
Impacts of Biochar and Vermicompost Addition on Physicochemical Characteristics, Metal Availability, and Microbial Communities in Soil Contaminated with Potentially Toxic Elements
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil, Biochar, and Vermicompost Preparation
2.2. Samples Collection and Parameters Determination
2.3. Quantitative PCR
2.4. High-Throughput Sequencing
2.5. Data Analysis
3. Results and Discussion
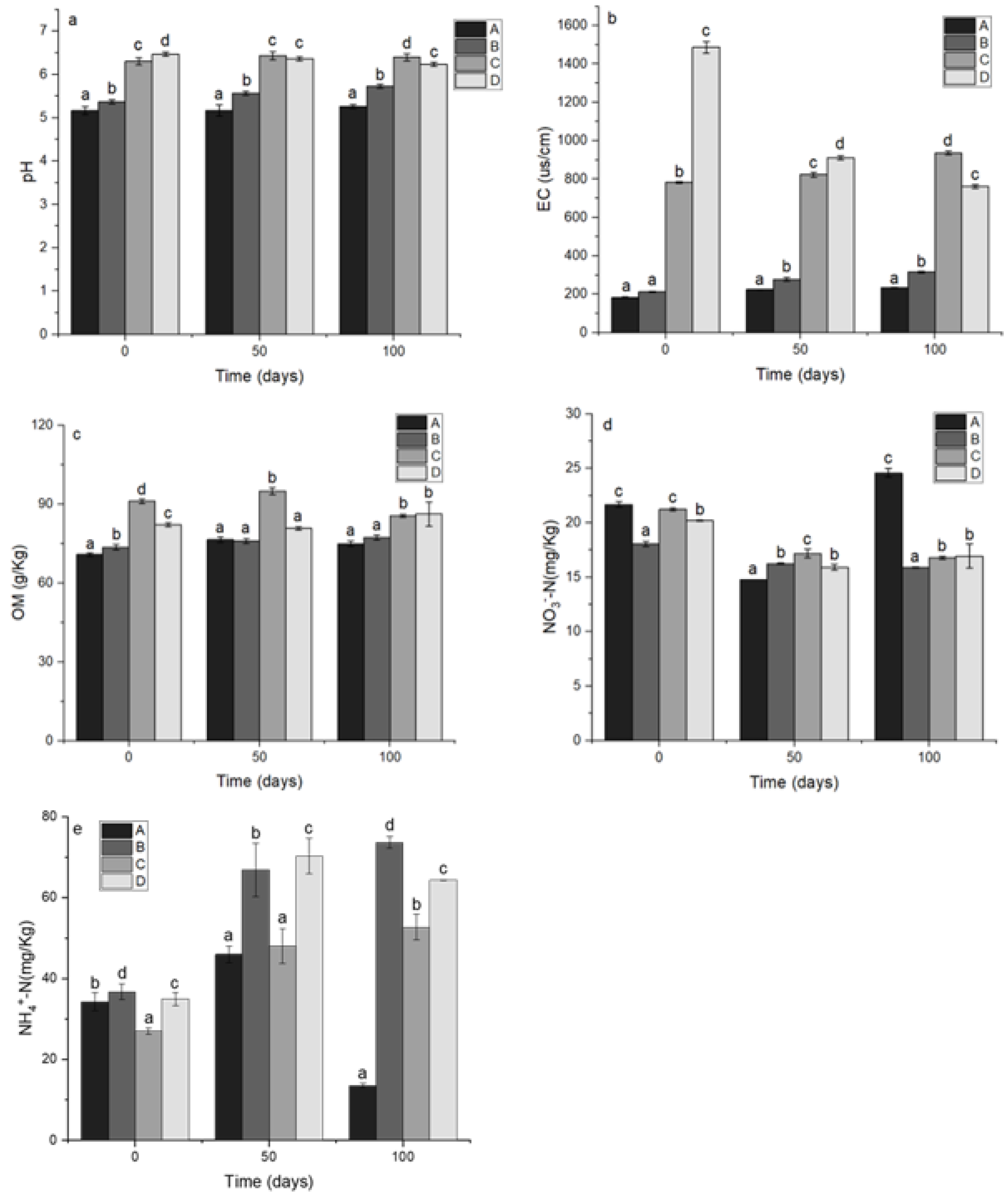
3.1. Physicochemical Parameters
3.2. Impacts on the Form of CaCl2-Extractable PTEs
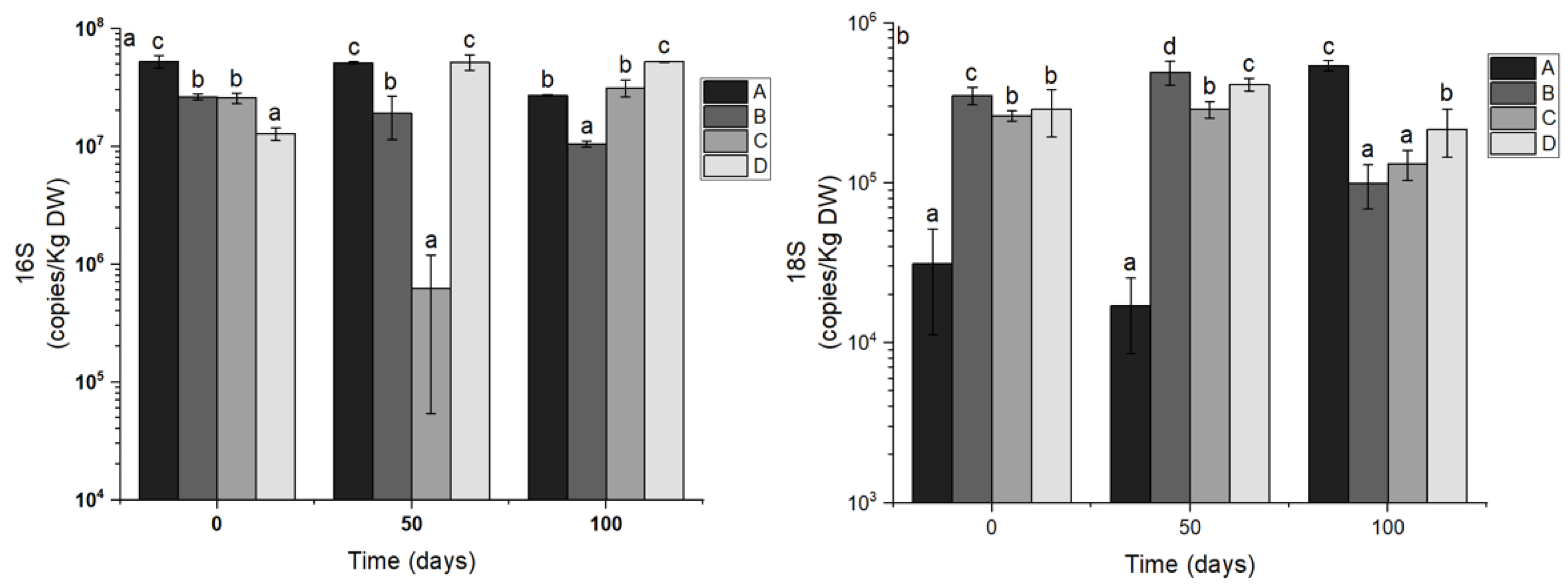
3.3. Abundance of 16S rRNA and 18S rRNA
3.4. AOA and AOB Gene Abundance and Structure
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yuan, X.; Xue, N.; Han, Z. A meta-analysis of heavy metals pollution in farmland and urban soils in China over the past 20 years-ScienceDirect. J. Environ. Sci. 2021, 101, 217–226. [Google Scholar] [CrossRef]
- Hou, D.; Ding, Z.; Li, G.; Wu, L.; Hu, P.; Guo, G.; Wang, X.; Ma, Y.; O’Connor, D.; Wang, X. A sustainability assessment framework for agricultural land remediation in China. Land Degrad. Dev. 2018, 29, 1005–1018. [Google Scholar] [CrossRef]
- Bolan, N.; Kunhikrishnan, A.; Thangarajan, R.; Kumpiene, J.; Park, J.; Makino, T.; Kirkham, M.B.; Scheckel, K. Remediation of heavy metal(loid)s contaminated soils—To mobilize or to immobilize. J. Hazard. Mater. 2014, 266, 141–166. [Google Scholar] [CrossRef] [PubMed]
- Kou, S.; Gilles, V.; Emmanuel, G.; Pitre, F.E.; Michel, L.; Brereton, N. The Response of a 16S Ribosomal RNA Gene Fragment Amplified Community to Lead, Zinc, and Copper Pollution in a Shanghai Field Trial. Front. Microbiol. 2018, 9, 366–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, X.; Wang, P.; Zweiten, L.V.; Bolan, N.; Wang, H.; Li, X.; Cheng, K.; Yang, Y.; Wang, M.; Liu, T.; et al. Towards a better understanding of the role of Fe cycling in soil for carbon stabilization and degradation. Carbon Res. 2022, 1, 5. [Google Scholar] [CrossRef]
- Hua, S.; Gong, J.L.; Zeng, G.M.; Yao, F.B.; Guo, M.; Ou, X.M. Remediation of organochlorine pesticides contaminated lake sediment using activated carbon and carbon nanotubes. Chemosphere 2017, 177, 65–76. [Google Scholar] [CrossRef]
- Zhai, X.; Li, Z.; Huang, B.; Luo, N.; Huang, M.; Zhang, Q.; Zeng, G. Remediation of multiple heavy metal-contaminated soil through the combination of soil washing and in situ immobilization. Sci. Total Environ. 2018, 635, 92–99. [Google Scholar] [CrossRef]
