
Bacterial Community Structure and Predicted Metabolic Function of Landfilled Municipal Solid Waste in China


MOE Key Laboratory of Soft Soils and Geoenvironmental Engineering, Institute of Geotechnical Engineering, Zhejiang University, Hangzhou 310000, China
*
Authors to whom correspondence should be addressed.
Sustainability 2022, 14(6), 3144; https://0-doi-org.brum.beds.ac.uk/10.3390/su14063144
Submission received: 16 December 2021
/
Revised: 19 February 2022
/
Accepted: 25 February 2022
/
Published: 8 March 2022
(This article belongs to the Topic Solid Waste Management)
Abstract
:The biodegradation behavior of municipal solid waste (MSW) depends on the diversity and metabolic function of bacterial communities, which are affected by environmental factors. However, the diversity of the bacterial communities and metabolic functions in MSW, as well as their influencing factors, remain unclear. In deep-aged MSW, the abovementioned deficiencies are more significant, and will effectively hamper landfill disposal. In this study, high-throughput sequencing was performed to examine the bacterial community structure and metabolic function from depths of 10 m to 40 m, of two large MSW landfills on the southeast coast of China. Thermotogota (1.6–32.0%), Firmicutes (44.2–77.1%), and Bacteroidota (4.0–34.3%) were the three dominant phyla among the 39 bacterial phyla identified in aged MSW samples. Bacterial genera associated with the degradation of many macromolecules, e.g., Defluviitoga, Hydrogenispora, and Lentimicrobium were abundantly detected in MSW samples, even in aged MSW. Redundancy analysis (RDA) showed that bacterial diversity in the landfills was most strongly correlated with electrical conductivity, age, and moisture content of the MSW. Tax4fun2 analysis predicted that there were abundant metabolism functions in aged MSW, especially functional enzymes (e.g., glycine dehydrogenase and cellulase) related to amino acids and cellulose degradation. This study increases our understanding of the bacterial diversity and functional characteristics in landfilled MSW.
1. Introduction
With the development of urbanism and the society–economy, a large amount of municipal solid waste (MSW) is produced globally every year [1,2,3]. Based on the latest available data, global waste generation in 2016 was estimated to reach 2.01 billion t [1]. Countries in East Asia and the Pacific regions produced the largest amount of MSW in the world, accounting for 23 percent by magnitude [1]. Around the world, almost 40 percent of MSW is disposed of in landfills [1]. For instance, over 69 percent of MSW is dumped in landfills in China [4]. In 2019, 200 large and medium-sized cities in China produced 236 million t of MSW, of which 109 million t were treated by landfilling [4].
The degradation and stabilization of landfilled MSW is a complex and long-term process of leading biological and physicochemical reactions [5]. It can be divided into three stages, a rapid degradation stage, a slow degradation stage, and a post-stabilization stage [5]. The stabilization process usually lasts for several decades under natural conditions [5,6]. For the slow degradation stage, the remaining organic matter from the previous stage is difficult to degrade [5]. Acceleration of the degradation process contributes to reducing the stock of waste as well as the pollution load [7,8,9]. Several methods, such as leachate recirculation, aeration, and the addition of exogenous microorganisms, have been proved to be effective in the acceleration of MSW degradation [7,8,9]. The degradation rate of MSW is closely related to the microbial community structure in landfills [10]. Therefore, it is of great significance to explore the bacterial community structure of landfill sites for a better understanding of MSW degradation.
The main components of aged MSW are soil-like materials and textiles; food waste is degraded in the rapid degradation stage [5,11]. Previous researchers have characterized bacterial strains and bacterial community composition in landfill leachate, solid waste, and cover soil using various approaches, including molecular library, denaturing gradient gel electrophoresis (DGGE), terminal restriction fragment length polymorphism (T-RFLP), and high-throughput sequencing [12,13,14,15,16,17,18,19,20,21,22,23,24]. As shown in Table 1, globally, Firmicutes, Proteobacteria, and Bacteroidetes are the dominant groups in solid waste [12,16,19,20,25,26,27,28,29,30,31]. Based on culture-independent approaches, previous studies have revealed that diverse bacterial genera associated with various functions, such as cellulolytic bacteria, acidifying bacteria and sulfate-reducing bacteria, were detected abundantly in landfill leachate [31]. Acidogen and acetogen groups of bacterial genera with different substrates were dominant in MSW samples that were landfilled to greater than 20 m [20]. However, the bacterial community composition in aged MSW in deep layers has yet to be fully elucidated because the thickness of landfill waste can reach tens of meters, making sample collection very difficult.
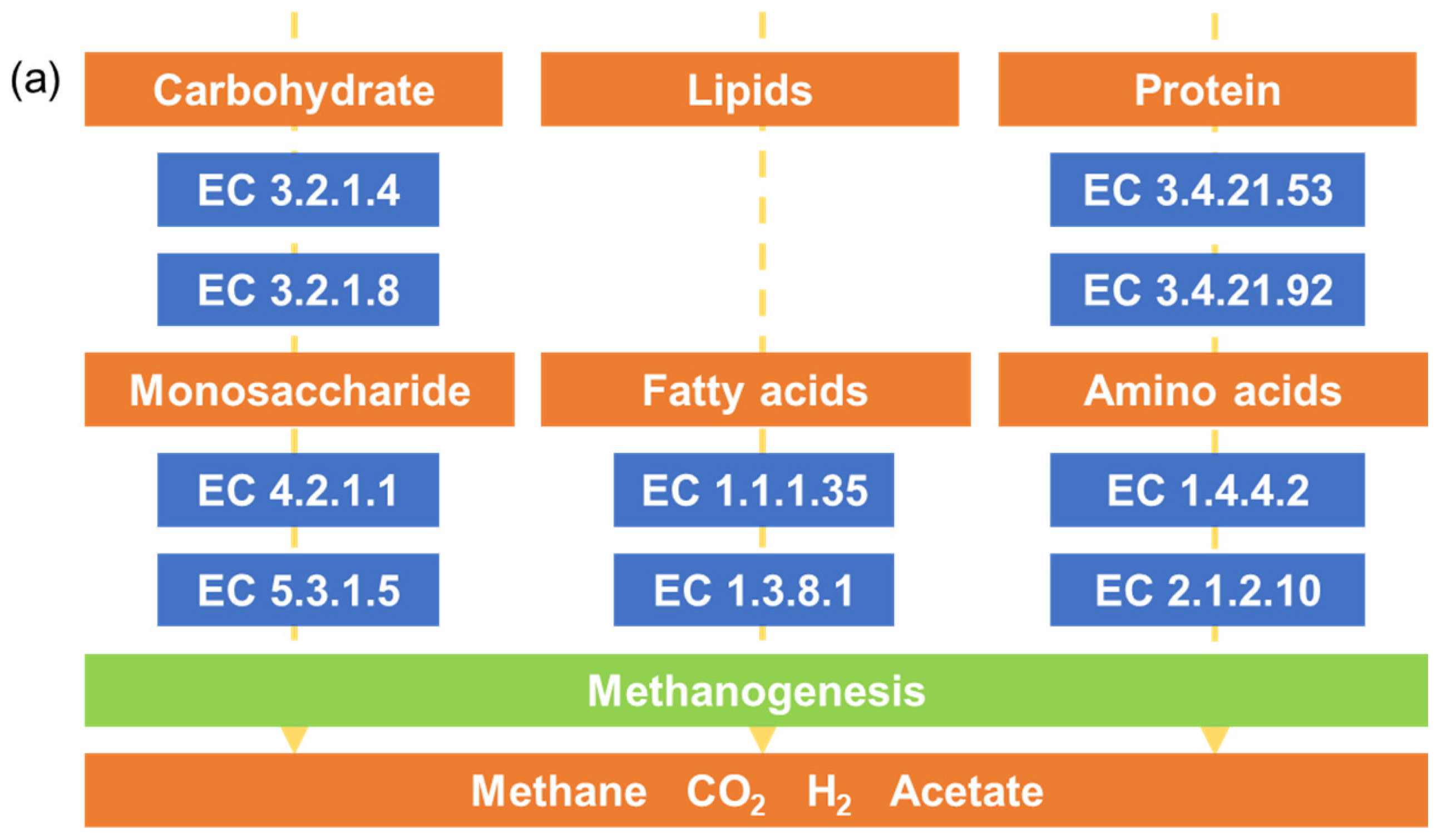
Bacterial metabolism and function are very important for the degradation of waste in landfills [10,31]. The pathways for processing macromolecular organic compounds, mainly contributed by carbohydrates, proteins, and lipids, into monosaccharides, amino acids, and fatty acids involve a variety of corresponding functional enzymes. The KEGG pathway of carbohydrate metabolism, amino acid metabolism, and energy metabolism were dominant in an anaerobic bioreactor [10,32]. Furthermore, glycine dehydrogenase (EC 1.4.4.2) and Endopeptidase La (EC 3.4.21.53), related to hydrolyzation of protein to amino acids, were more abundant than other functional enzymes [32]. Denitrification of functional genes was also abundant in a long-term operation lab-scale bioreactor [10]. Therefore, using bioinformatics tools to predict metabolic function during solid waste decomposition can provide more information on bacterial community activity during solid waste decomposition, which is useful for landfill management [10].
In this study, a high-throughput sequencing method based on the Illumina Novaseq6000 platform was used to assess bacterial diversity and predicted the functional capacity of aged MSW samples drilled from two typical landfill sites in China. The data were further analyzed using QIIME2 software to discuss the microbial community structure and were processed using the Tax4fun2 method to predict functional genes. This study aims at identifying the core bacterial community and the core functional genes of landfilled MSW in the middle and late degradation stages.
