
Aquatic Plants and Aquatic Animals in the Context of Sustainability: Cultivation Techniques, Integration, and Blue Revolution

 ,
,  ,
, 
Abstract
:1. Introduction
2. Cultivation Techniques

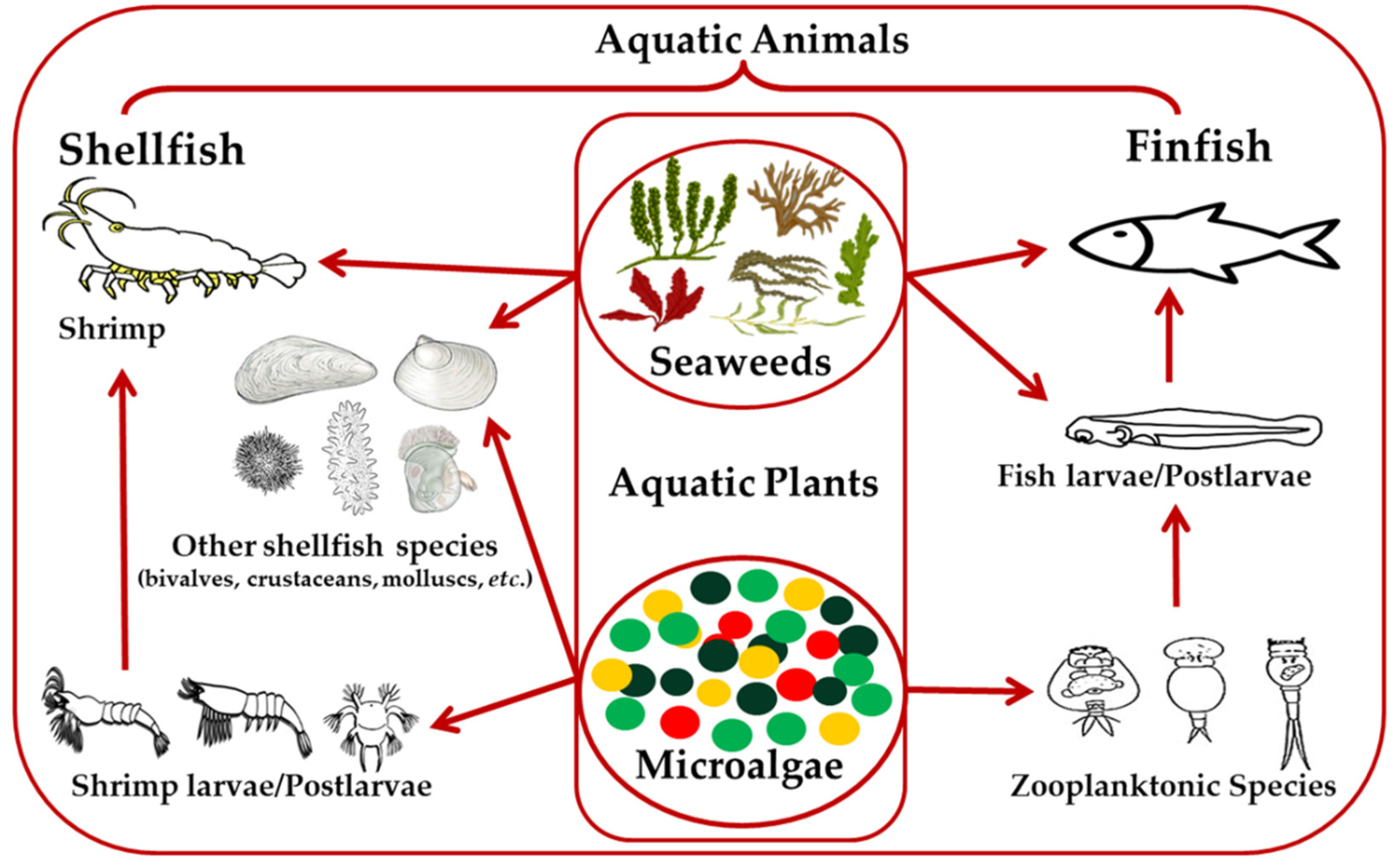{kind=link}
| Item | Biochemical Composition (%, Dry Weight Bases) | References | ||
|---|---|---|---|---|
| Protein | Carbohydrate | Lipid | ||
| The most Commercially Aquafeed Ingredients | ||||
| Fish meal | 63.00 | 11.00 | 12.5 | [54] |
| Soybean | 44.00 | 2.20 | 39.00 | [55] |
| Corn-gluten meal | 62.00 | 5.00 | 18.50 | [56] |
| Wheat meal | 12.20 | 2.90 | 69.00 | [57] |
| Yeast (Saccharomyces cerevisiae) | 50.10 | 1.80 | 4.6 | [58] |
| Hydrolyzed feather meal | 84.00 | 10.40 | - | [59] |
| Amphipod meal (Gammarus pulex) | 40.00 | 27.40 | 5.5.0 | [60] |
| Microalgae Species | ||||
| Arthrospira platensis | 50–65 | 8–14 | 4–9 | [61,62] |
| Nannochloropsis | 18–34 | 27–36 | 24–28 | [61,63,64] |
| Chlorella vulgaris | 51–58 | 2–17 | 14–22 | [65] |
| Tetraselmis chuii | 25 | 25 | 12 | [63,66] |
| T. suecica | 38.73 | 44.29 | 12.38 | [21] |
| Isochrysis galbana | 27 | 34 | 11 | [67] |
| Botryococcus braunii | 39–40 | 19–31 | 25–34 | [63] |
| Pavlova sp. | 24–29 | 6–9 | 9–14 | [65,66] |
| Dunaliella salina | 11–34 | 14–32 | 6–14 | [65,67,68] |
| Scenedesmus obliquus | 48–56 | 10–17 | 12–14 | [66,69] |
| Phaeodactylum tricornutum | 34–38 | 11–17 | 13–20 | [61,63] |
| Chlamydomonas reinhardtii | 48 | 17 | 21 | [66] |
| Haematococcus pluvialis | 17–45 | 20–37 | 15–40 | [70] |
| Skeletonema costatum | 25 | 4–6 | 10 | [66] |
| Chaetoceros muelleri | 59.00 | 10.00 | 31.00 | [71] |
| C. calcitrans | 40.00 | 37.00 | 3.00 | [71] |
| Chaetoceros sp. | 12–59 | 4–37 | 7–31 | [66] |
| Thalassiosira pseudonana | 34.00 | 9.00 | 19.00 | [66] |
| T. weissflogii | 13.2 | 10.00 | 20.00 | [72] |
| Porphyridium cruentum | 28–39 | 40–57 | 9–14 | [65] |
| Macroalgae Species | ||||
| Eucheuma sp. | 9.8–5.5 | 26.5–63 | 1–5 | [73,74] |
| Kappaphycus sp. | 3.4–6.2 | 65.2–55.7 | 1.1–1.5 | [75,76] |
| Sargassum sp. | 7.5–5.5 | 28–30 | 1.5–0.4 | [74,77,78] |
| Gracilaria sp. | 11–21 | 19–27 | 0.5–2.8 | [74,79] |
| Pterocladia capillacea | 17–20 | 47–51 | 1.7–2.5 | [80] |
| Jania rubens | 9–23 | 34–50 | 1–2.5 | [80] |
| Ulva lactuca | 12–20 | 42–46 | 2–4 | [80] |
| Dictyota dichotoma | 7–7.5 | 24–26 | 7–7.5 | [27] |
| Turbinaria decurrens | 32–33 | 2.5 | 5 | [27] |
| Laurencia obtusa | 21 | 4 | 3 | [81] |
| Porphyra sp. | 25–42 | 36 | 0.5 | [82,83] |
2.1. Microalgae as Livefeeds in Aquaculture Hatcheries
2.2. Microalgae Biomass
2.3. Algae as Aqua-Feed Additives and/or Replacement of Diet Ingredient
2.4. Algae as a Water Conditioner
2.5. Seaweed Co-Culture and Integration with Aquatic Animals
| Integration/Applications Type | Aquatic Animal Species | Aquatic Plant Species | Results/Interactions | References |
|---|---|---|---|---|
| Live Feeds | Zooplankton | |||
| Rotifers (B. plicatilis and B. rotundiformis), artemia (A. franciscana and Artemia sp.), and copepods (Bestiolina sp., T. stylifera, O. rigida, N. minor, Acartia spp., Paracalanus pas). | N. salina, N. oculata, N. gaditana, Nannochloropsis sp., Nannochloris sp., C. salina, Chlorella sp., Dunaleilla salina, C. calcitrans, Chaetoceros sp., I. galbana; T. chuii, T. suecica, P. lutheri, Coscinodiscus sp., S. costatum, Pseudo-nitzschia sp., Prorocentrum sp., Rhodomonas sp., and Navicula sp. | Improved egg production, hatchability, filtration and ingestion rates, productivity, population, sex ratio, growth performance, survival rates, reproductive behaviour. In addition, enhance the biochemical composition; PUFAs; HUFAs; EPA; and DHA contents. | [94,95,96,97,98,99,100,102,103] | |
| Shrimp | ||||
| Larvae of Pacific white leg shrimp, L. vannamei, Giant Tiger shrimp, P. monodon, and Kuruma shrimp, Marsupenaeus japonicus. | Chaetoceros, Skeletonema, Thalassiosira, Detonula, Phaeodactylum, Nitzschia, Navicula Isochrysis, Tetraselmis, Nannochloropsis, Chlorella, Dunaliella, and Tisochrysis | The larvae that fed on different diatom species achieved a higher survival rate, growth, body composition, development, digestive capability, and digestive enzyme activities, than the other microalgae genera. | [106,109,110,112,154,167,208,209] | |
| Larvae of Giant tiger shrimp, P. monodon | I. galbana and C. muelleri cultured with F/2 media supplemented with seaweed liquid extract (SWE) of U. lactuca, E. intestinalis, C. sinuosa, S. ilicifolium, G. corticata, and H. valentiae. | Microalgae cultured with F/2 media supplemented with SWE can be widely utilized as an alternative low-cost media in the production of live foods for the production of shrimp larvae P. monodon. | [108] | |
| Bivalve | ||||
| Larvae and juvenile of Pacific oyster, C. gigas, juvenile of scallop, Pecten fumatus, larvae and juvenile of Sydney rock oyster, S. glomerata, larvae of Manila clam, T. philippinarum, larvae of grooved carpet shell Ruditapes decussatus, juveniles of Pacific geoduck clam, Panopea generosa, larvae of Giant clam, Tridacna noae, larvae and juvenile of some important commercial bivalves of Ostrea edulis, Mercenaria mercenaria, T. semidecussata, and Mytilus galloprovincialis | Tetraselmis, Chaetoceros, Skeletonema, Isochrysis, Pavlova, Schizochytrium, Cyclotella, Hematococcus, Phaeodactylum, Tisochrysis, A. platensis, and Thalassiosira | Bivalves larvae and juveniles fed on the live form of microalgae showed increased survival rates, enhanced growth, nutritional value, body composition filtration, ingestion rates, and improved digestibility rates. | [113,114,115,116,117,118,119,120,121,122,123,124] | |
| Sea Cucumber | ||||
| Larvae of sandfish H. scabra | C. gracilis, I. galbana, and T. chuii, Rhodomonas sp., and T. tetrathele (single or mixed diets). | Increased the survival rate, enhanced metamorphosis, settlement, and improved the nutritional value of larvae. Improved the larval survival, development, and growth. | [128,131] | |
| Larvae of California sea cucumber, P. californicus | C. calcitrans, C. muelleri, D. tertiolecta, Isochrysis sp., P. lutheri, P. tricornutum, T. suecica, and T. pseudonana (single or mixed diets). | Among all the examined species, the diatom species C. calcitrans was the best species that achieved significant larval survival, growth, and metamorphosis. | [130] | |
| Juvenile of Selenka, A. japonicus | A. platensis, D. inornata, Cylindrotheca fusiformis, and N. Closterium. | The juvenile growth, energy contents, and nutritional value were improved. | [129] | |
| Adults red sea cucumber, P. tremulus | D. tertiolecta, I. galbana, and T. chuii (a mixture of them). | Low food availability (10–20 × 103 cell mL─1) resulted in high mortality. | [132] | |
| Sea Urchin | ||||
| Larvae of white-spined sea urchin, T. gratilla | Isochrysis sp., C. muelleri, S. pseudocostatum, P. lutheri, and T. suecica, | Isochrysis sp., C. muelleri, S. pseudocostatum, and are the best significant species for the rearing of larvae. | [135] | |
| Larvae of P. lividus | D. tertiolecta, T. suecica, C. elongata, and P. carterae. | When larvae fed on C. elongata, P. carterae, and D. tertiolecta, the metamorphosis was successfully enhanced. Larvae fed on C. elongata had 300% and 20% higher survival and development rates than larvae fed on P. carterae and D. tertiolecta. | [136] | |
| Larvae of A. crassispina. | C. gracilis, P. viridis, C. vulgaris, Platymonas subcordiformis, D. zhanjiangensis, and mixture of C. gracilis, and D. zhanjiangensis. | There were significant differences in the larvae’s survival and growth. The highest level was recorded by the larvae fed on D. zhanjiangensis followed by C. gracilis, P. viridis, C. vulgari, and, finally, the mixture of D. zhanjiangensis and C. gracilis. | [137] | |
| Seahorses | ||||
| Juveniles of longsnout seahorse, H. reidi. | N. oculate, and I. galbana. | The survival, ingestion rate, and growth of the juveniles were significantly higher in the treatment that fed on N. oculata or I. | [78,134,138,139,140] | |
| Biomass | Zooplankton | |||
| Rotifers, B. plicatilis and B. rotundiformis, used in larvae rearing of Gilthead seabream, S. aurata. | N. oculata (live and freeze-dried forms). | Applications of freeze-dried N. oculata can be used successful with 100% survival and without affecting water quality. | [93,178,210] | |
| Rotifer, B. plicatilis. | I. galbana, C. muelleri, P. lutheri, and Nannochloropsis sp. (live and frozen-concentrated forms). | All examined species could apply to relative enriched PUFA, EPA, and DHA levels in the rotifer, even in live and/or frozen-concentrated forms. | [104] | |
| Rotifer, B. plicatilis. | N. oculata (freeze-dried), A. platensis (dried), comparing to baker’s yeast S. cerevisiae (dried). | Unlive species of freeze-dried N. oculata and dried A. platensis resulted in an adequate rotifer population and population growth rate, comparing to yeast. | [142] | |
| Rotifer, B. plicatilis. | A. platensis NIOF17/003 (defatted, biodiesel by-product). | There was a significant increase in rotifer females carrying eggs and population when fed free lipid A. platensis NIOF17/003 at a level of 0.6 g L─1. | [62] | |
| Rotifer, B. plicatilis. | A. platensis NIOF17/003 (dried form), loaded with aquaculture effluent based ammonia or toxic dye (Ismate violet 2R, IV2R). | Rotifers are highly sensitive to the dried biomass of A. platensis loaded with ammonia levels or toxic dyes. Overall, the dried A. platensis loaded with ammonia levels is a potential feed source for rotifers. | [18,19] | |
| Artemia, A. franciscana. | N. oceanica NIOF15/001 (Defatted biodiesel by-product). | Applying defatted biomass of N. oceanica NIOF15/001 at 0.1 g L−1 significantly enhanced growth (40%) and survival (500%) of A. franciscana. | [64] | |
| Copepod, Cyclopina kasignete. | Melosira sp. and N. oculata (dried form), compared to I. lutea and N. oculata (live form). | The copepod C. kasignete that fed Melosira sp. (live or dried form) achieved higher EPA, DHA, ARA contents, trypsin, and protease activities than other microalga species. | [151] | |
| Shrimp | ||||
| Larvae and postlarval of Pacific white leg shrimp, L. vannamei. | Chaetoceros sp. (freeze-dried). | The survival, growth, and beneficial bacterial count in the gut of marine shrimp, L. vannamei, was significantly increased by the addition of freeze-dried Chaetoceros sp. directly in the culture water of propped. | [155] | |
| Larvae of Giant tiger shrimp, P. Monodon. | Sun-dried Chaetoceros sp., Isochrysis sp., and Tetraselmis, compared the live form of Chaetoceros sp. | Larvae fed the sun-dried Chaetoceros sp. and Tetraselmis sp. had good survival and growth performance, compared to the sun-dried Isochrysis sp. and the live form of Chaetoceros sp. | [152] | |
| Larvae of Giant tiger shrimp, P. monodon. | C. muelleri and T. weissflogii in live form comparing to Arthrospira powder. | Larval metamorphosis, survival, beneficial bacterial count and stability, digestive capability, and digestive enzyme activities of larvae fed the microalgae life form were significantly higher than those of the Arthrospira powder. | [154] | |
| Larvae of brown shrimp, Farfantepenaeus aztecus. | Replacement of live Chaetoceros sp. in partial or total replacement using their paste form (Chaetoceros 1000®, Premium Fresh Instant Algae™ paste) and other inert feeds. | Larvae growth performances were significantly lower than the control “live Chaetoceros sp.” | [153] | |
| Larvae of Indian prawn, P. indicus. | Partial replacement of Tetraselmis and Skeletonema with a freeze dried microencapsulated diets. | The microcapsules (application contains the extracts of Tetraselmis and Skeletonema) improved the survival and growth of the P. indicus larval stages, besides reducing their reliance on microalgae life forms. | [156] | |
| Bivalves | ||||
| Larvae of Pacific oyster, C. gigas. | P. lutheri (biomass compared to live form). | No significant differences in growth or survival between the larvae reared on the fresh and the preserved biomass. | [86] | |
