Response of Beech (Fagus sylvatica L.) Trees to Competition—New Insights from Using Fractal Analysis


Silviculture and Forest Ecology of the Temperate Zones, Faculty of Forest Sciences, Georg-August-Universität Göttingen, Büsgenweg 1, 37077 Göttingen, Germany
*
Author to whom correspondence should be addressed.
Remote Sens. 2019, 11(22), 2656; https://0-doi-org.brum.beds.ac.uk/10.3390/rs11222656
Submission received: 15 October 2019
/
Revised: 7 November 2019
/
Accepted: 12 November 2019
/
Published: 13 November 2019
(This article belongs to the Special Issue 3D Forest Structure Observation)
Abstract
:Individual tree architecture and the composition of tree species play a vital role for many ecosystem functions and services provided by a forest, such as timber value, habitat diversity, and ecosystem resilience. However, knowledge is limited when it comes to understanding how tree architecture changes in response to competition. Using 3D-laser scanning data from the German Biodiversity Exploratories, we investigated the detailed three-dimensional architecture of 24 beech (Fagus sylvatica L.) trees that grew under different levels of competition pressure. We created detailed quantitative structure models (QSMs) for all study trees to describe their branching architecture. Furthermore, structural complexity and architectural self-similarity were measured using the box-dimension approach from fractal analysis. Relating these measures to the strength of competition, the trees are exposed to reveal strong responses for a wide range of tree architectural measures indicating that competition strongly changes the branching architecture of trees. The strongest response to competition (rho = −0.78) was observed for a new measure introduced here, the intercept of the regression used to determine the box-dimension. This measure was discovered as an integrating descriptor of the size of the complexity-bearing part of the tree, namely the crown, and proven to be even more sensitive to competition than the box-dimension itself. Future studies may use fractal analysis to investigate and quantify the response of tree individuals to competition.

1. Introduction
In Europe, the conversion of pure stands into mixed stands is still ongoing [1,2,3]. The corresponding silvicultural interventions are made under the assumption that they result in ecologically and economically beneficial stands [4,5]. However, site conditions, competition processes and ontogenetic stage all affect the growth of an individual tree, be it in a mixed or pure stand [6,7]. Among the mentioned factors, competition is the only one that can be cost-efficiently influenced through silviculture, and it is therefore of special importance [8]. In mixed stands, the growth response of the individual tree depends on the species identity of the surrounding neighbors [9,10,11]. Accordingly, in order to understand and model the dynamics in mixed stands, it is necessary to look into the development of tree morphological characteristics and branching architecture under competition in mixed neighborhoods.
According to earlier research [12,13,14], there are two ways to quantify an individual tree’s competition, namely distance independent and distance dependent approaches. In distant-independent approaches, stand density measures are used to estimate the competition pressure of an individual tree, whereas the distant-dependent approaches use the competitive effect determined by the size, position and number of the neighbors [15]. Distant-dependent indices are particularly popular and they are therefore known to offer a reliable prognosis of single-tree growth [13,16]. Lately, new, spatially explicit competition indices that are based on 3D stand models from terrestrial laser scanning (TLS) have also been introduced and showed large potential to explain tree growth and tree shape in response to competition [17,18,19,20,21]. TLS is being increasingly used to measure and record high quality tree individual parameters such as stem volume and crown structure with high accuracy [22,23,24].
Despite these advancements, there was limited research focusing on the effects of competition on distinct tree characteristics within a species. The main reason was that a comprehensive measurement of branching pattern and detailed tree architecture was almost impossible based on conventional methods of forest inventory [25]. TLS, however, not only provides solid measures of competition [18,20], as mentioned above but also millimeter-level detail on the tree structure and ultimately, a rapid assessment of each individual tree in a stand [26].
Thus far, only a few recent pioneering studies used the technology to investigate responses of the branching architecture to competition [20,27,28]. Oliver et al. [20] used TLS data to overcome the limits of traditional canopy studies, when it comes to quantifying differences in tree crowns. For sugar maple (Acer saccharum, Marsh.) stands with different compositions and developmental stages, they found TLS-based competition indices to be better predictors of crown metric variability than stand type, also highlighting the potential of TLS data to quantify tree competition and space occupancy.
Juchheim et al. [28] used TLS data to calculate structural measures for European beech (Fagus sylvatica L.) trees that grew either in intra- or interspecific neighborhoods in unmanaged stands and their findings provide evidence of different phenotypic responses of European beech, as a consequence of different types of competition.
Again, through application of a TLS and a new point cloud skeletonization approach, Bayer et al. [27] determined structural crown properties of European beech trees and Norway spruce growing in mixed and pure stands. The results yielded detailed information on the morphological traits of the trees and revealed that different competitor species results in significantly different crown structures of the study trees.
Recently, the toolset for the analysis of tree architecture was further extended by methods of fractal analysis, for example the box dimension (abbreviation Db; cf. [21]). When fractal analysis became famous in the 1970s, the box-dimension was introduced by the mathematician Mandelbrot [29]. It is a holistic approach to the architectural complexity of trees [21] that is rested on observing the change in the number of virtual boxes one needs to enclose all parts of an object (its 3D model) in dependence of the size of the boxes one uses. Based on the relationship (regression line) between the number of boxes (y-axis) and their size (x-axis), fractal analysis allows conclusions to be drawn on the complexity of an object, its dimensions, and its geometrical self-similarity. Self-similarity can be understood as a measure of geometrical repetition across scales, meaning that similar architectural pattern, for example a y-shaped branch bifurcation occurs across several scales. Therefore, starting with the first bifurcation of the stem, the two major branches would bifurcate again in the same way, each into two more branches and with the same y-shape. This process would repeat down to the smallest twigs. Such trees can easily be created from computer models such as the famous Lindenmayer l-system [30]. Real plants however, deviate from perfect self-similarity to various degrees due to external or internal factors.
When it comes to the dimensional aspects of tree architecture, the intercept of the regression line for the relationship between number of boxes used to calculate the box-dimension and their size (see previous paragraph) was predicted to be a useful measure in an earlier study [31]
Using fractal analysis to address tree architecture bears great potential for the investigation of competition–architecture relationships. According to Seidel et al. [31], the box-dimension approach addresses a large number of architectural characteristics at once, including the tree crown lengths, crown diameter, branch angles, and others. In the same study, it was also shown that the branching patterns of the trees, together with the crown dimension, are solid surrogates for a tree structural complexity.
Here, we used the box-dimension, self-similarity derived from fractal analysis, as well as conventional topological measures of tree architecture to analyze the effect of competition on the shape of beech (Fagus sylvatica L.) trees.
