizing ancient and local olive germplasm from Montenegro. Sci. Hortic. 2016, 209, 117–123. [Google Scholar] [CrossRef]
- Trujillo, I.; Ojeda, M.A.; Urdiroz, N.M.; Potter, D.; Barranco, D.; Rallo, L.; Diez, C.M. Identification of the Worldwide Olive Germplasm Bank of Córdoba (Spain) using SSR and morphological markers. Tree Genet. Genomes 2014, 10, 141–155. [Google Scholar] [CrossRef]
- Fendri, M.; Trujillo, I.; Trigui, A.; Rodríguez-García, M.I.; Ramírez, J.D.D.A. Simple sequence repeats identification and endocarp characterization of olive tree accessions in a Tunisian germplasm collection. Hortscience 2010, 45, 1429–1436. [Google Scholar] [CrossRef]
- Mariotti, R.; Cultrera, N.G.; Díez, C.M.; Baldoni, L.; Rubini, A. Identification of new polymorphic regions and differentiation of cultivated olives (Olea europaea L.) through plastome sequence comparison. BMC Plant Biol. 2010, 10, 1–13. [Google Scholar] [CrossRef]
- Besnard, G.; Hernández, P.; Khadari, B.; Dorado, G.; Savolainen, V. Genomic profiling of plastid DNA variation in the Mediterranean olive tree. BMC Plant Biol. 2011, 11, 80. [Google Scholar] [CrossRef]
- Mariotti, R.; Belaj, A.; De La Rosa, R.; Leòn, L.; Brizioli, F.; Baldoni, L.; Mousavi, S. EST–SNP study of Olea europaea L. uncovers functional polymorphisms between cultivated and wild olives. Genes 2020, 11, 916. [Google Scholar] [CrossRef] [PubMed]
- Fenech, K. Human-Induced Changes in the Environment and Landscape of the Maltese Islands from the Neolithic to the 15th Century AD; BAR 1682; Archaeopress: Oxford, UK, 2007. [Google Scholar]
- Gambin, B.; Andrieu-Ponel, V.; Médail, F.; Marriner, N.; Peyron, O.; Montade, V.; Gambin, T.; Morhange, C.; Belkacem, D.; Djamali, M. 7300 years of vegetation history and climate for NW Malta: A Holocene perspective. Clim. Past. 2016, 12, 273–297. [Google Scholar] [CrossRef]
- Lageard, J.G.; Sultana, D.; Brearley, F.Q. Veteran trees in an historic landscape: The Bidnija olive grove, Malta. J. Archaeol. Sci. Rep. 2021, 38, 103094. [Google Scholar] [CrossRef]
- Mazzitelli, O.; Calleja, A.; Sardella, D.; Farrugia, C.; Zammit-Mangion, M. Analysis of the molecular diversity of Olea europaea in the Mediterranean Island of Malta. Genet. Resour. Crop Evol. 2015, 62, 1021–1027. [Google Scholar] [CrossRef]
- Baldoni, L.; Cultrera, N.G.; Mariotti, R.; Ricciolini, C.; Arcioni, S.; Vendramin, G.G.; Buonamici, A.; Porceddu, A.; Sarri, V.; Ojeda, M.A.; et al. A consensus list of microsatellite markers for olive genotyping. Mol. Plant Breed. 2009, 24, 213–231. [Google Scholar] [CrossRef]
- Sefc, K.M.; Lopes, M.S.; Mendonca, D.; Dos Santos, M.R.; Laimer, M.; da Câmara Machado, A. Identification of microsatellite loci in olive (Olea europaea) and their characterization in Italian and Iberian olive trees. Mol. Ecol. 2000, 9, 1171–1173. [Google Scholar] [CrossRef]
- De la Rosa, R.; James, C.M.; Tobutt, K.R. Isolation and characterization of polymorphic microsatellite in olive (Olea europaea L.) and their transferability to other genera in the Oleaceae. Mol. Ecol. 2002, 2, 265–267. [Google Scholar] [CrossRef]
- Carriero, F.; Fontanazza, G.; Cellini, F.; Giorio, G. Identification of simple sequence repeats (SSRs) in olive (Olea europaea L.). Theor. Appl. Genet. 2002, 104, 301–307. [Google Scholar] [CrossRef]
- Cipriani, G.; Marrazzo, M.T.; Marconi, R.; Cimato, A.; Testolin, R. Microsatellite markers isolated in olive (Olea europaea L.) are suitable for individual fingerprinting and reveal polymorphism within ancient cultivars. Theor. Appl. Genet. 2002, 104, 223–228. [Google Scholar] [CrossRef]
- Rotondi, A.; Beghè, D.; Fabbri, A.; Ganino, T. Olive oil traceability by means of chemical and sensory analyses: A comparison with SSR biomolecular profiles. Food. Chem. 2011, 129, 1825–1831. [Google Scholar] [CrossRef]
- Unver, T.; Wu, Z.; Sterck, L.; Turktas, M.; Lohaus, R.; Li, Z.; Yang, M.; He, L.; Deng, T.; Escalante, F.J.; et al. Genome of wild olive and the evolution of oil biosynthesis. Proc. Natl. Acad. Sci. USA 2017, 114, E9413–E9422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research an update. Bioinformatics 2013, 28, 2537–2539. [Google Scholar] [CrossRef]
- Marshall, T.C.; Slate, J.; Kruuk, L.E.B.; Pemberton, J.M. Statistical confidence for likelihood-based paternity inference in natural populations. Mol. Ecol. 1998, 7, 639–655. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.; Wen, X.; Falush, D. Documentation for Structure Software: Version 2.3. 2009. Available online: http://pritch.bsd.uchicago.edu/structure.html (accessed on 19 July 2022).
