
Comparative Study between Dietary Nanoelemental, Inorganic, and Organic Selenium in Broiler Chickens: Effects on Meat Fatty Acid Composition and Oxidative Stability


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Diets, and Experimental Procedures
2.2. Determination of Se Content
2.3. Determination of Iron-Induced Lipid Oxidation in Meat
2.4. Determination of Fatty Acid Composition
2.5. Statistical Analysis
3. Results
3.1. Growth Performance
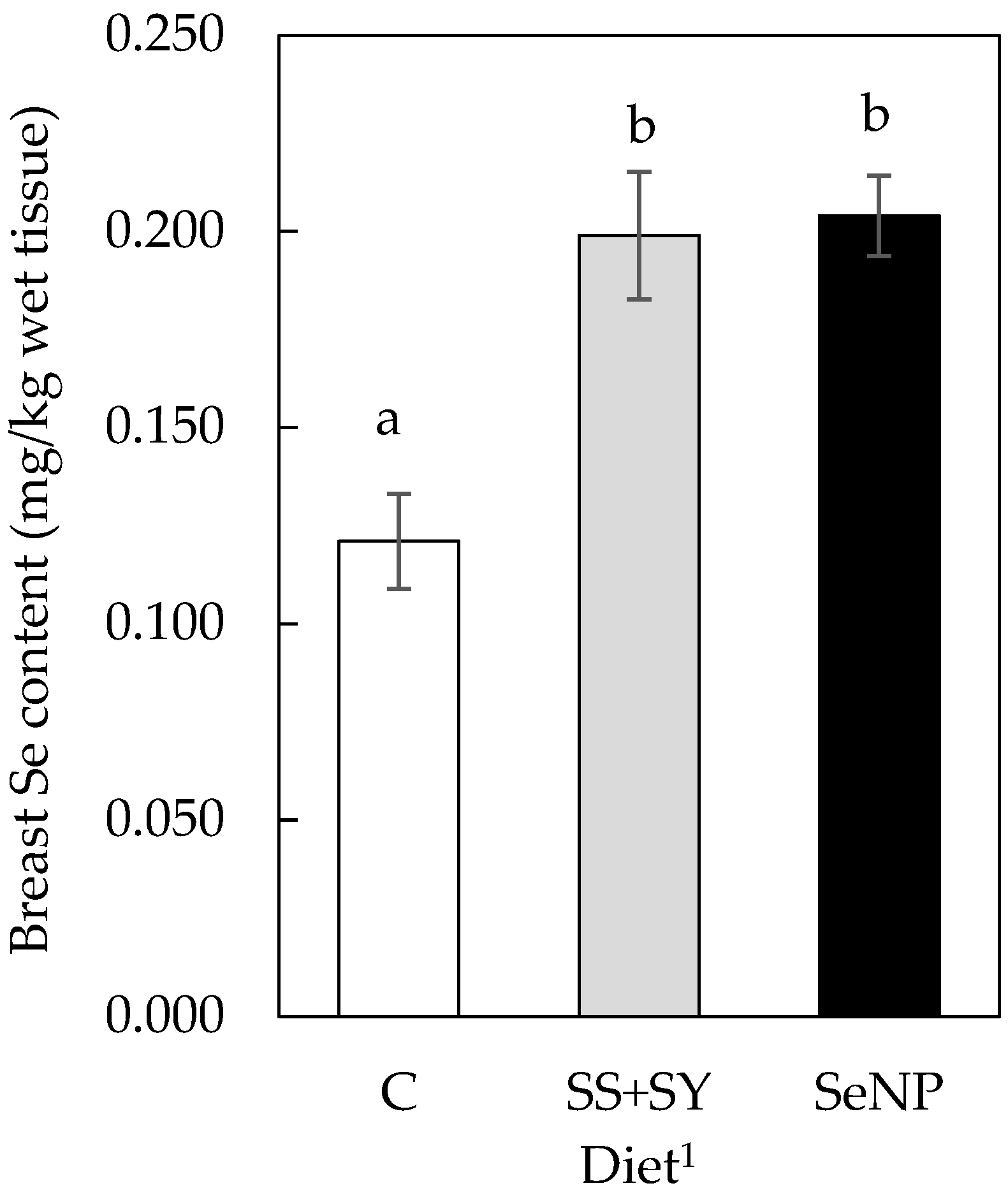
3.2. Breast Tissue Se Content
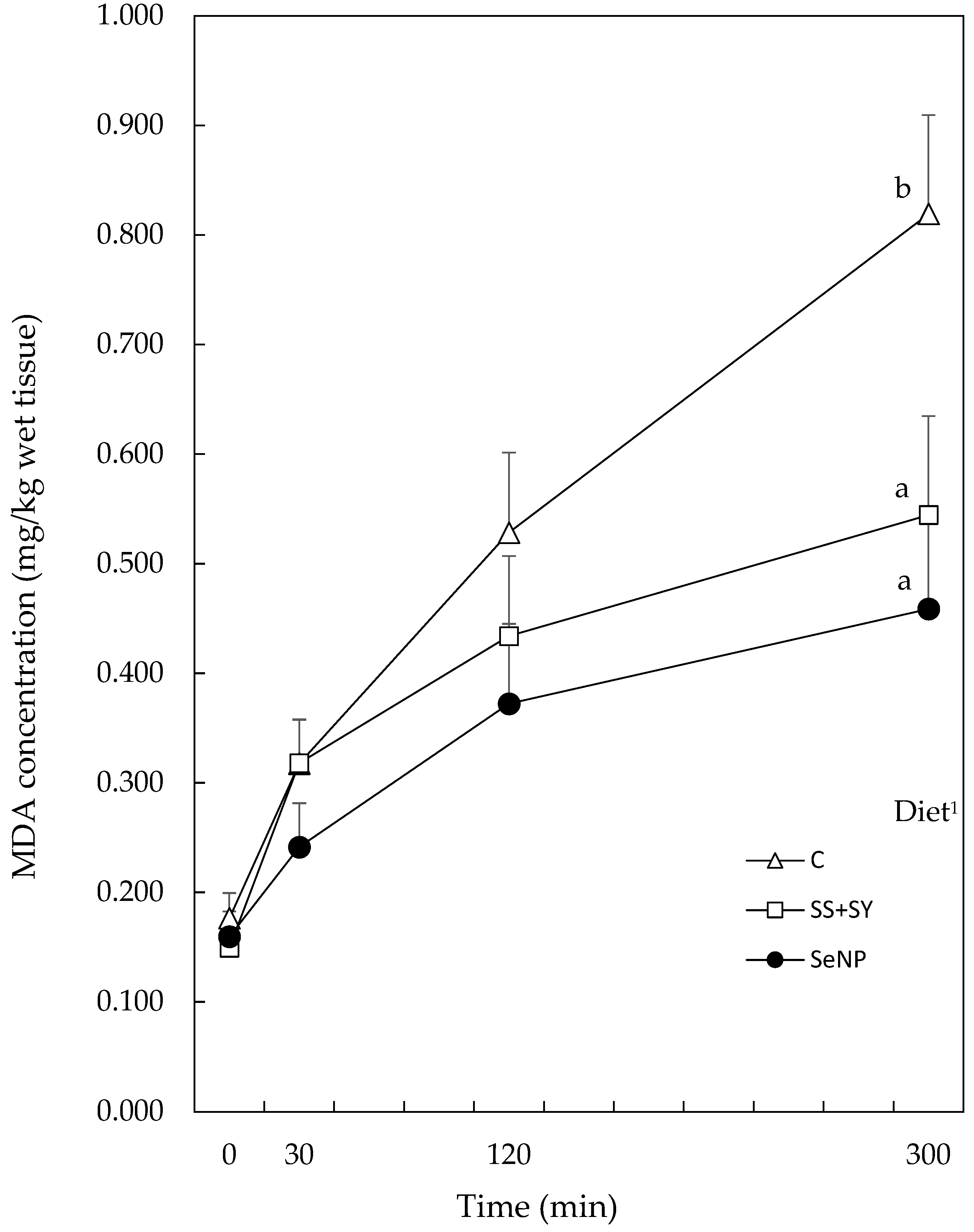
3.3. Breast Tissue Malondialdehyde Content
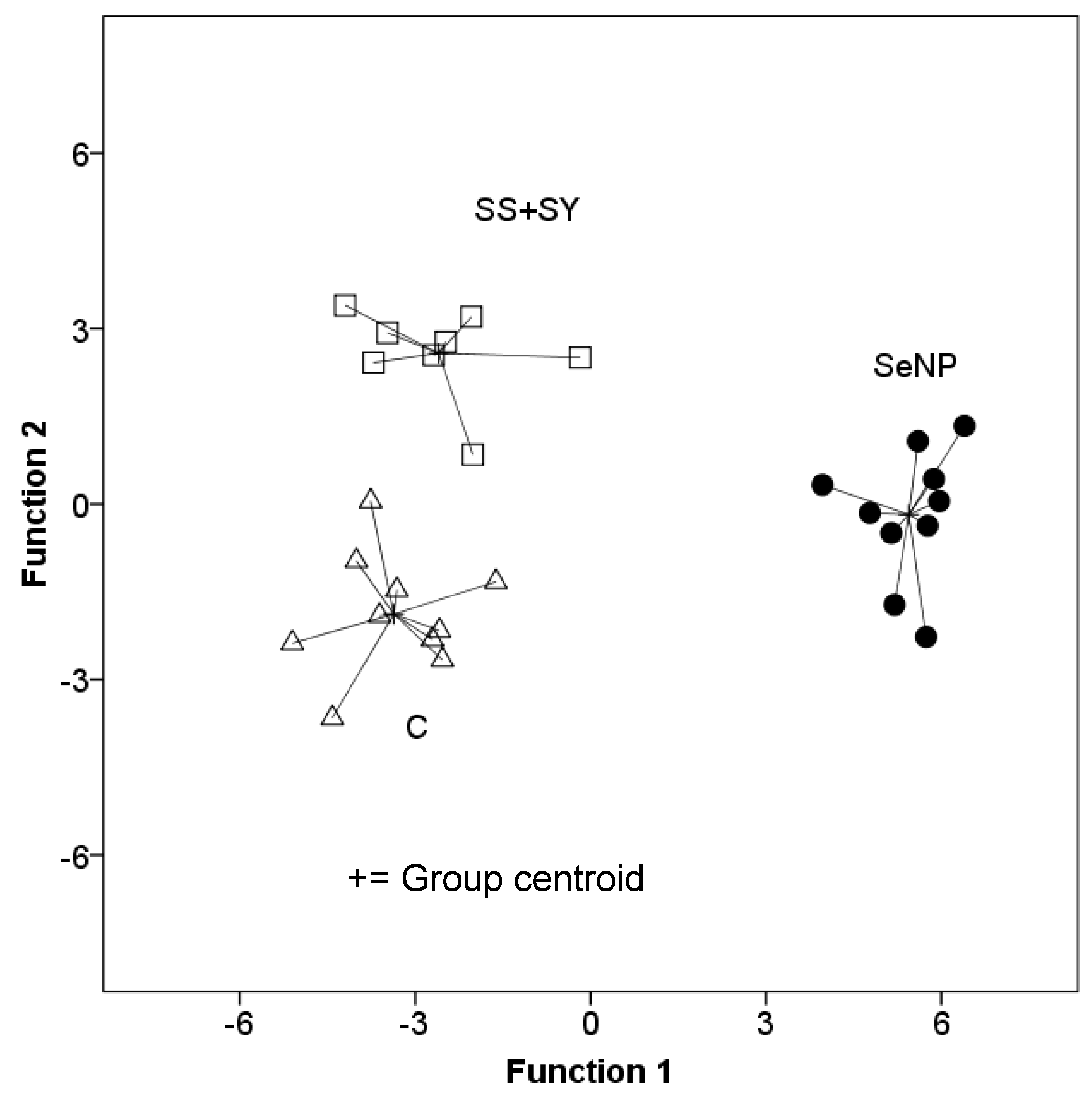
3.4. Breast Tissue Fatty Acid Composition
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kouba, M.; Mourot, J. A review of nutritional effects on fat composition of animal products with special emphasis on n-3 polyunsaturated fatty acids. Biochimie 2011, 93, 13–17. [Google Scholar] [CrossRef]
- Sadeghi, A.A.; Iravani, H.; Torshizi, M.K.; Chamani, M. Fatty acids profiles in meat of broiler chicks fed diet containing corn oil switched to fish oil at different weeks of age. World Appl. Sci. J. 2012, 18, 159–165. [Google Scholar]
- Jankowski, J.; Zdunczyk, Z.; Mikulski, D.; Juskiewicz, J.; Naczmanski, J.; Pomianowski, J.F.; Zdunczyk, P. Fatty acid profile, oxidative stability, and sensory properties of breast meat from turkeys fed diets with a different n-6/n-3 PUFA ratio. Eur. J. Lipid Sci. Technol. 2012, 114, 1025–1035. [Google Scholar] [CrossRef]
- Sohaib, M.; Anjum, F.M.; Arshad, M.S.; Imran, M.; Imran, A.; Hussain, S. Oxidative stability and lipid oxidation flavoring volatiles in antioxidants treated chicken meat patties during storage. Lipids Health Dis. 2017, 16, 16–27. [Google Scholar] [CrossRef] [Green Version]
- Surai, P.F. Natural antioxidants in poultry nutrition: New developments. In Proceedings of the 16th European Symposium on Poultry Nutrition, Strasbourg, France, 26–30 August 2007; pp. 669–676. [Google Scholar]
- Schweizer, U.; Streckfuß, F.; Pelt, P.; Carlson, B.A.; Hatfield, D.L.; Kohrle, J.; Schomburg, L. Hepatically derived selenoprotein P is a key factor for kidney but not for brain selenium supply. Biochem. J. 2005, 386, 221–226. [Google Scholar] [CrossRef]
- Yu, L.L.; Wang, R.; Zhang, Y.Z.; Kleemann, D.; Zhu, X.; Jia, Z. Effects of selenium supplementation on polyunsaturated fatty acid concentrations and antioxidant status in plasma and liver of lambs fed linseed oil or sunflower oil diets. Anim. Feed Sci. Technol. 2008, 140, 39–51. [Google Scholar] [CrossRef]