We hypothesized that competition strength affects the architecture of beech trees. More precisely, we hypothesized that competition affects (i) branching pattern, as well as (ii) measures of tree complexity derived from fractal analysis. We will explore the potential and meaning of measures from fractal analysis in the hope to gain new insights into competition–architecture relationships.
2. Materials and Methods
2.1. Study Sites and Objects
Our data were taken in the framework of the Biodiversity Exploratories [32]. Covering a gradient in precipitation and temperature across Germany, the three Exploratories were located in the Swabian Alb (South-West Germany), the Hainich–Dün (Central Germany) and the Schorfheide-Chorin (North-East Germany). The location of the study sites and the study design are shown in Figure 1.
In each area, we investigated the architecture of eight adult beech (Fagus sylvatica L.) trees (total = 3 × 8 = 24) that grow in the center of a quadrat. Each quadrat was defined by four competitor trees growing in the corners of the quadrat cornering the respective study tree in the center. In an earlier study, these tree quadrats were chosen, as they differed in species composition and hence competitive neighborhood of the subject beech trees [17]. The quadrats were intended to resemble a five-on-a-dice-like layout with the four competitor trees growing in the corners. Competitor trees comprised Scots pine (Pinus syvestris L.), Norway spruce (Picea abies L.), Sycamore maple (Acer pseudoplatanus L.), European ash (Fraxinus excelsior L.), oak (Quercus spp.), small-leaved lime (Tilia cordata Mill.), and hornbeam (Carpinus betulus L.). Due to some natural variation in distances, small deviations from the ideal spatial layout occurred. Details on the study sites can be found in Metz et al. [17], including climatic and soil-related properties. A summary of the main characteristics of the study trees is provided in Table 1. While Metz et al. [17] focused on the effect of competition on diameter growth, here we examine the effect of competition on crown structural characteristics.
2.2. Laser Scanning
Each study tree was scanned using a Zoller and Fröhlich Imager 5006 terrestrial laser scanner (Zoller and Fröhlich GmbH, Wangen i.A, Germany). The instrument was set to scan with a resolution of 0.036 degrees using a field of view of 310° (vertically) and 360° (horizontally). The maximum range of the instrument was 179 m and we conducted six or more scans (depending on stand density and visibility) surrounding the study trees and their immediate neighbors. All scans were conducted in March 2012, in leafless conditions to ensure free sight on the upper crowns [17]. We used 24 artificial chessboard targets that were distributed in the scanned scene as tie points for spatial co-registration of the individual scans. Scans were merged using Zoller and Fröhlich Laser Control 8.2 software (Zoller and Froehlich GmbH). As a result of the laser scanning procedure, each study trees and the surrounding forest area were available as virtual three-dimensional high-resolution (3D) point cloud representations of the real world.
2.3. Point Cloud Processing and Quantitative Structure Models
Each point cloud of a study tree and its surroundings was used to manually separate the study trees from the remaining forest. This was conducted manually using Leica Cyclone Software (Leica Geosysteme, Heerbrugg, Switzerland). After the study trees were available as single-tree point clouds (see Figure 2, left), they were processed using the CompuTree software [33] to create a quantitative structure model (abbreviation QSM; see Figure 2, right) for each individual. A QSM-model can be understood as a representation of the tree point cloud based on cylinders of various diameters and lengths [33]. We decided to use the same QSM-parameter settings for all trees in order to allow for a better reproducibility of our results. The parameters were 0.10 m for clustering tolerance, a maximum of 600-point clusters with at least 400 points each (automatically adjusted by the software if less clusters are found). Each cluster needed to contain at least 0.5% of all points in order to be created.