| Larvae of Pacific oyster, C. gigas. | P. lutheri (concentrated as replacing live C. calcitrans of 50–80%). | No significant differences in growth or survival compared to the control live form. | [113] | |
| Larvae of Pacific oyster, C. gigas. | T. suecica (concentrated compared to live form). | There were no significant differences in larval growth compared to the control live form. | [211] | |
| Larvae and juvenile of Pacific oyster, C. gigas. | C. calcitrans, S. costatum; I. galbana (concentrates as partial replacement or supplementation, compared to live form). | There were no significant differences in larvae and juvenile growth rates compared to the live forms. | [86] | |
| Juvenile of Pacific oyster, C. gigas | C. calcitrans, S. costatum (concentrated, compared to live form) | There were no significant differences in juvenile growth compared to the live forms. | [115] | |
| Juveniles Scallop, C. gigas, and Pecten fumatus. | C. muelleri (concentrated, compared to live form). | Improved the growth performances of juveniles. | [114] | |
| Larvae and juveniles of Sydney rock oyster, S. commercialis | P. lutheri, I. galbana, Tetraselmis sp., C. calcitrans, C. muellei (concentrated and storage for 6–8 weeks). | The survival rates and growth performance of bivalves (larval and juvenile) are equal to that achieved by the microalgal live forms. | [116] | |
| Larvae of Sydney rock oyster, S. commercialis | P. lutheri, I. galbana, C. calcitrans (mixed concentrated). | Similar and/or higher growth performance than larvae fed the control live forms. | [212] | |
| Larvae Manila clam, T. Philippinarum. | T. suecica; Nannochloris sp. (spray-dried) | Larval growth was similar to that of the same live species. | [118] | |
| Juvenile of five important commercial bivalve species (C. gigas, O. edulis, M. mercenaria, T. philippinarum, and T. decussata). | T. suecica (spray-dried), compared to live T. suecica; C. calcitrans. | The juvenile growth was similar to live Tetraselmis, while less than C. calcitrans. | [117] | |
| Juveniles C. gigas and T. Semidecussata. | Schizochytrium sp. (spray-dried as a 40% partial replacement of the live form). | Similar juvenile growth compared to the control live form. | [120] | |
| Juveniles of C. gigas, T. philippinarum, and O. edulis. | T. suecica; Cyclotella cryptica (mixed spray-dried, 70:30%). | There were no significant differences in juvenile growth compared to the live form. | [119] | |
| Juveniles of Mediterranean mussels M. galloprovincialis | Schizochytrium sp.; A. Platensis; Hematococcus pluvialis (spray-dried). | Growth performances of juvenile fed spray-dried algae were significantly higher than those of live feeds form. | [121] | |
| Larvae Ruditapes decussatus (Grooved carpet shell). | I. galbana; T. suecica; P. tricornutum (freeze-dried). | The growth of larvae fed freeze-dried algal species was significantly lower than that of live feeds. | [122] | |
| Juveniles of Pacific Geoduck Clam Panopea generosa. | C. muelleri; Tisochrysis lutea; Schizochytrium sp.; A. platensis (spray-dried as a different partial replacement of the live form). | Low significant growth rates of juveniles were obtained when replacing 25–50% of the live forms with spray-dried. | [123] | |
| Juveniles of black-lip pearl oyster, P. margaritifera. | T. suecica (dried in a mixture of 1:1 with the life form). | Growth of umbo larvae fed a 1:1 mixture of fresh algae and dried Tetraselmis was significantly greater than that of those fed live forms. | [213] | |
| Larvae of Giant clam, Tridacna noae. | Isochrysis sp., Pavlova sp., Tetraselmis sp., T. weissflogii. | All experimented microalgae were ingested and, thereafter, digested by larvae, confirming that the investigated algae concentrates were a good food source for the giant clam larvae. | [124] | |
| Seacucumber | ||||
| Larvae of sandfish, H. scabra. | Isochrysis sp., Pavlova sp., and T. weissflogii. | Significantly increased the survival rate, enhanced growth performance, and improved the nutritional value of larvae. Besides, it reduced larvae production cost. | [214] | |
| Larvae of sandfish, H. scabra | Isochrysis 1800® (Isochrysis sp.), and the product named Shellfish Diet 1800® (consisting of a mix of Isochrysis sp., T. pseudonana, Tetraselmis sp., Pavlova sp., T. weissflogii, and C. calcitrans (with a ratio: 30:30:19:13:6: and 3%, respectively, on a dry weight basis). | The examined microalgal concentrates confirm comparable results to those of the same live feeds traditionally utilized in the commercial production of H. scabra (sand-fish) in marine hatcheries. | [158] | |
| Larvae of sandfish, H. scabra. | Live microalgae Isochrysis sp. and C. muelleri compared to six concentrated algal diets (Instant Algae®, Reed Mariculture Inc.) which were Isochrysis sp. Pavlova sp., Tetraselmis sp., T. weissflogii, T. pseudonana, and a mixture of Isochrysis sp., Tetraselmis sp., T. pseudonana, and Pavlova sp. (Shellfish Diet 1800®). | All experimental algal species (even in live or concentrated form) were ingested by the juveniles. The live form of C. muelleri showed the highest juvenile growth. Overall, microalgae concentrates are suitable alternatives to live microalgae through the hatchery stages of sea cucumber, H. scabra. | [85,215] | |
| The orange-footed adult sea cucumber, Cucumaria frondose. | A commercial microalgae diet (Shellfish Diet 1800®: a mix of Isochrysis, Tetraselmis, T. pseudonana and Pavlova, with a ratio of 40:25:20:15% dry weight 8%), dried powder of seaweed (S. latissimi and A. nodosum) compared to the control (no diet supplementation). | Adult sea cucumbers fed with powdered macroalgae have similar physiological characteristics to those fed with commercial powdered microalgae diets, confirming that powdered macroalgae diets are a promising feed source for intensive aquaculture of adult individuals of C. frondosa. | [159] | |
| Sea-Urchin | ||||
| Larvae of P. lividus. | D. tertiolecta and a concentrated algal paste (Shellfish Diet 1800®). | There is no significant difference in survival and metamorphosis rates for larvae fed on live or paste microalgae. | [133] | |
| Seahorses | ||||
| Juvenile of newborn seahorse H. reidi. | N. oculata (live form, commercial paste, and flocculated paste). | No significant differences were recorded in the survival rates, dry weight, or ingestion rates of the juvenile seahorse. | [216] | |
| Aqua-Feed Additives and/or Replacement of Diet Ingredients | Fish | |||
| Nile tilapia, O. niloticus. | Defatted N. oculata, Schizochytrium sp., A. platensis, Chlorella sp., Anabaena sp., Dunaliella sp., G. arcuata, and seaweed liquid extract (TrueAlgaeMax, TAM®, prepared from U. lactuca, J. rubens, and P. capillacea). | Significantly increase productive performance nutrient utilization, survival rate, immune status, antioxidant enzymes, gene expression, histological status, and disease resistance. | [24,217,218,219,220,221,222] | |
| Juveniles of European sea bass, Dicentrarchus labrax. | I. lutea, T. suecica, P. viridis Nannochloropsis sp., N. oceanica, defatted Nannochloropsis sp., G. gracilis. | Significantly improve the muscle tissue composition, growth, nutrient utilization, survival, and nutritional quality. | [52,223,224,225,226] | |
| Juveniles of gilthead seabream S. aurata. | Defatted Tetraselmis sp., T. chuii, Cellulose hydrolyzed N. gaditana, Schizochytrium sp., P. tricornutum, N. gaditana, Navicula sp., hydrolyzed A. platensis, and heat-treated Gracilaria sp., and Ulva sp. | Significantly improve S. aurata weight gain, specific growth rate, feed conversion ratio, survival, nutritional quality, innate immune parameters, and acute hypoxia tolerance. | [175,227,228,229,230,231,232] | |
| Salmonids Atlantic salmon, Salmo salar L. | N. oceanica, defatted N. oceanica, Scenedesmus sp., Schizochytrium sp. | Significantly improve weight gain, specific growth rate, improve FCR, survival, and fatty acid profiles. | [233,234,235] | |
| Juveniles and adults Rrainbow trout, Oncorhynchus mykiss. | Nannochloropsis sp., Schizochytrium sp., Isochrysis sp., S. almeriensis, U. lactuca, E. linza | Significantly increase weight gain, improve feed conversion ratio and survival rate. | [236,237,238] | |
| Shellfish | ||||
| Pacific white shrimp, L. vannamei, black tiger shrimp, P. monodon, and freshwater prawn, M. rosenbergii. | T. suecica, Schizochytrium sp., Aurantiochytrium sp., C. calcitrans, defatted H. pluvialis, D. salina, C. vulgaris, A. platensis, U. lactuca and Gracilaria sp. | Significantly enhance the growth, immune, nutrient utilization, survival, gene expression, nutritional quality, disease resistance, and improve tolerance stress. | [21,167,217,239,240,241,242,243,244,245] | |
| Sea urchin, T. gratilla and Heliocidaris erythrogramma. | E. radiata, S. linearifolium, Sargassum spp., Gracilaria sp., Solieria robusta, L. kuetzingii, and U. lactuca. | Significantly increase growth performances, feed intake, energy, protein consumption, and improve gonad indices and of T. gratilla. | [176,177] | |
| Water Conditioner | Larval Gilthead seabream, S. aurata | N. oculata (live and freeze-dried forms). N. gaditana and I. galbana (live and freeze-dried forms). | Applications of different forms (live and/or freeze-dried) of N. oculata in the rearing tanks significantly improved growth, survival, and histological status. The complete replacement of live algae in larval rearing tanks with freeze-dried algae resulted in the same survival and growth of larvae reared for 43 days. | [93,178,246] |
| Tilapia (mosambique and hybrid tilapia) Nile tilapia, O. niloticus | Nitzschia, Pleurosigma, Nannochloropsis and Oscillatoria. Seaweed liquid extract (TrueAlgaeMax, TAM®, prepared from U. lactuca, J. rubens, and P. capillacea) | Application of green water is effective in controlling the luminous bacteria. The dominant species of Nitzschia, Pleurosigma, Nannochloropsis, and Oscillatoria have a positive effect on the pathogenic bacteria in ponds and hatcheries, in general. It significantly enhances growth performance, immune responses, feed utilization, and zooplankton community and diversity. | [39,185] | |
| Giant tiger shrimp, P. monodon | Chlorella sp. | Reduce the white spot disease load in culture ponds of P. monodon. | [187] | |
| Seaweed Co-CultureandIntegration with Aquatic Animals | Black tiger shrimp, P. monodon, Nile tilapia, O. niloticus, Mediterranean mussels, M. galloprovincialis, Pacific white shrimp, L. vannamei, sea cucumber, H. scabra. | G. tenuistipitata, G. verrucosa, G. corticata, Ulva, K. striatum. | Improved water quality, shrimp survival, growth, antibacterial activities, and disease resistance significantly. | [46,190,194,195,196,197,198] |
| Black rockfish, Sebastes schlegeli. | U. pertusa, S. japonica, and Gracilariopsis chorda. | Seaweed significantly removed large amounts of nutrients (NH4, NO3, NO2, and PO4) from the fish tank effluents. | [191] | |
| Gilthead seabream, S. aurata, and sea urchin, P. lividus. | U. lactuca. | The IMTA systems between aquatic animals (S. aurata and P. lividus) and aquatic plants (U. lactuca) resulted in a lower FCR, and decreased growth period, enhanced gonadal development and quality, improved economic return, bioremediation efficiency, and decreased the treatment cost of the effluent. | [247] | |
| Cobia fish, Rachycentron canadum and The Brown mussel, Perna perna. | U. lactuca. | The IMTA systems between aquatic animals (R. canadum and P. perna) and aquatic plants (U. lactuca) increased yield and bioremediation efficiency. | [248] | |
| Grunt cab, Isacia conteptionis, Pacific oyster, C. gigas, and Chilean sea urchin, Loxechinus albus. | G. chilensis. | G. chilensis was highly effective in bioremediation of the soluble nutrients. | [249] | |