2. Materials and Methods
2.1. Samples Collection
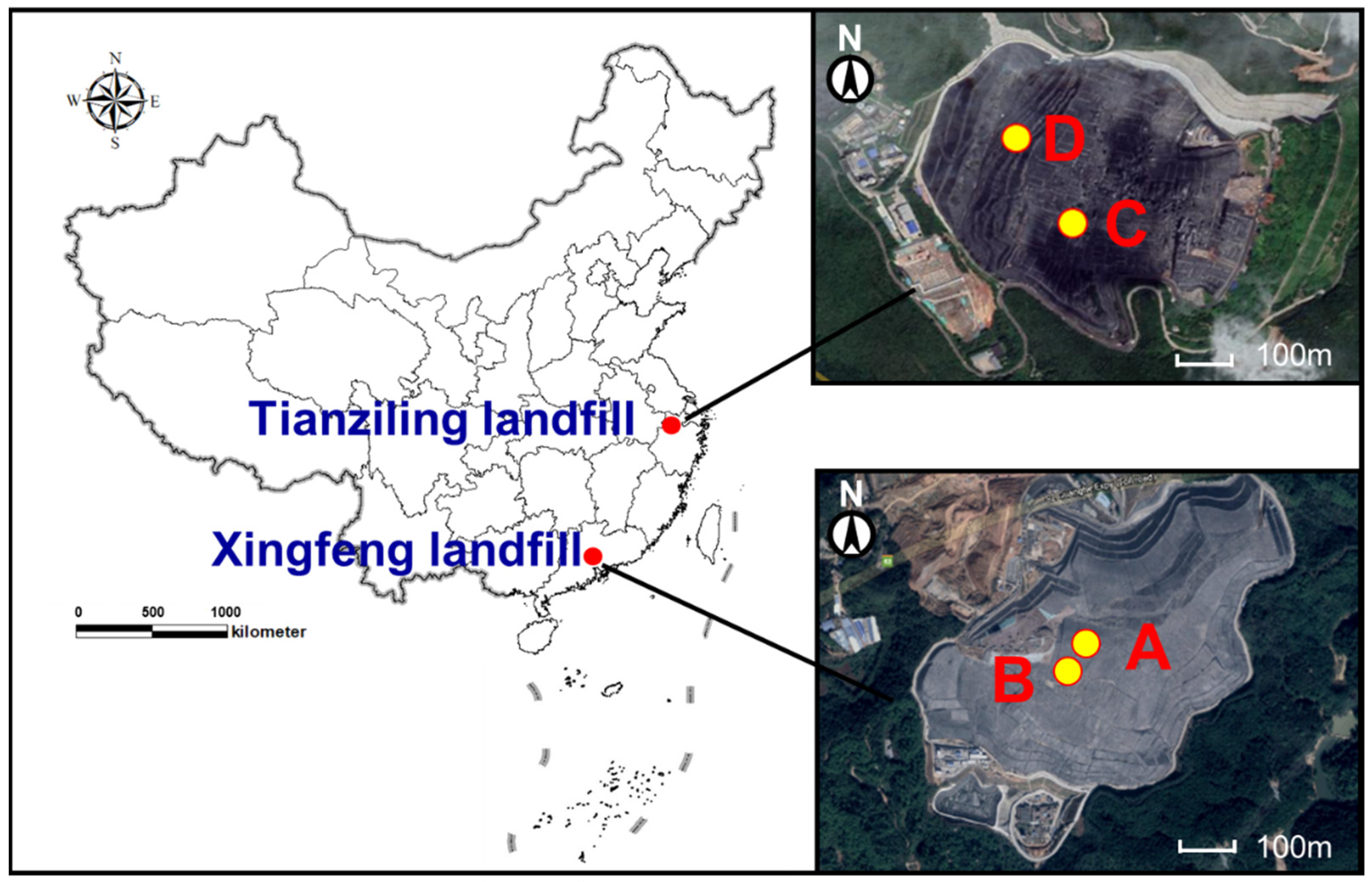
MSW samples were collected from Xingfeng landfill and Tianziling landfill, on the southeast coast of China (as shown in Figure 1). The Xingfeng landfill is located in the northern suburb of Guangzhou City (22°26′–23°56′ N, 112°57′–114°3′ E), in the south subtropical zone. It has a designed service life of 25 years and mainly collects MSW from the downtown area. This landfill was put into operation in 2002 and has a daily treatment capacity of 11,000 t [33]. Tianziling landfill is located in the eastern suburb of Hangzhou City (29°11′–30°33′ N, 118°21′–120°30′ E), in the mid-subtropical zone. This valley-type landfill was put into operation in 1991 and has a designed capacity of 6 billion m3. Recently, 4000–6000 t have been MSW transported and dumped per day [34]. As shown in Table 2, the proportion of food waste in these two large-scale landfills is greater than 50%, which indicates that they are typically high food-waste content municipal solid waste (HFWC-MSW) landfills. The remainder is mostly made up of plastics, paper, and textiles (more than 35%) [33,34].
For each landfill, samples were drilled from depths of 10 m to 40 m, at an interval of 10 m, at two different sites. Approximately 10 kg of MSW was collected at each sampling site, then subsampling was conducted using the quartering method. In total, 16 samples were collected, labeled A1, A2, A3, A4, B1, B2, B3, B4, C1, C2, C3, C4, D1, D2, D3, D4; labels “A, B, C, D” refer to site A, site B, site C, and site D; whereas labels “1, 2, 3, 4” refers to the depths of 10 m, 20 m, 30 m, and 40 m, respectively. Each sample was collected with three replicates. The subsamples were sorted to remove the inert components, including stone, metal, plastic fabrics, etc. [20]. A total of 100 g of biological samples, passed through a 10-mesh sieve, were put into centrifuge tubes, which were stored in a bucket filled with dry ice (temperature of −78.5 °C). The MSW samples, in triplicate, were transported to the laboratory and stored at a temperature of −80 °C, and prepared for sequencing. A landfill age of MSW > 5a was defined as aged MSW in this study.
2.2. Physical and Chemical Analyses
The physicochemical properties of all samples were analyzed, including moisture content, pH, electrical conductivity (EC), organic matter content, and ammonium. In order to measure the moisture content, MSW samples were dried at an oven temperature of 105 °C for 6 h to prevent ignition loss of organic matter, according to technical code CJ/T313-2009 [35]. The organic matter content (kg/kg, dry weight) of each specimen was measured using the burning loss method (i.e., heating at 600 °C for 6 h to achieve a constant mass) in accordance with technical code CJ/T96-2013 [36]. A soil–water (1:5, soil/water) suspension was prepared 30 min prior to EC and pH measurements. Ammonium in samples was displaced by 100 mL of 0.01 M CaCl2 for 60 min. The extract was filtered, and ammonia nitrogen was determined using Nessler’s reagent spectrophotometry [19,37].
2.3. DNA Extraction and Sequencing
The DNA in each sample was extracted using a MinkaGene Soil DNA Kit (MinkaGene, Guangzhou, China). PCR reactions were conducted in triplicate in a 50 uL mixture containing 25 uL premix (TaKaRa Premix Taq, Version 2.0, TaKaRa Biotechonlogy, Dalian Co., Dalian, China), 50 ng DNA templates, and 1 uL (20 mM) of forward and reverse primers. The mixtures were denatured at 94 °C for 5 min and were then amplified for 35 cycles of 94 °C for 30 s, 52 °C for 15 s, and 72 °C for 30 s, with a final extension at 72 °C for 10 min.
The V4–V5 region (420 bp) of the 16S rRNA gene was amplified by PCR using the following primers [38]:
Forward 515F: GTGCCAGCMGCCGCGGTAA
Reverse 907R: CCGTCAATTCMTTTRAGTTT
Equal quantities of the three PCR products from each sample were mixed for Pair-end sequencing, according to the NEBNext® Ultra™ II DNA Library Prep Kit for Illumina® (New England Biolabs, MA, USA). The library quality was assessed with Qubit@ 2.0 Fluorimeters (Thermo Fisher Scientific, Waltham, MA, USA). The standard process was used for database construction. Primers were removed using Cutadapt software, according to the primer information at the beginning and end of the sequence to obtain paired-end clean reads [39]. The standard process was used for database construction, and the raw data were deposited in the NCBI short reads archive database (accession number: PRJNA698338).
2.4. High-Throughout Data Processing
Analyses were performed in QIIME2 using standard procedures [40]. The data were imported using the import command with the EMPPairedEndSequences type using the demux command. The sequences were trimmed and quality-filtered using the DaDa2 plugin [40,41]. The 248 bp forward reads and 236 bp reverse reads were truncated with a high quality score (lower whisker was 25). The minimum number of sequence reads required to retain a sample in the study was set at 76,882 (to maximize the number of samples retained). An ASV table was generated, made up of rarefied counts. The align-to-tree-mafft-fasttree pipeline was then applied to build a phylogenetic tree. Diversity analyses, including α-diversity and β-diversity, were conducted using the Q2-diversity plugin and PAleontological STatistics (PAST) software (v4.02) [40]. The Q2-feature classifier plugin was used for the taxonomic composition of samples, based on the classifier trained on the Silva 138_99% operational taxonomic units (OTU) database [40].
In order to evaluate the influence of geographical location on bacterial community structure, the genera (with a relative abundance > 1.0% in at least one sample) were selected to draw a heat map. A Venn diagram was created to show both the unique and shared amplicon sequence variants (ASVs) of landfills at different sites. Based on the Bray–Curtis distance, principal coordinate analyses (PCoA) and adonis analyses were performed in R software (Version 4.1.2) using the Ape package (Version 5.6-1) [40,42,43]. Detrended correspondence analysis (DCA) and redundancy analysis (RDA) the used Vegan package (Version 2.5-6) in R software (Version 4.02) [43,44]. The species data (same with the heat map) were transformed using the Hellinger method. Since the length of axis 1 of DCA < 3.0, RDA was performed to analyze the impact of the physicochemical attributes on the variance in the community structure [45,46]. Environmental factors were selected based on the variation inflation factors (VIFs), which removed redundant attributes stepwise, resulting in VIFs < 10 [17,20]. The significance of the RDA model was tested using 999 Monte Carlo permutations [45]. A p value < 0.05 was considered statistically significant.
2.5. Bacterial Community Function Analysis
Tax4Fun2 was adopted to predict the functional capabilities of bacterial communities based on 16S rRNA gene amplicon data [47]. The functional annotation of Tax4fun2 predictions was obtained based on the Kyoto Encyclopedia of Genes and Genomes (KEGG) database [47]. Tax4Fun2 outputs relative abundances by default, so no transformations were required after producing the prediction profile [47]. A table of KEGG orthologs (KOs) and metabolic pathways were produced for each dataset. The relative abundance of functional enzymes shown in Table S1 were calculated in Microsoft Excel software [32].
3. Results
3.1. Physical and Chemical Properties
The physical and chemical properties of each sample are summarized in Table 3. Generally, based on the rapid degradation stage, which usually lasts for 2–3 years, samples with landfill age >5 years were separated as aged MSW in this study [5]. The average moisture content of aged MSW samples (31%) was significantly lower compared than those that were less than 5 years old (51%, average of A1, B1, D1). The average organic matter content of aged MSW samples (26%) was lower compared to those less than 5 years old (29%, average of A1, B1, D1). The pH of stored waste was neutral or slightly alkaline (6.6–8.0). The EC among samples from the Xingfeng landfill (28.0–35.1 ms/cm) was slightly higher than that of the Tianziling landfill (18.6–30.1 ms/cm) indicating more soluble salt concentration.