We decided for this set of parameters based on visual inspection of the resulting QSM models with the point clouds overlaid for quality assessment. We are not aware of any method that enables a less subjective quality control for QSM models. However, we argue, that a set of parameters used for all trees shall produce an objective and reproducible QSM model for our study trees.
Here, we applied QSM-models in order to obtain information on the branching pattern of the beech trees for branches up to the 3rd order. For all 24 study trees, the branching patterns were calculated using the same settings for the CompuTree software. These settings can theoretically be adapted to optimize the modeling for trees of different species, varying height, etc., but were kept the same to ensure repeatability in our study focusing on trees of only one species and of similar dimensions. We derived the mean branch volume of 1st, 2nd, and 3rd order branches, mean branch angle of 1st, 2nd, and 3rd order branches, range of branch angle of 1st, 2nd, and 3rd order branches, total branch length and mean branch length. In addition to these eleven measures, we also tested three measures obtained from fractal analysis. Those are explained in the following.
2.4. Box-dimension, Intercept and Self-Similarity
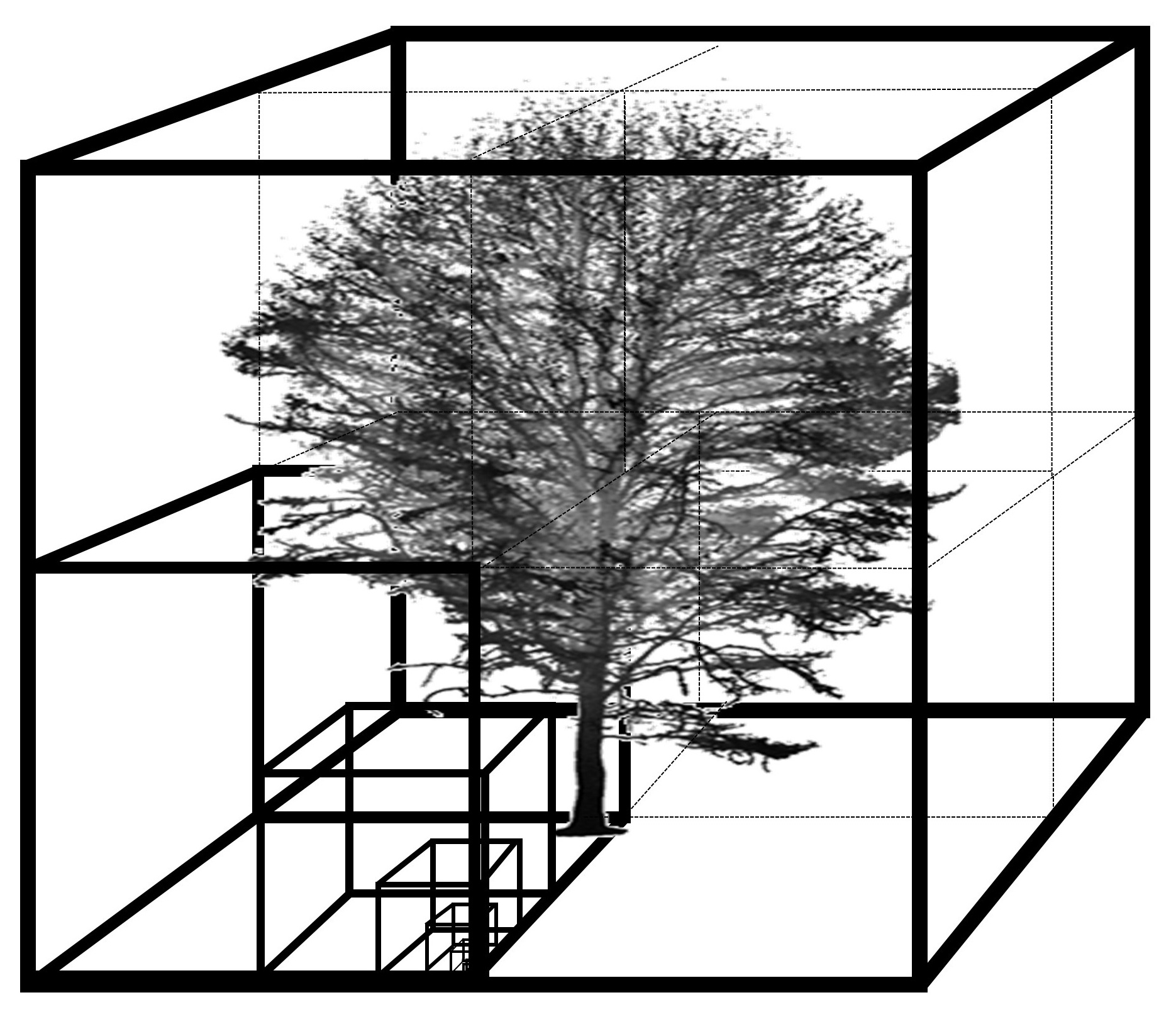
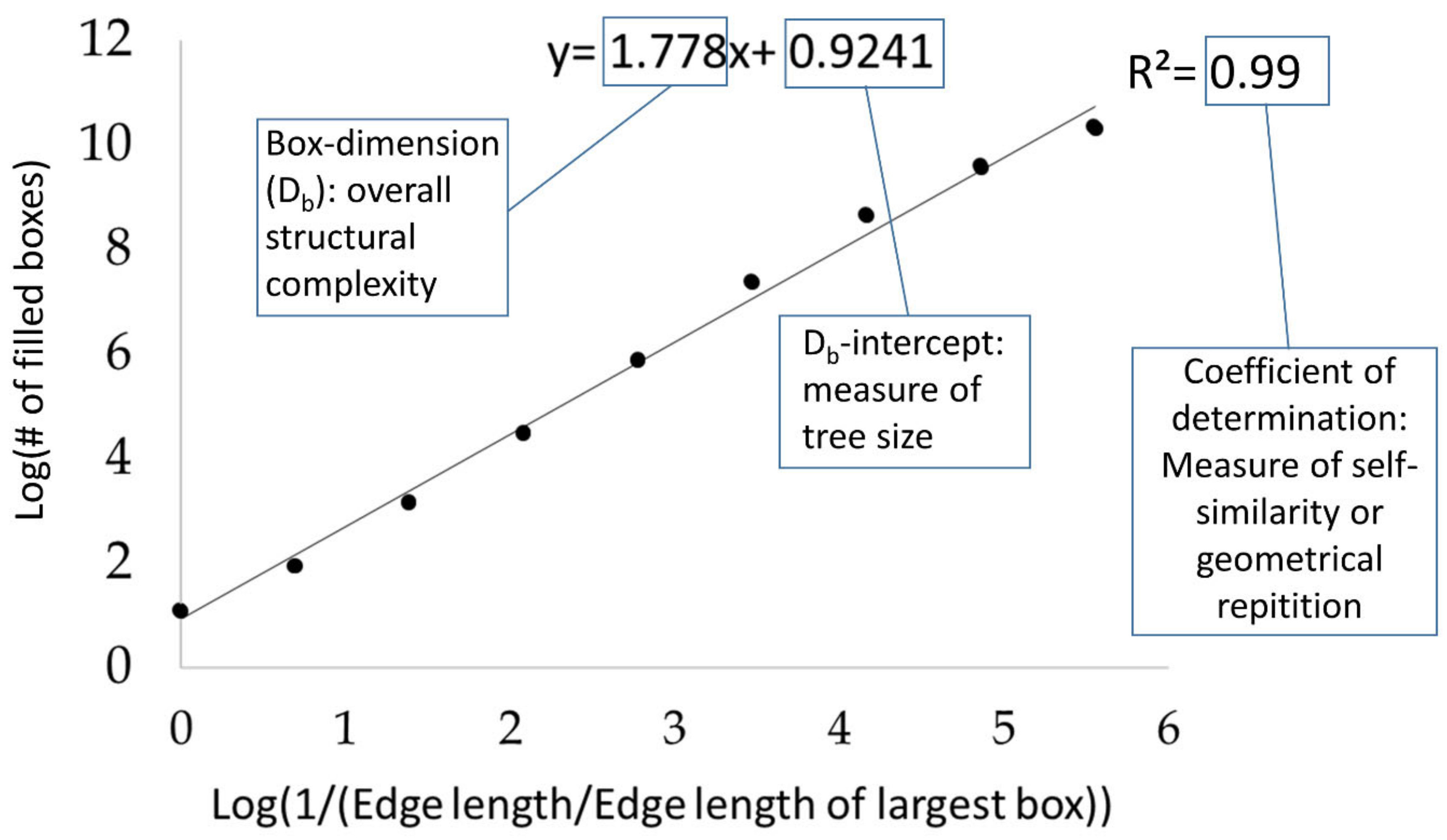
The box-dimension (Db) can be considered a measure of a tree’s architectural complexity. It was calculated from the single-tree point clouds following the approach introduced in Seidel [21]. This approach rests on the ideas of Sarkar and Chaudhuri [34] and the ground-breaking work of Mandelbrot [29]. In short, we determined Db by counting the number of virtual boxes of a given size needed to enclose all aboveground tree organs in the point cloud. This number of boxes is determined for different box-sizes, starting with the largest box, which is the minimum bounding box enclosing the full tree, and continuing with successively smaller boxes always being 1/8th the volume of the previous box size (or 1/2 the edge length). Then, a least-square line was fit through the scatterplot of the number of boxes needed (dependent variable) over the inverse of the box-size used (given in relation to the largest box used; explanatory variable). The slope of this line is defined as the Db, its coefficient of determination (R²) is defined as a measure of self-similarity of the tree architecture across the different tested scales (box-sizes) and the intercept of the regression line (Db-intercept) with the y-axis is a surrogate for trees size [31] (Figure 3 and Figure 4). Here, we will explore all three variables for their responsiveness to competition. First, we evaluated the intercept for correlation with established measures of tree size and correlation with competition strength. Since Mandelbrot [29] stated that the intercept is a measure of object dimension (size), we tested whether Db-intercept responded to competition strength and whether it was related more to conventional measures of tree size (total tree height [TTH]), diameter at breast height (DBH), or to crown-related measure of dimension (crown volume and crown radius). Then, in addition to Db as a dimensionless measure of tree architectural complexity, we evaluated whether self-similarity (R² of the regression) as a new architectural attribute may respond to competition.
