Using APAR to Predict Aboveground Plant Productivity in Semi-Arid Rangelands: Spatial and Temporal Relationships Differ

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
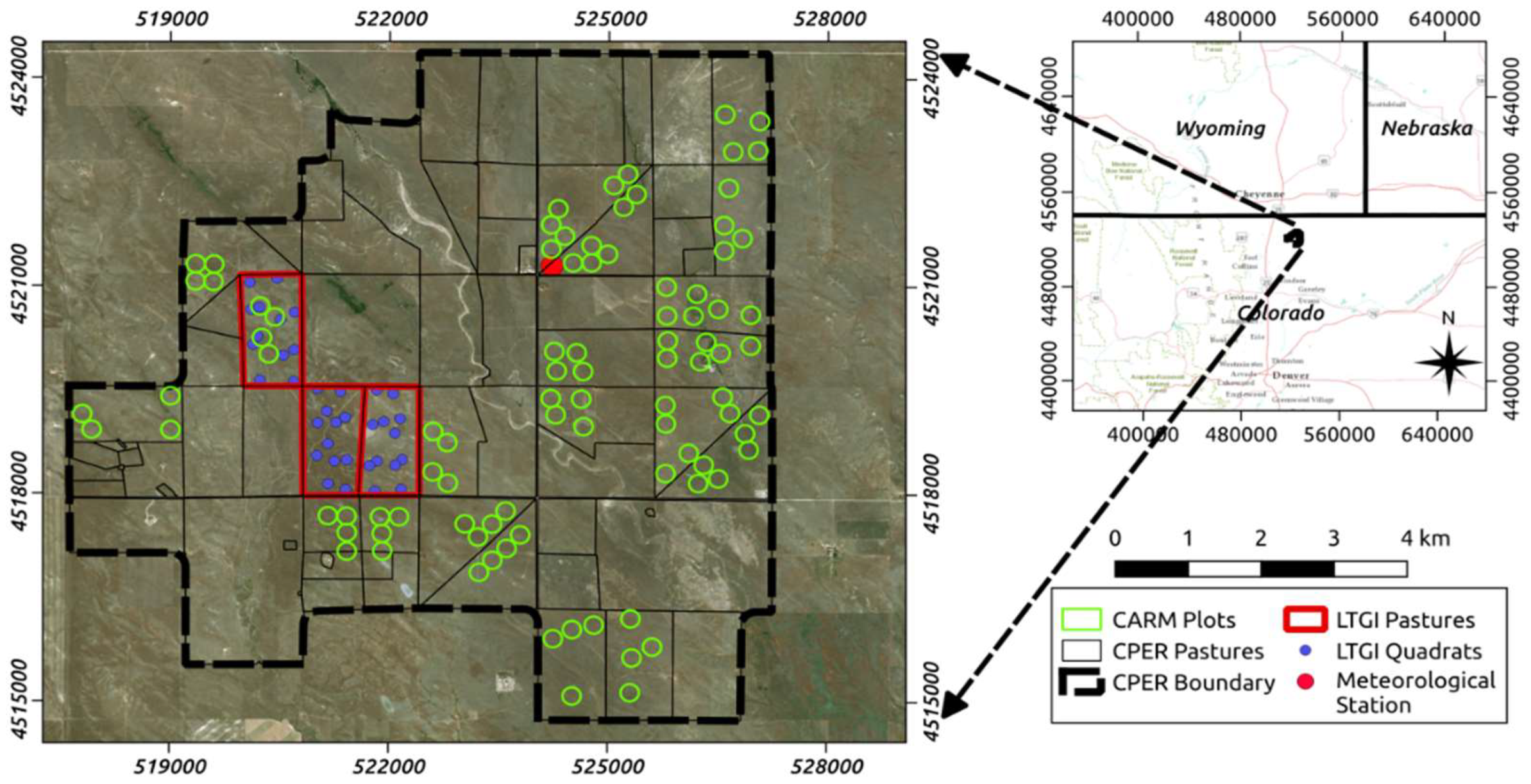
2.1. Study Area
2.2. Ground Based Observations
2.3. Remotely Sensed Observations and Aggregations
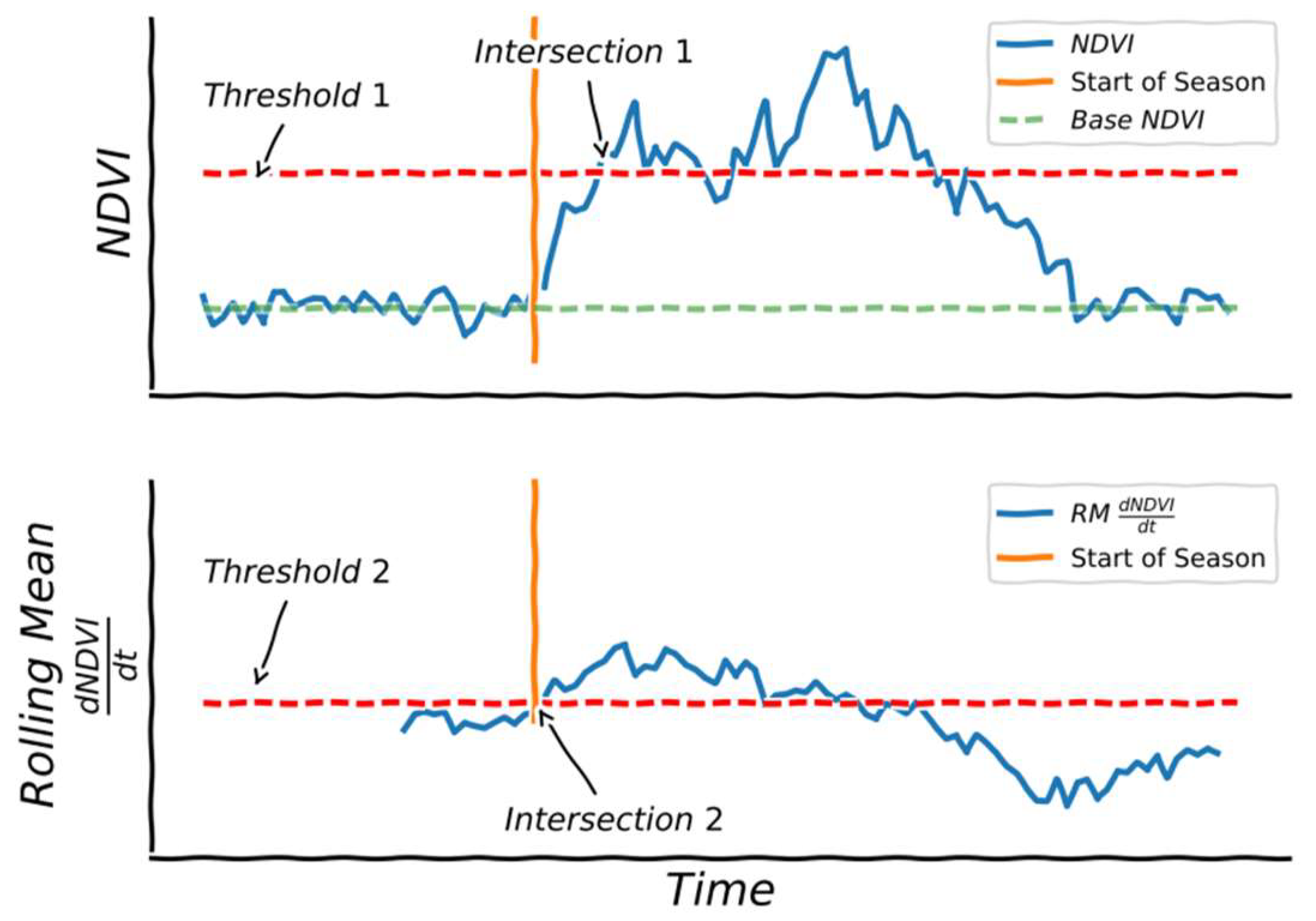
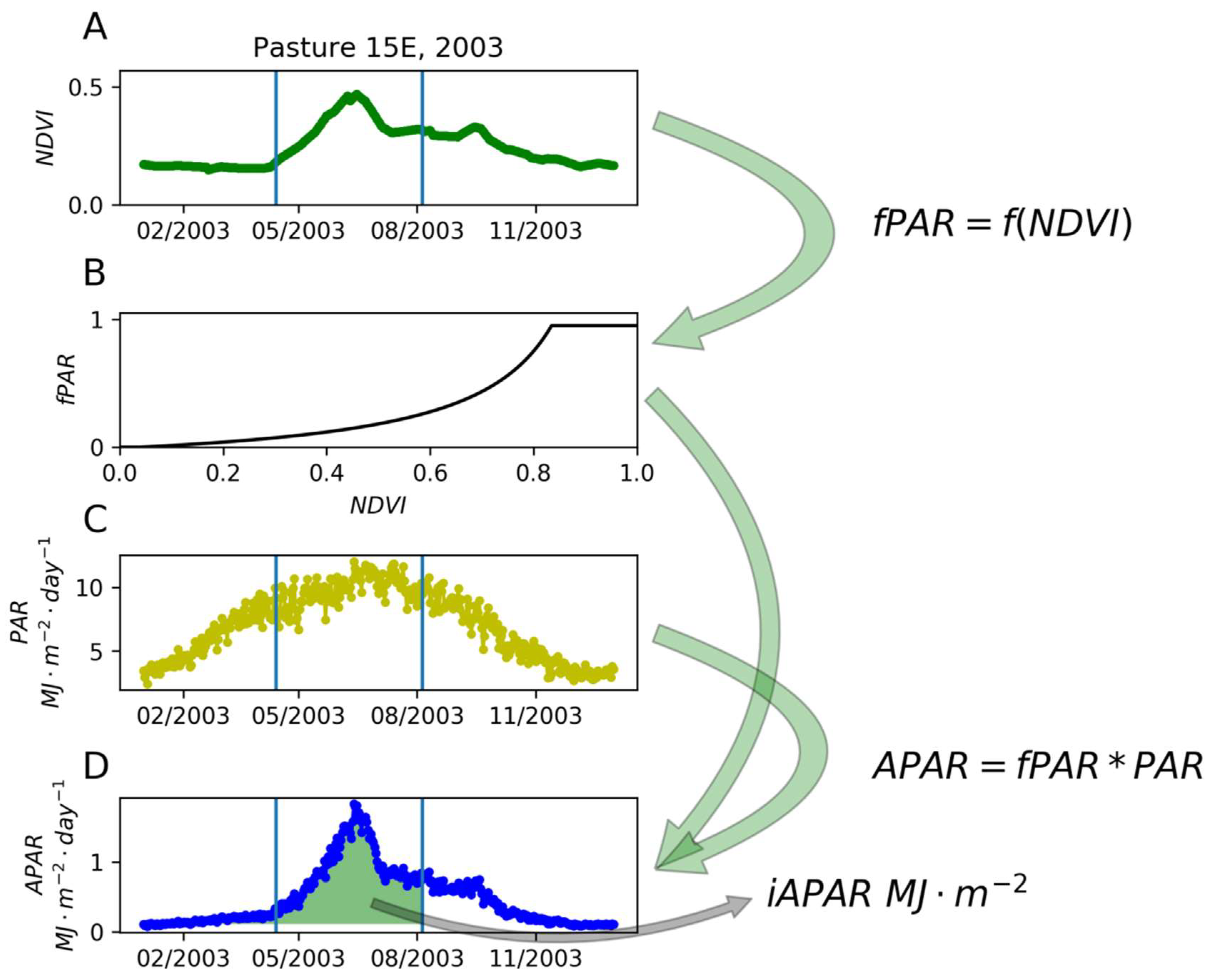
2.4. Phenological Calculations
2.5. Statistical Approaches
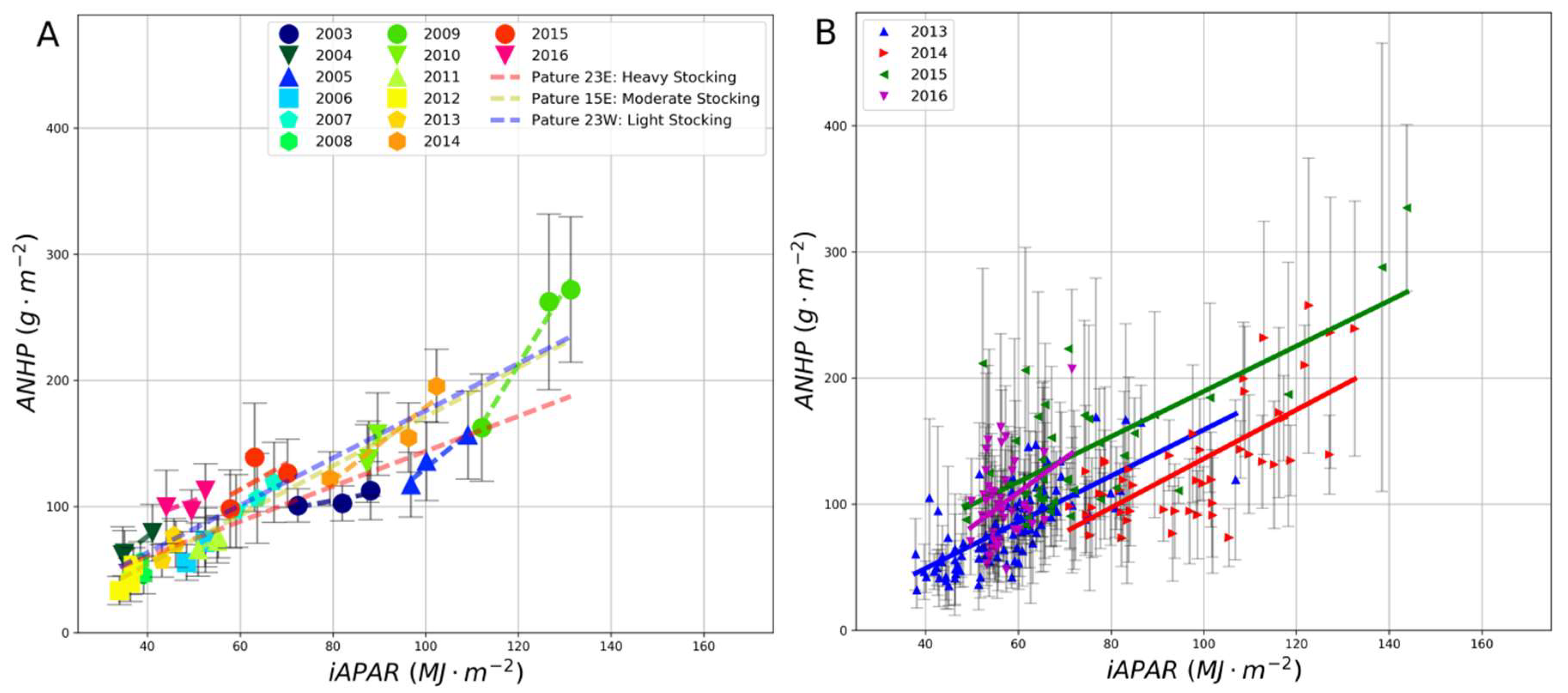
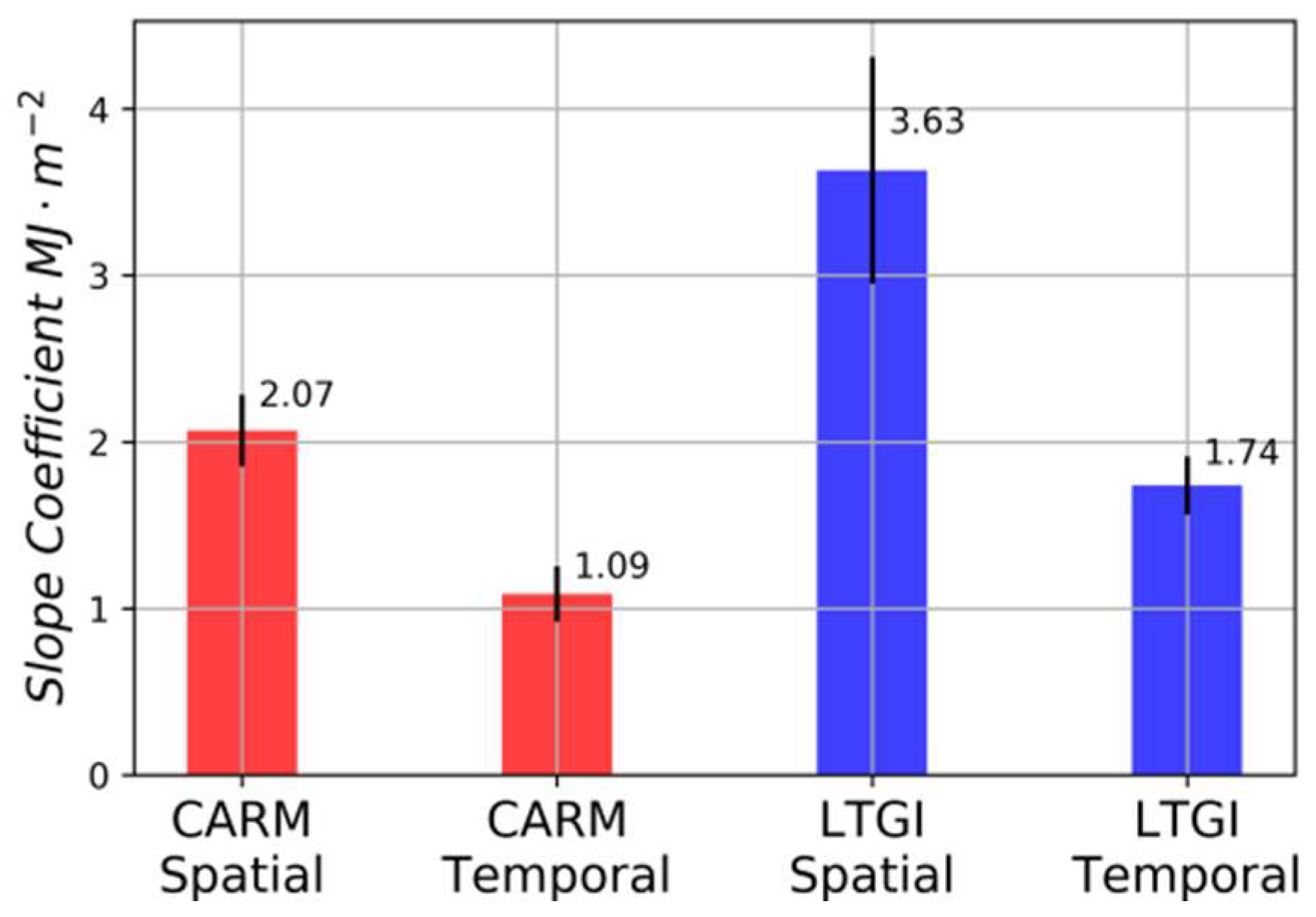
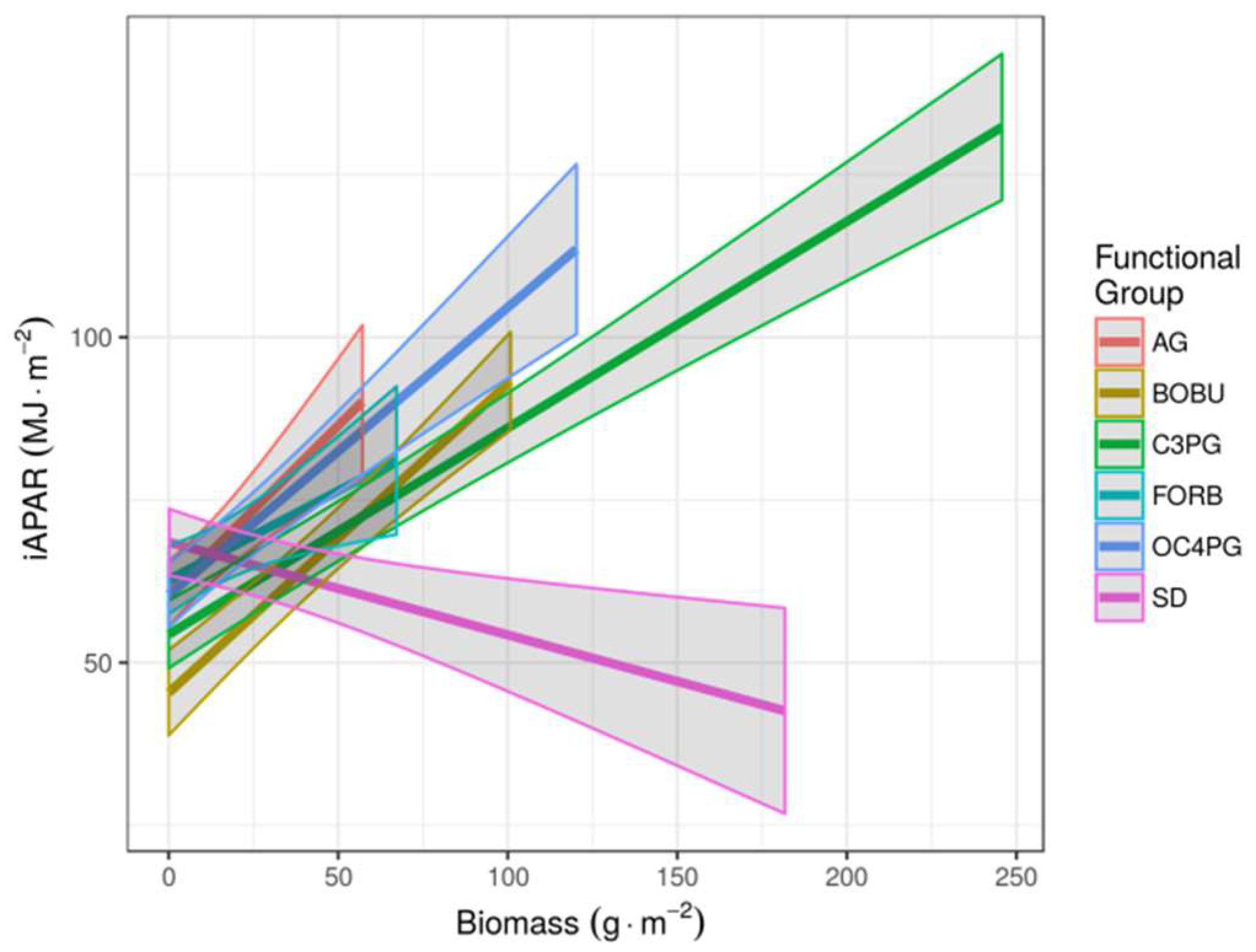
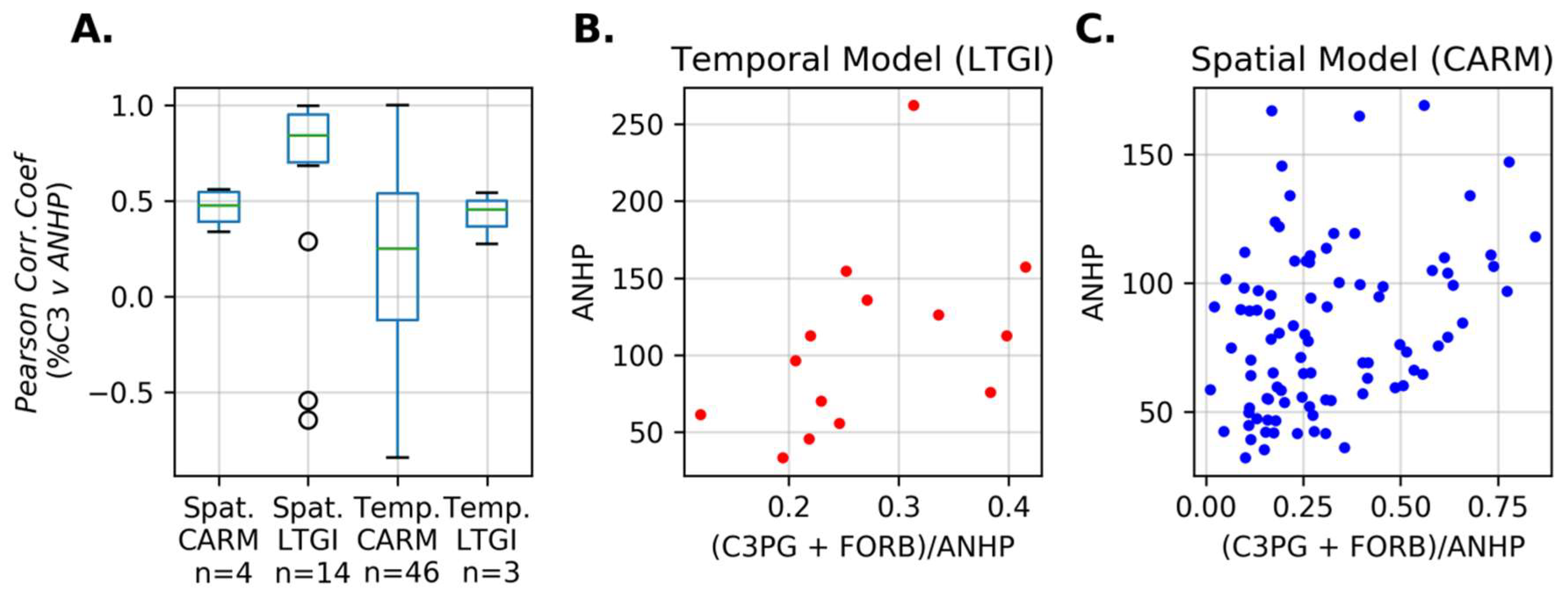
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bastian, C.T.; Ritten, J.P.; Derner, J.D. Ranch Profitability Given Increased Precipitation Variability and Flexible Stocking. J. Am. Soc. Farm Manag. Rural Apprais. 2018, 122–139. [Google Scholar]
- Derner, J.D.; Augustine, D.J. Adaptive Management for Drought on Rangelands. Rangelands 2016, 38, 211–215. [Google Scholar] [CrossRef]
- Sellers, P.J.; Berry, J.A.; Collatz, G.J.; Field, C.B.; Hall, F.G. Canopy reflectance, photosynthesis, and transpiration. III. A reanalysis using improved leaf models and a new canopy integration scheme. Remote Sens. Environ. 1992, 42, 187–216. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.; Gao, X.; Ferreira, L. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Di Bella, C.M.; Paruelo, J.M.; Becerra, J.E.; Bacour, C.; Baret, F. Effect of senescent leaves on NDVI-based estimates of f APAR: Experimental and modelling evidences. Int. J. Remote Sens. 2004, 25, 5415–5427. [Google Scholar] [CrossRef]