- Ringuet, M.T.; Hunne, B.; Lenz, M.; Bravo, D.M.; Furness, J.B. Analysis of bioavailability and induction of glutathione peroxidase by dietary nanoelemental, organic and inorganic selenium. Nutrients 2021, 13, 1073. [Google Scholar] [CrossRef]
- Rayman, M.P. Selenium intake, status, and health: A complex relationship. Hormones 2020, 19, 9–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khurana, A.; Tekula, S.; Saifi, M.A.; Venkatesh, P.; Godugu, C. Therapeutic applications of selenium nanoparticles. Biomed. Pharmacother. 2019, 111, 802–812. [Google Scholar] [CrossRef] [PubMed]
- Araujo, J.M.; Fortes-Silva, R.; Pola, C.C.; Yamamoto, F.Y.; Gatlin, D.M.; Gomes, C.L. Delivery of selenium using chitosan nanoparticles: Synthesis, characterization, and antioxidant and growth effects in Nile tilapia (Oreochromis niloticus). PLoS ONE 2021, 16, 0251786. [Google Scholar] [CrossRef] [PubMed]
- EC 2013. Commission Regulation (EC) No 427/2013 of 8 May 2013 concerning the authorisation of selenomethionine produced by Saccharomyces cerevisiae NCYC R646 as a feed additive for all Animal species and amending Regulations (EC) No 1750/2006, (EC) No 634/2007 and (EC) No 900/2009 as regards the maximum supplementation with selenised Yeast. Off. J. Eur. Union L 2013, 127, 20–22. [Google Scholar]
- Cai, S.J.; Wu, C.X.; Gong, L.M.; Song, T.; Wu, H.; Zhang, L.Y. Effects of nano—Selenium on performance, meat quality, immune function, oxidation resistance, and tissue selenium content in broilers. Poult. Sci. 2012, 91, 2532–2539. [Google Scholar] [CrossRef] [PubMed]
- Bień, D.; Michalczuk, M.; Szkopek, D.; Kinsner, M.; Konieczka, P. Changes in lipids metabolism indices as a result of different form of selenium supplementation in chickens. Sci. Rep. 2022, 12, 13817. [Google Scholar] [CrossRef] [PubMed]
- Samak, D.H.; El-Sayed, Y.S.; Shaheen, H.M.; El-Far, A.H.; Abd El-Hack, M.E.; Noreldin, A.E.; El-Naggar, K.; Abdelnour, S.A.; Saied, E.M.; El-Seedi, H.R.; et al. Developmental toxicity of carbon nanoparticles during embryogenesis in chicken. Environ. Sci. Pollut. Res. Int. 2020, 27, 19058–19072. [Google Scholar] [CrossRef]