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Earl, D.A. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Barker, G.; Gilbertson, D.; Jones, B.; Mattingly, D. Farming the Desert: The UNESCO Libyan Valleys Archaeological Survey; UNESCO: London, UK, 1996. [Google Scholar]
- Pantaléon-Cano, J.; Yll, E.I.; Pérez-Obiol, R.; Roure, J.M. Palynological evidence for vegetational history in semi-arid areas of the western Mediterranean (Almería, Spain). Holocene 2003, 13, 109–119. [Google Scholar] [CrossRef]
- Cicatelli, A.; Fortunati, T.; De Feis, I.; Castiglione, S. Oil composition and genetic biodiversity of ancient and new olive (Olea europaea L.) varieties and accessions of southern Italy. Plant Sci. 2013, 210, 82–92. [Google Scholar] [CrossRef]
- Harris, R.J. Root coverage with connective tissue grafts: An evaluation of short-and long-term results. J. Periodontol. 2002, 73, 1054–1059. [Google Scholar] [CrossRef]
- Anestiadou, K.; Nikoloudakis, N.; Hagidimitriou, M.; Katsiotis, A. Monumental olive trees of Cyprus contributed to the establishment of the contemporary olive germplasm. PLoS ONE. 2017, 12, e0187697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schicchi, R.; Speciale, C.; Amato, F.; Bazan, G.; Di Noto, G.; Marino, P.; Ricciardo, P.; Geraci, A. The monumental olive trees as biocultural heritage of Mediterranean landscapes: The case study of Sicily. Sustainability 2021, 13, 6767. [Google Scholar] [CrossRef]
- Gros-Balthazard, M.; Besnard, G.; Sarah, G.; Holtz, Y.; Leclercq, J.; Santoni, S.; Wegmann, D.; Glémin, S.; Khadari, B. Evolutionary transcriptomics reveals the origins of olives and the genomic changes associated with their domestication. Plant J. 2019, 100, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Foxhall, L: Olive Cultivation in Ancient Greece: Seeking the Ancient Economy; Oxford University Press: Oxford, UK, 2007.
- Connell, J.H.; Catlin, P.B. Root physiology and rootstock characteristics. In Olive Production Manual, 2nd ed.; Sibbett, G.S., Ferguson, L., Coviello, J.L., Lindstrand, M., Eds.; University of California, Agricultural and Natural Resources: Los Angeles, CA, USA, 2005; pp. 39–48. [Google Scholar]
- Barazani, O.; Waitz, Y.; Tugendhaft, Y.; Dorman, M.; Dag, A.; Hamidat, M.; Hijawi, T.; Kerem, Z.; Westberg, E.; Kadereit, J.W. Testing the potential significance of different scion/rootstock genotype combinations on the ecology of old cultivated olive trees in the southeast Mediterranean area. BMC Ecol. 2017, 17, 3. [Google Scholar] [CrossRef] [PubMed]
- Schembri, P.J. The Maltese Islands: Climate, vegetation and landscape. GeoJournal 1997, 41, 1–11. [Google Scholar] [CrossRef]
- di Rienzo, V.; Sion, S.; Taranto, F.; D’Agostino, N.; Montemurro, C.; Fanelli, V.; Sabetta, W.; Boucheffa, S.; Tamendjari, A.; Pasqualone, A.; et al. Genetic flow among olive populations within the Mediterranean basin. PeerJ 2018, 6, e5260. [Google Scholar] [CrossRef] [Green Version]