- Raklami, A.; Tahiri, A.i.; Bechtaoui, N.; Abdelhay, E.G.; Pajuelo, E.; Baslam, M.; Meddich, A.; Oufdou, K. Restoring the plant productivity of heavy metal-contaminated soil using phosphate sludge, marble waste, and beneficial microorganisms. J. Environ. Sci. 2021, 99, 210–221. [Google Scholar] [CrossRef] [PubMed]
- Kuppusamy, S.; Palanisami, T.; Megharaj, M.; Venkateswarlu, K.; Naidu, R. Ex-situ remediation technologies for environmental pollutants: A critical perspective. Rev. Environ. Contam. Toxi. 2016, 236, 117–192. [Google Scholar]
- Thakur, S.; Singh, L.; Wahid, Z.A.; Siddiqui, M.F.; Atnaw, S.M.; Din, M.F.M. Plant-driven removal of heavy metals from soil: Uptake, translocation, tolerance mechanism, challenges, and future perspectives. Environ. Monit. Assess. 2016, 188, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yang, X.; Yuan, X.; Tian, S.; Wang, X.; Zhang, H.; Han, L. Effect of pyrolysis temperature on composition, carbon fraction and abiotic stability of straw biochars: Correlation and quantitative analysis. Carbon Res. 2022, 1, 17. [Google Scholar] [CrossRef]
- Ding, Y.; Liu, Y.; Liu, S.; Li, Z.; Tan, X.; Huang, X.; Zeng, G.; Zhou, L.; Zheng, B. Biochar to improve soil fertility. A review. Agron. Sustain. Deve. 2016, 36, 36. [Google Scholar] [CrossRef] [Green Version]
- Kammann, C.; Ippolito, J.; Hagemann, N.; Borchard, N.; Cayuela, M.L.; Estavillo, J.M.; Fuertes-Mendizabal, T.; Jeffery, S.; Kern, J.; Novak, J.; et al. Biochar as a tool to reduce the agricultural greenhouse-gas burden–knowns, unknowns and future research needs. J. Environ. Eng. Landsc. Manag. 2017, 25, 114–139. [Google Scholar] [CrossRef] [Green Version]
- Beesley, L.; Moreno-Jiménez, E.; Gomez-Eyles, J.L.; Harris, E.; Robinson, B.; Sizmur, T. A review of biochars’ potential role in the remediation, revegetation and restoration of contaminated soils. Environ. Pollut. 2011, 159, 3269–3282. [Google Scholar] [CrossRef]
- Zhang, W.; Qiu, X.; Wang, C.; Zhong, L.; Fu, F.; Zhu, J.; Zhang, Z.; Qin, Y.; Yang, D.; Xu, C.C. Lignin derived carbon materials: Current status and future trends. Carbon Res. 2022, 1, 14. [Google Scholar] [CrossRef]
- Garg, P.; Gupta, A.; Satya, S. Vermicomposting of different types of waste using Eisenia foetida: A comparative study. Bioresour. Technol. 2006, 97, 391–395. [Google Scholar] [CrossRef]