3. Future Prospective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2020. Sustainability in Action; Food and Agriculture Organization of the United Nations: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- Ashour, M.; Abo-Taleb, H.; Abou-Mahmoud, M.; El-Feky, M. Effect of the integration between plankton natural productivity and environmental assessment of irrigation water, El-Mahmoudia Canal, on aquaculture potential of Oreochromis niloticus. Turk. J. Fish. Aquat. Sci. 2018, 18, 1163–1175. [Google Scholar] [CrossRef]
- Shaalan, M.; El-Mahdy, M.; Saleh, M.; El-Matbouli, M. Aquaculture in Egypt: Insights on the current trends and future perspectives for sustainable development. Rev. Fish. Sci. Aquac. 2018, 26, 99–110. [Google Scholar] [CrossRef]
- Kaleem, O.; Sabi, A.-F.B.S. Overview of aquaculture systems in Egypt and Nigeria, prospects, potentials, and constraints. Aquac. Fish. 2020, 6, 535–547. [Google Scholar] [CrossRef]
- Ashour, M. Current and future perspectives of microalgae-aquaculture in Egypt, case study: SIMAF-prototype-project. Egypt. J. Anim. Prod. 2020, 57, 163–170. [Google Scholar]
- Barkia, I.; Saari, N.; Manning, S.R. Microalgae for high-value products towards human health and nutrition. Mar. Drugs 2019, 17, 304. [Google Scholar] [CrossRef] [Green Version]
- Tocher, D.R. Omega-3 long-chain polyunsaturated fatty acids and aquaculture in perspective. Aquaculture 2015, 449, 94–107. [Google Scholar] [CrossRef]
- Limbu, S.M.; Zhou, L.; Sun, S.-X.; Zhang, M.-L.; Du, Z.-Y. Chronic exposure to low environmental concentrations and legal aquaculture doses of antibiotics cause systemic adverse effects in Nile tilapia and provoke differential human health risk. Environ. Int. 2018, 115, 205–219. [Google Scholar] [CrossRef]
- Naylor, R.L.; Goldburg, R.J.; Primavera, J.H.; Kautsky, N.; Beveridge, M.C.; Clay, J.; Folke, C.; Lubchenco, J.; Mooney, H.; Troell, M. Effect of aquaculture on world fish supplies. Nature 2000, 405, 1017–1024. [Google Scholar] [CrossRef] [Green Version]
- Naylor, R.L.; Hardy, R.W.; Buschmann, A.H.; Bush, S.R.; Cao, L.; Klinger, D.H.; Little, D.C.; Lubchenco, J.; Shumway, S.E.; Troell, M. A 20-year retrospective review of global aquaculture. Nature 2021, 591, 551–563. [Google Scholar] [CrossRef]
- Alam, M.A.; Xu, J.-L.; Wang, Z. Microalgae Biotechnology for Food, Health and High Value Products; Springer: New York, NY, USA, 2020. [Google Scholar]
- Javed, M.R.; Bilal, M.J.; Ashraf, M.U.F.; Waqar, A.; Mehmood, M.A.; Saeed, M.; Nashat, N. Microalgae as a feedstock for biofuel production: Current status and future prospects. Top 5 Contrib. Energy Res. Dev. 2019, 5, 1–39. [Google Scholar]
- Muhammad, G.; Alam, M.A.; Xiong, W.; Lv, Y.; Xu, J.-L. Microalgae biomass production: An overview of dynamic operational methods. In Microalgae Biotechnology for Food, Health and High Value Products; Springer: Cham, Switzerland, 2020; pp. 415–432. [Google Scholar] [CrossRef]
- Kamani, M.H.; Eş, I.; Lorenzo, J.M.; Remize, F.; Roselló-Soto, E.; Barba, F.J.; Clark, J.; Khaneghah, A.M. Advances in plant materials, food by-products, and algae conversion into biofuels: Use of environmentally friendly technologies. Green Chem. 2019, 21, 3213–3231. [Google Scholar] [CrossRef]
- Santhakumaran, P.; Ayyappan, S.M.; Ray, J.G. Nutraceutical applications of twenty-five species of rapid-growing green-microalgae as indicated by their antibacterial, antioxidant and mineral content. Algal Res. 2020, 47, 101878. [Google Scholar] [CrossRef]
- Smita, P.; Gunjan, P.; Lali, A.M. Reduced chlorophyll antenna mutants of Chlorella saccharophila for higher photosynthetic efficiency and biomass productivity under high light intensities. J. Appl. Phycol. 2020, 32, 1559–1567. [Google Scholar]
- Tolboom, S.N.; Carrillo-Nieves, D.; de Jesús Rostro-Alanis, M.; de la Cruz Quiroz, R.; Barceló, D.; Iqbal, H.M.; Parra-Saldivar, R. Algal-based removal strategies for hazardous contaminants from the environment—A review. Sci. Total Environ. 2019, 665, 358–366. [Google Scholar] [CrossRef]
- Alprol, A.E.; Heneash, A.M.M.; Ashour, M.; Abualnaja, K.M.; Alhashmialameer, D.; Mansour, A.T.; Sharawy, Z.Z.; Abu-Saied, M.A.; Abomohra, A.E. Potential Applications of Arthrospira platensis Lipid-Free Biomass in Bioremediation of Organic Dye from Industrial Textile Effluents and Its Influence on Marine Rotifer (Brachionus plicatilis). Materials 2021, 14, 4446. [Google Scholar] [CrossRef]
- Ashour, M.; Alprol, A.E.; Heneash, A.M.M.; Saleh, H.; Abualnaja, K.M.; Alhashmialameer, D.; Mansour, A.T. Ammonia Bioremediation from Aquaculture Wastewater Effluents Using Arthrospira platensis NIOF17/003: Impact of Biodiesel Residue and Potential of Ammonia-Loaded Biomass as Rotifer Feed. Materials 2021, 14, 5460. [Google Scholar] [CrossRef]
- Abo-Taleb, H.A.; El-feky, M.M.M.; Azab, A.M.; Mabrouk, M.M.; Elokaby, M.A.; Ashour, M.; Mansour, A.T.; Abdelzaher, O.F.; Abualnaja, K.M.; Sallam, A.E. Growth Performance, Feed Utilization, Gut Integrity, and Economic Revenue of Grey Mullet, Mugil cephalus, Fed an Increasing Level of Dried Zooplankton Biomass Meal as Fishmeal Substitutions. Fishes 2021, 6, 38. [Google Scholar] [CrossRef]
- Sharawy, Z.Z.; Ashour, M.; Abbas, E.; Ashry, O.; Helal, M.; Nazmi, H.; Kelany, M.; Kamel, A.; Hassaan, M.; Rossi, W.; et al. Effects of dietary marine microalgae, Tetraselmis suecica, on production, gene expression, protein markers and bacterial count of Pacific white shrimp Litopenaeus vannamei. Aquacult. Res. 2020, 51, 2216–2228. [Google Scholar] [CrossRef]
- Sharawy, Z.Z.; Ashour, M.; Labena, A.; Alsaqufi, A.S.; Mansour, A.T.; Abbas, E.M. Effects of dietary Arthrospira platensis nanoparticles on growth performance, feed utilization, and growth-related gene expression of Pacific white shrimp, Litopenaeus vannamei. Aquaculture 2022, 551, 737905. [Google Scholar] [CrossRef]
- Abomohra, A.E.-F.; Almutairi, A.W. A close-loop integrated approach for microalgae cultivation and efficient utilization of agar-free seaweed residues for enhanced biofuel recovery. Bioresour. Technol. 2020, 317, 124027. [Google Scholar] [CrossRef]
- Ashour, M.; Mabrouk, M.M.; Ayoub, H.F.; El-Feky, M.M.; Zaki, S.Z.; Hoseinifar, S.H.; Rossi, W.; Van Doan, H.; El-Haroun, E.; Goda, A. Effect of dietary seaweed extract supplementation on growth, feed utilization, hematological indices, and non-specific immunity of Nile Tilapia, Oreochromis niloticus challenged with Aeromonas hydrophila. J. Appl. Phycol. 2020, 32, 3467–3479. [Google Scholar] [CrossRef]
- Barra, L.; Chandrasekaran, R.; Corato, F.; Brunet, C. The challenge of ecophysiological biodiversity for biotechnological applications of marine microalgae. Mar. Drugs 2014, 12, 1641–1675. [Google Scholar] [CrossRef]
- Elshobary, M.E.; El-Shenody, R.A.; Ashour, M.; Zabed, H.M.; Qi, X. Antimicrobial and antioxidant characterization of bioactive components from Chlorococcum minutum. Food Biosci. 2020, 35, 100567. [Google Scholar] [CrossRef]
- El-Shenody, R.A.; Ashour, M.; Ghobara, M.M.E. Evaluating the chemical composition and antioxidant activity of three Egyptian seaweeds: Dictyota dichotoma, Turbinaria decurrens, and Laurencia obtusa. Braz. J. Food Technol. 2019, 22, 1–15. [Google Scholar] [CrossRef] [Green Version]
- El-Shouny, W.; Sharaf, M.; Abomohra, A.; Abo-Eleneen, M. Production enhancement of some valuable compounds of Arthrospira platensis. J. Basic Environ. Sci. 2015, 2, 74–83. [Google Scholar]
- Choi, Y.Y.; Hong, M.-E.; Chang, W.S.; Sim, S.J. Autotrophic biodiesel production from the thermotolerant microalga Chlorella sorokiniana by enhancing the carbon availability with temperature adjustment. Biotechnol. Bioprocess Eng. 2019, 24, 223–231. [Google Scholar] [CrossRef]
- Abualnaja, K.M.; Alprol, A.E.; Abu-Saied, M.A.; Ashour, M.; Mansour, A.T. Removing of Anionic Dye from Aqueous Solutions by Adsorption Using of Multiwalled Carbon Nanotubes and Poly (Acrylonitrile-styrene) Impregnated with Activated Carbon. Sustainability 2021, 13, 7077. [Google Scholar] [CrossRef]
- Abualnaja, K.M.; Alprol, A.E.; Ashour, M.; Mansour, A.T. Influencing Multi-Walled Carbon Nanotubes for the Removal of Ismate Violet 2R Dye from Wastewater: Isotherm, Kinetics, and Thermodynamic Studies. Appl. Sci. 2021, 11, 4786. [Google Scholar] [CrossRef]
- Abualnaja, K.M.; Alprol, A.E.; Abu-Saied, M.; Mansour, A.T.; Ashour, M. Studying the Adsorptive Behavior of Poly (Acrylonitrile-co-Styrene) and Carbon Nanotubes (Nanocomposites) Impregnated with Adsorbent Materials towards Methyl Orange Dye. Nanomaterials 2021, 11, 1144. [Google Scholar] [CrossRef] [PubMed]
- Alprol, A.E. Study of environmental concerns of dyes and recent textile effluents treatment technology: A Review. Asian J. Fish. Aquat. Res. 2019, 3, 1–18. [Google Scholar] [CrossRef]
- Alprol, A.E.; Abd El Azzem, M.; Amer, A.; El-Metwally, M.E.; Abd El-Hamid, H.T.; El-Moselhy, K.M. Adsorption of cadmium (II) ions from aqueous solution onto mango leaves. Asian J. Phys. Chem. Sci. 2017, 2, 1–11. [Google Scholar] [CrossRef]
- Geddie, A.W.; Hall, S.G. Development of a suitability assessment model for the cultivation of intertidal macroalgae in the United States. Sci. Total Environ. 2020, 699, 134327. [Google Scholar] [CrossRef]
- Geddie, A.W.; Hall, S.G. The effect of salinity and alkalinity on growth and the accumulation of copper and zinc in the Chlorophyta Ulva fasciata. Ecotoxicol. Environ. Saf. 2019, 172, 203–209. [Google Scholar] [CrossRef]
- Geddie, A.W.; Hall, S.G. An introduction to copper and zinc pollution in macroalgae: For use in remediation and nutritional applications. J. Appl. Phycol. 2019, 31, 691–708. [Google Scholar] [CrossRef]
- Ashour, M.; Hassan, S.M.; Elshobary, M.E.; Ammar, G.A.G.; Gaber, A.; Alsanie, W.F.; Mansour, A.T.; El-Shenody, R. Impact of Commercial Seaweed Liquid Extract (TAM®) Biostimulant and Its Bioactive Molecules on Growth and Antioxidant Activities of Hot Pepper (Capsicum annuum). Plants 2021, 10, 1045. [Google Scholar] [CrossRef]
- Hassan, S.M.; Ashour, M.; Sakai, N.; Zhang, L.; Hassanien, H.A.; Gaber, A.; Ammar, G. Impact of Seaweed Liquid Extract Biostimulant on Growth, Yield, and Chemical Composition of Cucumber (Cucumis sativus). Agriculture 2021, 11, 320. [Google Scholar] [CrossRef]
- Hassan, S.M.; Ashour, M.; Soliman, A.A.F.; Hassanien, H.A.; Alsanie, W.F.; Gaber, A.; Elshobary, M.E. The Potential of a New Commercial Seaweed Extract in Stimulating Morpho-Agronomic and Bioactive Properties of Eruca vesicaria (L.) Cav. Sustainability 2021, 13, 4485. [Google Scholar] [CrossRef]
- Hassan, S.M.; Ashour, M.; Soliman, A. Anticancer Activity, Antioxidant Activity, Mineral Contents, Vegetative and Yield of Eruca sativa Using Foliar Application of Autoclaved Cellular Extract of Spirulina platensis Extract, Comparing to NPK Fertilizers. J. Plant Prod. 2017, 8, 529–536. [Google Scholar] [CrossRef] [Green Version]
- Vega, J.; Álvarez-Gómez, F.; Güenaga, L.; Figueroa, F.L.; Gómez-Pinchetti, J.L. Antioxidant activity of extracts from marine macroalgae, wild-collected and cultivated, in an integrated multi-trophic aquaculture system. Aquaculture 2020, 522, 735088. [Google Scholar] [CrossRef]
- Ardiles, P.; Cerezal-Mezquita, P.; Salinas-Fuentes, F.; Órdenes, D.; Renato, G.; Ruiz-Domínguez, M.C. Biochemical Composition and Phycoerythrin Extraction from Red Microalgae: A Comparative Study Using Green Extraction Technologies. Processes 2020, 8, 1628. [Google Scholar] [CrossRef]
- Araújo, R.; Vázquez Calderón, F.; Sánchez López, J.; Azevedo, I.C.; Bruhn, A.; Fluch, S.; Garcia Tasende, M.; Ghaderiardakani, F.; Ilmjärv, T.; Laurans, M. Current status of the algae production industry in Europe: An emerging sector of the Blue Bioeconomy. Front. Mar. Sci. 2021, 7, 1247. [Google Scholar] [CrossRef]
- Araújo, R.; Sanchez Lopez, J.; Landa, L.; Lusser, M. Report on the community of practice workshop: Algae production in europe: Status, challenges and future developments. In Proceedings of the European Commission’s Knowledge Centre for Bioeconomy, Brussels, Belgium, 27 February 2019; Volume 3, pp. 1–27. Available online: https://knowledge4policy.ec.europa.eu/publication/report-community-practice-workshop-algae-production-europe-status-challenges-future_en (accessed on 28 February 2022).