2.5. Calculation of Competitive Pressure
Competition strength enforced on the study trees was determined using the cumulative crown surface area (CCSA) of the competitor trees. For the trees investigated in our study, Metz et al. [17] showed that CCSA is a good predictor of the competitive situation the trees were facing, thereby explaining tree growth (R² = 0.34; cf. [17]). The crown surface area of each competitor tree was determined using the convex hull polygon approach and the total for the four competitors was calculated per study tree. Using an algorithm written in Mathematica (Wolfram Research, Champaign, USA), we determined the surface of the convex hull polygons from the triangle points building the convex hull by applying Heron’s formula.
2.6. Statistical Analysis
Using the statistical software R [35], we tested for relationships between all structural attributes and competition strength using correlation analysis (Spearman’ rank rho). Level of significance was < 0.05 in all tests. The within data trend of selected significant correlations was further analyzed using non-linear Generalized Additive Modeling (GAM) techniques because assumptions for linear regressions were not met. Also, to avoid over-fitting, the effective degrees of freedom (EDF) were set to 3 with smoothing then chosen automatically through cross-validation [36]. Models evaluation occurred through interpreting the EDF value as complexity of the smoothing function, p-values of the smoothing function, and the deviance explained by the GAM. The EDF values were all one, which means the within data trend suggests linearity (comp. results and discussion).
3. Results
Out of the 14 tested structural attributes, five were significantly correlated to competition strength (Table 2).
We found significant correlations between competition strength and the following measures; total branch length 1st order (rho: −0.53; p < 0.01), mean branch length 1st order (rho: –0.52; p < 0.01), and the range of branch angles 2nd order (rho: −0.62; p < 0.01) (Table 2).
Competition not only reduced branch lengths, but it also reduced the variability in branching angles (Figure 5A,B) and it reduced the box-dimension of the trees (Figure 5C).
The intercept of the Db-regression line was the attribute that was most sensitive to competition strength and the relationship was again negative (rho: −0.78; p < 0.001), with the GAM also resulting in the highest explained deviance of all tested measures (Figure 5D and Table 2).
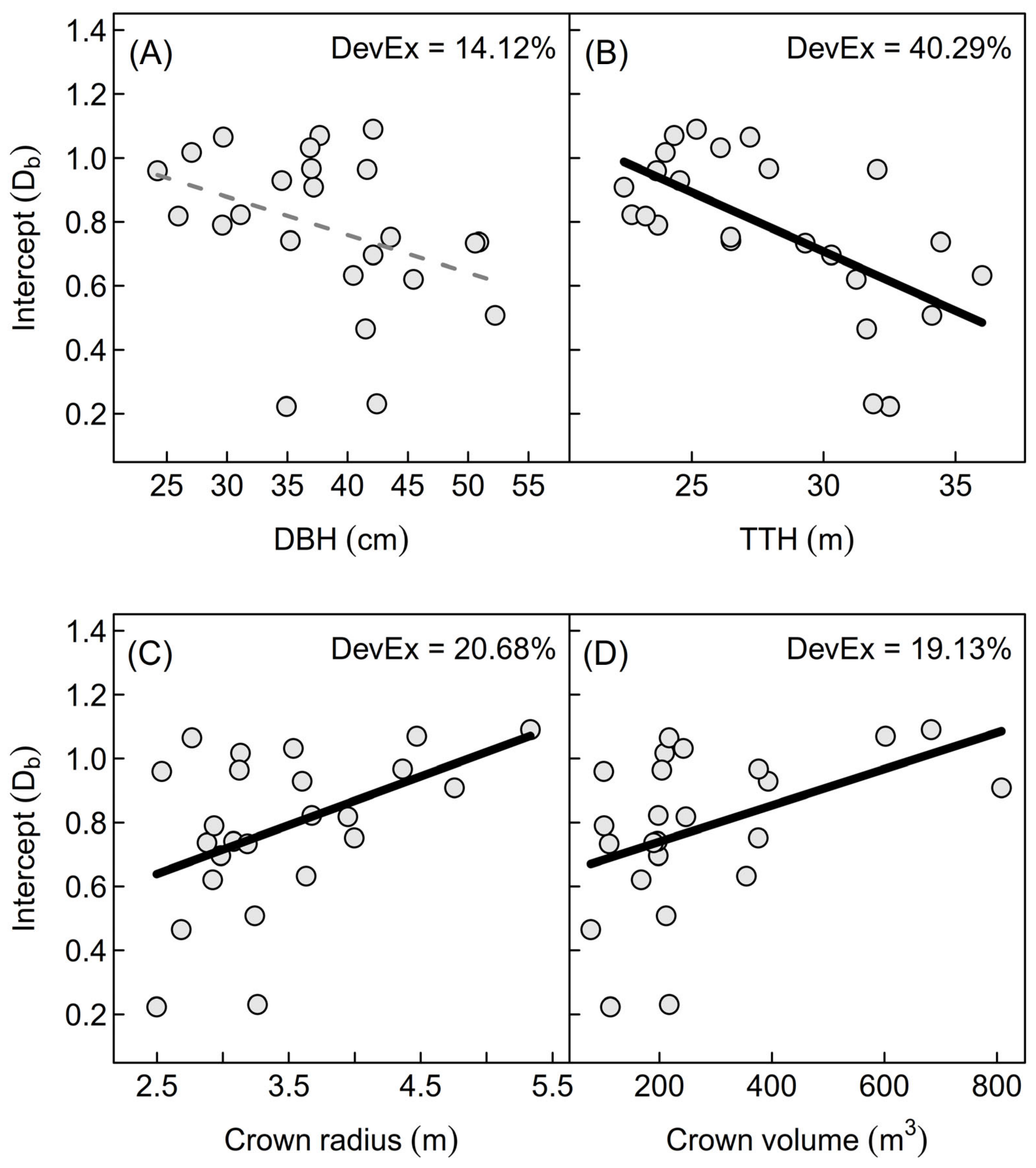
We also discovered positive correlations between the Db-regression and crown morphological variables, like crown radius (rho: 0.42; p < 0.05) and crown volume (rho: 0.50; p < 0.05) (comp. Figure 6C,D). The two conventional measures of tree size, diameter at breast height (rho: −0.44; p < 0.05) and total tree height (rho: −0.59; p < 0.05) were, however, negatively correlated with Db-intercept (Figure 6A,B).