3.2. Sequencing Results and Microbial Diversity
In total, 1,230,112 effective sequences were obtained from high-throughput sequencing and a total of 1113 ASVs were assigned. The ASV coverage rate exceeded 99% for each sample, indicating that exhaustive information on the bacterial communities was obtained. In order to indicate the diversity and coverage of samples, the α-diversity of each sample was calculated (as listed in Table 4). The species richness was estimated using Shannon and Chao-1 diversity indices. The Shannon diversity index varied greatly among the samples, where sample D1 had the highest Shannon diversity index value (4.87) and sample C2 had the lowest (2.51). The community evenness of the 16 samples was estimated using Simpson and evenness diversity indices. Sample D1 had the highest Simpson (0.983) and evenness (0.281) index values, whereas sample D4 had the lowest Simpson (0.868) and evenness (0.079) index values. The α-diversity of samples at different depths varied greatly. Samples at 20 m had the lowest Shannon diversity index values (3.54, average of A2, B2, C2, and D2), Simpson value (0.876, average), and evenness diversity index value (0.110, average). Whereas samples at 30 m had the highest Chao-1 diversity index value (434.7, average of A3, B3, C3, and D3) and Simpson diversity index value (0.950, average).
3.3. Bacterial Taxonomic Composition
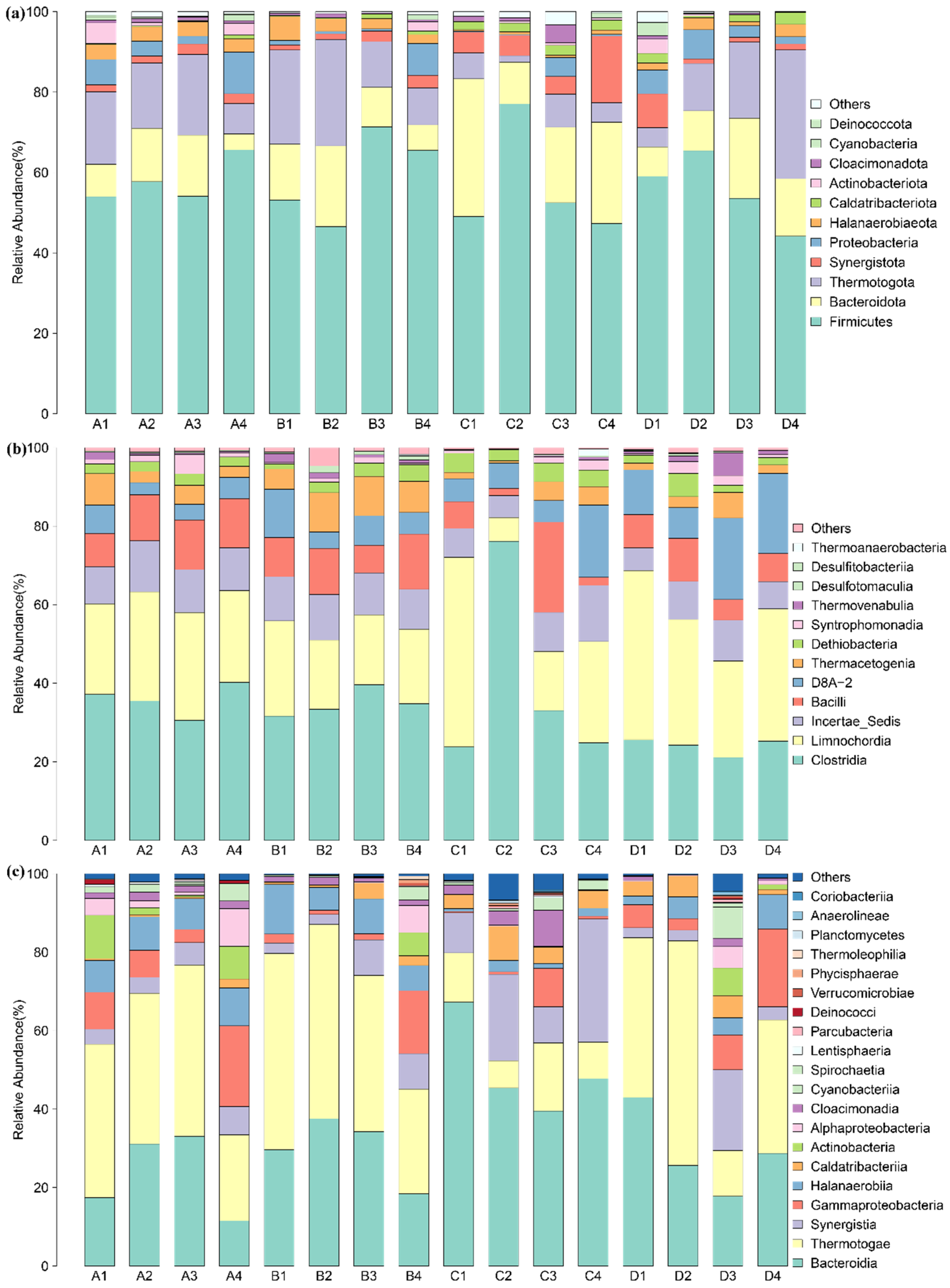
High-throughput sequencing revealed the diversity of the microbial community in different samples at the phylum level. In total, 39 phyla were found in these two landfills. As shown in Figure 2a, the dominant phyla across all samples, with an average relative abundance ≥ 1.0%, were Firmicutes (44.2–77.1%), Bacteroidota, (4.0–34.3%), Thermotogota (1.6–32.0%), Synergistota (1.2–16.6%), Proteobacteria (0.0–9.8%), Halanaerobiaeota (0.2–3.0%), Caldatribacteriota (0.2–3.0%) and Actinobacteriota (0.0–5.2%). Firmicutes, Bacteroidota, and Thermotogota were the most abundant phyla in all samples, accounting for 71.1–93.1% of the total sequences in the different samples. Others, such as Proteobacteria and Synergistota, were low in all the samples. Samples at 10 m had the lowest relative abundance of Firmicutes (53.8%, average of A1, B1, C1, and D1) and the lowest relative abundance of Thermotogota (13.2%). Samples at 20 m had the highest relative abundance of Firmicutes (61.7%, average of A2, B2, C2, and D2).
A total of 88 classes were detected. As shown in Figure 2b, Clostridia (11.2–58.3%) accounted for the greatest part of phylum Firmicutes. Thermotogota consists of Thermotogae. As shown in Figure 2c, Bacteroidia (3.9–34.0%) accounted for the largest proportion among other phyla. Synergistia accounted for more in the Tianziling C samples than in other samples, while samples from the Xingfeng landfill showed more Halanaerobiia. Petrotogales (accounting for 99.9% of class Thermotogae) and Kosmotogales were determined. Defluviitoga (accounting for 99.9% of order Petrotogales) accounted for the vast majority while Mesotoga only accounted for a minor part.
3.4. Impact Factors on Bacterial Community Structure
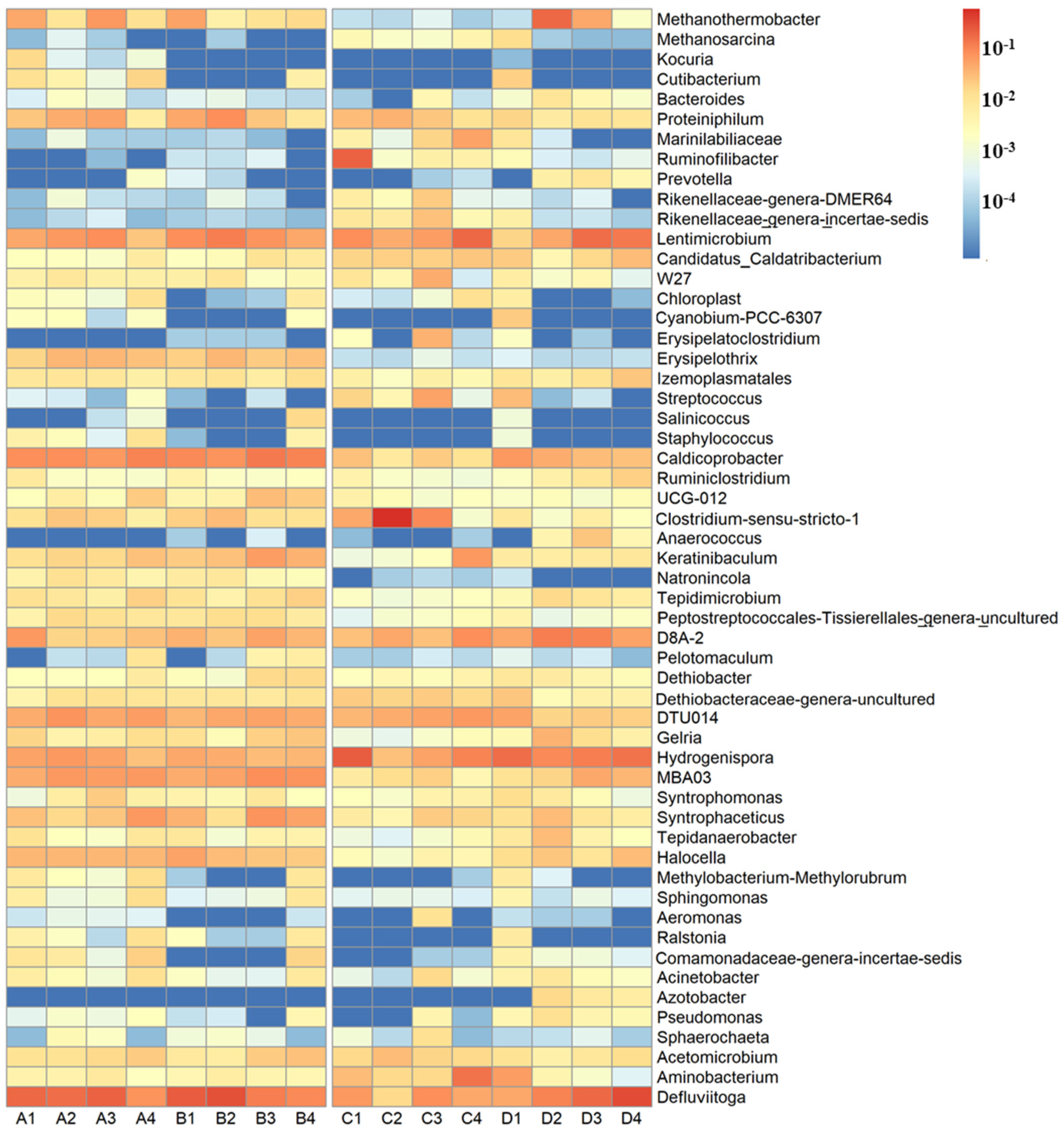
The impact of physicochemical properties, deposit age, and geographical location on bacterial community structure was analyzed using a heat map, PCoA, Venn diagram, and RDA analysis. A total of 471 genera were detected, of which 47 genera were found in all the samples, accounting for 68.9–94.9% of the total sequences. A total of 98 genera were only determined at the Xingfeng landfill, accounting for 0.0–2.5% of the total sequences, whereas 67 genera were only determined at the Tianziling landfill, accounting for 0.0–3.0% of the total sequences. Figure 3 shows the genera with a relative abundance > 1.0% in at least one sample, revealed by the universal primer pair. For example, Defluviitoga (1.6–31.8%), Hydrogenispora (2.8–21.6%), Lentimicrobium (1.8–17.3%), and Caldicoprobacter (0.8–12.5%) were abundant in all samples. The relative abundances of Sphingomonas (0–1.4%) and Ralstonia (0–1.2%) in samples from the Xingfeng landfill were much higher than samples from the Tianziling landfill (lower than 0.5%). The relative abundances of Marinilabiliaceae (1.4–5.3%) and Ruminiclostridium (0.1–2.0%) in samples from the Tianziling landfill were much higher than samples from the Xingfeng landfill (all of them were lower than 0.1%).