- Edwards, C.; Burrows, I. The potential of earthworms composts as plant growth media. In Earthworms in Waste and Environmental Management; Edward, C.A., Neuhauser, E.F., Eds.; SPB Academic Publishing: The Hague, The Netherlands, 1988; pp. 211–220. [Google Scholar]
- Ndegwa, P.; Thompson, S. Integrating composting and vermicomposting in the treatment and bioconversion of biosolids. Bioresour. Technol. 2001, 76, 107–112. [Google Scholar] [CrossRef]
- Pereira, M.G.; Arruda, M.A. Vermicompost as a natural adsorbent material: Characterization and potentialities for cadmium adsorption. J. Brazil. Chem. Soc. 2003, 14, 39–47. [Google Scholar] [CrossRef] [Green Version]
- Vetrova, O.V.; Konovalov, K.B.; Gavrilenko, M.A. Application of Humic Sorbents for Pb2+, Cu2+ and Hg2+ ions preconcentration from aqueous solutions. Procedia Chem. 2014, 10, 120–126. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-Miceli, F.A.; Santiago-Borraz, J.; Molina, J.A.M.; Nafate, C.C.; Abud-Archila, M.; Llaven, M.A.O.; Rincón-Rosales, R.; Dendooven, L. Vermicompost as a soil supplement to improve growth, yield and fruit quality of tomato (Lycopersicum esculentum). Bioresour. Technol. 2007, 98, 2781–2786. [Google Scholar] [CrossRef]
- Gil-Sotres, F.; Trasar-Cepeda, C.; Leirós, M.C.; Seoane, S. Different approaches to evaluating soil quality using biochemical properties. Soil Biol. Biochem. 2004, 37, 877–887. [Google Scholar] [CrossRef]
- Lagomarsino, A.; Moscatelli, M.C.; Tizio, A.D.; Mancinelli, R.; Grego, S.; Marinari, S. Soil biochemical indicators as a tool to assess the short-term impact of agricultural management on changes in organic C in a Mediterranean environment. Ecol. Indic. 2008, 9, 518–527. [Google Scholar] [CrossRef]
- Zhang, C.; Mora, P.; Dai, J.; Chen, X.; Giusti-Miller, S.; Ruiz-Camacho, N.; Velasquez, E.; Lavelle, P. Earthworm and organic amendment effects on microbial activities and metal availability in a contaminated soil from China. Appl. Soil Ecol. 2016, 104, 54–66. [Google Scholar] [CrossRef]
- Lu, H.; Yan, M.; Wong, M.H.; Mo, W.Y.; Wang, Y.; Chen, X.W.; Wang, J.J. Effects of biochar on soil microbial community and functional genes of a landfill cover three years after ecological restoration. Sci. Total Environ. 2020, 717, 137133. [Google Scholar] [CrossRef] [PubMed]