- Kumbhar, S.; Khan, A.Z.; Parveen, F.; Nizamani, Z.A.; Siyal, F.A.; Abd El-Hack, M.E.; Gan, F.; Liu, Y.; Hamid, M.; Nido, S.A.; et al. Impacts of selenium and vitamin E supplementation on mRNA of heat shock proteins, selenoproteins and antioxidants in broilers exposed to high temperature. AMB Express 2018, 8, 112. [Google Scholar] [CrossRef] [PubMed]
- Abdul, H.; Sivaraj, R.; Venckatesh, R. Green synthesis and characterization of zinc oxide nanoparticles from Ocimum basilicum L. var purpurascens Benth.-lamiaceae leaf extract. Mater. Lett. 2014, 131, 16–18. [Google Scholar] [CrossRef]
- Gangadoo, S.; Stanley, D.; Hughes, R.J.; Moore, R.J.; Chapman, J. Nanoparticles in feed: Progress and prospects in poultry research. Trends Food Sci. Technol. 2016, 58, 115–126. [Google Scholar] [CrossRef]
- Li, H.; Liu, D.; Li, S.; Xue, C. Synthesis and cytotoxicity of selenium nanoparticles stabilized by α-D-glucan from Castanea mollissima Blume. Int. J. Biol. Macromol. 2019, 129, 818–826. [Google Scholar] [CrossRef]
- Zhang, J.S.; Wang, X.; Xu, T. Elemental selenium at nano size (Nano-Se) as a potential chemopreventive agent with reduced risk of selenium toxicity: Comparison with Se-Methylselenocysteine in mice. Toxicol. Sci. 2008, 101, 22–31. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zhang, J.; Yu, H. Elemental selenium at nano size possesses lower toxicity without compromising the fundamental effect on selenoenzymes: Comparison with selenomethionine in mice. Free Radic. Biol. Med. 2007, 42, 1524–1533. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, Y. Influence of dietary nano elemental selenium on growth performance, tissue selenium distribution, meat quality and glutathione peroxidase activity in Guangxi Yellow Chicken. Poult. Sci. 2011, 90, 680–686. [Google Scholar] [CrossRef]
- Saleh, A.A.; Ebeid, T.A. Feeding sodium selenite and nano-selenium stimulates growth and oxidation resistance in broilers. S. Afr. J. Anim. Sci. 2019, 49, 176–184. [Google Scholar] [CrossRef] [Green Version]
- Aparna, N.; Karunakaran, R. Effect of selenium nanoparticles supplementation on oxidation resistance of broiler chicken. Indian J. Sci. Technol. 2016, 9, 1–5. [Google Scholar] [CrossRef]
- Li, B.; Li, D.; Jing, W.; Fan, J.; Dahms, H.U.; Lee, S.C.; Wang, L. Biogenic selenium and its hepatoprotective activity. Sci. Rep. 2017, 7, 15627. Available online: https://0-www-nature-com.brum.beds.ac.uk/articles/s41598-017-13636-1 (accessed on 7 April 2023). [CrossRef] [PubMed] [Green Version]