- Paruelo, J.M.; Epstein, H.E.; Lauenroth, W.K.; Burke, I.C. ANPP Estimates From NDVI For The Central Grassland Region of the United States. Ecology 1997, 78, 953–958. [Google Scholar] [CrossRef]
- Liu, Z.; Huang, M. Assessing spatio-temporal variations of precipitation-use efficiency over Tibetan grasslands using MODIS and in-situ observations. Front. Earth Sci. 2016, 10, 784–793. [Google Scholar] [CrossRef]
- Piñeiro, G.; Oesterheld, M.; Paruelo, J.M. Seasonal Variation in Aboveground Production and Radiation-use Efficiency of Temperate rangelands Estimated through Remote Sensing. Ecosystems 2006, 9, 357–373. [Google Scholar] [CrossRef]
- Chen, M.; Parton, W.J.; Del Grosso, S.J.; Hartman, M.D.; Day, K.A.; Tucker, C.J.; Derner, J.D.; Knapp, A.K.; Smith, W.K.; Ojima, D.S.; et al. The signature of sea surface temperature anomalies on the dynamics of semiarid grassland productivity. Ecosphere 2017, 8, e02069. [Google Scholar] [CrossRef] [Green Version]
- Tucker, C.J.; Vanpraet, C.L.; Sharman, M.J.; Ittersum, G.V. Satellite remote sensing of total herbaceous biomass production in the senegalese sahel: 1980–1984. Remote Sens. Environ. 1985, 17, 233–249. [Google Scholar] [CrossRef]