- Zeng, G.; Zhang, J.; Chen, Y.; Yu, Z.; Yu, M.; Li, H.; Liu, Z.; Chen, M.; Lu, L.; Hu, C. Relative contributions of archaea and bacteria to microbial ammonia oxidation differ under different conditions during agricultural waste composting. Bioresour. Technol. 2011, 102, 9026–9032. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Cai, C.; Zhang, J.; Yang, Y.; Wu, G.; Luo, L.; Huang, H.; Zhou, Y.; Qin, P.; Yu, M. Key environmental factors to variation of ammonia-oxidizing archaea community and potential ammonia oxidation rate during agricultural waste composting. Bioresour. Technol. 2018, 270, 278–285. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Sun, H.; Min, L.; Ren, C. Biochars change the sorption and degradation of thiacloprid in soil: Insights into chemical and biological mechanisms. Environ. Pollut. 2018, 236, 158–167. [Google Scholar] [CrossRef]
- Yuan, J.H.; Xu, R.K.; Hong, Z. The forms of alkalis in the biochar produced from crop residues at different temperatures. Bioresour. Technol. 2011, 102, 3488–3497. [Google Scholar] [CrossRef]
- Zheng, R.; Chen, Z.; Cai, C.; Tie, B.; Liu, X.; Reid, B.J.; Huang, Q.; Lei, M.; Sun, G.; Baltrėnaitė, E. Mitigating heavy metal accumulation into rice (Oryza sativa L.) using biochar amendment—A field experiment in Hunan, China. Environ. Sci. Pollut. Res. 2015, 22, 11097–11108. [Google Scholar] [CrossRef] [PubMed]
- Glaser, B.; Wiedner, K.; Seelig, S.; Schmidt, H.P.; Gerber, H. Biochar organic fertilizers from natural resources as substitute for mineral fertilizers. Agron. Sustain. Deve. 2015, 35, 667–678. [Google Scholar] [CrossRef]
- Arif, M.S.; Riaz, M.; Shahzad, S.M.; Yasmeen, T.; Ashraf, M.; Siddique, M.; Mubarik, M.S.; Bragazza, L.; Buttler, A. Fresh and composted industrial sludge restore soil functions in surface soil of degraded agricultural land. Sci. Total Environ. 2018, 619, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Clark, I.; Sánchez-Monedero, M.; Shea, S.; Meier, S.; Bolan, N. Maturity indices in co-composting of chicken manure and sawdust with biochar. Bioresour. Technol. 2014, 168, 245–251. [Google Scholar] [CrossRef]
- Malińska, K.; Zabochnicka-WiTek, M.; Dach, J. Effects of biochar amendment on ammonia emission during composting of sewage sludge. Ecol. Eng. 2014, 71, 474–478. [Google Scholar] [CrossRef]
- Zhang, L.; Dong, H.; Zhang, J.; Chen, Y.; Zeng, G.; Yuan, Y.; Cao, W.; Fang, W.; Hou, K.; Wang, B. Influence of FeONPs amendment on nitrogen conservation and microbial community succession during composting of agricultural waste: Relative contributions of ammonia-oxidizing bacteria and archaea to nitrogen conservation. Bioresour. Technol. 2019, 287, 121463. [Google Scholar] [CrossRef]