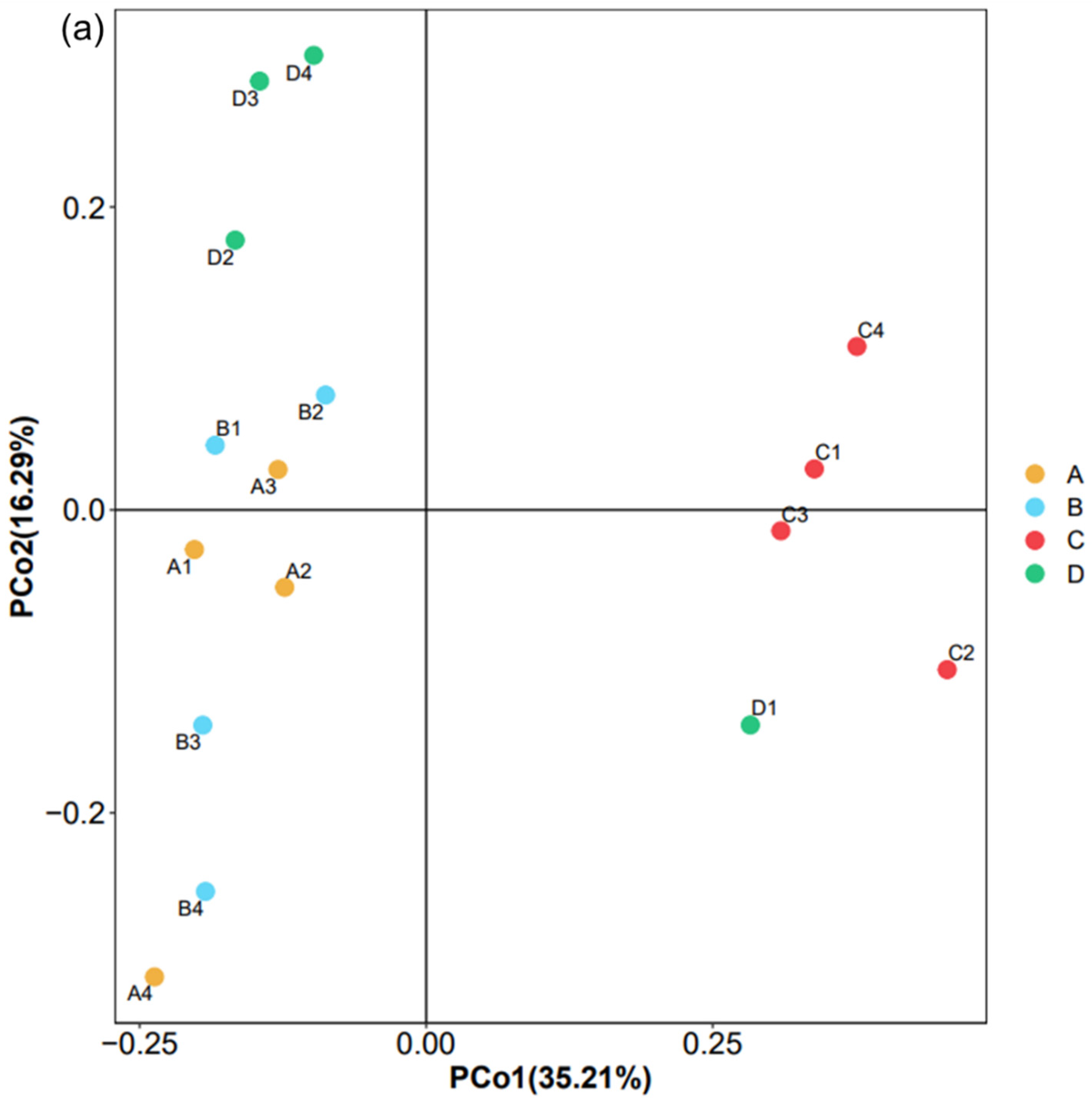
PCoA was conducted to show the relationship between samples and species. As shown in Figure 4a, the two axes explained 51.50% of the variance in microbial community abundance. Adonis analysis indicated that the geographical regions affected the bacterial community composition to a certain extent (adonis: R2 = 0.422, p = 0.001). Most of the samples from the same sampling site could be clustered together, indicating that the microbial diversity and structure of the landfill were more affected by the sampling site than the deposit depth. Samples from a site with an older age may form unique bacterial communities, e.g., samples from site C were isolated from others. While other samples from both the same or different landfills with a similar history might have a certain similarity, such as sites A, B, and D.
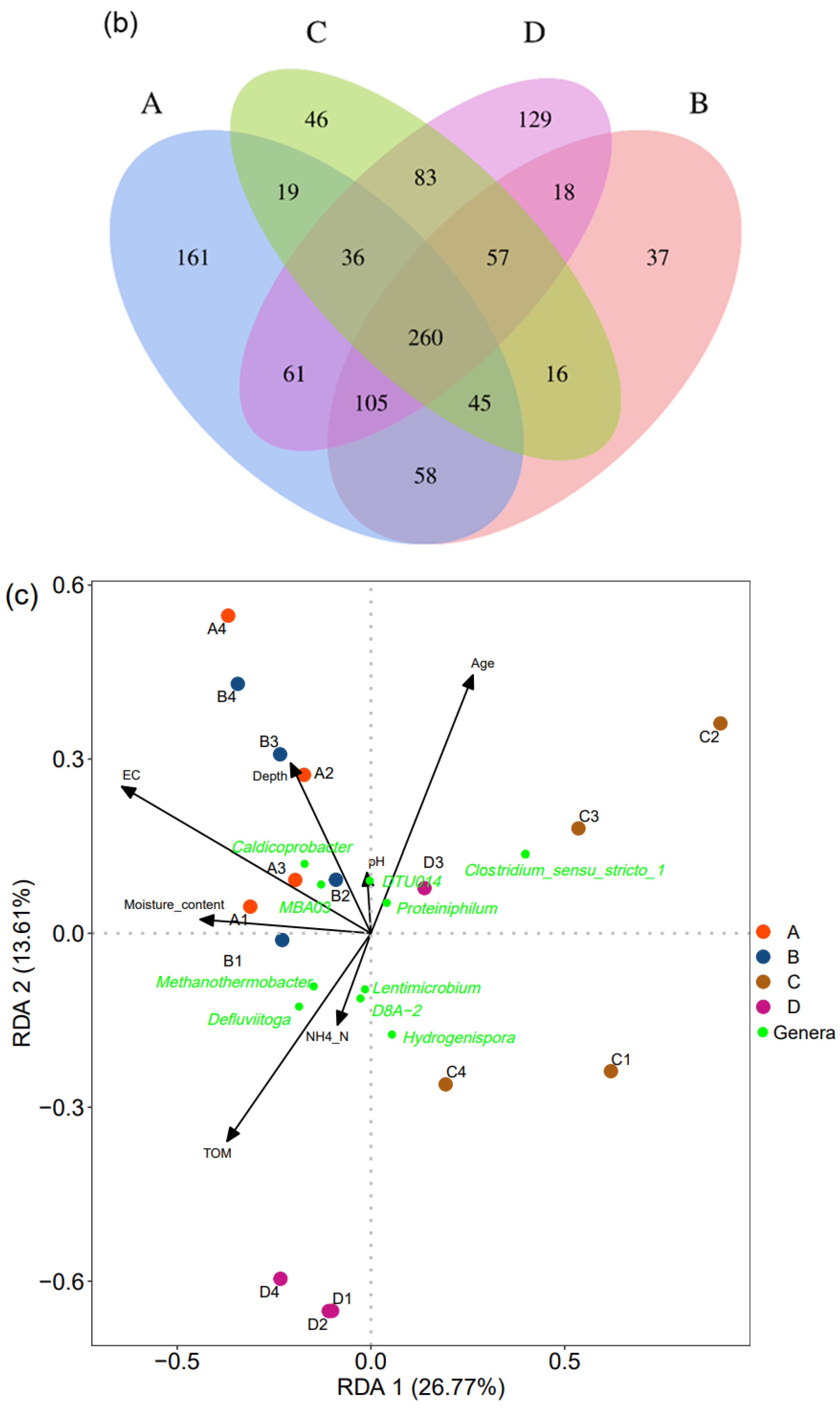
Venn diagram showed that the four stands shared 260 ASVs, accounting for 88.7% effective sequences (Figure 4b). The two stands in Xingfeng landfill shared 468 ASVs and those in Tianziling landfill shared 436 ASVs. Although in different landfills, Groups A and D shared 462 ASVs, while A and C only shared 360 ASVs.
In order to further discern the possible relationship between physicochemical properties of MSW and bacterial community variance, RDA (Figure 4c) was conducted. It represented the correlation degree between environmental factors and bacterial community structure. In total, seven physicochemical attributes, organic matter content, depth, ammonium, moisture content, pH, and EC, listed in Table 3, were selected for RDA. The first axis explained 26.8% of the variance of the species–environment relation, and the 40.4% variance was explained by the first and second axes. Based on 999 permutations of the Monte Carlo test, these factors were significantly correlated with microbial communities (F = 1.99, p = 0.002). Among the selected attributes, EC, age, and moisture content (p < 0.05) were considered to affect the bacterial community structure most significantly. Five genera, including Lentimicrobium and D8A-2 in the third and delta quadrant, were strongly correlated with age. Clostridia_MBA03 and Caldicoprobacter were strongly correlated with EC.