- Lo Seen Chong, D.; Mougin, E.; Gastellu-Etchegorry, J.P. Relating the Global Vegetation Index to net primary productivity and actual evapotranspiration over Africa. Int. J. Remote Sens. 1993, 14, 1517–1546. [Google Scholar] [CrossRef]
- Garbulsky, M.F.; Peñuelas, J.; Gamon, J.; Inoue, Y.; Filella, I. The photochemical reflectance index (PRI) and the remote sensing of leaf, canopy and ecosystem radiation use efficiencies: A review and meta-analysis. Remote Sens. Environ. 2011, 115, 281–297. [Google Scholar] [CrossRef]
- Gamon, J.A.; Field, C.B.; Goulden, M.L.; Griffin, K.L.; Hartley, A.E.; Joel, G.; Penuelas, J.; Valentini, R. Relationships Between NDVI, Canopy Structure, and Photosynthesis in Three Californian Vegetation Types. Ecol. Appl. 1995, 5, 28–41. [Google Scholar] [CrossRef] [Green Version]
- Grigera, G.; Oesterheld, M.; Pacín, F. Monitoring forage production for farmers’ decision making. Agric. Syst. 2007, 94, 637–648. [Google Scholar] [CrossRef]
- Hermance, J.F.; Augustine, D.J.; Derner, J.D. Quantifying characteristic growth dynamics in a semi-arid grassland ecosystem by predicting short-term NDVI phenology from daily rainfall: A simple four parameter coupled-reservoir model. Int. J. Remote Sens. 2015, 36, 5637–5663. [Google Scholar] [CrossRef]
- Moran, M.S.; Ponce-Campos, G.E.; Huete, A.; McClaran, M.P.; Zhang, Y.; Hamerlynck, E.P.; Augustine, D.J.; Gunter, S.A.; Kitchen, S.G.; Peters, D.P.; et al. Functional response of US grasslands to the early 21st-century drought. Ecology 2014, 95, 2121–2133. [Google Scholar] [CrossRef] [PubMed]