- Sundaray, S.K.; Nayak, B.B.; Lin, S.; Bhatta, D. Geochemical speciation and risk assessment of heavy metals in the river estuarine sediments—A case study: Mahanadi basin, India. J. Hazard. Mater. 2011, 186, 1837–1846. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.; Yang, X.; Gielen, G.; Bolan, N.; Wang, H. Effect of bamboo and rice straw biochars on the mobility and redistribution of heavy metals (Cd, Cu, Pb and Zn) in contaminated soil. J. Environ. Manag. 2017, 186, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Yang, Z.; Tang, L.; Zeng, G.; Yu, M.; Li, X.; Wu, H.; Qian, Y.; Li, X.; Luo, Y. Changes in heavy metal mobility and availability from contaminated wetland soil remediated with combined biochar-compost. Chemosphere 2017, 181, 281–288. [Google Scholar] [CrossRef]
- Li, D.; Ma, J.; Xu, H.; Xu, X.; Qiu, H.; Cao, X.; Zhao, L. Recycling waste nickel-laden biochar to pseudo-capacitive material by hydrothermal treatment: Roles of nickel-carbon interaction. Carbon Res. 2022, 1, 16. [Google Scholar] [CrossRef]
- Sizmur, T.; Hodson, M.E. Do earthworms impact metal mobility and availability in soil?—A review. Environ. Pollut. 2009, 157, 1981–1989. [Google Scholar] [CrossRef] [Green Version]
- Shan, J.; Wang, Y.F.; Wang, L.H.; Yan, X.Y.; Ji, R. Effects of the geophagous earthworm Metaphire guillelmi on sorption, mineralization, and bound-residue formation of 4-nonylphenol in an agricultural soil. Environ. Pollut. 2014, 189, 202–207. [Google Scholar] [CrossRef]
- Senesi, N. Binding mechanisms of pesticides to soil humic substances. Sci. Total Environ. 1992, 123, 63–76. [Google Scholar] [CrossRef]
- Liang, J.; Tang, S.; Gong, J.; Zeng, G.; Luo, Y. Responses of enzymatic activity and microbial communities to biochar/compost amendment in sulfamethoxazole polluted wetland soil. J. Hazard. Mater. 2019, 385, 121533. [Google Scholar] [CrossRef] [PubMed]
- Xiao, R.; Awasthi, M.K.; Li, R.; Park, J.; Pensky, S.M.; Wang, Q.; Wang, J.J.; Zhang, Z. Recent developments in biochar utilization as an additive in organic solid waste composting: A review. Bioresour. Technol. 2017, 246, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Wang, S.; Tian, L.; Li, S.; Li, X.; Shen, Y.; Tian, C. Long-term biochar application influences soil microbial community and its potential roles in semiarid farmland. Appl. Soil Ecol. 2017, 117, 10–15. [Google Scholar] [CrossRef]