The third measure derived from fractal analysis, namely self-similarity, showed no significant correlation with competition strength (rho: 0.31; p = 0.142).
4. Discussion
We investigated whether the branching architecture of beech trees responds to competition strength, as was indicated in earlier studies on European beech [27,28]. We hypothesized that competition strength affects the architecture of beech trees in terms of branching pattern, as already indicated by Bayer et al. [27], as well as measures of tree structural complexity derived from fractal analysis. Our results strongly support this assumption, as we identified a significant response of branch length, branch angles as well as the box-dimension and Db-intercept of the target trees to competition enforced on them by the neighboring trees.
Not all tree species may be as responsive to competition strength as beech. Its high adaptive capacity is clearly expressed in its phenotypic plasticity and was reported repeatedly in the literature [37,38]. For example, the study by Bayer et al. [27] showed first results on changes in the branching angles of beech in dependence of the species of the neighboring trees. Similarly, Juchheim et al. [28] showed that the structure of the crown of European beech, assessed using QSM-model like in our present study, significantly varied depending on competition type (intra vs. interspecific competition). Our research added knowledge on the general response of beech architecture to competition strength (irrespective of competition type), and most importantly, that the entire structural complexity of the trees is negatively affected by competition.
Together with the crown size, the distribution of the branches and leaves is an important aspect of productivity [39,40] and it is therefore not surprising that competition effects directly relate to the difference in a tree’s productivity, as already shown for our study trees by Metz et al. [17]. Earlier studies using fractal analysis also confirmed that a reduction in architectural complexity comes at the cost of productivity [21]. It is also known that structural complexity, assessed as box-dimension, translates well to the growing efficiency of a tree [41]. Summarizing, competition enforced on a tree results in adaptations of the tree’s growth pattern (branching pattern), which in turn results in lower Db-values and consequently in a lower growth efficiency.
Using only crown shapes and dimensions to evaluate the effects of competition strength may lead to false conclusions, as changes in the branching architecture may result in changes in the light transmissivity of tree crowns and consequently forest canopies [42]. Those are not accounted for when only the dimensional extent of a crown (e.g., crown radius) is considered. Here, fractal analysis provides summarizing measures that address the tree’s architecture more comprehensively (branches and stem architecture together) and that should respond to competition as well. In fact, we found that the box-dimension and Db-intercept showed a strong negative correlation with competition strength (see Table 2 and Figure 5C,D), indicating that competitive pressure indeed reduces the structural complexity of beech trees. This supports the findings of an earlier study focusing on European beech [41].
We argue that the intercept of the Db-regression line is actually a proxy for the dimension of the complexity-bearing part of a tree, more precisely the tree crown (comp. Figure 6). This was confirmed by the positive correlations with crown morphological variables, like crown radius and crown volume (comp. Figure 6C,D). The two conventional measures of tree size, diameter at breast height and total tree height were, however, negatively correlated with Db-intercept. Despite the significant correlation, the smoothing term of the GAM model for the regression between DBH and Db-intercept was not significant (Figure 6A). This indicates that the DBH per se is not necessarily a complexity-bearing measure for a tree, meaning that trees with the same DBH can be of greatly varying complexity in architecture. Surprisingly, the Db-intercept decreased with increasing total tree height (Figure 6B). Actually, a similar result as described for the DBH would have been expected, namely that height alone is not related to complexity, but if at all one would have expected a positive relationship between the two variables. The only explanation for the negative trend is that the higher intraspecific competition led to slightly higher TTH across the sites due to increased height growth. As increased height growth resulted in reduced canopy dimensions (horizontal extent), it also resulted in a lower complexity of the postulated complexity-bearing tree part (crown).
In the dataset used here, self-similarity could not be confirmed as a measure that responded to competition but one has to consider that this may be due to our small sample size. The rho-value of 0.31 (Table 2) points towards some correlation but it is too weak to draw conclusions from it. In a future study, we will address this measure in more detail.
The advent of TLS provides the avenue to measure details of the tree architecture thereby giving new insight into the effects of competition on tree morphology and branching pattern [43]. Such knowledge is crucial to understand structure–productivity relationships [27,41], but also to get a better understanding of structure-biodiversity relationships [44,45], as they all depend on the architecture of individual trees.
5. Conclusions
Our study based on high-resolution terrestrial laser scanning, delved further into the response of branching architecture and fractal characteristics of crown structures to competition. The objective was to provide a deeper understanding on how tree architecture changes in response to competition.
Using terrestrial laser scanning data of individual trees, we determined quantitative structure models to derive branching pattern and applied fractal analysis to describe the tree architectural complexity. We showed that crowns of European beech are highly responsive to competition, changing not only dimensions and branching pattern, as shown in early work, but also the entire structural complexity.
We discovered the intercept of the regression line of the box-dimension to be strongly related to competition pressure. We argue that this is because it integrates several dimensional measures of the tree crown that respond strongly to competition.
Future studies may use fractal analysis to investigate and quantify the response of tree individuals to competition, as we discovered a strong architectural response of beech trees to competition pressure.
Author Contributions
Conceptualization: Y.D., P.A., C.A., D.S.; methodology: Y.D., D.S.; software: D.S.; formal analysis: Y.D., D.S., P.A.; resources: D.S., C.A.; writing-original draft preparation: Y.D., D.S.; writing- review and editing: Y.D., P.A., C.A., D.S.; visualization: Y.D., D.S., P.A.; supervision: D.S., C.A., P.A.; project administration: D.S.; funding acquisition: D.S., C.A.
Funding
The German Research Foundation (DFG) is acknowledged for funding this research through grant SE2383/5-1 provided to Dominik Seidel. Part of this work was also supported by funds of the German Government´s Special Purpose Fund held at Landwirtschaftliche Rentenbank (844732).