- Scott Butterfield, H.; Malmstrom, C.M. Experimental Use of Remote Sensing by Private Range Managers and Its Influence on Management Decisions. Rangel. Ecol. Manag. 2006, 59, 541–548. [Google Scholar] [CrossRef]
- Washington-Allen, R.A.; West, N.E.; Ramsey, R.D.; Efroymson, R.A. A Protocol for Retrospective Remote Sensing–Based Ecological Monitoring of Rangelands. Rangel. Ecol. Manag. 2006, 59, 19–29. [Google Scholar] [CrossRef]
- Tebbs, E.; Rowland, C.; Smart, S.; Maskell, L.; Norton, L. Regional-Scale High Spatial Resolution Mapping of Aboveground Net Primary Productivity (ANPP) from Field Survey and Landsat Data: A Case Study for the Country of Wales. Remote Sens. 2017, 9, 801. [Google Scholar] [CrossRef]
- Blanco, L.J.; Ferrando, C.A.; Biurrun, F.N. Remote Sensing of Spatial and Temporal Vegetation Patterns in Two Grazing Systems. Rangel. Ecol. Manag. 2009, 62, 445–451. [Google Scholar] [CrossRef]
- Porensky, L.M.; Derner, J.D.; Augustine, D.J.; Milchunas, D.G. Plant Community Composition After 75 Yr of Sustained Grazing Intensity Treatments in Shortgrass Steppe. Rangel. Ecol. Manag. 2017, 70, 456–464. [Google Scholar] [CrossRef]
- Augustine, D.J.; Derner, J.D.; Milchunas, D.; Blumenthal, D.; Porensky, L.M. Grazing moderates increases in C3 grass abundance over seven decades across a soil texture gradient in shortgrass steppe. J. Veg. Sci. 2017, 28, 562–572. [Google Scholar] [CrossRef]