3.5. Functional Analysis of Bacterial Community
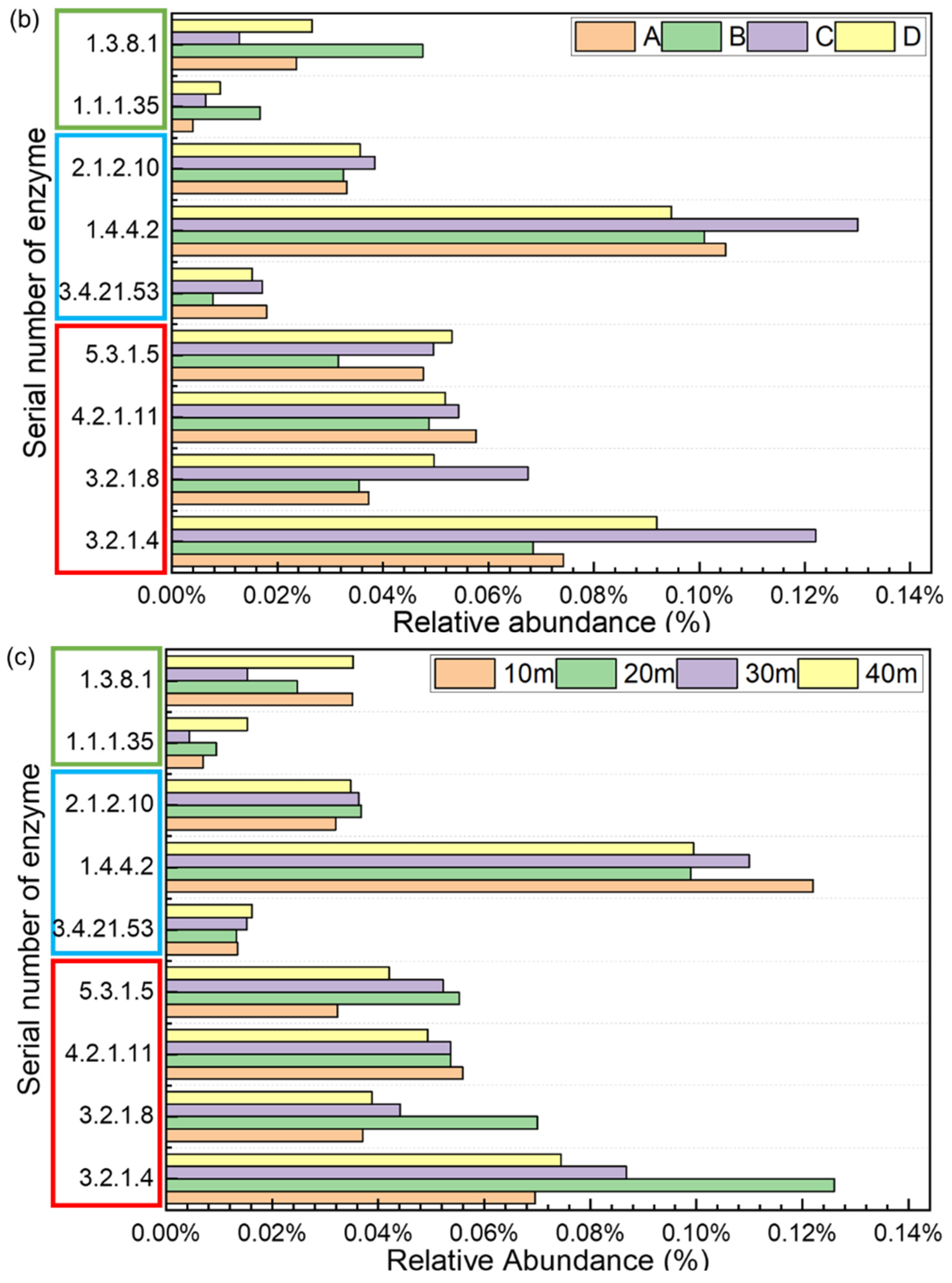
Tax4fun2 was used to predict the metagenomics functional composition, based on 16S rRNA marker genes. The predicted sequences mainly belonged to metabolism (71.22–77.05%), environmental information processing (8.26–12.27%), genetic information processing (4.39–6.09%), cellular processes (5.38–7.76%), human diseases (2.09–3.08%) and organismal systems (0.86–1.32%) categories. In the metabolism category at KEGG level 2, three subfunctions, namely, amino acid metabolism (5.7–8.0%), carbohydrate metabolism (9.7–11.9%), and energy metabolism (3.7–4.8%), showed the highest abundances compared to those of the other subfunctions. The pathways for processing carbohydrates, proteins, and lipids into monosaccharides, amino acids, and fatty acids to generate methane are shown in Figure 5a [32]. Figure 5b,c show the relative abundances of the corresponding functional enzymes in the three pathways. Among them, glycine dehydrogenase (EC 1.4.4.2) and cellulase (EC 3.2.1.4) were more abundant than the other functional enzymes. The 20 m group had the highest relative abundance of cellulase (EC 3.2.1.4), endo-1,4-beta-xylanase (EC 3.2.1.8), and xylose isomerase (EC 5.3.1.5) among groups with different depths. Samples from the Tianziling landfill (Group C and D) had more corresponding functional enzymes related to carbohydrate degradation than samples from the Xingfeng landfill (Group A and B). In addition, as shown in Table S2, denitrification related functional genes were abundant, e.g., nar (0.003–0.038%), nir (0.007–0.093%), nor (0.001–0.016%), and nos (0–0.003%). Samples in Group D had the highest average of relative abundance of nos (0.0016%), nar (0.0272%), and nir (0.0532%).
4. Discussion
Most samples had the characteristics of aged MSW. The physicochemical properties of MSW varied with storage depth and age, especially in terms of organic matter content. The rapid degradation stage lasts for about 2–3 years and then the slow degradation stage lasts for about 20–30 years [5,6,48]. Therefore, according to age, the degradation stage of most samples was categorized into the slow degradation stage, while A1, B1, and D1 were in the rapid degradation stage [5,6,48]. The fine fractions in MSW were formed by complex biochemical reactions from the degradation of organic waste over an extended period of time [49]; it had the properties of black soil and increased with deposit age. Due to the leachate generated by degradation, the moisture content of fresh HFWC-MSW is generally high and, differently, the moisture content of aged MSW varied with deposit depth in the two landfills differently, which may be due to the main leachate layer and partially blocked leachate mound in the waste body. The EC of stored waste was lower than that which has been reported previously [19].
Previous research found that Firmicutes, Bacteroidota, and Proteobacteria are the three most abundant phyla in landfills, accounting for 66–90% of total sequences [18,19,20]. Different from the results obtained by previous scholars, in this study, Firmicutes, Bacteroidota, and Thermotogota were the three main phyla. Firmicutes are mainly responsible for hydrolysis and fermentation in landfills. They promote the metabolism of cellulose, proteins, lignin, and lipids [20,31]. Bacteroidota is known for its excellent hydrolysis ability [17]. The biological function of Bacteroidota mainly includes hydrolytic and fermentation, which can degrade carbohydrates, proteins, and lipids [50]. The low relative abundance of Proteobacteria may be due to these bacteria being more likely to be enriched in leachate rather than landfill waste [20,25]. A high temperature may be the reason for the increase in the relative abundance of Thermotogota. Gas well temperature test results of 27 landfills in Florida showed that 74% of gas wells have a temperature higher than 55 °C [51]. The test results of the temperature of the Laogang landfill (Shanghai, China), showed that the MSW temperature reached 42–55 °C. A high-temperature MSW could lead to a decrease in other bacteria and the proliferation of thermophilic bacteria. The high temperature was caused by heat generation from the process of waste degradation and external heat due to the subtropical monsoon climate. In addition, the deep MSW layer could play an important part in heat insulation. Most of the species of Thermotogota were isolated from high-temperature environments, such as wastewater, underground gas reservoirs, coastal sediments, etc., in which optimal growth temperature can be as high as 55 °C. Most of these can degrade carbohydrates extensively, reduce sulfur compounds (thiosulfate and sulfur) to hydrogen sulfide and produce yeast extract [52,53]. However, the temperature of samples could not be measured because of some technical limitations during the sampling progress. The growth of Defluviitoga could create a positive response to the increasing temperature during the landfill progress [53,54]. Some members of Defluviitoga in the thermophilic anaerobic digestion system could effectively degrade organic materials, metabolize sugars, and generate hydrogen [53,55,56].
Generally, geographical locations and heterogeneous physicochemical properties of MSW are considered the main factors affecting the bacterial community structure in landfills [17,20,31]. The factors contributing to the geographical differences may depend on factors, such as climatic conditions, waste composition, and landfill procedures [20]. In this study, climatic conditions and composition of MSW in two landfills were similar. The age of the MSW at a corresponding depth was similar at site A, site B, and site D. The similarity in bacterial composition also suggests that the samples had similar ages, based on studies showing that differently aged MSWs harbor distinct bacterial communities [10,20]. With increasing landfill age, the degradation of macromolecules can influence the response of the microbial communities [31]. However, the difference in bacterial composition between sites C and D seems slightly greater, partly due to the different ages of MSW at the top layer. The separation or clustering between samples depended on the unique or shared abundant OTUs in MSW samples [18]. For instance, the clustering of A, B, and D may be driven by their shared abundant ASVs, belonging to Firmicutes, Bacteroidota, and Thermotogota. The Venn diagram also confirmed that the vast majority of ASVs were shared in four sites. The ASVs that were only detected in one site were rare ASVs. Although there were still many kinds of unique ASVs in each site, the relative abundance was low, suggesting that the composition of bacteria in different sites was similar.
The factors above could result in variations in MSW physicochemical properties, such as EC, age, and moisture content in different landfills. The RDA revealed some statistically significant relationships between the bacterial community and physicochemical attributes of samples. The results showed that the community compositions in this study were significantly affected by three physicochemical attributes. Among these attributes, electrical conductivity (EC) appeared to be the most significant environmental factor (p < 0.05), followed by age and moisture content. Similar results have been obtained in previous studies [19]. The EC of MSW leachate had a high linear correlation with its salt content, which affects the survival of many species. Most of the research showed that salt affects the availability of water or the metabolic process of bacteria [19,57,58,59]. The addition of copper salts during thermophilic anaerobic digestion of swine manure resulted in Clostridia_MBA03 enriched in the end, which is in line with the RDA analysis [60]. Age and moisture content are also considered to be key factors affecting the distribution of microorganisms in landfills [16,20]. The moisture content can influence the enzyme activities directly, thereby influencing the bacterial community structure. In addition, NH4+-N, NO2−N, pH, and dissolved organic carbon (DOC) were also considered to be significant environmental factors [20]. The total nitrogen was found to be related to some specific taxa and increasing NH4+-N significantly influenced the community structures of the total bacteria [20,61,62]. The pH is believed to be an independent driver of bacterial diversity, and it can serve as a predictor of the bacterial community structure in soil. The intracellular pH of most bacteria is usually within 1 pH unit of neutral [63]. In this study, the pH was neutral or slightly alkaline (6.6–8.0), and this may be the reason why the impact of pH was not significant. DOC is an actively cycled carbon pool, which is the direct precursor of bacteria growth and activity, thereby driving the decomposition processes [20].