- Bai, K.; Hong, B.; He, J.; Hong, Z.; Tan, R. Preparation and antioxidant properties of selenium nanoparticles-loaded chitosan microspheres. Int. J. Nanomed. 2017, 12, 4527–4539. [Google Scholar] [CrossRef] [Green Version]
- Ren, L.; Wu, Z.; Ma, Y.; Jian, W.; Xiong, H.; Zhou, L. Preparation and growth-promoting effect of selenium nanoparticles capped by polysaccharide-protein complexes on tilapia. J. Sci. Food Agric. 2021, 101, 476–485. [Google Scholar] [CrossRef] [PubMed]
- Filipović, N.; Ušjak, D.; Milenković, M.T.; Zheng, K.; Liverani, L.; Boccaccini, A.R.; Stevanović, M.M. Comparative study of the antimicrobial activity of selenium nanoparticles with different surface chemistry and structure. Front. Bioeng. Biotechnol. 2021, 8, 624621. [Google Scholar] [CrossRef]
- Zanini, S.F.; Torres, C.A.A.; Bragagnolo, N.; Turatti, J.M.; Silva, M.G.; Zanini, M.S. Effect of oil sources and vitamin E levels in the diet on the composition of fatty acids in rooster thigh and chest meat. J. Sci. Food Agric. 2004, 84, 672–682. [Google Scholar] [CrossRef]
- González, E.; Tejeda, J.F. Effects of dietary incorporation of different antioxidant extracts and free-range rearing on fatty acid composition and lipid oxidation of Iberian pig meat. Animal 2007, 1, 1060–1067. [Google Scholar] [CrossRef] [Green Version]
- Pappas, A.C.; Zoidis, E.; Papadomichelakis, G.; Fegeros, K. Supranutritional selenium level affects fatty acid composition and oxidative stability of chicken breast muscle tissue. J. Anim. Physiol. Anim. Nutr. 2011, 96, 385–394. [Google Scholar] [CrossRef]
- Pereira, A.S.C.; Santos, M.V.; Aferri, G.; Silva Corte, R.R.P.; Silva, S.L.; de Freitas, J.E., Jr.; Leme, P.R.; Renno, F.P. Lipid and selenium sources on fatty acid composition of intramuscular fat and muscle selenium concentration of Nellore steers. Rev. Bras. Zootec. 2012, 41, 2357–2363. [Google Scholar] [CrossRef] [Green Version]
- Schäfer, K.; Kyriakopoulos, A.; Gessner, H.; Grune, T.; Behne, D. Effects of selenium deficiency on fatty acid metabolism in rats fed fish oil-enriched diets. J. Trace Elem. Med. Biol. 2004, 18, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Netto, A.S.; Zanetti, M.A.; Ribeiro del Claro, G.; Pires de Melo, M.; Garcia Vilela, F.; Bertonha Correa, L. Effects of copper and selenium supplementation on performance and lipid metabolism in confined brangus bulls. Asian-Austral. J. Anim. Sci. 2014, 27, 