- Hart, R.H.; Ashby, M.M. Grazing intensities, vegetation, and heifer gains: 55 years on shortgrass. J. Range Manag. 1998, 51, 392–398. [Google Scholar] [CrossRef]
- Klipple, G.E.; Costello, D.F. Vegetation and cattle responses to different intensities of grazing on short-grass ranges on the Central Great Plains. Tech. Bull. US. Dep. Agric. 1960, N, 82. [Google Scholar]
- Wilmer, H.; Fernandez-Gimenez, M.E.; Derner, J.D.; Briske, D.D.; Augustine, D.J.; Porensky, L.M.; Tate, K.W.; Roche, L.M. Adaptive Grazing Management for Multiple Ecosystem Goods and Services: Does it Enhance Effective Decision-Making? 10th Int. Rangel. Congr. 2016, 1108. [Google Scholar]
- Didan, K. MOD13Q1 MODIS/Terra Vegetation Indices 16-Day L3 Global 250m SIN Grid V006 [Data set]. NASA EOSDIS LP DAAC 2015. Available online: doi:10.5067/MODIS/MOD13Q1.006 (accessed on 5 July 2018).
- Didan, K. MYD13Q1 MODIS/Aqua Vegetation Indices 16-Day L3 Global 250m SIN Grid V006 [Data set]. NASA EOSDIS LP DAAC 2015. Available online: doi:10.5067/MODIS/MYD13Q1.006 (accessed on 5 July 2018).
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Gao, F.; Hilker, T.; Zhu, X.; Anderson, M.; Masek, J.; Wang, P.; Yang, Y. Fusing Landsat and MODIS Data for Vegetation Monitoring. IEEE Geosci. Remote Sens. Mag. 2015, 3, 47–60. [Google Scholar] [CrossRef]