The degradation of MSW is a complex reaction process. Cellulolytic bacteria such as Clostridium depend on the hydrolysis of cellulose into usable sugars by cellulase [64]. Acidifying bacteria, such as Prevotella, play an important role in the acidogenic fermentation of organic solid waste and contributes significantly to the production of volatile fatty acids [65]. The hydrogen-producing bacteria, e.g., Candidatus Cloacamonas, Syntrophomonas and Petrimonas produce acetate, hydrogen, and CO2 by degrading amino acids, lactate, succinate, propionate, etc. [31]. Finally, methanogens, e.g., Methanosarcina and Methanomassiliicoccus, utilize hydrogen and formate or acetic acid to produce methane in cooperation with other bacteria [31,66]. Landfilled waste contains a large amount of cellulose, which provides a sufficient carbon source for the growth and reproduction of microorganisms [6,67]. Similar to what was found by other researchers, a majority of the metabolic pathways were associated with carbohydrate metabolism, amino acid metabolism, and energy metabolism in the MSW samples [10]. Moreover, functional enzymes corresponding to cellulose degradation (e.g., cellulase) were still abundant in aged MSW, which agreed well with the decomposition behavior of cellulose [6,64]. The landfill over 5 years, or even 10 years, still has a relatively high abundance of cellulose [6,67,68]. The stabilization of a landfill may be over 20 years [67]. Therefore, even in aged MSW, abundant cellulase may suggest that degradation may be still relatively active, which is in line with the high temperature in aged landfills reported elsewhere [51]. In addition, the nitrogen microbial cycle is one of the most important element cycles during solid waste decomposition, and nitrogen management in landfills is essential for downstream nitrogen removal in leachate and green gas (N2O) emission control [10]. The abundant nar, nir, nor, and nos, suggesting denitrification, may be still abundant in MSW samples. Many previously reported denitrifying bacteria belong to phylum Proteobacteria, e.g., Pseudomonas and Acinetobacter [69]. Groups D also had the highest average of Pseudomonas (0.6%) and Acinetobacter (0.5%) which is in line with the higher relative abundance of denitrification genes. The results provide the potential functional genes information of carbon and nitrogen elements recycled in landfills.
Although plenty of work has been done on the bacterial diversity and metabolic function of landfilled MSW, there are still some limitations in this study. For instance, due to the difficulty in field sampling and the lack of instruments, some environmental factors such as the temperature are not tested. The influence of temperature on the bacterial community structure is worthy of further study.
5. Conclusions
Through high-throughput sequencing, the structure and functional compositions of the in situ microbial communities of aged MSW were determined. Firmicutes, Bacteroidota, and Thermotogota were identified as the main phyla in landfills, accounting for 71.1–93.1% of the representative sequences. Due to the similar composition of MSW, landfill history, and climate conditions, the bacteria community structure in different sub-region or landfills showed a high similarity. The RDA analysis further demonstrated that electrical conductivity, age, and moisture content were the main factors affecting the community. Among these, electrical conductivity, corresponding to salinity content, was the most important factor. Tax4fun2 analysis predicted that there were still abundant metabolism functions in aged MSW samples, especially amino acid metabolism, carbohydrate metabolism, and energy metabolism. Abundant carbohydrate metabolism genes regulate the degradation of lignin and cellulose in landfills, which is of great significance to the stabilization of landfilled MSW. This study enhances our understanding of the composition and functional characteristics of the bacterial communities in landfill MSW and provides guidance for the management of landfilled MSW.
Supplementary Materials
The following supporting information can be downloaded at: https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/su14063144/s1, Table S1: The functional enzymes related to macromolecular metabolism; Table S2: The relative abundance of functional genes and enzymes related to the nitrogen cycle or macromolecular metabolism in different sites.
Author Contributions
H.K.: conceptualization, supervision, resources. J.L.: investigation, software, methodology, writing—original draft. X.Z.: data curation. J.H.: visualization, formal analysis, writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This work is funded by the National Key Research and Development Program of China (Grant No. 2019YFC1806000), the National Natural Science Foundation of China (Grant No. 52108348), and the China Postdoctoral Science Foundation (Grant No. 2021M692836), which are gratefully acknowledged.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated or analyzed during this study are included in this published article.
Acknowledgments
We thank to Jingyu Peng, Yingyue Han, Yuxiang Zhang, Siyu Zhang, graduate students of Zhejiang University, for their assistance in the sampling and testing process. We are also grateful to the Guangdong Magigene Biotechnology Co., Ltd. China for assisting in the high-throughput sequencing.
Conflicts of Interest
The authors declare no conflict of interest.
References
- World Bank. What a Waste2.0: A Global Snapshot of Solid Waste Management to 2050. Available online: http://hdl.handle.net/10986/30317 (accessed on 24 February 2022).
- Jingxia, Y. Municipal solid waste (MSW)-to-energy in China: Challenges and cost analysis. Energy Sources Part B Econ. Plan. Policy 2017, 13, 116–120. [Google Scholar] [CrossRef]
- Saini, J.K.; Kumar, G.; Singh, S.; Kuhad, R.C. Municipal solid waste biorefinery for sustainable production of bioenergy. In Waste Biorefinery; Elsevier: Amsterdam, The Netherlands, 2021. [Google Scholar]
- National Bureau of Statistics, China. Annual Report; National Bureau of Statistics: Beijing, China, 2020. (In Chinese)
- Chen, Y.; Xu, W.; Ling, D.; Zhan, L.; Gao, W. A degradation–consolidation model for the stabilization behavior of landfilled municipal solid waste. Comput. Geotech. 2020, 118, 103341. [Google Scholar] [CrossRef]
- Barlaz, M.A. Forest products decomposition in municipal solid waste landfills. Waste Manag. 2006, 26, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Ge, S.; Ma, J.; Liu, L.; Yuan, Z. The impact of exogenous aerobic bacteria on sustainable methane production associated with municipal solid waste biodegradation: Revealed by high-throughput sequencing. Sustainability 2020, 12, 1815. [Google Scholar] [CrossRef] [Green Version]
- Reinhart, D.R.; McCreanor, P.T.; Townsend, T. The bioreactor landfill: Its status and future. Waste Manag. Res. J. Sustain. Circ. Econ. 2002, 20, 172–186. [Google Scholar] [CrossRef]
- Valencia, R.; van der Zon, W.; Woelders, H.; Lubberding, H.; Gijzen, H. The effect of hydraulic conditions on waste stabilisation in bioreactor landfill simulators. Bioresour. Technol. 2009, 100, 1754–1761. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Song, L. Succession of bacterial community structure and metabolic function during solid waste decomposition. Bioresour. Technol. 2019, 291, 121865. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Wu, T.; Wang, X.; Qiu, Z.; Lan, J. Study on compressibility and settlement of a landfill with aged municipal solid waste: A case study in taizhou. Sustainability 2021, 13, 4831. [Google Scholar] [CrossRef]
- Gomez, A.M.; Yannarell, A.C.; Sims, G.K.; Cadavid-Restrepo, G.; Herrera, C.X.M. Characterization of bacterial diversity at different depths in the Moravia Hill landfill site at Medellín, Colombia. Soil Biol. Biochem. 2011, 43, 1275–1284. [Google Scholar] [CrossRef]
- Uchida, M.; Hatayoshi, H.; Syuku-Nobe, A.; Shimoyama, T.; Nakayama, T.; Okuwaki, A.; Nishino, T.; Hemmi, H. Polymerase chain reaction-denaturing gradient gel electrophoresis analysis of microbial community structure in landfill leachate. J. Hazard. Mater. 2009, 164, 1503–1508. [Google Scholar] [CrossRef]
- Sawamura, H.; Yamada, M.; Endo, K.; Soda, S.; Ishigaki, T.; Ike, M. Characterization of microorganisms at different landfill depths using carbon-utilization patterns and 16S rRNA gene based T-RFLP. J. Biosci. Bioeng. 2010, 109, 130–137. [Google Scholar] [CrossRef]
- Song, J.; Zhang, W.; Gao, J.; Hu, X.; Zhang, C.; He, Q.; Yang, F.; Wang, H.; Wang, X.; Zhan, X. A pilot-scale study on the treatment of landfill leachate by a composite biological system under low dissolved oxygen conditions: Performance and microbial community. Bioresour. Technol. 2020, 296, 122344. [Google Scholar] [CrossRef]
- Song, L.; Yang, S.; Liu, H.; Xu, J. Geographic and environmental sources of variation in bacterial community composition in a large-scale municipal landfill site in China. Appl. Microbiol. Biotechnol. 2016, 101, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Wang, Y.; Zhao, H.; Long, D.T. Composition of bacterial and archaeal communities during landfill refuse decomposition processes. Microbiol. Res. 2015, 181, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Wang, Y.; Tang, W.; Lei, Y. Bacterial community diversity in municipal waste landfill sites. Appl. Microbiol. Biotechnol. 2015, 99, 7745–7756. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Cao, A.; Zhao, G.; Zhou, C.; Xu, R. Microbial community structure and diversity in a municipal solid waste landfill. Waste Manag. 2017, 66, 79–87. [Google Scholar] [CrossRef]
- Xu, S.; Lu, W.; Liu, Y.; Ming, Z.; Liu, Y.; Meng, R.; Wang, H. Structure and diversity of bacterial communities in two large sanitary landfills in China as revealed by high-throughput sequencing (MiSeq). Waste Manag. 2017, 63, 41–48. [Google Scholar] [CrossRef]
- Chien, P.C.T.; Yoo, H.S.; Dykes, G.A.; Lee, S.M. Isolation and characterization of cellulose degrading ability in Paenibacillus isolates from landfill leachate. Malays. J. Microbiol. 2015, 11, 185–194. [Google Scholar]
- Imron, M.F.; Kurniawan, S.B.; Soegianto, A. Characterization of mercury-reducing potential bacteria isolated from Keputih non-active sanitary landfill leachate, Surabaya, Indonesia under different saline conditions. J. Environ. Manag. 2019, 241, 113–122. [Google Scholar] [CrossRef]