Acknowledgments
We are grateful to Jeromé Metz who provided us the scan data of the trees. We also thank the managers of the three Exploratories, Kirsten Reichel-Jung, Swen Renner, Katrin Hartwich, Sonja Gockel, Kerstin Wiesner, and Martin Gorke for their work in maintaining the plot and project infrastructure; Christiane Fischer and Simone Pfeiffer for giving support through the central office, Michael Owonibi for managing the central data base, and Markus Fischer, Eduard Linsenmair, Dominik Hessenmöller, Jens Nieschulze, Daniel Prati, Ingo Schöning, François Buscot, Ernst-Detlef Schulze, Wolfgang W. Weisser and the late Elisabeth Kalko for their role in setting up the Biodiversity Exploratories project. We thank Falk Hänsel, Stephan Wöllauer, Frank Suschke, Mathias Groß, Martin Fellendorf and Thomas Nauss for operation and maintenance of meteorological stations in the research plots.
Conflicts of Interest
The authors declare no conflict of interest
References
- Von Lüpke, B.; Ammer, C.; Braciamacchie, M.; Brunner, A.; Ceitel, J.; Collet, C.; Deuleuze, C.; Di Placido, J.; Huss, J.; Jankovic, J.; et al. Silvicultural strategies for conversion. In Norway Spruce Conversion—Options and Consequences; European Forest Institute Research Report 18; Spiecker, H., Hansen, J., Klimo, E., Skovsgaard, J.P., Sterba, H., von Teuffel, K., Eds.; Brill: Leiden, The Netherlands; Boston, MA, USA, 2004; pp. 121–164. [Google Scholar]
- Filipescu, C.N.; Comeau, P.G. Competitive interactions between aspen and white spruce vary with stand age in boreal mixed woods. For. Ecol. Manag. 2007, 247, 175–184. [Google Scholar] [CrossRef]
- Ammer, C.; Bickel, E.; Kolling, C. Converting Norway spruce stands with beech-a review of arguments and techniques. Aust. J. For. Sci. 2008, 125, 3–26. [Google Scholar]
- Knoke, T.; Ammer, C.; Stimm, B.; Mosandl, R. Admixing broadleaved to coniferous tree species: a review on yield, ecological stability and economics. Eur. J. For. Res. 2008, 127, 89–101. [Google Scholar] [CrossRef]
- Juchheim, J.; Ammer, C.; Schall, P.; Seidel, D. Canopy space filling rather than conventional measures of structural diversity explains productivity of beech stands. For. Ecol. Manag. 2017, 395, 19–26. [Google Scholar] [CrossRef]
- Shi, H.; Zhang, L. Local analysis of tree competition and growth. For. Sci. 2003, 49, 938–955. [Google Scholar] [CrossRef]
- Ledermann, T. Evaluating the performance of semi-distance-independent competition indices in predicting the basal area growth of individual trees. Can. J. For. Res. 2010, 40, 796–805. [Google Scholar] [CrossRef]
- Ammer, C. Konkurrenzsteuerung–Anmerkungen zu einer Kernaufgabe des Waldbaus beim Aufbau vielfältiger Wälder. Eberswalder Forstl. Schriftenreihe 2008, 36, 21–26. [Google Scholar]
- Canham, C.D.; LePage, P.T.; Coates, K.D. A neighborhood analysis of canopy tree competition: effects of shading versus crowding. Can. J. For. Res. 2004, 34, 778–787. [Google Scholar] [CrossRef]
- Zhao, D.; Borders, B.; Wilson, M.; Rathbun, S.L. Modeling neighborhood effects on the growth and survival of individual trees in a natural temperate species rich-forest. Ecol. Model. 2006, 196, 90–102. [Google Scholar] [CrossRef]
- Pretzsch, H.; Schütze, G. Transgressive overyielding in mixed compared with pure stands of Norway spruce and European beech in Central Europe: evidence on stand level and explanation on individual tree level. Eur. J. For. Res. 2009, 128, 183–204. [Google Scholar] [CrossRef]
- Munro, D.D. Forest Growth Models-A Prognosis. In Growth Models for Tree and Stand Simulation, Proceedings of the Congress IUFRO S4.01-4, September 1974 Stockholm, Sweden; Fries, J., Ed.; Department of Forest Yield Research, Royal College of Forestry: Stockholm, Sweden, 1974; Volume 30, pp. 1–21. [Google Scholar]
- Biging, G.S.; Dobbertin, M. Evaluation of competition indices in individual tree growth models. For. Sci. 1995, 41, 360–377. [Google Scholar] [CrossRef]
- Porté, A.; Bartelink, H. Modelling mixed forest growth: a review of models for forest management. Ecol. Model. 2002, 150, 141–188. [Google Scholar] [CrossRef]
- Tomé, M.; Burkhart, H.E. Distance-dependent competition measures for predicting growth of individual trees. For. Sci. 1989, 35, 816–831. [Google Scholar] [CrossRef]
- Bachmann, M. Indizes zur Erfassung der Konkurrenz von Einzelbäumen: Methodische Untersuchung in Bergmischwäldern; Forstliche Forschungsberichte München 171; Schriftenreihe der Forstwissenschaftlichen Fakultät der Universität München und der Bayerischen Landesanstalt für Wald und Forstwirtschaft: München, Germany, 1998. [Google Scholar]
- Metz, J.; Seidel, D.; Scheffer, D.; Schulze, E.D.; Ammer, C. Crown modeling by terrestrial laser scanning as an approach to assess the effect of aboveground intra-and interspecific competition on tree growth. For. Ecol. Manag. 2013, 310, 275–288. [Google Scholar] [CrossRef]
- Seidel, D.; Hoffmann, N.; Ehbrecht, M.; Juchheim, J.; Ammer, C. How neighborhood affects tree diameter increment–New insights from terrestrial laser scanning and some methodical considerations. For. Ecol. Manag. 2015, 336, 119–128. [Google Scholar] [CrossRef]
- Seidel, D.; Schall, P.; Gille, M.; Ammer, C. Relationship between tree growth and physical dimensions of Fagus sylvatica crowns assessed from terrestrial laser scanning. IForest 2015, 8, 735–742. [Google Scholar] [CrossRef]
- Olivier, M.D.; Robert, S.; Fournier, R.A. Response of sugar maple (Acer saccharum, Marsh.) tree crown structure to competition in pure versus mixed stands. For. Ecol. Manag. 2016, 374, 20–32. [Google Scholar] [CrossRef]
- Seidel, D. A holistic approach to determine tree structural complexity based on laser scanning data and fractal analysis. Ecol. Evol. 2018, 8, 128–134. [Google Scholar] [CrossRef]