- Gao, F.; Masek, J.; Schwaller, M.; Hall, F. On the blending of the Landsat and MODIS surface reflectance: predicting daily Landsat surface reflectance. IEEE Trans. Geosci. Remote Sens. 2006, 44, 2207–2218. [Google Scholar] [CrossRef]
- Caride, C.; Piñeiro, G.; Paruelo, J.M. How does agricultural management modify ecosystem services in the argentine Pampas? The effects on soil C dynamics. Agric. Ecosyst. Environ. 2012, 154, 23–33. [Google Scholar] [CrossRef]
- Verón, S.R.; Oesterheld, M.; Paruelo, J.M. Production as a Function of Resource Availability: Slopes and Efficiencies Are Different. J. Veg. Sci. 2005, 16, 351–354. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. R Core Team nlme: Linear and Nonlinear Mixed Effects Models. Available online: https://cran.r-project.org/web/packages/nlme/index.html (accessed on 5 July 2018).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Reed, B.C.; Brown, J.F.; VanderZee, D.; Loveland, T.R.; Merchant, J.W.; Ohlen, D.O. Measuring phenological variability from satellite imagery. J. Veg. Sci. 1994, 5, 703–714. [Google Scholar] [CrossRef]
- White, M.A.; Nemani, R.R.; Thornton, P.E.; Running, S.W. Satellite Evidence of Phenological Differences Between Urbanized and Rural Areas of the Eastern United States Deciduous Broadleaf Forest. Ecosystems 2002, 5, 260–273. [Google Scholar] [CrossRef]
- GEOGLAM RAPP Rangeland and Pasture Productivity. Available online: http://www.geo-rapp.org/ (accessed on 24 June 2018).