- Yang, X.; Tsibart, A.; Nam, H.; Hur, J.; El-Naggar, A.; Tack, F.M.; Wang, C.H.; Lee, Y.H.; Tsang, D.C.; Ok, Y.S. Effect of gasification biochar application on soil quality: Trace metal behavior, microbial community, and soil dissolved organic matter. J. Hazard. Mater. 2019, 365, 684–694. [Google Scholar] [CrossRef]
- Novak, J.M.; Busscher, W.J.; Laird, D.L.; Ahmedna, M.; Watts, D.W.; Niandou, M.A. Impact of biochar amendment on fertility of a southeastern coastal plain soil. Soil Sci. 2009, 174, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Haynes, R.J.; Fraser, P.M.; Williams, P.H. Earthworm population size and composition, and microbial biomass: Effect—of pastoral and arable management in Canterbury, New Zealand. In The Significance and Regulation of Soil Biodiversity; Collins, H.P., Robertson, G.P., Klug, M.J., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1995; pp. 279–285. [Google Scholar]
- Merino-Trigo, A.; Sampedro, L.; Rodríguez-Berrocal, F.J.; Mato, S.; Cadena, M. Activity and partial characterisation of xylanolytic enzymes in the earthworm Eisenia andrei fed on organic wastes. Soil Biol. Biochem. 1999, 31, 1735–1740. [Google Scholar] [CrossRef]
- Zhang, H.; Li, J.; Zhang, Y.; Huang, K. Quality of vermicompost and microbial community diversity affected by the contrasting temperature during vermicomposting of dewatered sludge. Int. J. Environ. Res. Public Health 2020, 17, 1748. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Wang, H.; Fu, X.; Huang, K. Effect of vermicomposting using Eisenia foetida on properties of sewage sludge. Chinese J. Environ. Eng. 2010, 4, 1421–1425. [Google Scholar]
- Harter, J.; Krause, M.H.; Schuettler, S.; Ruser, R.; Fromme, M.; Scholten, T.; Kappler, A.; Behrens, S. Linking N2O emissions from biochar-amended soil to the structure and function of the N-cycling microbial community. ISEM J. 2014, 8, 660–674. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.Y.; Bai, S.H.; Tang, L.; Zhang, Y.L.; Teng, Y.; Xu, Z.H. Linking potential nitrification rates, nitrogen cycling genes and soil properties after remediating the agricultural soil contaminated with heavy metal and fungicide. Chemosphere 2017, 184, 892. [Google Scholar] [CrossRef]