- McDonald, J.E.; Lockhart, R.J.; Cox, M.J.; Allison, H.E.; McCarthy, A.J. Detection of novel Fibrobacter populations in landfill sites and determination of their relative abundance via quantitative PCR. Environ. Microbiol. 2008, 10, 1310–1319. [Google Scholar] [CrossRef]
- Pérez-Leblic, M.I.; Turmero, A.; Hernández, M.; Hernández, A.J.; Pastor, J.; Ball, A.S.; Rodríguez, J.; Arias, M.E. Influence of xenobiotic contaminants on landfill soil microbial activity and diversity. J. Environ. Manag. 2012, 95, S285–S290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bareither, C.A.; Wolfe, G.L.; McMahon, K.D.; Benson, C.H. Microbial diversity and dynamics during methane production from municipal solid waste. Waste Manag. 2013, 33, 1982–1992. [Google Scholar] [CrossRef] [PubMed]
- Köchling, T.; Sanz, J.L.; Gavazza, S.; Florencio, L. Analysis of microbial community structure and composition in leachates from a young landfill by 454 pyrosequencing. Appl. Microbiol. Biotechnol. 2015, 99, 5657–5668. [Google Scholar] [CrossRef]
- Mwaikono, K.S.; Maina, S.; Sebastian, A.; Schilling, M.; Kapur, V.; Gwakisa, P. High-throughput sequencing of 16S rRNA Gene Reveals Substantial Bacterial Diversity on the Municipal Dumpsite. BMC Microbiol. 2016, 16, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamps, B.W.; Lyles, C.N.; Suflita, J.M.; Masoner, J.R.; Cozzarelli, I.M.; Kolpin, D.W.; Stevenson, B.S. Municipal Solid Waste Landfills Harbor Distinct Microbiomes. Front. Microbiol. 2016, 7, 534. [Google Scholar] [CrossRef] [Green Version]
- Zainun, M.Y.; Simarani, K. Metagenomics profiling for assessing microbial diversity in both active and closed landfills. Sci. Total Environ. 2018, 616–617, 269–278. [Google Scholar] [CrossRef]
- Yadav, B.; Johri, A.K.; Dua, M. Metagenomic Analysis of the Microbial Diversity in Solid Waste from Okhla Landfill, New Delhi, India. Microbiol. Resour. Announc. 2020, 9, e00921-20. [Google Scholar] [CrossRef]
- Zhao, R.; Liu, J.; Feng, J.; Li, X.; Li, B. Microbial community composition and metabolic functions in landfill leachate from different landfills of China. Sci. Total Environ. 2020, 767, 144861. [Google Scholar] [CrossRef]
- Chen, K.; Deng, X.; Wang, L.; He, R.; Yang, Y.; Jiang, J.; Zhang, C.; Yang, S.; Fan, L.; Wang, L.; et al. Effects of different functional strains on key metabolic pathways of methanogenesis in the domestic waste fermentation reactor. Biomass Bioenergy 2021, 146, 105995. [Google Scholar] [CrossRef]
- Ma, P.C.; Ke, H.; Lan, J.W.; Chen, Y.M.; He, H.J. Field measurement of pore pressures and liquid-gas distribution using drilling and ERT in a high food waste content MSW landfill in Guangzhou, China. Eng. Geol. 2019, 250, 21–33. [Google Scholar] [CrossRef]
- Xu, H.; Zhan, L.-T.; Li, H.; Lan, J.-W.; Chen, Y.-M.; Zhou, H.-Y. Time-and stress-dependent model for predicting moisture retention capacity of high-food-waste-content municipal solid waste: Based on experimental evidence. J. Zhejiang Univ. A 2016, 17, 525–540. [Google Scholar] [CrossRef] [Green Version]
- MOHURD (Ministry of Housing and Urban-Rural Development of the People’s Republic of China). Sampling and Analysis Methods for Domestic Waste; CJ/T 313-2009; Standards Press of China: Beijing, China, 2009. (In Chinese)
- MOHURD (Ministry of Housing and Urban-Rural Development of the People’s Republic of China). General Detecting Methods for the Chemical Characteristic of Domestic Refuse; CJ/T 96-2013; Standards Press of China: Beijing, China, 2013. (In Chinese)
- MEE (Ministry of Ecology and Environment of the People’s Republic of China). Water Quality-Determination of Ammonia Nitrogen Nessler’s Spectrophotometry; HJ 535-2009; Standards Press of China: Beijing, China, 2009. (In Chinese) [Google Scholar]
- Ye, J.; Song, Z.; Wang, L.; Zhu, J. Metagenomic analysis of microbiota structure evolution in phytoremediation of a swine lagoon wastewater. Bioresour. Technol. 2016, 219, 439–444. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Caporaso, J.G. Author Correction: Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Paradis, E.; Schliep, K. ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Jari Oksanen, F.; Blanchet, G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. 2019. Available online: https://CRAN.R-project.org/package=vegan (accessed on 24 February 2022).
- Song, L.; Wang, Y.; Tang, W.; Lei, Y. Archaeal community diversity in municipal waste landfill sites. Appl. Microbiol. Biotechnol. 2015, 99, 6125–6137. [Google Scholar] [CrossRef]
- Hill, M.O.; Gauch, H.G. Detrended correspondence analysis: An improved ordination technique. Vegetatio 1980, 42, 47–58. [Google Scholar] [CrossRef]
- Wemheuer, F.; Taylor, J.A.; Daniel, R.; Johnston, E.; Meinicke, P.; Thomas, T.; Wemheuer, B. Tax4Fun2: Prediction of habitat-specific functional profiles and functional redundancy based on 16S rRNA gene sequences. Environ. Microbiome 2020, 15, 11. [Google Scholar] [CrossRef]
- Zhan, L.T.; Xu, H.; Chen, Y.M.; Lü, F.; Lan, J.W.; Shao, L.M.; Lin, W.A.; He, P.J. Biochemical, hydrological and mechanical behaviors of high food waste content MSW landfill: Preliminary findings from a large-scale experiment. Waste Manag. 2017, 63, 27–40. [Google Scholar] [CrossRef]
- Zhao, Y.; Song, L.; Huang, R.; Song, L.; Li, X. Recycling of aged refuse from a closed landfill. Waste Manag. Res. J. Sustain. Circ. Econ. 2007, 25, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Nakasaki, K.; Tran, L.T.H.; Idemoto, Y.; Abe, M.; Rollon, A. Comparison of organic matter degradation and microbial community during thermophilic composting of two different types of anaerobic sludge. Bioresour. Technol. 2009, 100, 676–682. [Google Scholar] [CrossRef]
- Reinhart, D.; Joslyn, R.; Emrich, C.T. Characterization of Florida, U.S. landfills with elevated temperatures. Waste Manag. 2020, 118, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Nesbø, C.L.; Bradnan, D.M.; Adebusuyi, A.; Dlutek, M.; Petrus, A.K.; Foght, J.; Doolittle, W.F.; Noll, K.M. Mesotoga prima gen. nov., sp. nov., the first described mesophilic species of the Thermotogales. Extremophiles 2012, 16, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Ben Hania, W.; Ghodbane, R.; Postec, A.; Hamdi, M.; Ollivier, B.; Fardeau, M.-L. Defluviitoga tunisiensis gen. nov., sp. nov., a thermophilic bacterium isolated from a mesothermic and anaerobic whey digester. Int. J. Syst. Evol. Microbiol. 2012, 62, 1377–1382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Li, F.; Kuroki, A.; Loh, K.C.; Wang, C.H.; Dai, Y.; Tong, Y.W. Methane yield enhancement of mesophilic and thermophilic anaerobic co-digestion of algal biomass and food waste using algal biochar: Semi-continuous operation and microbial community analysis. Bioresour. Technol. 2020, 302, 122892. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Wang, C.; Sun, F.; Zhu, W.; Wu, W. A comparison of microbial characteristics between the thermophilic and mesophilic anaerobic digesters exposed to elevated food waste loadings. Bioresour. Technol. 2014, 152, 420–428. [Google Scholar] [CrossRef]
- Kim, E.; Lee, J.; Han, G.; Hwang, S. Comprehensive analysis of microbial communities in full-scale mesophilic and thermophilic anaerobic digesters treating food waste-recycling wastewater. Bioresour. Technol. 2018, 259, 442–450. [Google Scholar] [CrossRef]
- Ghollarata, M.; Raiesi, F. The adverse effects of soil salinization on the growth of Trifolium alexandrinum L. and associated microbial and biochemical properties in a soil from Iran. Soil Biol. Biochem. 2007, 39, 1699–1702. [Google Scholar] [CrossRef]
- Sardinha, M.; Müller, T.; Schmeisky, H.; Joergensen, R.G. Microbial performance in soils along a salinity gradient under acidic conditions. Appl. Soil Ecol. 2003, 23, 237–244. [Google Scholar] [CrossRef]
- Townsend, T.G.; Miller, W.L.; Lee, H.-J.; Earle, J.F.K. Acceleration of Landfill Stabilization Using Leachate Recycle. J. Environ. Eng. 1996, 122, 263–268. [Google Scholar] [CrossRef]
- Wu, X.; Tian, Z.; Lv, Z.; Chen, Z.; Liu, Y.; Yong, X.; Zhou, J.; Xie, X.; Jia, H.; Wei, P. Effects of copper salts on performance, antibiotic resistance genes, and microbial community during thermophilic anaerobic digestion of swine manure. Bioresour. Technol. 2020, 300, 122728. [Google Scholar] [CrossRef]
- Jetten, M.S.M. The microbial nitrogen cycle. Environ. Microbiol. 2008, 10, 2903–2909. [Google Scholar] [CrossRef]
- Fierer, N.; Lauber, C.L.; Ramirez, K.S.; Zaneveld, J.; Bradford, M.A.; Knight, R. Comparative metagenomic, phylogenetic and physiological analyses of soil microbial communities across nitrogen gradients. ISME J. 2012, 6, 1007–1017. [Google Scholar] [CrossRef] [Green Version]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westlake, K.; Archer, D.; Boone, D. Diversity of cellulolytic bacteria in landfill. J. Appl. Bacteriol. 1995, 79, 73–78. [Google Scholar] [CrossRef]
- Inaba, T.; Su, T.; Aoyagi, T.; Aizawa, H.; Sato, Y.; Suh, C.; Lee, J.H.; Hori, T.; Ogata, A.; Habe, H. Microbial community in an anaerobic membrane bioreactor and its performance in treating organic solid waste under controlled and deteriorated conditions. J. Environ. Manag. 2020, 269, 110786. [Google Scholar] [CrossRef] [PubMed]
- Conklin, A.; Stensel, H.D.; Ferguson, J. Growth Kinetics and Competition between Methanosarcina and Methanosaeta in Mesophilic Anaerobic Digestion. Water Environ. Res. 2006, 78, 486–496. [Google Scholar] [CrossRef] [PubMed]
- Youcai, Z.; Zhugen, C.; Qingwen, S.; Renhua, H. Monitoring and long-term prediction of refuse compositions and settlement in large-scale landfill. Waste Manag. Res. 2001, 19, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Ximenes, F.; Gardner, W.; Cowie, A. The decomposition of wood products in landfills in Sydney, Australia. Waste Manag. 2008, 28, 2344–2354. [Google Scholar] [CrossRef] [PubMed]
- Bothe, H.; Ferguson, S.; Newton, W.E. Biology of the Nitrogen Cycle; Elsevier: Amsterdam, The Netherlands, 2007; pp. 3–20. [Google Scholar]
Figure 1.
The location of both landfills and their sampling points. Label A and B refer to the sampling points in the Xingfeng landfill; Label C and D refer to the sampling points in the Tianziling landfill.
Figure 1.