488–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papadomichelakis, G.; Zoidis, E.; Pappas, A.C.; Mountzouris, K.C.; Fegeros, K. Effects of increasing dietary organic selenium levels on meat fatty acid composition and oxidative stability in growing rabbits. Meat Sci. 2017, 131, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Fortatos, E.; Giamouri, E.; Pappas, A.C.; Yannopoulos, S.N.; Papadomichelakis, G. Selenium nanoparticles-loaded chitosan microspheres as a dietary selenium source in rabbits: Impact on meat selenium content and oxidative stability. Acta Sci. Vet. Sci. 2021, 3, 27–38. Available online: https://actascientific.com/ASVS/ASVS-03-0237.php (accessed on 2 February 2023). [CrossRef]
- Pappas, A.C.; Acamovic, T.; Sparks, N.H.C.; Surai, P.F.; McDevitt, R.M. Effects of supplementing broiler breeder diets with organoselenium compounds and polyunsaturated fatty acids on hatchability. Poult. Sci. 2006, 85, 1584–1593. [Google Scholar] [CrossRef]
- Terevinto, A.; Ramos, A.; Castroman, G.; Cabrera, M.C.; Saadoun, A. Oxidative status, in vitro iron-induced lipid oxidation and superoxide dismutase, catalase and glutathione peroxidase activities in rhea meat. Meat Sci. 2010, 84, 706–710. [Google Scholar] [CrossRef]
- O’Fallon, J.V.; Busboom, J.R.; Nelson, M.L.; Gaskins, C.T. A direct method for fatty acid methyl-ester synthesis: Application to wet meat tissues, oils, and feedstuffs. J. Anim. Sci. 2007, 85, 1511–1521. [Google Scholar] [CrossRef] [Green Version]
- IBM Corp. Released, IBM SPSS Statistics for Windows, Version 23.0; IBM Corp.: Armonk, NY, USA, 2015. [Google Scholar]
- Templeton, G.F. A two-step approach for transforming continuous variables to normal: Implications and recommendations for IS research. Commun. Assoc. Inf. Syst. 2011, 28, 41–58. [Google Scholar] [CrossRef] [Green Version]
- National Research Council (NRC). Nutrient Requirements of Poultry, 9th ed.; National Academies Press: Washington, DC, USA, 1994. [Google Scholar]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I.; Velichko, O.A. Selenium in poultry nutrition: From sodium selenite to organic selenium sources. J. Poult. Sci. 2018, 55, 79–93. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.H.; Li, Y.L.; Xiong, L.; Zhang, H.M.; Song, J.; Xia, M.S. Comparative effects of nano elemental selenium and sodium selenite on selenium retention in broiler chickens. Anim. Feed Sci. Technol. 2012, 177, 204–210. [Google Scholar] [CrossRef]