- Ding, L.J.; An, X.L.; Li, S.; Zhang, G.L.; Zhu, Y.G. Nitrogen loss through anaerobic ammonium oxidation coupled to iron reduction from paddy soils in a chronosequence. Environ. Sci. Technol. 2014, 48, 10641–10647. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhang, J.; Yang, X.; Zhou, Y.; Zhang, L.; Yang, Y.; Luo, L.; Yan, Q. Responses of ammonia-oxidizing microorganisms to biochar and compost amendments of heavy metals-polluted soil. J. Environ. Sci. 2021, 102, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Enwall, K.; Nyberg, K.; Bertilsson, S.; Cederlund, H.; Stenstr, J.; Hallin, S. Long-term impact of fertilization on activity and composition of bacterial communities and metabolic guilds in agricultural soil. Soil Biol. Biochem. 2007, 39, 106–115. [Google Scholar] [CrossRef]
- Stephen, J.R.; Mccaig, A.E.; Smith, Z.; Prosser, J.I.; Embley, T.M. Molecular diversity of soil and marine 16S rRNA gene sequences related to beta-subgroup ammonia-oxidizing bacteria. Appl. Environ. Microbiol. 1996, 62, 4147–4154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, C.J.; Harris, D.; Dollhopf, S.L.; Gross, K.L.; Prosser, J.I.; Paul, E.A. Effects of Agronomic Treatments on Structure and Function of Ammonia-Oxidizing Communities. Appl. Environ. Microbiol. 2001, 66, 5410–5418. [Google Scholar] [CrossRef] [Green Version]
- Mendum, A.T.; Hirsch, P.R. Changes in the population structure of b-group autotrophic ammonia oxidising bacteria in arable soils in response to agricultural practice. Soil Biol. Biochem. 2002, 34, 1479–1485. [Google Scholar] [CrossRef]
- Lu, H.; Xu, C.; Zhang, J.; Du, C.; Wu, G.; Luo, L. The characteristics of alkaline phosphatase activity and phoD gene community in heavy-metal contaminated soil remediated by biochar and compost. Bull. Environ. Contam. Toxicol. 2022, 109, 298–303. [Google Scholar] [CrossRef]
- Zhao, K.; Wang, N.; Jiang, S.; Li, F.; Luo, S.; Chen, A.; Li, H.; Lin, X.; Zhang, J.; Zhang, L.; et al. Potential implications of biochar and compost on the stoichiometry-based assessments of soil enzyme activity in heavy metal-polluted soils. Carbon Res. 2022. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Properties | Soil | Biochar | Vermicompost |
|---|---|---|---|
| EC (ds·m–1) | 0.18 ± 0.01 | 0.16 ± 0.01 | 0.13 ± 0.01 |
| pH (H2O) | 5.16 ± 0.14 | 9.10 ± 0.02 | 8.10 ± 0.02 |
| OM (g·kg–1) | 70.8 ± 0.8 | 816.20 ± 4.23 | 316.60 ± 5.63 |
| NH4+-N (mg·kg–1) | 34.32 ± 2.2 | 42.20 ± 1.39 | 64.20 ± 2.23 |
| NO3–-N (mg·kg–1) | 21.67 ± 0.23 | 6.98 ± 1.59 | 26.48 ± 1.27 |
| Total As (mg·kg–1) | 72.4 ± 2.01 | 4.25 ± 0.16 | 44.25 ± 0.25 |
| Total Cd (mg·kg–1) | 0.51 ± 0.01 | 0.15 ± 0.01 | 0.23± 0.02 |