- Yu, X.; Liang, X.; Hyyppä, J.; Kankare, V.; Vastaranta, M.; Holopainen, M. Stem biomass estimation based on stem reconstruction from terrestrial laser scanning point clouds. Remote Sens. Lett. 2013, 4, 344–353. [Google Scholar] [CrossRef]
- Astrup, R.; Ducey, M.J.; Granhus, A.; Ritter, T.; von Lüpke, N. Approaches for estimating stand-level volume using terrestrial laser scanning in a single-scan mode. Can. J. For. Res. 2014, 44, 666–676. [Google Scholar] [CrossRef]
- Newnham, G.J.; Armston, J.D.; Calders, K.; Disney, M.I.; Lovell, J.L.; Schaaf, C.B.; Strahler, A.H.; Danson, F.M. Terrestrial laser scanning for plot-scale forest measurement. Curr. Forest. Rep. 2015, 1, 239–251. [Google Scholar] [CrossRef]
- Seidel, D.; Leuschner, C.; Müller, A.; Krause, B. Crown plasticity in mixed forests—Quantifying asymmetry as a measure of competition using terrestrial laser scanning. For. Ecol. Manag. 2011, 261, 2123–2132. [Google Scholar] [CrossRef]
- Liang, X.; Hyyppä, J.; Kaartinen, H.; Lehtomäki, M.; Pyörälä, J.; Pfeifer, N.; Holopainen, M.; Brolly, G.; Francesco, P.; Hackenberg, J.; et al. International benchmarking of terrestrial laser scanning approaches for forest inventories. ISPRS J. Photogramm. Remote Sens. 2018, 144, 137–179. [Google Scholar] [CrossRef]
- Bayer, D.; Seifert, S.; Pretzsch, H. Structural crown properties of Norway spruce (Picea abies [L.] Karst.) and European beech (Fagus sylvatica [L.]) in mixed versus pure stands revealed by terrestrial laser scanning. Trees 2013, 27, 1035–1047. [Google Scholar] [CrossRef]
- Juchheim, J.; Annighöfer, P.; Ammer, C.; Calders, K.; Raumonen, P.; Seidel, D. How management intensity and neighborhood composition affect the structure of beech (Fagus sylvatica L.) trees. Trees 2017, 31, 1723–1735. [Google Scholar] [CrossRef]
- Mandelbrot, B.B. The Fractal Geometry of Nature; W.H. Freeman Company: New York, NY, USA, 1977; p. 468. [Google Scholar]
- Prusinkiewicz, P.; Runions, A. Computational models of plant development and form. New Phytol. 2012, 193, 549–569. [Google Scholar] [CrossRef]
- Seidel, D.; Ehbrecht, M.; Dorji, Y.; Jambay, J.; Ammer, C.; Annighöfer, P.J. Identifying architectural characteristics that determine tree structural complexity. Trees 2019, 33, 911–919. [Google Scholar] [CrossRef]
- Fischer, M.; Bossdorf, O.; Gockel, S.; Hansel, F.; Hemp, A.; Hessenmoller, D.; Korte, G.; Nieschulze, J.; Pfeiffer, S.; Prati, D.; et al. Implementing large-scale and long-term functional biodiversity research: The Biodiversity Exploratories. Basic Appl. Ecol. 2010, 11, 473–485. [Google Scholar] [CrossRef]
- Piboule, A.; Krebs, M.; Esclatine, L.; Hervé, J.-C. Computree: a collaborative platform for use of terrestrial lidar in dendrometry. In Proceedings of the International IUFRO Conference MeMoWood, Nancy, France, 1–4 October 2013. [Google Scholar]
- Sarkar, N.; Chaudhuri, B.B. An efficient differential box-counting approach to compute fractal dimension of image. IEEE T. Syst. Man Cyb. 1994, 24, 115–120. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; p. 201. [Google Scholar]
- Wood, S.N. Generalized Additive Models: An Introduction with R, 2nd ed.; Taylor Francis: New York, NY, USA, 2017; p. 476. [Google Scholar] [CrossRef]
- Schröter, M.; Härdtle, W.; Oheimb, G. Crown plasticity and neighborhood interactions of European beech (Fagus sylvatica L.) in an old-growth forest. Eur. J. For. Res. 2012, 131, 787–798. [Google Scholar] [CrossRef]
- Valladares, F.; Gianoli, E.; Gómez, J.M. Ecological limits to plant phenotypic plasticity. New Phytol. 2007, 176, 749–763. [Google Scholar] [CrossRef] [PubMed]
- Maguire, D.A.; Bennett, W.S. Patterns in vertical distribution of foliage in young coastal Douglas-fir. Can. J. For. Res. 1996, 26, 1991–2005. [Google Scholar] [CrossRef]
- Jucker, T.; Bouriaud, O.; Coomes, D.A. Crown plasticity enables trees to optimize canopy packing in mixed-species forests. Funct. Ecol. 2015, 29, 1078–1086. [Google Scholar] [CrossRef] [Green Version]
- Seidel, D.; Annighöfer, P.; Stiers, M.; Zemp, C.D.; Burkardt, K.; Ehbrecht, M.; Ammer, C. How a measure of tree structural complexity relates to architectural benefit-to-cost ratio, light availability, and growth of trees. Ecol. Evol. 2019, 9, 7134–7142. [Google Scholar] [CrossRef] [Green Version]
- Oliver, C.D.; Larson, B.C. Forest Stand Dynamics: Updated Edition; John Wiley and Sons: New York, NY, USA, 1996; p. 520. [Google Scholar]
- Henning, J.G.; Radtke, P.J. Ground-based laser imaging for assessing three dimensional forest canopy structure. Photogramm. Eng. Remote Sens. 2006, 72, 1349–1358. [Google Scholar] [CrossRef]
- Bazzaz, F.A. Plant Species Diversity in Old-Field Successional Ecosystems in Southern Illinois. Ecology 1975, 56, 485–488. [Google Scholar] [CrossRef]
- Tews, J.; Brose, U.; Grimm, V.; Tielbörger, K.; Wichmann, M.C.; Schwager, M.; Jeltsch, F. Animal species diversity driven by habitat heterogeneity/diversity: the importance of keystone structures. J. Biogeogr. 2004, 31, 79–92. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Study areas within Germany (left side) and sampling design (right side).

Figure 2.
Example of a study tree point cloud (left), the corresponding quantitative structure model (middle) and a close-up of the quantitative structure model (right).
Figure 2.
Example of a study tree point cloud (left), the corresponding quantitative structure model (middle) and a close-up of the quantitative structure model (right).

Figure 3.
Exemplary three-dimensional tree point clouds with a high box-dimension (left: Db: 2.02) and a low box-dimension (right: Db: 1.50). Box-dimension is considered a measure of structural complexity.
Figure 3.
Exemplary three-dimensional tree point clouds with a high box-dimension (left: Db: 2.02) and a low box-dimension (right: Db: 1.50). Box-dimension is considered a measure of structural complexity.