- Ahmad, H.; Tian, J.; Wang, J.; Khan, M.A.; Wang, Y.; Zhang, L.; Wang, T. Effects of Dietary Sodium Selenite and Selenium Yeast on Antioxidant Enzyme Activities and Oxidative Stability of Chicken Breast Meat. J. Agric. Food Chem. 2012, 60, 7111–7120. [Google Scholar] [CrossRef]
- Bien, D.; Michalczuk, M.; Łysek-Gładysinsk, M.; Józwik, A.; Wieczorek, A.; Matuszewski, A.; Kinsner, M.; Konieczka, P. Nano-sized selenium maintains performance and improves health status and antioxidant potential while not compromising ultrastructure of breast muscle and liver in chickens. Antioxidants 2023, 12, 905. [Google Scholar] [CrossRef]
- Suzuki, K.T.; Ogra, Y. Metabolic pathway for selenium in the body: Speciation by HPLC-ICP MS with enriched Se. Food Addit. Contam. 2022, 19, 974–983. [Google Scholar] [CrossRef] [PubMed]
- Gangadoo, S.; Dinev, I.; Chapman, J.; Hughes, R.J.; Hao Van, T.T.; Moore, R.; Stanley, D. Selenium nanoparticles in poultry feed modify gut microbiota and increase abundance of Faecalibacterium prausnitzii. Appl. Microbiol. Biotechnol. 2018, 102, 1455–1466. Available online: https://0-link-springer-com.brum.beds.ac.uk/article/10.1007%2Fs00253-017-8688-4 (accessed on 4 March 2023). [CrossRef]
- Visha, P.; Nanjappan, K.; Selvaraj, P.; Jayachandran, S.; Thavasiappan, V. Influence of dietary nanoselenium supplementation on the meat characteristics of broiler chickens. Int. J. Curr. Microbiol. App. Sci. 2017, 6, 340–347. [Google Scholar] [CrossRef]
- Botsoglou, N.A.; Florou-Paneri, P.; Christaki, E.; Fletouris, D.J.; Spais, A.B. Effect of dietary oregano essential oil on performance of chickens and on iron-induced lipid oxidation of breast, thigh and abdominal fat tissues. Br. Poult. Sci. 2002, 43, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Voss, A.; Reinhart, M.; Sankarappa, S.; Sprecher, H. The metabolism of 7,10,13,16,19-docosapentaenoic acid to 4,7,10,13,16,19-docosahexaenoic acid in rat liver is independent of a delta-4-desaturase. J. Biol. Chem. 1991, 266, 19995–20000. [Google Scholar] [CrossRef]
- Sprecher, H.; Luthria, D.L.; Mohammed, B.S.; Baykousheva, S.P. Re-evaluation of the pathways for the biosynthesis of polyunsaturated fatty acids. J. Lipid Res. 1995, 36, 2471–2477. [Google Scholar] [CrossRef]
- Glauert, H.P.; Beaty, M.M.; Clark, T.D.; Greenwell, W.S.; Chow, C.K. Effect of dietary selenium on the induction of altered hepatic foci and hepatic tumors by the peroxisome proliferator ciprofibrate. Nutr. Cancer 1990, 14, 261–271. [Google Scholar] [CrossRef]