| Total Cu (mg·kg–1) | 52.6 ± 2.29 | 301.62 ± 0.89 | 418.62 ± 3.32 |
| Total Zn (mg·kg–1) | 144.9 ± 3.13 | 483.47 ± 13.75 | 283.47 ± 5.37 |
| Ash content (%) | - | 49.52 ± 1.23% | - |
| Moisture (%) | 18.65 | 7.12 | 22.43 |
| Treatment | Soil | Biochar | Vermicompost |
|---|---|---|---|
| A | 2.0 kg | - | - |
| B | 2.0 kg | - | 2%, 40 g |
| C | 2.0 kg | 2%, 40 g | - |
| D | 2.0 kg | 2%, 40 g | 2%, 40 g |
| pH | EC | OM | NO3−-N | NH4+-N | As | Cu | Zn | Cd | 16S | 18S | AOB | AOA | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| pH | 1 | ||||||||||||
| EC | 0.904 ** | 1 | |||||||||||
| OM | 0.836 ** | 0.653 * | 1 | ||||||||||
| NO3−-N | –0.187 | –0.032 | –0.133 | 1 | |||||||||
| NH4+-N | 0.276 | 0.067 | 0.080 | –0.849 ** | 1 | ||||||||
| As | –0.222 | –0.365 | –0.235 | –0.066 | 0.218 | 1 | |||||||
| Cu | –0.548 | –.385 | –0.497 | 0.366 | –0.412 | –0.055 | 1 | ||||||
| Zn | –0.958 ** | –0.871 ** | –0.838 ** | 0.099 | –0.134 | 0.181 | 0.618 * | 1 | |||||
| Cd | –0.948 ** | –0.836 ** | –0.867 ** | 0.137 | –0.178 | 0.276 | 0.641 * | 0.975 ** | 1 | ||||
| 16S | –0.281 | –0.213 | –0.345 | –0.074 | 0.067 | 0.158 | –0.046 | 0.140 | 0.074 | 1 | |||
| 18S | 0.092 | 0.063 | 0.006 | 0.297 | –0.147 | –0.466 | 0.066 | –0.050 | –0.095 | –0.293 | 1 | ||
| AOB | 0.486 | 0.417 | 0.210 | –0.247 | 0.449 | –0.013 | –0.568 | –0.553 | –0.586 * | 0.533 | 0.158 | 1 | |
| AOA | 0.561 | 0.681 * | 0.088 | –0.207 | 0.369 | –0.033 | –0.284 | –0.486 | –0.399 | –0.101 | 0.030 | 0.488 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Z.; He, W. Impacts of Biochar and Vermicompost Addition on Physicochemical Characteristics, Metal Availability, and Microbial Communities in Soil Contaminated with Potentially Toxic Elements. Sustainability 2023, 15, 790. https://0-doi-org.brum.beds.ac.uk/10.3390/su15010790
Huang Z, He W. Impacts of Biochar and Vermicompost Addition on Physicochemical Characteristics, Metal Availability, and Microbial Communities in Soil Contaminated with Potentially Toxic Elements. Sustainability. 2023; 15(1):790. https://0-doi-org.brum.beds.ac.uk/10.3390/su15010790
Chicago/Turabian StyleHuang, Zhiyue, and Wenjuan He. 2023. "Impacts of Biochar and Vermicompost Addition on Physicochemical Characteristics, Metal Availability, and Microbial Communities in Soil Contaminated with Potentially Toxic Elements" Sustainability 15, no. 1: 790. https://0-doi-org.brum.beds.ac.uk/10.3390/su15010790