- An, B.N.T.; Anh, N.T.N. Co-culture of Nile tilapia (Oreochromis niloticus) and red seaweed (Gracilaria tenuistipitata) under different feeding rates: Effects on water quality, fish growth and feed efficiency. J. Appl. Phycol. 2020, 32, 2031–2040. [Google Scholar] [CrossRef]
- Ferdouse, F.; Holdt, S.L.; Smith, R.; Murúa, P.; Yang, Z. The global status of seaweed production, trade and utilization. Globefish Res. Programme 2018, 124, I. [Google Scholar]
- Shields, R.; Lupatsch, I. 5 Algae for aquaculture and animal feeds. In Microalgal Biotechnology: Integration and Economy; De Gruyter: Berlin, Germany, 2012; pp. 79–100. [Google Scholar] [CrossRef]
- Kaparapu, J. Application of microalgae in aquaculture. Phykos 2018, 48, 21–26. [Google Scholar]
- Wikfors, G.H.; Ohno, M. Impact of algal research in aquaculture. J. Phycol. 2001, 37, 968–974. [Google Scholar] [CrossRef]
- Delgado, E.; Valles-Rosales, D.J.; Flores, N.C.; Reyes-Jáquez, D. Evaluation of fish oil content and cottonseed meal with ultralow gossypol content on the functional properties of an extruded shrimp feed. Aquacult. Rep. 2021, 19, 100588. [Google Scholar] [CrossRef]
- Cardinaletti, G.; Messina, M.; Bruno, M.; Tulli, F.; Poli, B.; Giorgi, G.; Chini-Zittelli, G.; Tredici, M.; Tibaldi, E. Effects of graded levels of a blend of Tisochrysis lutea and Tetraselmis suecica dried biomass on growth and muscle tissue composition of European sea bass (Dicentrarchus labrax) fed diets low in fish meal and oil. Aquaculture 2018, 485, 173–182. [Google Scholar] [CrossRef]
- Yang, W.; Zheng, Z.; Lu, K.; Zheng, C.; Du, Y.; Wang, J.; Zhu, J. Manipulating the phytoplankton community has the potential to create a stable bacterioplankton community in a shrimp rearing environment. Aquaculture 2020, 520, 734789. [Google Scholar] [CrossRef]
- Hodar, A.; Vasava, R.; Mahavadiya, D.; Joshi, N. Fish meal and fish oil replacement for aqua feed formulation by using alternative sources: A review. J. Exp. Zool. India 2020, 23, 13–21. [Google Scholar]
- El-Sayed, A.-F.M. Evaluation of soybean meal, spirulina meal and chicken offal meal as protein sources for silver seabream (Rhabdosargus sarba) fingerlings. Aquaculture 1994, 127, 169–176. [Google Scholar] [CrossRef]
- Liu, W.-Y.; Fang, L.; Feng, X.-W.; Li, G.-M.; Gu, R.-Z. In vitro antioxidant and angiotensin I-converting enzyme inhibitory properties of peptides derived from corn gluten meal. Eur. Food Res. Technol. 2020, 246, 2017–2027. [Google Scholar] [CrossRef]
- Sørensen, M.; Morken, T.; Kosanovic, M.; Øverland, M. Pea and wheat starch possess different processing characteristics and affect physical quality and viscosity of extruded feed for Atlantic salmon. Aquacult. Nutr. 2011, 17, e326–e336. [Google Scholar] [CrossRef]
- Blomqvist, J.; Pickova, J.; Tilami, S.K.; Sampels, S.; Mikkelsen, N.; Brandenburg, J.; Sandgren, M.; Passoth, V. Oleaginous yeast as a component in fish feed. Sci. Rep. 2018, 8, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, R.; Cao, H.; Huang, Y.; Peng, M.; Kajbaf, K.; Kumar, V.; Tao, Z.; Yang, G.; Wen, C. The effects of partial replacement of fishmeal protein by hydrolysed feather meal protein in the diet with high inclusion of plant protein on growth performance, fillet quality and physiological parameters of Pengze crucian carp (Carassius auratus var. Pengze). Aquacult. Res. 2020, 51, 636–647. [Google Scholar]
- Abo-Taleb, H.A.; Zeina, A.F.; Ashour, M.; Mabrouk, M.M.; Sallam, A.E.; El-feky, M.M. Isolation and cultivation of the freshwater amphipod Gammarus pulex (Linnaeus, 1758), with an evaluation of its chemical and nutritional content. Egypt. J. Aquat. Biol. Fish. 2020, 24, 69–82. [Google Scholar] [CrossRef] [Green Version]
- Niccolai, A.; Zittelli, G.C.; Rodolfi, L.; Biondi, N.; Tredici, M.R. Microalgae of interest as food source: Biochemical composition and digestibility. Algal Res. 2019, 42, 101617. [Google Scholar] [CrossRef]
- Zaki, M.A.; Ashour, M.; Heneash, A.M.M.; Mabrouk, M.M.; Alprol, A.E.; Khairy, H.M.; Nour, A.M.; Mansour, A.T.; Hassanien, H.A.; Gaber, A.; et al. Potential Applications of Native Cyanobacterium Isolate (Arthrospira platensis NIOF17/003) for Biodiesel Production and Utilization of Its Byproduct in Marine Rotifer (Brachionus plicatilis) Production. Sustainability 2021, 13, 1769. [Google Scholar] [CrossRef]
- Tibbetts, S.M.; Milley, J.E.; Lall, S.P. Chemical composition and nutritional properties of freshwater and marine microalgal biomass cultured in photobioreactors. J. Appl. Phycol. 2015, 27, 1109–1119. [Google Scholar] [CrossRef] [Green Version]
- Ashour, M.; Elshobary, M.E.; El-Shenody, R.; Kamil, A.-W.; Abomohra, A.E.-F. Evaluation of a native oleaginous marine microalga Nannochloropsis oceanica for dual use in biodiesel production and aquaculture feed. Biomass Bioenergy 2019, 120, 439–447. [Google Scholar] [CrossRef]
- Becker, E.W. Micro-algae as a source of protein. Biotechnol. Adv. 2007, 25, 207–210. [Google Scholar] [CrossRef]
- Brown, M.R. The amino-acid and sugar composition of 16 species of microalgae used in mariculture. J. Exp. Mar. Biol. Ecol. 1991, 145, 79–99. [Google Scholar] [CrossRef]
- da Silva Gorg, C.M.; Aranda, D.A.G. Morphological and chemical aspects of Chlorella pyrenoidosa, Dunaliella tertiolecta, Isochrysis galbana and Tetraselmis gracilis microalgae. Nat. Sci. 2013, 5, 783–791. [Google Scholar] [CrossRef] [Green Version]
- Becker, E.W. Microalgae: Biotechnology and Microbiology; Cambridge University Press: Cambridge, UK, 1994; Volume 10. [Google Scholar]
- López, C.V.G.; García, M.d.C.C.; Fernández, F.G.A.; Bustos, C.S.; Chisti, Y.; Sevilla, J.M.F. Protein measurements of microalgal and cyanobacterial biomass. Bioresour. Technol. 2010, 101, 7587–7591. [Google Scholar] [CrossRef]
- Shah, M.; Mahfuzur, R.; Liang, Y.; Cheng, J.J.; Daroch, M. Astaxanthin-producing green microalga Haematococcus pluvialis: From single cell to high value commercial products. Front. Plant Sci. 2016, 7, 531. [Google Scholar] [CrossRef] [Green Version]
- Velasco, L.A.; Carrera, S.; Barros, J. Isolation, culture and evaluation of Chaetoceros muelleri from the Caribbean as food for the native scallops, Argopecten nucleus and Nodipecten nodosus. Lat. Am. J. Aquat. Res. 2016, 44, 557–568. [Google Scholar] [CrossRef]
- Tam, L.T.; Van Cong, N.; Thom, L.T.; Ha, N.C.; Hang, N.T.M.; Van Minh, C.; Vien, D.T.H.; Hong, D.D. Cultivation and biomass production of the diatom Thalassiosira weissflogii as a live feed for white-leg shrimp in hatcheries and commercial farms in Vietnam. J. Appl. Phycol. 2021, 33, 1559–1577. [Google Scholar] [CrossRef]
- Kawaroe, M.; Salundik, R.W.; Lestari, D.F. Comparison of Biogas Productionfrom Macroalgae Eucheuma cottonii in Anaerobic Degradation under Different Salinity Conditions. World Appl. Sci. J. 2017, 35, 344–351. [Google Scholar]
- Muraguri, E.N.; Wakibia, J.G.; Kinyuru, J.N. Chemical composition and functional properties of selected seaweeds from the Kenya coast. J. Food Res. 2016, 5, 114. [Google Scholar] [CrossRef]
- Khalil, H.A.; Yap, S.W.; Tye, Y.Y.; Tahir, P.M.; Rizal, S.; Fazita, M.N. Effects of corn starch and Kappaphycus alvarezii seaweed blend concentration on the optical, mechanical, and water vapor barrier properties of composite films. BioResources 2018, 13, 1157–1173. [Google Scholar]
- Xiren, G.; Aminah, A. Proximate composition and total amino acid composition of Kappaphycus alvarezii found in the waters of Langkawi and Sabah, Malaysia. Int. Food Res. J. 2017, 24, 1255. [Google Scholar]
- Arguelles, E.; Monsalud, R.G.; Sapin, A.B. Chemical composition and in vitro antioxidant and antibacterial activities of Sargassum vulgare C. Agardh from Lobo, Batangas, Philippines. J. Int. Soc. Southeast Asian Agric. Sci. (ISSAAS) 2019, 25, 112–122. [Google Scholar]
- Kasimala, M.; Mogos, G.; Negasi, K.; Bereket, G.; Abdu, M.; Melake, H. Biochemical Composition of Selected Seaweeds from Intertidal Shallow Waters of Southern Red Sea, Eritrea; NISCAIR-CSIR: New Delhi, India, 2020. [Google Scholar]
- Benjama, O.; Masniyom, P. Biochemical composition and physicochemical properties of two red seaweeds (Gracilaria fisheri and G. tenuistipitata) from the Pattani Bay in Southern Thailand. Sonklanakarin J. Sci. Technol. 2012, 34, 223. [Google Scholar]
- Khairy, H.M.; El-Shafay, S.M. Seasonal variations in the biochemical composition of some common seaweed species from the coast of Abu Qir Bay, Alexandria, Egypt. Oceanologia 2013, 55, 435–452. [Google Scholar] [CrossRef] [Green Version]
- Crespi, V.; New, M. Cultured Aquatic Species Fact Sheets; FAO: Rome, Italy, 2009. [Google Scholar]
- Admassu, H.; Abera, T.; Abraha, B.; Yang, R.; Zhao, W. Proximate, mineral and amino acid composition of dried laver (Porphyra spp.) seaweed. J. Acad. Ind. Res. (JAIR) 2018, 6, 149. [Google Scholar]
- Cho, T.J.; Rhee, M.S. Health functionality and quality control of laver (Porphyra, Pyropia): Current issues and future perspectives as an edible seaweed. Mar. Drugs 2020, 18, 14. [Google Scholar] [CrossRef] [Green Version]
- Spolaore, P.; Joannis-Cassan, C.; Duran, E.; Isambert, A. Commercial applications of microalgae. J. Biosci. Bioeng. 2006, 101, 87–96. [Google Scholar] [CrossRef] [Green Version]
- Duy, N.D.Q.; Francis, D.S.; Southgate, P.C. The nutritional value of live and concentrated micro-algae for early juveniles of sandfish, Holothuria scabra. Aquaculture 2017, 473, 97–104. [Google Scholar] [CrossRef]
- Brown, M.; Robert, R. Preparation and assessment of microalgal concentrates as feeds for larval and juvenile Pacific oyster (Crassostrea gigas). Aquaculture 2002, 207, 289–309. [Google Scholar] [CrossRef]
- Alprol, A.E.; Ashour, M.; Mansour, A.T.; Alzahrani, O.M.; Mahmoud, S.F.; Gharib, S.M. Assessment of Water Quality and Phytoplankton Structure of Eight Alexandria Beaches, Southeastern Mediterranean Sea, Egypt. J. Mar. Sci. Eng. 2021, 9, 1328. [Google Scholar] [CrossRef]
- Abo-Taleb, H.; Ashour, M.; El-Shafei, A.; Alataway, A.; Maaty, M.M. Biodiversity of Calanoida Copepoda in Different Habitats of the North-Western Red Sea (Hurghada Shelf). Water 2020, 12, 656. [Google Scholar] [CrossRef] [Green Version]
- Alprol, A.E.; Heneash, A.M.M.; Soliman, A.M.; Ashour, M.; Alsanie, W.F.; Gaber, A.; Mansour, A.T. Assessment of Water Quality, Eutrophication, and Zooplankton Community in Lake Burullus, Egypt. Diversity 2021, 13, 268. [Google Scholar] [CrossRef]