- Charles, G.K.; Porensky, L.M.; Riginos, C.; Veblen, K.E.; Young, T.P. Herbivore effects on productivity vary by guild: cattle increase mean productivity while wildlife reduce variability. Ecol. Appl. 2017, 27, 143–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derner, J.D.; Hess, B.W.; Olson, R.A.; Schuman, G.E. Functional Group and Species Responses to Precipitation in Three Semi-Arid Rangeland Ecosystems. Arid Land Res. Manag. 2008, 22, 81–92. [Google Scholar] [CrossRef]
- Albrizio, R.; Steduto, P. Resource use efficiency of field-grown sunflower, sorghum, wheat and chickpea. Agric. For. Meteorol. 2005, 130, 254–268. [Google Scholar] [CrossRef]
- Blanco, L.J.; Paruelo, J.M.; Oesterheld, M.; Biurrun, F.N. Spatial and temporal patterns of herbaceous primary production in semi-arid shrublands: A remote sensing approach. J. Veg. Sci. 2016, 27, 716–727. [Google Scholar] [CrossRef]
- Fuhlendorf, S.D.; Harrell, W.C.; Engle, D.M.; Hamilton, R.G.; Davis, C.A.; Leslie, D.M. Should Heterogeneity be the Basis for Conservation? Grassland Bird Response to Fire and Grazing. Ecol. Appl. 2006, 16, 1706–1716. [Google Scholar] [CrossRef]
- Petrie, M.D.; Peters, D.P.C.; Yao, J.; Blair, J.M.; Burruss, N.D.; Collins, S.L.; Derner, J.D.; Gherardi, L.A.; Hendrickson, J.R.; Sala, O.E.; et al. Regional grassland productivity responses to precipitation during multiyear above- and below-average rainfall periods. Glob. Change Biol. 2018, 24, 1935–1951. [Google Scholar] [CrossRef] [PubMed]
- Sellers, P.J. Canopy reflectance, photosynthesis and transpiration. Int. J. Remote Sens. 1985, 6, 1335–1372. [Google Scholar] [CrossRef] [Green Version]
- Asner, G.P. Biophysical and Biochemical Sources of Variability in Canopy Reflectance. Remote Sens. Environ. 1998, 64, 234–253. [Google Scholar] [CrossRef]
- van Leeuwen, W.J.D.; Huete, A.R. Effects of standing litter on the biophysical interpretation of plant canopies with spectral indices. Remote Sens. Environ. 1996, 55, 123–138. [Google Scholar] [CrossRef]
- Phenocam. Available online: https://phenocam.sr.unh.edu/webcam/ (accessed on 24 June 2018).










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | R2 | Slope | Intercept | Type | Dataset | Model | R2 | Slope | Intercept | Type | Dataset | Model | R2 | Slope | Intercept | Type | Dataset | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2003 | 0.76 | 0.70 | 48.75 | sp | LTGI | 15E P3 | 0.01 | −0.07 | 80.14 | temp | CARM | 25SE P1 | 0.50 | 1.14 | 19.17 | temp | CARM | ||
| 2004 | 1.00 | 2.76 | −33.96 | sp | LTGI | 15E P4 | 0.01 | −0.03 | 93.45 | temp | CARM | 25SE P2 | 0.94 | 2.01 | −38.79 | temp | CARM | ||
| 2005 | 0.95 | 3.01 | −170.62 | sp | LTGI | 17N P1 | 0.34 | 3.39 | −100.82 | temp | CARM | 25SE P3 | 0.81 | 1.56 | 7.58 | temp | CARM | ||
| 2006 | 1.00 | 4.23 | −149.40 | sp | LTGI | 17N P2 | 0.68 | 1.50 | −9.24 | temp | CARM | 25SE P4 | 0.81 | 0.92 | 29.02 | temp | CARM | ||
| 2007 | 0.98 | 2.73 | −66.00 | sp | LTGI | 17N P3 | 0.69 | 1.78 | 45.12 | temp | CARM | 26E P1 | 0.87 | 2.03 | −41.87 | temp | CARM | ||
| 2008 | 0.14 | 1.64 | −13.53 | sp | LTGI | 17N P4 | 1.00 | 2.08 | 0.50 | temp | CARM | 26E P2 | 0.94 | 1.29 | −12.66 | temp | CARM | ||
| 2009 | 0.98 | 5.98 | −505.20 | sp | LTGI | 17N P5 | 0.47 | 1.12 | 13.27 | temp | CARM | 26E P3 | 1.00 | 1.27 | −3.34 | temp | CARM | ||
| 2010 | 1.00 | 10.34 | −769.89 | sp | LTGI | 17N P6 | 0.91 | 2.17 | −39.75 | temp | CARM | 26E P4 | 0.43 | 1.77 | −33.68 | temp | CARM | ||
| 2011 | 0.91 | 2.00 | −35.98 | sp | LTGI | 19N P1 | 0.73 | 1.48 | 38.26 | temp | CARM | 31E P1 | 0.60 | 1.28 | −4.22 | temp | CARM | ||