Figure 4.
Explanatory graph on the calculation of the three tested measures from fractal analysis, namely box-dimension (Db), intercept of the regression line (Db-intercept) and the coefficient of determination of the regression line (self-similarity).
Figure 4.
Explanatory graph on the calculation of the three tested measures from fractal analysis, namely box-dimension (Db), intercept of the regression line (Db-intercept) and the coefficient of determination of the regression line (self-similarity).

Figure 5.
Scatterplots of architectural features (A) = total length of 1st order branches; (B) = range of 2nd order branch angles; (C) = box dimension (Db); (D) = intercept of box dimension) with GAM and explained deviance (DevEx), respectively, of beech trees in dependence of competition strength as cumulative crown surface area (CCSA [m²]). Black solid lines show significant GAM models, the effective degrees of freedom (EDF) for all models was 1, suggesting linear within-data relationships.
Figure 5.
Scatterplots of architectural features (A) = total length of 1st order branches; (B) = range of 2nd order branch angles; (C) = box dimension (Db); (D) = intercept of box dimension) with GAM and explained deviance (DevEx), respectively, of beech trees in dependence of competition strength as cumulative crown surface area (CCSA [m²]). Black solid lines show significant GAM models, the effective degrees of freedom (EDF) for all models was 1, suggesting linear within-data relationships.

Figure 6.
Scatterplots of Db-Intercept against conventional measures of tree size (A) = diameter at breast height (DBH (cm)); (B) = total tree height (TTH (m)) and crown morphology (C) = Crown radius (m); (D) = Crown volume (m³)) with GAM and explained deviance (DevEx), respectively, of beech trees. Black solid lines show significant GAM models, dashed grey line shows insignificant model. The effective degrees of freedom (EDF) for all models was 1, suggesting linear within data relationships.
Figure 6.
Scatterplots of Db-Intercept against conventional measures of tree size (A) = diameter at breast height (DBH (cm)); (B) = total tree height (TTH (m)) and crown morphology (C) = Crown radius (m); (D) = Crown volume (m³)) with GAM and explained deviance (DevEx), respectively, of beech trees. Black solid lines show significant GAM models, dashed grey line shows insignificant model. The effective degrees of freedom (EDF) for all models was 1, suggesting linear within data relationships.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Major characteristics of the 24 investigated beech trees (adapted from [17]). DBH = Diameter at breast height (1.3 m). HAI = Hainich Dün, SCH = Schorfheide Chorin, ALB = Swabian Alb.
Table 1.
Major characteristics of the 24 investigated beech trees (adapted from [17]). DBH = Diameter at breast height (1.3 m). HAI = Hainich Dün, SCH = Schorfheide Chorin, ALB = Swabian Alb.
| Target Tree ID | Exploratory | Height (m) | DBH (cm) | Competing Species |
|---|---|---|---|---|
| BE 1 | HAI | 32.03 | 41.60 | ash |
| BE 2 | HAI | 31.24 | 45.50 | beech |
| BE 3 | HAI | 34.44 | 50.40 | beech |
| BE 4 | HAI | 31.63 | 41.60 | beech |
| BE 5 | HAI | 31.88 | 42.70 | beech |
| BE 6 | HAI | 22.72 | 31.30 | maple, ash |
| BE 7 | HAI | 29.30 | 51.50 | maple, lime, oak, hornbeam |
| BE 8 | HAI | 23.72 | 30.30 | ash |
| BE 9 | SCH | 27.92 | 37.20 | pine |
| BE10 | SCH | 23.25 | 26.20 | pine |
| BE11 | SCH | 25.18 | 42.30 | pine |
| BE12 | SCH | 36.01 | 40.00 | beech |
| BE13 | SCH | 34.11 | 50.10 | beech |
| BE14 | SCH | 24.33 | 37.30 | pine |
| BE15 | SCH | 26.47 | 43.30 | beech |
| BE16 | SCH | 26.09 | 37.00 | beech |
| BE17 | ALB | 27.21 | 30.00 | beech |
| BE18 | ALB | 32.51 | 34.70 | beech |
| BE19 | ALB | 30.29 | 42.00 | beech |
| BE20 | ALB | 23.67 | 22.10 | spruce |
| BE21 | ALB | 22.43 | 37.70 | spruce |
| BE22 | ALB | 24.55 | 35.20 | spruce |
| BE23 | ALB | 26.49 | 34.70 | beech |
| BE24 | ALB | 24.00 | 27.30 | spruce |
Table 2.
All tested attributes of tree architecture and their relationship with competition strength.
Table 2.
All tested attributes of tree architecture and their relationship with competition strength.
| Architectural Attribute | rho | p-value |
|---|---|---|
| Branch volume 1st order | −0.26 | 0.227 |
| Branch volume 2nd order | −0.20 | 0.355 |
| Branch volume 3rd order | −0.25 | 0.237 |
| Total branch length 1st order | −0.53 | 0.007 |
| Mean branch length 1st order | −0.52 | 0.009 |
| Mean branch angle 1st order | 0.11 | 0.624 |
| Mean branch angle 2nd order | −0.07 | 0.755 |
| Mean branch angle 3rd order | −0.22 | 0.291 |
| Range of branch angles 1st order | −0.21 | 0.323 |
| Range of branch angles 2nd order | −0.62 | 0.002 |
| Range of branch angles 3rd order | −0.28 | 0.179 |
| Db (box dimension) | −0.65 | 0.006 |
| Intercept of Db-regression | −0.78 | <0.001 |
| Self-similarity | 0.31 | 0.142 |
(p-values in bold indicate statistically significant relationships).
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dorji, Y.; Annighöfer, P.; Ammer, C.; Seidel, D. Response of Beech (Fagus sylvatica L.) Trees to Competition—New Insights from Using Fractal Analysis. Remote Sens. 2019, 11, 2656. https://0-doi-org.brum.beds.ac.uk/10.3390/rs11222656
AMA Style
Dorji Y, Annighöfer P, Ammer C, Seidel D. Response of Beech (Fagus sylvatica L.) Trees to Competition—New Insights from Using Fractal Analysis. Remote Sensing. 2019; 11(22):2656. https://0-doi-org.brum.beds.ac.uk/10.3390/rs11222656
Chicago/Turabian StyleDorji, Yonten, Peter Annighöfer, Christian Ammer, and Dominik Seidel. 2019. "Response of Beech (Fagus sylvatica L.) Trees to Competition—New Insights from Using Fractal Analysis" Remote Sensing 11, no. 22: 2656. https://0-doi-org.brum.beds.ac.uk/10.3390/rs11222656
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.