- Abo-Taleb, H.A.; Ashour, M.; Elokaby, M.A.; Mabrouk, M.M.; El-feky, M.M.M.; Abdelzaher, O.F.; Gaber, A.; Alsanie, W.F.; Mansour, A.T. Effect of a New Feed Daphnia magna (Straus, 1820), as a Fish Meal Substitute on Growth, Feed Utilization, Histological Status, and Economic Revenue of Grey Mullet, Mugil cephalus (Linnaeus 1758). Sustainability 2021, 13, 7093. [Google Scholar] [CrossRef]
- Abdel-Rahim, M.M.; Mansour, A.T.; Mona, M.H.; El-Gamal, M.M.; El Atafy, M.M. To what extent can maternal inherited immunity acquired from a crustacean-enhanced diet improve the performance and vitality of the offspring and enhance profitability of European Sea bass (Dicentrarchus labrax)? J. World Aquacult. Soc. 2019, 50, 550–574. [Google Scholar] [CrossRef]
- El-Gamal, M.M.; Othman, S.I.; Abdel-Rahim, M.M.; Mansour, A.T.; Alsaqufi, A.S.; El Atafy, M.M.; Mona, M.H.; Allam, A.A. Palaemon and artemia supplemented diet enhances sea bass, Dicentrarchus labrax, broodstock reproductive performance and egg quality. Aquacult. Rep. 2020, 16, 100290. [Google Scholar] [CrossRef]
- Navarro, N.; Sarasquete, C. Use of freeze-dried microalgae for rearing gilthead seabream, Sparus aurata, larvae: I. Growth, histology and water quality. Aquaculture 1998, 167, 179–193. [Google Scholar] [CrossRef]
- Zaki, M.; Saad, H. Comparative study on growth and survival of larval and juvenile Dicentrarchus labrax rearing on rotifer and Artemia enriched with four different microalgae species. Afr. J. Biotechnol. 2010, 9, 3676–3688. [Google Scholar]
- Camus, T.; Zeng, C. Roles of microalgae on total egg production over female lifespan and egg incubation time, naupliar and copepodite survival, sex ratio and female life expectancy of the copepod Bestiolina similis. Aquacult. Res. 2010, 41, 1717–1726. [Google Scholar] [CrossRef]
- Ianora, A.; Romano, G.; Carotenuto, Y.; Esposito, F.; Roncalli, V.; Buttino, I.; Miralto, A. Impact of the diatom oxylipin 15S-HEPE on the reproductive success of the copepod Temora stylifera. Hydrobiologia 2011, 666, 265–275. [Google Scholar] [CrossRef]
- Ananth, S.; Santhanam, P. Intensive Culture, Biochemical Composition Analysis, and Use of Zooplankton Tisbe sp.(Copepoda: Harpacticoida) as an Alternative Live Feed for Shrimp Larviculture. In Basic and Applied Zooplankton Biology; Springer: New York, NY, USA, 2019; pp. 329–362. [Google Scholar] [CrossRef]
- Jothiraj, K.; Santhanam, P. Optimisation of the Culture Conditions of Nannocalanus minor (Copepoda: Calanoida). In Basic and Applied Zooplankton Biology; Springer: New York, NY, USA, 2019; pp. 225–246. [Google Scholar] [CrossRef]
- Schipp, G.R.; Bosmans, J.M.; Marshall, A.J. A method for hatchery culture of tropical calanoid copepods, Acartia spp. Aquaculture 1999, 174, 81–88. [Google Scholar] [CrossRef]
- Altaff, K.; Vijayaraj, R. Micro-Algal Diet for Copepod Culture with Reference to Their Nutritive Value—A Review. Int. J. Cur. Res. Rev. Vol. 2021, 13, 86. [Google Scholar]
- Magouz, F.I.; Essa, M.A.; Matter, M.; Mansour, A.T.; Gaber, A.; Ashour, M. Effect of Different Salinity Levels on Population Dynamics and Growth of the Cyclopoid Copepod Oithona nana. Diversity 2021, 13, 190. [Google Scholar] [CrossRef]
- Ferreira, M.; Coutinho, P.; Seixas, P.; Fábregas, J.; Otero, A. Enriching rotifers with “premium” microalgae. Nannochloropsis gaditana. Mar. Biotechnol. 2009, 11, 585–595. [Google Scholar] [CrossRef]
- Palmtag, M.R.; Faulk, C.K.; Holt, G. Highly unsaturated fatty acid composition of rotifers (Brachionus plicatilis) and Artemia fed various enrichments. J. World Aquacult. Soc. 2006, 37, 126–131. [Google Scholar] [CrossRef]
- Seychelles, L.; Audet, C.; Tremblay, R.; Fournier, R.; Pernet, F. Essential fatty acid enrichment of cultured rotifers (Brachionus plicatilis, Müller) using frozen-concentrated microalgae. Aquacult. Nutr. 2009, 15, 431–439. [Google Scholar] [CrossRef]
- Abbas, E.M.; Ali, F.S.; Desouky, M.G.; Ashour, M.; El-Shafei, A.; Maaty, M.M.; Sharawy, Z.Z. Novel Comprehensive Molecular and Ecological Study Introducing Coastal Mud Shrimp (Solenocera Crassicornis) Recorded at the Gulf of Suez, Egypt. J. Mar. Sci. Eng. 2020, 9, 9. [Google Scholar] [CrossRef]
- Sivakumar, N.; Sundararaman, M.; Selvakumar, G. Efficacy of micro algae and cyanobacteria as a live feed for juveniles of shrimp Penaeus monodon. Afr. J. Biotechnol. 2011, 10, 11594–11599. [Google Scholar]
- Rodríguez-Núñez, K.; Toledo-Aguero, P. Fatty acids profile and nutritional composition of two tropical diatoms from the Costa Rican Pacific Coast. Grasas Aceites 2017, 68, e209. [Google Scholar] [CrossRef]
- Rohani-Ghadikolaei, K.; Abdolalian, E.; Hojatollah, F.; Masoud, G.; Ng, W.K. The nutritional effect of Isochrysis galbana and Chaetoceros muelleri cultured with different seaweed extracts on the larval development, growth and survival of the marine shrimp, Penaeus indicus. Aquacult. Res. 2015, 46, 1444–1454. [Google Scholar] [CrossRef]
- Abdel Rahman, S.; Abdel Razek, F.; AbouZeid, A.; Ashour, M. Optimum growth conditions of three isolated diatoms species; Skeletonema costatum, Chaetoceros calcitrans and Detonula confervacea and their utilization as feed for marine penaeid shrimp larvae. Egypt. J. Aquat. Res 2010, 36, 161–183. [Google Scholar]
- Pratiwi, D.Y. A mini review-effect of Dunaliella salina on growth and health of shrimps. Int. J. Fish. Aquat. Stud. 2020, 8, 317–319. [Google Scholar]
- Muller-Feuga, A.; Robert, R.; Cahu, C.; Robin, J.; Divanach, P. Uses of microalgae in aquaculture. Live Feed. Mar. Aquac. 2003, 1, 253–299. [Google Scholar]
- Mai, T.D.; Lee-Chang, K.J.; Jameson, I.D.; Hoang, T.; Cai, N.B.A.; Pham, H.Q. Fatty acid profiles of selected microalgae used as live feeds for shrimp postlarvae in Vietnam. Aquac. J. 2021, 1, 26–38. [Google Scholar] [CrossRef]
- Ponis, E.; Robert, R.; Parisi, G.; Tredici, M. Assessment of the performance of Pacific oyster (Crassostrea gigas) larvae fed with fresh and preserved Pavlova lutheri concentrates. Aquacult. Int. 2003, 11, 69–79. [Google Scholar] [CrossRef]
- Knuckey, R.M.; Brown, M.R.; Robert, R.; Frampton, D.M. Production of microalgal concentrates by flocculation and their assessment as aquaculture feeds. Aquacult. Eng. 2006, 35, 300–313. [Google Scholar] [CrossRef] [Green Version]
- McCausland, M.A.; Brown, M.R.; Barrett, S.M.; Diemar, J.A.; Heasman, M.P. Evaluation of live microalgae and microalgal pastes as supplementary food for juvenile Pacific oysters (Crassostrea gigas). Aquaculture 1999, 174, 323–342. [Google Scholar] [CrossRef]
- Heasman, M.; Diemar, J.; O’connor, W.; Sushames, T.; Foulkes, L. Development of extended shelf-life microalgae concentrate diets harvested by centrifugation for bivalve molluscs—A summary. Aquacult. Res. 2000, 31, 637–659. [Google Scholar]
- Laing, I.; Verdugo, C.G. Nutritional value of spray-dried Tetraselmis suecica for juvenile bivalves. Aquaculture 1991, 92, 207–218. [Google Scholar] [CrossRef]
- Laing, I.; Child, A.R.; Janke, A. Nutritional value of dried algae diets for larvae of Manila clam (Tapes philippinarum). J. Mar. Biol. Assoc. UK 1990, 70, 1–12. [Google Scholar] [CrossRef]
- Laing, I.; Millican, P.F. Indoor nursery cultivation of juvenile bivalve molluscs using diets of dried algae. Aquaculture 1992, 102, 231–243. [Google Scholar] [CrossRef]
- Boeing, P.; Escondido, C. Use of spray-dried Schizochytrium sp. as a partial algal replacement for juvenile bivalves. J. Shellfish Res 1997, 16, 284. [Google Scholar]
- Langdon, C.; Önal, E. Replacement of living microalgae with spray-dried diets for the marine mussel Mytilus galloprovincialis. Aquaculture 1999, 180, 283–294. [Google Scholar] [CrossRef]
- Albentosa, M.; Pérez-Camacho, A.; Labarta, U.; Fernández-Reiriz, M.J. Evaluation of freeze-dried microalgal diets for the seed culture of Ruditapes decussatus using physiological and biochemical parameters. Aquaculture 1997, 154, 305–321. [Google Scholar] [CrossRef] [Green Version]
- Arney, B.; Liu, W.; Forster, I.P.; McKinley, R.S.; Pearce, C.M. Feasibility of dietary substitution of live microalgae with spray-dried Schizochytrium sp. or Spirulina in the hatchery culture of juveniles of the Pacific geoduck clam (Panopea generosa). Aquaculture 2015, 444, 117–133. [Google Scholar] [CrossRef]
- Militz, T.A.; Braley, R.D.; Southgate, P.C. Factors influencing the capacity for pediveliger larvae of the giant clam, Tridacna noae, to ingest and digest cells of microalgae concentrates. Aquaculture 2021, 533, 736121. [Google Scholar] [CrossRef]
- Mélo, R.C.S.; Santos, L.P.d.S.; Brito, A.P.M.; Gouveia, A.d.A.; Marçal, C.; Cavalli, R.O. Use of the microalga Nannochloropsis occulata in the rearing of newborn longsnout seahorse Hippocampus reidi (Syngnathidae) juveniles. Aquacult. Res. 2016, 47, 3934–3941. [Google Scholar] [CrossRef]
- Martínez-Fernández, E.; Southgate, P.C. Use of tropical microalgae as food for larvae of the black-lip pearl oyster Pinctada margaritifera. Aquaculture 2007, 263, 220–226. [Google Scholar] [CrossRef]
- Bordbar, S.; Anwar, F.; Saari, N. High-value components and bioactives from sea cucumbers for functional foods—A review. Mar. Drugs 2011, 9, 1761–1805. [Google Scholar] [CrossRef] [Green Version]
- Abdelaty, M.; Al-Solami, L.; Al-Harbi, M.; Abu El-Regal, M. Utilization of Different Types of Microalgae to Improve Hatcheries Production of the Sea Cucumber Holothuria scabra Jaeger, 1833 in the Red Sea, Egypt. Egypt. J. Aquat. Biol. Fish. 2021, 25, 193–204. [Google Scholar] [CrossRef]
- Shi, C.; Dong, S.; Wang, F.; Gao, Q.; Tian, X. Effects of four fresh microalgae in diet on growth and energy budget of juvenile sea cucumber Apostichopus japonicus (Selenka). Aquaculture 2013, 416, 296–301. [Google Scholar] [CrossRef]
- Ren, Y.; Liu, W.; Dong, S.; Pearce, C. Effects of mono-species and bi-species microalgal diets on the growth, survival and metamorphosis of auricularia larvae of the California sea cucumber, Parastichopus californicus (Stimpson, 1857). Aquacult. Nutr. 2016, 22, 304–314. [Google Scholar] [CrossRef]
- Sibonga, R.C.; Laureta, L.V.; Lebata-Ramos, M.J.H.; Nievales, M.F.J.; Pedroso, F.L. Single and mixed species of microalgae as larval food for the tropical sea cucumber Holothuria scabra. J. Appl. Phycol. 2021, 33, 3103–3112. [Google Scholar] [CrossRef]
- Schagerström, E.; Christophersen, G.; Sunde, J.; Bakke, S.; Matusse, N.R.; Dupont, S.; Sundell, K.S. Controlled spawning and rearing of the sea cucumber, Parastichopus tremulus. J. World Aquacult. Soc. 2021, 53, 224–240. [Google Scholar] [CrossRef]
- Liu, H.; Kelly, M.S.; Cook, E.J.; Black, K.; Orr, H.; Zhu, J.X.; Dong, S.L. The effect of diet type on growth and fatty-acid composition of sea urchin larvae, I. Paracentrotus lividus (Lamarck, 1816) (Echinodermata). Aquaculture 2007, 264, 247–262. [Google Scholar] [CrossRef]