| 2012 | 0.72 | 5.65 | −160.24 | sp | LTGI | 19N P2 | 0.27 | 0.64 | 72.57 | temp | CARM | 31E P2 | 0.23 | 0.86 | 33.23 | temp | CARM | ||
| 2013 | 0.80 | 5.54 | −181.35 | sp | LTGI | 19N P3 | 0.75 | 1.08 | 14.18 | temp | CARM | 31E P3 | 0.30 | 1.27 | 13.01 | temp | CARM | ||
| 2014 | 0.89 | 2.92 | −113.06 | sp | LTGI | 19N P4 | 0.23 | 0.67 | 72.36 | temp | CARM | 31E P4 | 0.42 | 1.44 | −21.30 | temp | CARM | ||
| 2015 | 0.37 | 2.06 | −9.93 | sp | LTGI | 20SE P1 | 0.97 | 1.87 | 21.17 | temp | CARM | 5E P1 | 0.00 | −0.07 | 109.40 | temp | CARM | ||
| 2016 | 0.42 | 1.29 | 39.91 | sp | LTGI | 20SE P2 | 0.97 | 1.04 | 86.10 | temp | CARM | 5E P2 | 0.40 | −3.30 | 330.10 | temp | CARM | ||
| 23E (heavy) | 0.87 | 1.39 | 4.63 | temp | LTGI | 20SE P3 | 0.35 | 0.84 | 69.29 | temp | CARM | 5E P3 | 0.13 | −1.71 | 242.81 | temp | CARM | ||
| 23W (light) | 0.84 | 1.87 | −11.60 | temp | LTGI | 20SE P4 | 0.61 | 1.24 | 51.89 | temp | CARM | 5E P4 | 0.11 | 0.35 | 84.09 | temp | CARM | ||
| 15E (moderate) | 0.86 | 1.95 | −23.92 | temp | LTGI | 20SE P5 | 0.13 | 0.38 | 115.12 | temp | CARM | 7NW P1 | 0.33 | 0.56 | 76.13 | temp | CARM | ||
| 2013 | 0.49 | 1.83 | −24.19 | sp | CARM | 20SE P6 | 0.77 | 1.46 | 9.28 | temp | CARM | 7NW P2 | 0.58 | 0.89 | 53.24 | temp | CARM | ||
| 2014 | 0.48 | 1.94 | −58.15 | sp | CARM | 24W P1 | 0.08 | 0.77 | 42.38 | temp | CARM | 7NW P3 | 0.43 | 0.73 | 64.69 | temp | CARM | ||
| 2015 | 0.47 | 1.80 | 9.76 | sp | CARM | 24W P2 | 0.00 | −0.05 | 85.75 | temp | CARM | 7NW P4 | 0.88 | 1.72 | −20.76 | temp | CARM | ||
| 2016 | 0.12 | 2.71 | −53.30 | sp | CARM | 24W P3 | 0.62 | 0.74 | 33.82 | temp | CARM | 7NW P5 | 0.13 | 1.00 | 65.64 | temp | CARM | ||
| 15E P1 | 0.54 | 0.85 | 35.52 | temp | CARM | 24W P4 | 0.05 | 0.31 | 77.34 | temp | CARM | 7NW P6 | 0.00 | 0.01 | 111.12 | temp | CARM | ||
| 15E P2 | 0.95 | 1.05 | 17.53 | temp | CARM |
| LTGI | CARM | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Term | Type | Estimate | Std Error | T-Value | P-value | Term | Type | Estimate | Std Error | T-Value | P-value | |
| Intercept | - | −59.75 | 55.99 | −1.07 | 0.2939 | Intercept | - | 25.52 | 4.32 | 5.90 | 0.0000 | |
| AG | C4 Short | 4.81 | 1.45 | 3.32 | 0.0022 | AG | C4 Short | 0.52 | 0.11 | 4.53 | 0.0000 | |
| BOBU | C4 Short | 6.28 | 1.09 | 5.76 | 0.0000 | BOBU | C4 Short | 0.48 | 0.05 | 8.88 | 0.0000 | |
| C3PG | C3 Tall | 7.15 | 2.95 | 2.42 | 0.0213 | C3PG | C3 Tall | 0.32 | 0.03 | 12.03 | 0.0000 | |
| FORB | C3 Tall | 4.81 | 1.44 | 3.35 | 0.0021 | FORB | C3 Tall | 0.27 | 0.10 | 2.75 | 0.0064 | |
| OC4PG | C4 Short | 12.10 | 3.61 | 3.35 | 0.0021 | OC4PG | C4 Short | 0.44 | 0.06 | 7.49 | 0.0000 | |
| SD | - | −3.60 | 2.26 | −1.60 | 0.1203 | SD | - | −0.14 | 0.05 | −2.94 | 0.0036 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaffney, R.; Porensky, L.M.; Gao, F.; Irisarri, J.G.; Durante, M.; Derner, J.D.; Augustine, D.J. Using APAR to Predict Aboveground Plant Productivity in Semi-Arid Rangelands: Spatial and Temporal Relationships Differ. Remote Sens. 2018, 10, 1474. https://0-doi-org.brum.beds.ac.uk/10.3390/rs10091474
Gaffney R, Porensky LM, Gao F, Irisarri JG, Durante M, Derner JD, Augustine DJ. Using APAR to Predict Aboveground Plant Productivity in Semi-Arid Rangelands: Spatial and Temporal Relationships Differ. Remote Sensing. 2018; 10(9):1474. https://0-doi-org.brum.beds.ac.uk/10.3390/rs10091474
Chicago/Turabian StyleGaffney, Rowan, Lauren M. Porensky, Feng Gao, J. Gonzalo Irisarri, Martín Durante, Justin D. Derner, and David J. Augustine. 2018. "Using APAR to Predict Aboveground Plant Productivity in Semi-Arid Rangelands: Spatial and Temporal Relationships Differ" Remote Sensing 10, no. 9: 1474. https://0-doi-org.brum.beds.ac.uk/10.3390/rs10091474