- Lee, J.M.; Lee, H.; Kang, S.B.; Park, W.J. Fatty acid desaturases, polyunsaturated fatty acid regulation, and biotechnological advances. Nutrients 2016, 8, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Basal Diets | ||||
|---|---|---|---|---|
| Starter (0–10 d) | Grower (11–24 d) | Finisher (25–42 d) | ||
| Ingredient | ||||
| Maize | 485 | 521 | 576 | |
| Soybean meal, 450 g CP/kg | 428 | 390 | 334 | |
| Soybean oil | 44.7 | 51.7 | 56.0 | |
| Monocalcium phosphate | 14.3 | 12.3 | 10.6 | |
| Limestone | 14.1 | 12.8 | 11.6 | |
| Sodium chloride | 4.0 | 4.0 | 4.0 | |
| DL-methionine, 99% | 3.6 | 3.1 | 2.8 | |
| L-lysine HCl, 80% | 2.5 | 1.7 | 1.8 | |
| L-threonine | 1.0 | 0.7 | 0.4 | |
| Premix 1 | 2.0 | 2.0 | 2.0 | |
| Choline | 0.8 | 0.7 | 0.8 | |
| Calculated chemical composition | ||||
| Dry matter | 880 | 880 | 880 | |
| Crude protein | 230 | 215 | 195 | |
| Ether extract | 69 | 77 | 82 | |
| Lysine | 14.4 | 12.9 | 11.6 | |
| Methionine + cystine | 10.8 | 9.9 | 9.1 | |
| Threonine | 9.7 | 8.8 | 7.8 | |
| Calcium | 9.6 | 8.7 | 7.8 | |
| Available phosphorus | 4.8 | 4.4 | 3.9 | |
| Metabolizable energy, MJ/kg | 12.6 | 13.0 | 13.4 | |
| Fatty acid composition | ||||
| 12:0 | 0.16 | |||
| 14:0 | 0.62 | |||
| 15:0 | 0.13 | |||
| C16:0 | 27.33 | |||
| C16:1 | 0.34 | |||
| 17:0 | 0.17 | |||
| C18:0 | Not determined | Not determined | 19.98 | |
| C18:1n − 7 | 11.90 | |||
| C18:2n − 6 | 30.10 | |||
| C18:3n − 3 | 4.15 | |||
| C20:0 | 0.49 | |||
| C20:1n − 9 | 0.43 | |||
| C20:2n − 6 | 0.06 | |||
| C22:0 | 0.50 | |||
| C22:1 | 0.36 | |||
| C23:0 | 0.15 | |||
| C24:0 | 0.35 | |||
| C24:1 | 0.08 | |||
| Se content (mg/kg) | ||||
| Experimental diets | Added 2 | Determined 3 | ||
| C | Se not determined | - | 0.117 ± 0.020 | |
| SS+SY | 0.40 | 0.492 ± 0.049 | ||
| SeNP | 0.40 | 0.488 ± 0.045 | ||
| Diet 1 | SEM 2 | p-Value 3 | |||
|---|---|---|---|---|---|
| C | SS+SY | SeNP | |||
| Total FA weights | 1258 | 1353 | 1285 | 88.5 | 0.552 |
| 14:0 | 0.28 | 0.27 | 0.26 | 0.010 | 0.159 |
| 15:0 | 0.05 a | 0.06 ab | 0.07 b | 0.008 | 0.032 |
| 16:0 | 16.51 | 16.43 | 16.21 | 0.240 | 0.451 |
| 16:1n − 9 | 0.25 | 0.24 | 0.25 | 0.019 | 0.824 |
| 16:1n − 7 | 1.15 | 1.15 | 0.99 | 0.120 | 0.310 |
| 17:0 | 0.17 | 0.16 | 0.17 | 0.007 | 0.397 |
| 17:1 | 0.60 | 0.67 | 0.74 | 0.056 | 0.068 |