- Willadino, L.; Souza-Santos, L.P.; Mélo, R.C.; Brito, A.P.; Barros, N.C.; Araújo-Castro, C.M.; Galvão, D.B.; Gouveia, A.; Regis, C.G.; Cavalli, R.O. Ingestion rate, survival and growth of newly released seahorse Hippocampus reidi fed exclusively on cultured live food items. Aquaculture 2012, 360, 10–16. [Google Scholar] [CrossRef]
- Scholtz, R.; Bolton, J.; Macey, B. Effects of different microalgal feeds and their influence on larval development in the white-spined sea urchin Tripneustes gratilla. Afr. J. Mar. Sci. 2013, 35, 25–34. [Google Scholar] [CrossRef]
- Carboni, S.; Vignier, J.; Chiantore, M.; Tocher, D.R.; Migaud, H. Effects of dietary microalgae on growth, survival and fatty acid composition of sea urchin Paracentrotus lividus throughout larval development. Aquaculture 2012, 324, 250–258. [Google Scholar] [CrossRef]
- Shigai, X.; Chuanxin, Q.; Zhenhua, M.; Gang, Y.; Jinhui, S.; Wanni, P.; Tao, Z.; Hongmei, M.; Wentao, Z. Effects of dietary microalgae on growth and survival of larval development of sea urchin (Anthocidaris crassispina). South China Fish. Sci. 2020, 16, 115–120. [Google Scholar]
- Koldewey, H. Syngnathid Husbandry in Public Aquariums 2005 Manual; Project Seahorse: London, UK, 2005. [Google Scholar]
- Olivotto, I.; Avella, M.A.; Sampaolesi, G.; Piccinetti, C.; Ruiz, P.N.; Carnevali, O. Breeding and rearing the longsnout seahorse Hippocampus reidi: Rearing and feeding studies. Aquaculture 2008, 283, 92–96. [Google Scholar] [CrossRef]
- Pham, N.K.; Lin, J. The effects of different feed enrichments on survivorship and growth of early juvenile longsnout seahorse, Hippocampus reidi. J. World Aquacult. Soc. 2013, 44, 435–446. [Google Scholar] [CrossRef]
- Oostlander, P.; van Houcke, J.; Wijffels, R.H.; Barbosa, M.J. Microalgae production cost in aquaculture hatcheries. Aquaculture 2020, 525, 735310. [Google Scholar] [CrossRef]
- Heneash, A.; Ashour, M.; Matar, M. Effect of Un-live Microalgal diet, Nannochloropsis oculata and Arthrospira (Spirulina) platensis, comparing to yeast on population of rotifer, Brachionus plicatilis. Mediterr. Aquac. J. 2015, 7, 48–54. [Google Scholar] [CrossRef]
- Amjad, S.; Jones, D. An evaluation of artificial larval diets used in the culture of penaeid shrimp larvae Penaeus monodon (Fabricius). Pak. J. Zool. 1992, 24, 135. [Google Scholar]
- Medina-Reyna, C.E.; Ronson-Paulin, J.A.; Hernandez-Rojas, F.; Santiago-Morales, I.; Pedroza-Islas, R.; Vernon-Carter, E.J. Dual benefits of whey protein concentrate in a microencapsulated diet for larval white shrimp, Litopenaeus vannamei. J. World Aquacult. Soc. 2005, 36, 401–410. [Google Scholar] [CrossRef]
- Raja, R.; Coelho, A.; Hemaiswarya, S.; Kumar, P.; Carvalho, I.S.; Alagarsamy, A. Applications of microalgal paste and powder as food and feed: An update using text mining tool. Beni-Suef Univ. J. Basic Appl. Sci. 2018, 7, 740–747. [Google Scholar] [CrossRef]
- Nunes, M.; Pereira, A.; Ferreira, J.F.; Yasumaru, F. Evaluation of the microalgae paste viability produced in a mollusk hatchery in Southern Brazil. J. World Aquacult. Soc. 2009, 40, 87–94. [Google Scholar] [CrossRef]
- Metwally, A.S.; El-Naggar, H.A.; El-Damhougy, K.A.; Bashar, M.A.E.; Ashour, M.; Abo-Taleb, H.A.H. GC-MS analysis of bioactive components in six different crude extracts from the Soft Coral (Sinularia maxim) collected from Ras Mohamed, Aqaba Gulf, Red Sea, Egypt. Egypt. J. Aquat. Biol. Fish 2020, 24, 425–434. [Google Scholar] [CrossRef]
- Rani, A.; Saini, K.C.; Bast, F.; Mehariya, S.; Bhatia, S.K.; Lavecchia, R.; Zuorro, A. Microorganisms: A potential source of bioactive molecules for antioxidant applications. Molecules 2021, 26, 1142. [Google Scholar] [CrossRef]
- Knuckey, R.M.; Semmens, G.L.; Mayer, R.J.; Rimmer, M.A. Development of an optimal microalgal diet for the culture of the calanoid copepod Acartia sinjiensis: Effect of algal species and feed concentration on copepod development. Aquaculture 2005, 249, 339–351. [Google Scholar] [CrossRef]
- Puello-Cruz, A.; Mezo-Villalobos, S.; Gonzalez-Rodriguez, B.; Voltolina, D. Culture of the calanoid copepod Pseudodiaptomus euryhalinus (Johnson 1939) with different microalgal diets. Aquaculture 2009, 290, 317–319. [Google Scholar] [CrossRef]
- Rasdi, N.W.; Qin, J.G.; Li, Y. Effects of dietary microalgae on fatty acids and digestive enzymes in copepod C yclopina kasignete, a potential live food for fish larvae. Aquacult. Res. 2016, 47, 3254–3264. [Google Scholar] [CrossRef]
- Millamena, O.M.; Aujero, E.J.; Borlcmgan, I.G. Techniques on algae harvesting and preservation for use in culture and as larval food. Aquacult. Eng. 1990, 9, 295–304. [Google Scholar] [CrossRef]
- Robinson, C.; Samocha, T.; Fox, J.; Gandy, R.; McKee, D. The use of inert artificial commercial food sources as replacements of traditional live food items in the culture of larval shrimp, Farfantepenaeus aztecus. Aquaculture 2005, 245, 135–147. [Google Scholar] [CrossRef]
- Tang, Y.; Wang, R.; Tan, L.; Guo, L.; Duan, Y.; Yang, L.; Jiang, S.; Zhou, F.; Jiang, S.; Huang, J. Effects of live microalgae and algae powder on microbial community, survival, metamorphosis and digestive enzyme activity of Penaeus monodon larvae at different growth stages. Aquaculture 2020, 526, 735344. [Google Scholar] [CrossRef]
- Nimrat, S.; Boonthai, T.; Vuthiphandchai, V. Effects of probiotic forms, compositions of and mode of probiotic administration on rearing of Pacific white shrimp (Litopenaeus vannamei) larvae and postlarvae. Anim. Feed Sci. Technol. 2011, 169, 244–258. [Google Scholar] [CrossRef]
- Kumlu, M.; Jones, D. The effect of live and artificial diets on growth, survival, and trypsin activity in larvae of Penaeus indicus. J. World Aquacult. Soc. 1995, 26, 406–415. [Google Scholar] [CrossRef]
- Ponis, E.; Parisi, G.; Zittelli, G.C.; Lavista, F.; Robert, R.; Tredici, M. Pavlova lutheri: Production, preservation and use as food for Crassostrea gigas larvae. Aquaculture 2008, 282, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Militz, T.A.; Leini, E.; Duy, N.D.Q.; Southgate, P.C. Successful large-scale hatchery culture of sandfish (Holothuria scabra) using micro-algae concentrates as a larval food source. Aquacult. Rep. 2018, 9, 25–30. [Google Scholar] [CrossRef]
- Yu, Z.; Robinson, S.; MacDonald, B.; Lander, T.; Smith, C. Effect of diets on the feeding behavior and physiological properties of suspension-feeding sea cucumber Cucumaria frondosa. J. Oceanol. Limnol. 2020, 38, 883–893. [Google Scholar] [CrossRef]
- Floreto, E.A.; Teshima, S.-I.; Ishikawa, M. The Effects of seaweed diets on the growth, lipid and acids of juveniles of the white sea Urchin Tripneustes gratilla. Fish. Sci. 1996, 62, 589–593. [Google Scholar] [CrossRef] [Green Version]
- Lyons, D.A.; Scheibling, R.E. Effect of dietary history and algal traits on feeding rate and food preference in the green sea urchin Strongylocentrotus droebachiensis. J. Exp. Mar. Biol. Ecol. 2007, 349, 194–204. [Google Scholar] [CrossRef]
- Sarker, P.K.; Kapuscinski, A.R.; Bae, A.Y.; Donaldson, E.; Sitek, A.J.; Fitzgerald, D.S.; Edelson, O.F. Towards sustainable aquafeeds: Evaluating substitution of fishmeal with lipid-extracted microalgal co-product (Nannochloropsis oculata) in diets of juvenile Nile tilapia (Oreochromis niloticus). PLoS ONE 2018, 13, e0201315. [Google Scholar] [CrossRef] [PubMed]
- Mabrouk, M.M.; Ashour, M.; Labena, A.; Zaki, M.A.A.; Abdelhamid, A.F.; Gewaily, M.S.; Dawood, M.A.O.; Abualnaja, K.M.; Ayoub, H.F. Nanoparticles of Arthrospira platensis improves growth, antioxidative and immunological responses of Nile tilapia (Oreochromis niloticus) and its resistance to Aeromonas hydrophila. Aquacult. Res. 2022, 53, 125–135. [Google Scholar] [CrossRef]
- Nagappan, S.; Das, P.; AbdulQuadir, M.; Thaher, M.; Khan, S.; Mahata, C.; Al-Jabri, H.; Vatland, A.K.; Kumar, G. Potential of microalgae as a sustainable feed ingredient for aquaculture. J. Biotechnol. 2021, 341, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Skrede, A.; Mydland, L.; Ahlstrøm, Ø.; Reitan, K.; Gislerød, H.; Øverland, M. Evaluation of microalgae as sources of digestible nutrients for monogastric animals. J. Anim. Feed. Sci. 2011, 20, 131–142. [Google Scholar] [CrossRef]
- Souza, F.P.d.; Lima, E.C.S.d.; Urrea-Rojas, A.M.; Suphoronski, S.A.; Facimoto, C.T.; Bezerra Júnior, J.d.S.; Oliveira, T.E.S.d.; Pereira, U.d.P.; Santis, G.W.D.; Oliveira, C.A.L.d. Effects of dietary supplementation with a microalga (Schizochytrium sp.) on the hemato-immunological, and intestinal histological parameters and gut microbiota of Nile tilapia in net cages. PLoS ONE 2020, 15, e0226977. [Google Scholar] [CrossRef] [Green Version]
- Medina-Félix, D.; López-Elías, J.A.; Martínez-Córdova, L.R.; López-Torres, M.A.; Hernández-López, J.; Rivas-Vega, M.E.; Mendoza-Cano, F. Evaluation of the productive and physiological responses of Litopenaeus vannamei infected with WSSV and fed diets enriched with Dunaliella sp. J. Invertebr. Pathol. 2014, 117, 9–12. [Google Scholar] [CrossRef]
- Maliwat, G.C.; Velasquez, S.; Robil, J.L.; Chan, M.; Traifalgar, R.F.; Tayamen, M.; Ragaza, J.A. Growth and immune response of giant freshwater prawn Macrobrachium rosenbergii (De Man) postlarvae fed diets containing Chlorella vulgaris (Beijerinck). Aquacult. Res. 2017, 48, 1666–1676. [Google Scholar] [CrossRef]
- Cerezuela, R.; Guardiola, F.A.; Meseguer, J.; Esteban, M.A. Enrichment of gilthead seabream (Sparus aurata L.) diet with microalgae: Effects on the immune system. Fish Physiol. Biochem. 2012, 38, 1729–1739. [Google Scholar] [CrossRef]
- Das, B.K.; Pradhan, J.; Sahu, S. The effect of Euglena viridis on immune response of rohu, Labeo rohita (Ham.). Fish Shellfish Immunol. 2009, 26, 871–876. [Google Scholar] [CrossRef]
- Ansarifard, F.; Rajabi Islami, H.; Shamsaie Mehrjan, M.; Soltani, M. Effects of Arthrospira platensis on growth, skin color and digestive enzymes of Koi, Cyprinus carpio. Iran. J. Fish. Sci. 2018, 17, 381–393. [Google Scholar]
- Dineshbabu, G.; Goswami, G.; Kumar, R.; Sinha, A.; Das, D. Microalgae–nutritious, sustainable aqua-and animal feed source. J. Funct. Foods 2019, 62, 103545. [Google Scholar] [CrossRef]
- Liu, C.; Li, Y.; Chen, Z.; Yuan, L.; Liu, H.; Han, D.; Jin, J.; Yang, Y.; Hu, Q.; Zhu, X. Effects of dietary whole and defatted Arthrospira platensis (Cyanobacterium) on growth, body composition and pigmentation of the yellow catfish Pelteobagrus fulvidraco. J. Appl. Phycol. 2021, 33, 2251–2259. [Google Scholar] [CrossRef]