- Camps, G. Massinissa, ou les Débuts de L’histoire; Imprimerie Officielle: Alger, Algeria, 1961.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus | Na | Ne | I | Ho | He | F | Fnull | PIC | PA | PAf |
|---|---|---|---|---|---|---|---|---|---|---|
| DCA3 | 9.00 | 4.36 | 1.70 | 0.91 | 0.77 | −0.18 | −0.10 | 0.74 | - | - |
| DCA5 | 8.00 | 3.28 | 1.46 | 0.78 | 0.69 | −0.13 | −0.07 | 0.65 | - | - |
| DCA9 | 20.00 | 5.10 | 2.03 | 0.95 | 0.80 | −0.19 | −0.10 | 0.78 | 164, 170 | 0.022, 0.011 |
| DCA16 | 20.00 | 5.75 | 2.13 | 0.94 | 0.83 | −0.14 | −0.08 | 0.81 | 216 | 0.011 |
| DCA18 | 12.00 | 4.25 | 1.68 | 0.94 | 0.76 | −0.23 | −0.12 | 0.73 | 157 | 0.011 |
| EMO90 | 7.00 | 2.50 | 1.14 | 0.41 | 0.60 | 0.32 | 0.22 | 0.53 | - | - |
| GAPU71B | 6.00 | 3.39 | 1.35 | 0.83 | 0.71 | −0.18 | −0.10 | 0.66 | - | - |
| GAPU101 | 9.00 | 3.10 | 1.50 | 0.57 | 0.68 | 0.15 | 0.13 | 0.65 | - | - |
| GAPU103A | 11.00 | 3.29 | 1.47 | 0.78 | 0.70 | −0.12 | −0.07 | 0.65 | - | - |
| UDO-043 | 14.00 | 3.09 | 1.65 | 0.48 | 0.68 | 0.29 | 0.21 | 0.66 | 184 | 0.011 |
| Mean | 11.60 | 3.81 | 1.61 | 0.76 | 0.72 | −0.04 | −0.01 | 0.69 |
| POP1 | POP2 | POP3 | ||
|---|---|---|---|---|
| Dest | POP1 | 0.000 | 0.001 | 0.001 |
| POP2 | 0.333 | 0.000 | 0.001 | |
| POP3 | 0.316 | 0.380 | 0.000 | |
| G’st (Hed) | POP1 | 0.000 | 0.002 | 0.002 |
| POP2 | 0.372 | 0.000 | 0.002 | |
| POP3 | 0.348 | 0.413 | 0.000 | |
| Fst * | POP1 | 0.000 | 0.001 | 0.001 |
| POP2 | 0.077 | 0.000 | 0.001 | |
| POP3 | 0.066 | 0.066 | 0.000 |
| POP | Na | Ho | He | F | nPA | PAf | |
|---|---|---|---|---|---|---|---|
| Malta | Mean | 11.60 | 0.80 | 0.81 | 0.01 | 5 | 0.013 |
| SE | 1.59 | 0.04 | 0.02 | 0.03 | |||
| POP1 | Mean | 17.10 | 0.84 | 0.82 | −0.02 | 15 | 0.006 |
| SE | 2.10 | 0.02 | 0.02 | 0.03 | |||
| POP2 | Mean | 18.80 | 0.83 | 0.82 | −0.01 | 33 | 0.007 |
| SE | 1.87 | 0.03 | 0.03 | 0.01 | |||
| POP3 | Mean | 14.40 | 0.83 | 0.73 | −0.13 | 7 | 0.004 |
| SE | 1.63 | 0.07 | 0.06 | 0.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valeri, M.C.; Mifsud, D.; Sammut, C.; Pandolfi, S.; Lilli, E.; Bufacchi, M.; Stanzione, V.; Passeri, V.; Baldoni, L.; Mariotti, R.; et al. Exploring Olive Genetic Diversity in the Maltese Islands. Sustainability 2022, 14, 10684. https://0-doi-org.brum.beds.ac.uk/10.3390/su141710684
Valeri MC, Mifsud D, Sammut C, Pandolfi S, Lilli E, Bufacchi M, Stanzione V, Passeri V, Baldoni L, Mariotti R, et al. Exploring Olive Genetic Diversity in the Maltese Islands. Sustainability. 2022; 14(17):10684. https://0-doi-org.brum.beds.ac.uk/10.3390/su141710684
Chicago/Turabian StyleValeri, Maria Cristina, David Mifsud, Clayton Sammut, Saverio Pandolfi, Emanuele Lilli, Marina Bufacchi, Vitale Stanzione, Valentina Passeri, Luciana Baldoni, Roberto Mariotti, and et al. 2022. "Exploring Olive Genetic Diversity in the Maltese Islands" Sustainability 14, no. 17: 10684. https://0-doi-org.brum.beds.ac.uk/10.3390/su141710684