| 18:0 | 9.25 | 9.21 | 9.84 | 0.391 | 0.212 |
| 18:1n − 9 | 22.47 | 22.22 | 21.54 | 0.739 | 0.424 |
| 18:1n − 7 | 1.66 | 1.67 | 1.65 | 0.060 | 0.965 |
| 18:2n − 6 | 31.10 | 31.33 | 30.66 | 0.899 | 0.757 |
| 18:3n − 6 | 0.20 | 0.20 | 0.20 | 0.012 | 0.790 |
| 18:3n − 3 | 2.72 | 2.80 | 2.70 | 0.157 | 0.799 |
| 20:1n − 9 | 0.21 | 0.21 | 0.21 | 0.008 | 0.685 |
| 20:2n − 6 | 0.71 | 0.74 | 0.83 | 0.061 | 0.165 |
| 20:3n − 6 | 0.74 | 0.67 | 0.94 | 0.184 | 0.342 |
| 20:4n − 6 | 4.59 | 5.02 | 5.64 | 0.628 | 0.266 |
| 20:3n − 3 | 0.05 | 0.07 | 0.01 | 0.031 | 0.171 |
| 20:5n − 3 | 0.23 | 0.23 | 0.28 | 0.032 | 0.216 |
| 22:4n − 6 | 1.37 | 1.32 | 1.45 | 0.125 | 0.552 |
| 22:5n − 3 | 1.03 | 1.09 | 1.22 | 0.107 | 0.233 |
| 22:6n − 3 | 0.63 a | 0.65 a | 0.86 b | 0.082 | 0.017 |
| ΣSFA 4 | 26.24 | 26.04 | 26.55 | 0.477 | 0.565 |
| ΣMUFA 4 | 26.31 | 26.27 | 25.40 | 0.799 | 0.434 |
| ΣPUFA 4 | 43.38 | 44.12 | 44.79 | 0.795 | 0.223 |
| ΣPUFA/ΣSFA | 1.66 | 1.70 | 1.69 | 0.049 | 0.650 |
| Σn – 6 5 | 38.00 | 38.53 | 38.90 | 0.752 | 0.500 |
| Σn – 3 5 | 4.66 a | 4.84 a | 5.07 b | 0.093 | <0.001 |
| ΣLCn – 3 6 | 1.95 | 2.04 | 2.37 | 0.185 | 0.077 |
| Σn − 6/Σn − 3 | 8.17 b | 7.97 ab | 7.69 a | 0.180 | 0.038 |
| Δ9-desaturase index 7 | 0.48 | 0.47 | 0.46 | 0.013 | 0.520 |
| Δ5,6-desaturase index 8 | 0.18 | 0.19 | 0.21 | 0.020 | 0.230 |
| Elongase index 9 | 0.56 | 0.56 | 0.61 | 0.024 | 0.093 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giamouri, E.; Fortatos, E.; Pappas, A.C.; Papadomichelakis, G. Comparative Study between Dietary Nanoelemental, Inorganic, and Organic Selenium in Broiler Chickens: Effects on Meat Fatty Acid Composition and Oxidative Stability. Sustainability 2023, 15, 9762. https://0-doi-org.brum.beds.ac.uk/10.3390/su15129762
Giamouri E, Fortatos E, Pappas AC, Papadomichelakis G. Comparative Study between Dietary Nanoelemental, Inorganic, and Organic Selenium in Broiler Chickens: Effects on Meat Fatty Acid Composition and Oxidative Stability. Sustainability. 2023; 15(12):9762. https://0-doi-org.brum.beds.ac.uk/10.3390/su15129762
Chicago/Turabian StyleGiamouri, Elisavet, Efstathios Fortatos, Athanasios C. Pappas, and George Papadomichelakis. 2023. "Comparative Study between Dietary Nanoelemental, Inorganic, and Organic Selenium in Broiler Chickens: Effects on Meat Fatty Acid Composition and Oxidative Stability" Sustainability 15, no. 12: 9762. https://0-doi-org.brum.beds.ac.uk/10.3390/su15129762