- Sun, X.; Chang, Y.; Ye, Y.; Ma, Z.; Liang, Y.; Li, T.; Jiang, N.; Xing, W.; Luo, L. The effect of dietary pigments on the coloration of Japanese ornamental carp (koi, Cyprinus carpio L.). Aquaculture 2012, 342, 62–68. [Google Scholar] [CrossRef]
- Ribeiro, A.R.; Gonçalves, A.; Barbeiro, M.; Bandarra, N.; Nunes, M.L.; Carvalho, M.L.; Silva, J.; Navalho, J.; Dinis, M.T.; Silva, T. Phaeodactylum tricornutum in finishing diets for gilthead seabream: Effects on skin pigmentation, sensory properties and nutritional value. J. Appl. Phycol. 2017, 29, 1945–1956. [Google Scholar] [CrossRef]
- Dworjanyn, S.A.; Pirozzi, I.; Liu, W. The effect of the addition of algae feeding stimulants to artificial diets for the sea urchin Tripneustes gratilla. Aquaculture 2007, 273, 624–633. [Google Scholar] [CrossRef]
- Warren-Myers, F.; Turchini, G.; Swearer, S.E.; Francis, D.; Dempster, T. The balancing act: Protein, lipid and seaweed dietary levels to maximize gonad quantity in a wild-caught sea urchin. Aquacult. Nutr. 2021, 27, 1019–1030. [Google Scholar] [CrossRef]
- Navarro, N.; Yúfera, M.; García-Gallego, M. Use of freeze-dried microalgae for rearing gilthead seabream, Sparus aurata L., larvae. II. Biochemical composition. Hydrobiologia 2001, 452, 69–77. [Google Scholar] [CrossRef]
- Kokou, F.; Makridis, P.; Kentouri, M.; Divanach, P. Antibacterial activity in microalgae cultures. Aquacult. Res. 2012, 43, 1520–1527. [Google Scholar] [CrossRef]
- Natrah, F.M.; Bossier, P.; Sorgeloos, P.; Yusoff, F.M.; Defoirdt, T. Significance of microalgal–bacterial interactions for aquaculture. Rev. Aquacult. 2014, 6, 48–61. [Google Scholar] [CrossRef]
- Tendencia, E.A.; dela Peña, M. Investigation of some components of the greenwater system which makes it effective in the initial control of luminous bacteria. Aquaculture 2003, 218, 115–119. [Google Scholar] [CrossRef]
- Biao, X.; Kaijin, Y. Shrimp farming in China: Operating characteristics, environmental impact and perspectives. Ocean Coast. Manage. 2007, 50, 538–550. [Google Scholar] [CrossRef]
- Neori, A. “Green water” microalgae: The leading sector in world aquaculture. J. Appl. Phycol. 2011, 23, 143–149. [Google Scholar] [CrossRef]
- Suárez-Puerto, B.; Delgadillo-Díaz, M.; Sánchez-Solís, M.J.; Gullian-Klanian, M. Analysis of the cost-effectiveness and growth of Nile tilapia (Oreochromis niloticus) in biofloc and green water technologies during two seasons. Aquaculture 2021, 538, 736534. [Google Scholar] [CrossRef]
- Jimenez, B.D.; Gaid, R.D.; Jimenez, C.R. Microalgal profile and anti-Vibrio activity of crude extracts from green water reservoir stocked with tilapia. Int. J. Adv. Agric. Environ. Eng. 2016, 3, 259–262. [Google Scholar]
- Chithambaran, S.; Harbi, M.; Broom, M.; Khobrani, K.; Ahmad, O.; Fattani, H.; Sofyani, A.; Ayaril, N. Green water technology for the production of Pacific white shrimp Penaeus vannamei (Boone, 1931). Indian J. Fish 2017, 64, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Tendencia, E.A.; Bosma, R.H.; Sorio, L.R. Effect of three innovative culture systems on water quality and whitespot syndrome virus (WSSV) viral load in WSSV-fed Penaeus monodon cultured in indoor tanks. Aquaculture 2012, 350, 169–174. [Google Scholar] [CrossRef]
- Palmer, P.J.; Burke, M.J.; Palmer, C.J.; Burke, J.B. Developments in controlled green-water larval culture technologies for estuarine fishes in Queensland, Australia and elsewhere. Aquaculture 2007, 272, 1–21. [Google Scholar] [CrossRef]
- Kalasariya, H.S.; Patel, R.V.; Pandya, K.Y.; Jasrai, R.; Brahmbhatt, N.H. A review on nutritional facets of seaweeds. Int. J. Chem. Sci. Technol. 2016, 1, 27–32. [Google Scholar]
- Anh, N.T.N.; Vinh, N.H.; An, B.N.T.; Lan, L.M.; Hai, T.N. Polyculture culture of black tiger shrimp Penaeus monodon and red seaweed Gracilaria tenuistipitata under different densities: Effects on water quality, post-larvae performance and their resistance against Vibrio parahaemolyticus. J. Appl. Phycol. 2020, 32, 4333–4345. [Google Scholar] [CrossRef]
- Kang, Y.H.; Park, S.R.; Chung, I.K. Biofiltration efficiency and biochemical composition of three seaweed species cultivated in a fish-seaweed integrated culture. Algae 2011, 26, 97–108. [Google Scholar]
- Yang, Y.; Chai, Z.; Wang, Q.; Chen, W.; He, Z.; Jiang, S. Cultivation of seaweed Gracilaria in Chinese coastal waters and its contribution to environmental improvements. Algal Res. 2015, 9, 236–244. [Google Scholar] [CrossRef]
- Ajjabi, L.C.; Abaab, M.; Segni, R. The red macroalga Gracilaria verrucosa in co-culture with the Mediterranean mussels Mytilus galloprovincialis: Productivity and nutrient removal performance. Aquacult. Int. 2018, 26, 253–266. [Google Scholar] [CrossRef]
- Fourooghifard, H.; Matinfar, A.; Mortazavi, M.S.; Roohani Ghadikolaee, K.; Roohani Ghadikolaee, M. Nitrogen and phosphorous budgets for integrated culture of whiteleg shrimp Litopenaeus vannamei with red seaweed Gracilaria corticata in zero water exchange system. Iran. J. Fish. Sci. 2018, 17, 471–486. [Google Scholar]
- Brito, L.O.; Arantes, R.; Magnotti, C.; Derner, R.; Pchara, F.; Olivera, A.; Vinatea, L. Water quality and growth of Pacific white shrimp Litopenaeus vannamei (Boone) in co-culture with green seaweed Ulva lactuca (Linaeus) in intensive system. Aquacult. Int. 2014, 22, 497–508. [Google Scholar] [CrossRef]
- Elizondo-González, R.; Quiroz-Guzmán, E.; Escobedo-Fregoso, C.; Magallón-Servín, P.; Peña-Rodríguez, A. Use of seaweed Ulva lactuca for water bioremediation and as feed additive for white shrimp Litopenaeus vannamei. PeerJ 2018, 6, e4459. [Google Scholar] [CrossRef] [Green Version]
- Pallaoro, M.F.; do Nascimento Vieira, F.; Hayashi, L. Ulva lactuca (Chlorophyta Ulvales) as co-feed for Pacific white shrimp. J. Appl. Phycol. 2016, 28, 3659–3665. [Google Scholar] [CrossRef]
- Beltran-Gutierrez, M.; Ferse, S.C.; Kunzmann, A.; Stead, S.M.; Msuya, F.E.; Hoffmeister, T.S.; Slater, M.J. Co-culture of sea cucumber Holothuria scabra and red seaweed Kappaphycus striatum. Aquacult. Res. 2016, 47, 1549–1559. [Google Scholar] [CrossRef]
- Chávez-Crooker, P.; Obreque-Contreras, J. Bioremediation of aquaculture wastes. Curr. Opin. Biotechnol. 2010, 21, 313–317. [Google Scholar] [CrossRef]
- Granada, L.; Sousa, N.; Lopes, S.; Lemos, M.F. Is integrated multitrophic aquaculture the solution to the sectors’ major challenges?—A review. Rev. Aquacult. 2016, 8, 283–300. [Google Scholar] [CrossRef]
- Namvar, F.; Mohamed, S.; Fard, S.G.; Behravan, J.; Mustapha, N.M.; Alitheen, N.B.M.; Othman, F. Polyphenol-rich seaweed (Eucheuma cottonii) extract suppresses breast tumour via hormone modulation and apoptosis induction. Food Chem. 2012, 130, 376–382. [Google Scholar] [CrossRef]
- García-Poza, S.; Leandro, A.; Cotas, C.; Cotas, J.; Marques, J.C.; Pereira, L.; Gonçalves, A.M. The evolution road of seaweed aquaculture: Cultivation technologies and the industry 4.0. Int. J. Env. Res. Public Health 2020, 17, 6528. [Google Scholar] [CrossRef]
- Abreu, M.H.; Pereira, R.; Yarish, C.; Buschmann, A.H.; Sousa-Pinto, I. IMTA with Gracilaria vermiculophylla: Productivity and nutrient removal performance of the seaweed in a land-based pilot scale system. Aquaculture 2011, 312, 77–87. [Google Scholar] [CrossRef]
- Tanaka, Y.; Ashaari, A.; Mohamad, F.S.; Lamit, N. Bioremediation potential of tropical seaweeds in aquaculture: Low-salinity tolerance, phosphorus content, and production of UV-absorbing compounds. Aquaculture 2020, 518, 734853. [Google Scholar] [CrossRef]
- Zheng, Y.; Jin, R.; Zhang, X.; Wang, Q.; Wu, J. The considerable environmental benefits of seaweed aquaculture in China. Stoch. Environ. Res. Risk Assess. 2019, 33, 1203–1221. [Google Scholar] [CrossRef]
- Freeman, S.; Angel, D.; Freeman, S. Integrated aquaculture (INTAQ) as a tool for an ecosystem approach to the marine farming sector in the Mediterranean Sea. Integr. Maric. 2019, 529, 133–183. [Google Scholar]
- Machado, M.; Machado, S.; Pimentel, F.B.; Freitas, V.; Alves, R.C.; Oliveira, M. Amino acid profile and protein quality assessment of macroalgae produced in an integrated multi-trophic aquaculture system. Foods 2020, 9, 1382. [Google Scholar] [CrossRef]
- Pérez-Morales, A.; Band-Schmidt, C.J.; Martínez-Díaz, S.F. Changes in mortality rates during the larval stage of the Pacific white shrimp (Litopenaeus vannamei) on the basis of algal (Chaetoceros calcitrans or Tetraselmis suecica) food density. Ecosistemas Y. Recur. Agropecu. 2016, 3, 415–420. [Google Scholar]
- Jamali, H.; Ahmadifard, N.; Abdollahi, D. Evaluation of growth, survival and body composition of larval white shrimp (Litopenaeus vannamei) fed the combination of three types of algae. Int. Aquat. Res. 2015, 7, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Navarro, N. Feeding behaviour of the rotifers Brachionus plicatilis and Brachionus rotundiformis with two types of food: Live and freeze-dried microalgae. J. Exp. Mar. Biol. Ecol. 1999, 237, 75–87. [Google Scholar] [CrossRef]
- Robert, R.; Parisi, G.; Rodolfi, L.; Poli, B.M.; Tredici, M.R. Use of fresh and preserved Tetraselmis suecica for feeding Crassostrea gigas larvae. Aquaculture 2001, 192, 333–346. [Google Scholar] [CrossRef]
- Nell, J.A.; O’Connor, W.A. The evaluation of fresh algae and stored algal concentrates as a food source for Sydney rock oyster, Saccostrea commercialis (Iredale & Roughley), larvae. Aquaculture 1991, 99, 277–284. [Google Scholar]
- Southgate, P.C.; Beer, A.C.; Duncan, P.F.; Tamburri, R. Assessment of the nutritional value of three species of tropical microalgae, dried Tetraselmis and a yeast-based diet for larvae of the blacklip pearl oyster, Pinctada margaritifera (L.). Aquaculture 1998, 162, 247–257. [Google Scholar] [CrossRef]
- Duy, N.D.Q.; Francis, D.S.; Pirozzi, I.; Southgate, P.C. Use of micro-algae concentrates for hatchery culture of sandfish, Holothuria scabra. Aquaculture 2016, 464, 145–152. [Google Scholar] [CrossRef]
- Duy, N.D.Q.; Pirozzi, I.; Southgate, P.C. Ingestion and digestion of live microalgae and microalgae concentrates by sandfish, Holothuria scabra, larvae. Aquaculture 2015, 448, 256–261. [Google Scholar] [CrossRef]
- Sales, R.; Mélo, R.C.S.; de Moraes Junior, R.M.; da Silva, R.C.S.; Cavalli, R.O.; Navarro, D.M.d.A.F.; de Souza Santos, L.P. Production and use of a flocculated paste of Nannochloropsis oculata for rearing newborn seahorse Hippocampus reidi. Algal Res. 2016, 17, 142–149. [Google Scholar] [CrossRef]