The location of both landfills and their sampling points. Label A and B refer to the sampling points in the Xingfeng landfill; Label C and D refer to the sampling points in the Tianziling landfill.

Figure 2.
Relative abundance of microorganisms at different levels: (a) relative abundance of different phyla in the Tianziling and Xingfeng landfills, (b) relative abundance of different classes among Firmicutes, and (c) relative abundance of different classes among other phyla.
Figure 2.
Relative abundance of microorganisms at different levels: (a) relative abundance of different phyla in the Tianziling and Xingfeng landfills, (b) relative abundance of different classes among Firmicutes, and (c) relative abundance of different classes among other phyla.

Figure 3.
Heat map of the genera with a relative abundance > 1.0% in at least one sample. A total of 55 genera were selected from all of the samples. The color intensity (logarithmic scale) in each cell shows the percentage of a genus in a sample, referring to the color key on the right.
Figure 3.
Heat map of the genera with a relative abundance > 1.0% in at least one sample. A total of 55 genera were selected from all of the samples. The color intensity (logarithmic scale) in each cell shows the percentage of a genus in a sample, referring to the color key on the right.

Figure 4.
Different factors influencing the bacterial community in landfilled waste. Group A includes samples A1, A2, A3, and A4; Group B includes samples B1, B2, B3, and B4; Group C includes samples C1, C2, C3, and C4; Group D includes samples D1, D2, D3, and D4. (a) PCoA based on “Bray–Curtis” distance showing the relationship among samples (Adonis: R2 = 0.422, p = 0.001). Each circle dot represents a different sample, (b) the Venn diagram showing the number of shared and unique ASVs between different sites, and (c) RDA of the bacterial community and the physicochemical attributes selected by VIFs. The arrows indicate the direction and magnitude of the measurable variables associated with bacterial community structures. Green dots represent different genera, and dots of other colors represent different samples.
Figure 4.
Different factors influencing the bacterial community in landfilled waste. Group A includes samples A1, A2, A3, and A4; Group B includes samples B1, B2, B3, and B4; Group C includes samples C1, C2, C3, and C4; Group D includes samples D1, D2, D3, and D4. (a) PCoA based on “Bray–Curtis” distance showing the relationship among samples (Adonis: R2 = 0.422, p = 0.001). Each circle dot represents a different sample, (b) the Venn diagram showing the number of shared and unique ASVs between different sites, and (c) RDA of the bacterial community and the physicochemical attributes selected by VIFs. The arrows indicate the direction and magnitude of the measurable variables associated with bacterial community structures. Green dots represent different genera, and dots of other colors represent different samples.


Figure 5.
Analysis of macromolecular hydrolysis pathways based on Tax4fun2: (a) three paths for macromolecular hydrolysis. (b) Relative abundance of macromolecular metabolic functional enzymes in different sites (red box: carbohydrate, blue box: protein, and green box: lipids). (c) Relative abundance of macromolecular metabolic functional enzymes in different depths (red box: carbohydrate, blue box: protein, and green box: lipids).
Figure 5.
Analysis of macromolecular hydrolysis pathways based on Tax4fun2: (a) three paths for macromolecular hydrolysis. (b) Relative abundance of macromolecular metabolic functional enzymes in different sites (red box: carbohydrate, blue box: protein, and green box: lipids). (c) Relative abundance of macromolecular metabolic functional enzymes in different depths (red box: carbohydrate, blue box: protein, and green box: lipids).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Characterization of microbial communities in landfill leachate and waste.
| Region | Phase | Bacteria Phylum | Dominant Genus | Reference |
|---|---|---|---|---|
| UK | Leachate | Fibrobacters | NA | [23] |
| Western Japan | Solid waste | Firmicutes, Bacteroidota, Proteobacteria | NA | [14] |
| Medellín, Colombia | Solid waste | Bacteroidota, Proteobacteria | Acinetobacter | [12] |
| Madrid, Spain | Solid waste | Proteobacteria, Acidobacteria, Actinobacteria | Sphingosinicella, Microvirga, Amaricoccus, Conexibacter, Marmoricola, Pseudonocardina | [24] |
| USA | Solid waste/leachate | Proteobacteria, Firmicutes, Bacteroidota | Pseudomanas, Arcobacter, Acinetobacter | [25] |
| Brazil | Solid waste | Firmicutes, Bacteroidota, Proteobacteria | Syntrophomonas, Pseudomanas, Clostridium, Petrimonas, Proteiniphilum | [26] |
| Arusha, Tanzania | Solid waste | Firmicutes, Bacteroidota, Proteobacteria, Acidobacteria, Actinobacteria, Spirochaetes | Acinetobacter, Clostridium, Proteinclasticum, Lactobacillus | [27] |
| USA | Leachate | Firmicutes, Proteobacteria | NA | [28] |
| Beijing and Xi’an, China | Solid waste | Firmicutes, Bacteroidota, Proteobacteria | Proteiniphilum, Atopostipes, Petrimonas | [20] |
| Yangzhou, China | Solid waste | Firmicutes, Bacteroidota, Proteobacteria | Halanaerobium, Halanaerobium, Halocella | [19] |
| Malaysia | Solid waste | Firmicutes, Bacteroidota, Proteobacteria, Acidobacteria, Actinobacteria, Spirochaetes | NA | [29] |
| Xi’an, China | Solid waste | Firmicutes, Bacteroidota, Proteobacteria | Olivibacter, Petrimonas | [16] |
| New Delhi, India | Solid waste | Actinobacteria, Proteobacteria, Firmicutes, Bacteroidota, Planctomycetes, Cyanobacteria | NA | [30] |
| 11 cities, China | Leachate | Bacteroidota, Firmicutes, Proteobacteria | Candidatus Cloacamonas, Defluviitoga, Sphaerochaeta, Arcobacter, Syntrophomonas, and Petrimonas | [31] |
Table 2.
Composition of MSW from the downtown area of Guangzhou and Hangzhou (wet basis).
| Component | Landfill | Food Waste | Plastic | Paper | Textile | Wood | Glass and Metal | Cinder and Dust | Others |
|---|---|---|---|---|---|---|---|---|---|
| Mass proportion (100%) | Xingfeng | 54.0 | 19.9 | 12.3 | 4.6 | 3.0 | 3.3 | 1.6 | 1.3 |
| Tianziling | 59.4 | 18.0 | 13.5 | 2.0 | 1.1 | 2.3 | 3.7 | 0.0 |
Table 3.
Physicochemical properties of MSW samples in the Xingfeng and Tianziling landfills.
| Sample ID | Site | Depth (m) | Age (a) | Moisture Content (%) | Organic Matter Content (%) | pH | EC (ms/cm) | Ammonia (mg/kg) |
|---|---|---|---|---|---|---|---|---|
| A1 | Xingfeng landfill, Guangzhou | 10 | 2~3 | 49% | 26.73% | 7.7 | 35.1 | 108.15 |
| A2 | 20 | 10~11 | 25% | 23.68% | 6.6 | 32.7 | 165.45 | |
| A3 | 30 | 10~11 | 33% | 22.55% | 8.0 | 28.5 | 180.75 | |
| A4 | 40 | 10~11 | 41% | 19.93% | 8.0 | 27.8 | 166.35 | |
| B1 | 10 | 2~3 | 51% | 28.01% | 7.6 | 33.5 | 49.05 | |
| B2 | 20 | 10~11 | 27% | 32.82% | 6.8 | 33.4 | 150.45 | |
| B3 | 30 | 10~11 | 37% | 28.55% | 8.0 | 28.0 | 166.65 | |
| B4 | 40 | 10~11 | 42% | 26.59% | 8.0 | 29.1 | 183.75 | |
| C1 | Tianziling landfill, Hangzhou | 10 | 7~9 | 33% | 19.88% | 7.8 | 24.5 | 123.75 |
| C2 | 20 | 9~11 | 32% | 22.93% | 7.7 | 18.6 | 187.05 | |
| C3 | 30 | 11~13 | 24% | 21.61% | 7.9 | 24.1 | 30.03 | |
| C4 | 40 | 13~15 | 21% | 15.56% | 7.7 | 24.9 | 299.25 | |
| D1 | 10 | 1~2 | 52% | 32.39% | 7.5 | 30.1 | 169.05 | |
| D2 | 20 | 5~7 | 32% | 32.53% | 7.4 | 24.3 | 226.95 | |
| D3 | 30 | 7~9 | 30% | 35.04% | 7.0 | 28.3 | 28.05 | |
| D4 | 40 | 9~11 | 25% | 29.95% | 7.8 | 19.1 | 116.70 |
Each sample in triplicate was taken from the corresponding depth of the target sampling point.
Table 4.
The α-diversity of different samples.
| Sample ID | Chao-1 | Shannon | Evenness | Simpson |
|---|---|---|---|---|
| A1 | 451.7 | 4.22 | 0.195 | 0.955 |
| A2 | 497.8 | 4.19 | 0.160 | 0.956 |
| A3 | 603.5 | 3.98 | 0.103 | 0.944 |
| A4 | 362 | 4.37 | 0.264 | 0.975 |
| B1 | 334.9 | 3.72 | 0.126 | 0.930 |
| B2 | 345.9 | 3.47 | 0.094 | 0.905 |
| B3 | 305.8 | 3.91 | 0.164 | 0.959 |
| B4 | 380.2 | 4.39 | 0.247 | 0.973 |
| C1 | 370.4 | 3.54 | 0.094 | 0.921 |
| C2 | 277 | 2.51 | 0.045 | 0.690 |
| C3 | 479.9 | 4.48 | 0.184 | 0.971 |
| C4 | 324.2 | 3.60 | 0.116 | 0.937 |
| D1 | 509.5 | 4.87 | 0.281 | 0.983 |
| D2 | 402 | 4.01 | 0.140 | 0.955 |
| D3 | 349.6 | 3.60 | 0.105 | 0.924 |
| D4 | 312.7 | 3.16 | 0.079 | 0.868 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ke, H.; Li, J.; Zhang, X.; Hu, J. Bacterial Community Structure and Predicted Metabolic Function of Landfilled Municipal Solid Waste in China. Sustainability 2022, 14, 3144. https://0-doi-org.brum.beds.ac.uk/10.3390/su14063144
AMA Style
Ke H, Li J, Zhang X, Hu J. Bacterial Community Structure and Predicted Metabolic Function of Landfilled Municipal Solid Waste in China. Sustainability. 2022; 14(6):3144. https://0-doi-org.brum.beds.ac.uk/10.3390/su14063144
Chicago/Turabian StyleKe, Han, Jinghang Li, Xuanqi Zhang, and Jie Hu. 2022. "Bacterial Community Structure and Predicted Metabolic Function of Landfilled Municipal Solid Waste in China" Sustainability 14, no. 6: 3144. https://0-doi-org.brum.beds.ac.uk/10.3390/su14063144
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.