- Sarker, P.K.; Kapuscinski, A.R.; McKuin, B.; Fitzgerald, D.S.; Nash, H.M.; Greenwood, C. Microalgae-blend tilapia feed eliminates fishmeal and fish oil, improves growth, and is cost viable. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Fadl, S.E.; ElGohary, M.; Elsadany, A.Y.; Gad, D.M.; Hanaa, F.F.; El-Habashi, N.M. Contribution of microalgae-enriched fodder for the Nile tilapia to growth and resistance to infection with Aeromonas hydrophila. Algal Res. 2017, 27, 82–88. [Google Scholar] [CrossRef]
- Fadl, S.E.; Elsadany, A.Y.; El-Shenawy, A.M.; Sakr, O.A.; El Gammal, G.A.; Gad, D.M.; Norag, M.A.A.; Eissa, I. Efficacy of cyanobacterium Anabaena sp. as a feed supplement on productive performance and immune status in cultured Nile tilapia. Aquacult. Rep. 2020, 17, 100406. [Google Scholar] [CrossRef]
- Fadl, S.E.; El-Habashi, N.; Gad, D.M.; Elkassas, W.M.; Elbialy, Z.I.; Abdelhady, D.H.; Hegazi, S.M. Effect of adding Dunaliella algae to fish diet on lead acetate toxicity and gene expression in the liver of Nile tilapia. Toxin Rev. 2021, 40, 1155–1171. [Google Scholar] [CrossRef]
- Younis, E.-S.M.; Al-Quffail, A.S.; Al-Asgah, N.A.; Abdel-Warith, A.-W.A.; Al-Hafedh, Y.S. Effect of dietary fish meal replacement by red algae, Gracilaria arcuata, on growth performance and body composition of Nile tilapia Oreochromis niloticus. Saudi J. Biol. Sci. 2018, 25, 198–203. [Google Scholar] [CrossRef]
- Mansour, A.T.; Alsaqufi, A.S.; Alkhamis, Y.A.; Al-Gazar, F.F.; Zaki, M.A.; Nour, A.A.M.; Ramada, K.M.A. The evaluation of Arthrospira platensis bioactivity and their dietary supplementation to Nile tilapia vegetarian diet on growth performance, feed utilization, body composition and hemato-biochemical parameters Ann. Anim. Sci. 2021, 21, 1061–1080. [Google Scholar] [CrossRef]
- Pascon, G.; Messina, M.; Petit, L.; Valente, L.M.P.; Oliveira, B.; Przybyla, C.; Dutto, G.; Tulli, F. Potential application and beneficial effects of a marine microalgal biomass produced in a high-rate algal pond (HRAP) in diets of European sea bass, Dicentrarchus labrax. Environ. Sci. Pollut. Res. 2021, 28, 62185–62199. [Google Scholar] [CrossRef] [PubMed]
- Haas, S.; Bauer, J.; Adakli, A.; Meyer, S.; Lippemeier, S.; Schwarz, K.; Schulz, C. Marine microalgae Pavlova viridis and Nannochloropsis sp. as n-3 PUFA source in diets for juvenile European sea bass (Dicentrarchus labrax L.). J. Appl. Phycol. 2016, 28, 1011–1021. [Google Scholar] [CrossRef]
- Valente, L.M.P.; Custódio, M.; Batista, S.; Fernandes, H.; Kiron, V. Defatted microalgae (Nannochloropsis sp.) from biorefinery as a potential feed protein source to replace fishmeal in European sea bass diets. Fish Physiol. Biochem. 2019, 45, 1067–1081. [Google Scholar] [CrossRef]
- Batista, S.; Pereira, R.; Oliveira, B.; Baião, L.F.; Jessen, F.; Tulli, F.; Messina, M.; Silva, J.L.; Abreu, H.; Valente, L.M. Exploring the potential of seaweed Gracilaria gracilis and microalga Nannochloropsis oceanica, single or blended, as natural dietary ingredients for European seabass Dicentrarchus labrax. J. Appl. Phycol. 2020, 32, 2041–2059. [Google Scholar] [CrossRef]
- Reyes-Becerril, M.; Guardiola, F.; Rojas, M.; Ascencio-Valle, F.; Esteban, M.Á. Dietary administration of microalgae Navicula sp. affects immune status and gene expression of gilthead seabream (Sparus aurata). Fish Shellfish Immunol. 2013, 35, 883–889. [Google Scholar] [CrossRef]
- Cerezuela, R.; Guardiola, F.A.; González, P.; Meseguer, J.; Esteban, M.Á. Effects of dietary Bacillus subtilis, Tetraselmis chuii, and Phaeodactylum tricornutum, singularly or in combination, on the immune response and disease resistance of sea bream (Sparus aurata L.). Fish Shellfish Immunol. 2012, 33, 342–349. [Google Scholar] [CrossRef]
- Pereira, H.; Sardinha, M.; Santos, T.; Gouveia, L.; Barreira, L.; Dias, J.; Varela, J. Incorporation of defatted microalgal biomass (Tetraselmis sp. CTP4) at the expense of soybean meal as a feed ingredient for juvenile gilthead seabream (Sparus aurata). Algal Res. 2020, 47, 101869. [Google Scholar] [CrossRef]
- Seong, T.; Matsutani, H.; Haga, Y.; Kitagima, R.; Satoh, S. First step of non-fish meal, non-fish oil diet development for red seabream,(Pagrus major), with plant protein sources and microalgae Schizochytrium sp. Aquacult. Res. 2019, 50, 2460–2468. [Google Scholar] [CrossRef]
- Galafat, A.; Vizcaíno, A.J.; Sáez, M.I.; Martínez, T.F.; Arizcun, M.; Chaves-Pozo, E.; Alarcón, F.J. Assessment of dietary inclusion of crude or hydrolysed Arthrospira platensis biomass in starter diets for gilthead seabream (Sparus aurata). Aquaculture 2022, 548, 737680. [Google Scholar] [CrossRef]
- Magnoni, L.J.; Martos-Sitcha, J.A.; Queiroz, A.; Calduch-Giner, J.A.; Gonçalves, J.F.M.; Rocha, C.M.; Abreu, H.T.; Schrama, J.W.; Ozorio, R.O.; Pérez-Sánchez, J. Dietary supplementation of heat-treated Gracilaria and Ulva seaweeds enhanced acute hypoxia tolerance in gilthead sea bream (Sparus aurata). Biol. Open 2017, 6, 897–908. [Google Scholar]
- Gong, Y.; Bandara, T.; Huntley, M.; Johnson, Z.I.; Dias, J.; Dahle, D.; Sørensen, M.; Kiron, V. Microalgae Scenedesmus sp. as a potential ingredient in low fishmeal diets for Atlantic salmon (Salmo salar L.). Aquaculture 2019, 501, 455–464. [Google Scholar] [CrossRef]
- Gong, Y.; Sørensen, S.L.; Dahle, D.; Nadanasabesan, N.; Dias, J.; Valente, L.M.; Sørensen, M.; Kiron, V. Approaches to improve utilization of Nannochloropsis oceanica in plant-based feeds for Atlantic salmon. Aquaculture 2020, 522, 735122. [Google Scholar] [CrossRef]
- Wei, M.; Parrish, C.C.; Guerra, N.I.; Armenta, R.E.; Colombo, S.M. Extracted microbial oil from a novel Schizochytrium sp.(T18) as a sustainable high DHA source for Atlantic salmon feed: Impacts on growth and tissue lipids. Aquaculture 2021, 534, 736249. [Google Scholar] [CrossRef]
- Sarker, P.K.; Kapuscinski, A.R.; Vandenberg, G.W.; Proulx, E.; Sitek, A.J.; Thomsen, L. Towards sustainable and ocean-friendly aquafeeds: Evaluating a fish-free feed for rainbow trout (Oncorhynchus mykiss) using three marine microalgae species. Elem. Sci. Anthr. 2020, 8, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Tomás-Almenar, C.; Larrán, A.; de Mercado, E.; Sanz-Calvo, M.; Hernández, D.; Riaño, B.; García-González, M. Scenedesmus almeriensis from an integrated system waste-nutrient, as sustainable protein source for feed to rainbow trout (Oncorhynchus mykiss). Aquaculture 2018, 497, 422–430. [Google Scholar] [CrossRef]
- Yıldırım, Ö.; Ergün, S.; Yaman, S.; Türker, A. Effects of two seaweeds (Ulva lactuca and Enteromorpha linza) as a feed additive in diets on growth performance, feed utilization, and body composition of rainbow trout (Oncorhynchus mykiss). Kafkas Univ. Vet. Fak. Derg. 2009, 15, 455–460. [Google Scholar]
- Allen, K.M.; Habte-Tsion, H.-M.; Thompson, K.R.; Filer, K.; Tidwell, J.H.; Kumar, V. Freshwater microalgae (Schizochytrium sp.) as a substitute to fish oil for shrimp feed. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Guimarães, A.M.; Dias Schleder, D.; Nagata, M.; Nóbrega, R.O.; Fracalossi, D.M.; Quadros Seiffert, W.; do Nascimento Vieira, F. Aurantiochytrium sp. meal can replace fish oil in practical diets for the juvenile Pacific white shrimp. Aquacult. Nutr. 2019, 25, 798–807. [Google Scholar] [CrossRef]
- Rodríguez-González, H.; Orduña-Rojas, J.; Villalobos-Medina, J.P.; García-Ulloa, M.; Polanco-Torres, A.; López-Álvarez, E.S.; Montoya-Mejía, M.; Hernández-Llamas, A. Partial inclusion of Ulva lactuca and Gracilaria parvispora meal in balanced diets for white leg shrimp (Litopenaeus vannamei). J. Appl. Phycol. 2014, 26, 2453–2459. [Google Scholar] [CrossRef]
- Seraspe, E.B.; Gabotero, S.; Milagros, R.; Pahila, I.G.; Amar, E.C. Evaluation of dietary freeze-dried Chaetoceros calcitrans supplementation to control Vibrio harveyi infection on Penaeus monodon juvenile. Aquaculture 2014, 432, 212–216. [Google Scholar] [CrossRef]
- Ju, Z.Y.; Deng, D.-F.; Dominy, W. A defatted microalgae (Haematococcus pluvialis) meal as a protein ingredient to partially replace fishmeal in diets of Pacific white shrimp (Litopenaeus vannamei, Boone, 1931). Aquaculture 2012, 354, 50–55. [Google Scholar] [CrossRef]
- Radhakrishnan, S.; Belal, I.E.; Seenivasan, C.; Muralisankar, T.; Bhavan, P.S. Impact of fishmeal replacement with Arthrospira platensis on growth performance, body composition and digestive enzyme activities of the freshwater prawn, Macrobrachium rosenbergii. Aquacult. Rep. 2016, 3, 35–44. [Google Scholar] [CrossRef] [Green Version]
- Maliwat, G.C.F.; Velasquez, S.F.; Buluran, S.M.D.; Tayamen, M.M.; Ragaza, J.A. Growth and immune response of pond-reared giant freshwater prawn Macrobrachium rosenbergii post larvae fed diets containing Chlorella vulgaris. Aquac. Fish. 2021, 6, 465–470. [Google Scholar] [CrossRef]
- Cañavate, J.P.; Fernández-Díaz, C. Pilot evaluation of freeze-dried microalgae in the mass rearing of gilthead seabream (Sparus aurata) larvae. Aquaculture 2001, 193, 257–269. [Google Scholar] [CrossRef]
- Shpigel, M.; Shauli, L.; Odintsov, V.; Ben-Ezra, D.; Neori, A.; Guttman, L. The sea urchin, Paracentrotus lividus, in an Integrated Multi-Trophic Aquaculture (IMTA) system with fish (Sparus aurata) and seaweed (Ulva lactuca): Nitrogen partitioning and proportional configurations. Aquaculture 2018, 490, 260–269. [Google Scholar] [CrossRef]
- Nardelli, A.E.; Chiozzini, V.G.; Braga, E.S.; Chow, F. Integrated multi-trophic farming system between the green seaweed Ulva lactuca, mussel, and fish: A production and bioremediation solution. J. Appl. Phycol. 2019, 31, 847–856. [Google Scholar] [CrossRef]
- Chow, F.; Macchiavello, J.; Cruz, S.S.; Fonck, E.; Olivares, J. Utilization of Gracilaria chilensis (Rhodophyta: Gracilariaceae) as a biofilter in the depuration of effluents from tank cultures of fish, oysters, and sea urchins. J. World Aquacult. Soc. 2001, 32, 215–220. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mansour, A.T.; Ashour, M.; Alprol, A.E.; Alsaqufi, A.S. Aquatic Plants and Aquatic Animals in the Context of Sustainability: Cultivation Techniques, Integration, and Blue Revolution. Sustainability 2022, 14, 3257. https://0-doi-org.brum.beds.ac.uk/10.3390/su14063257
Mansour AT, Ashour M, Alprol AE, Alsaqufi AS. Aquatic Plants and Aquatic Animals in the Context of Sustainability: Cultivation Techniques, Integration, and Blue Revolution. Sustainability. 2022; 14(6):3257. https://0-doi-org.brum.beds.ac.uk/10.3390/su14063257
Chicago/Turabian StyleMansour, Abdallah Tageldein, Mohamed Ashour, Ahmed E. Alprol, and Ahmed Saud Alsaqufi. 2022. "Aquatic Plants and Aquatic Animals in the Context of Sustainability: Cultivation Techniques, Integration, and Blue Revolution" Sustainability 14, no. 6: 3257. https://0-doi-org.brum.beds.ac.uk/10.3390/su14063257