Monitoring and Characterizing Temporal Patterns of a Large Colony of Tadarida brasiliensis (Chiroptera: Molossidae) in Argentina Using Field Observations and the Weather Radar RMA1


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Characterization and Monitoring of the Colony
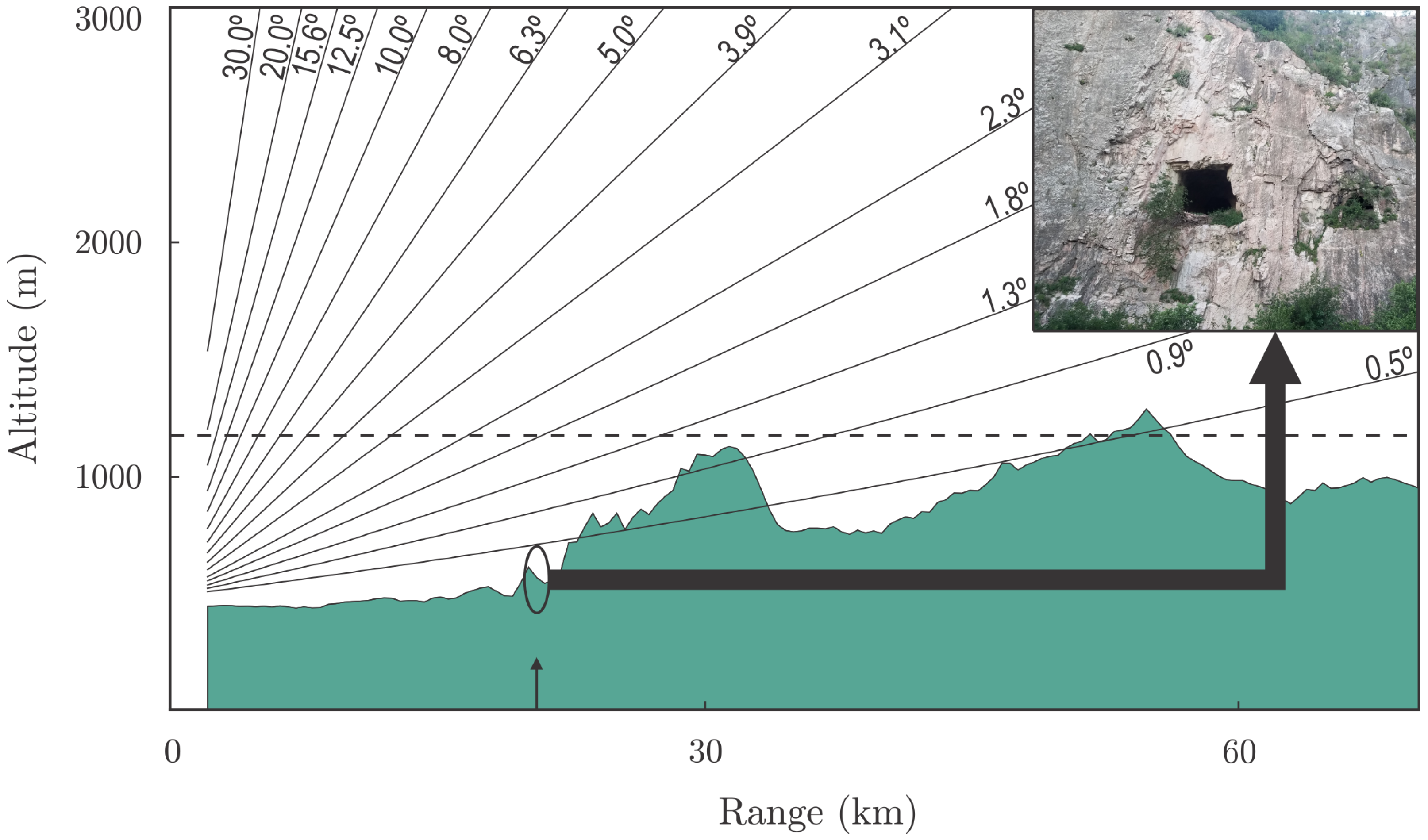
2.3. Radar Meteorológico Argentino 1 (RMA1)
2.4. Radar Sampling Data
2.5. Bat Detection
3. Results
3.1. Characterization and Monitoring of the Colony
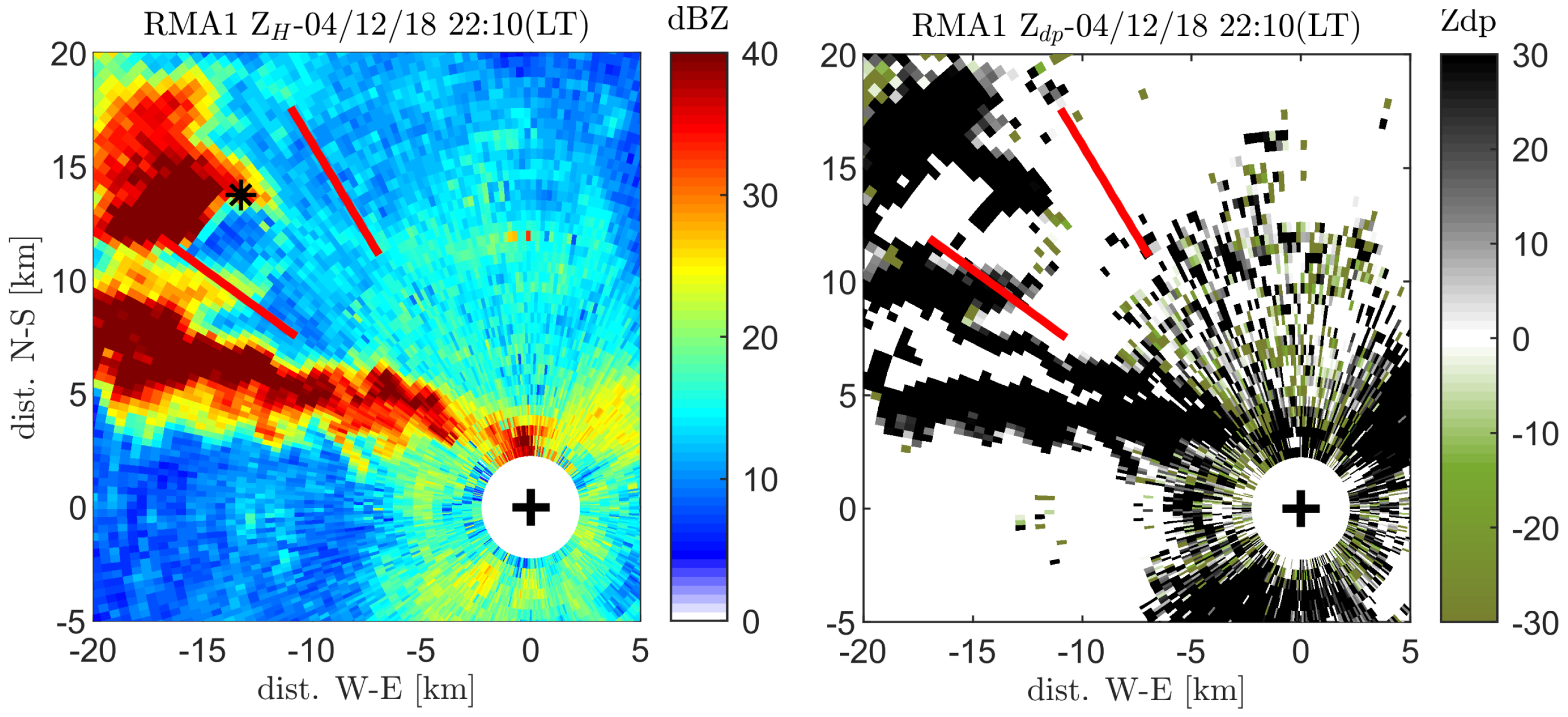
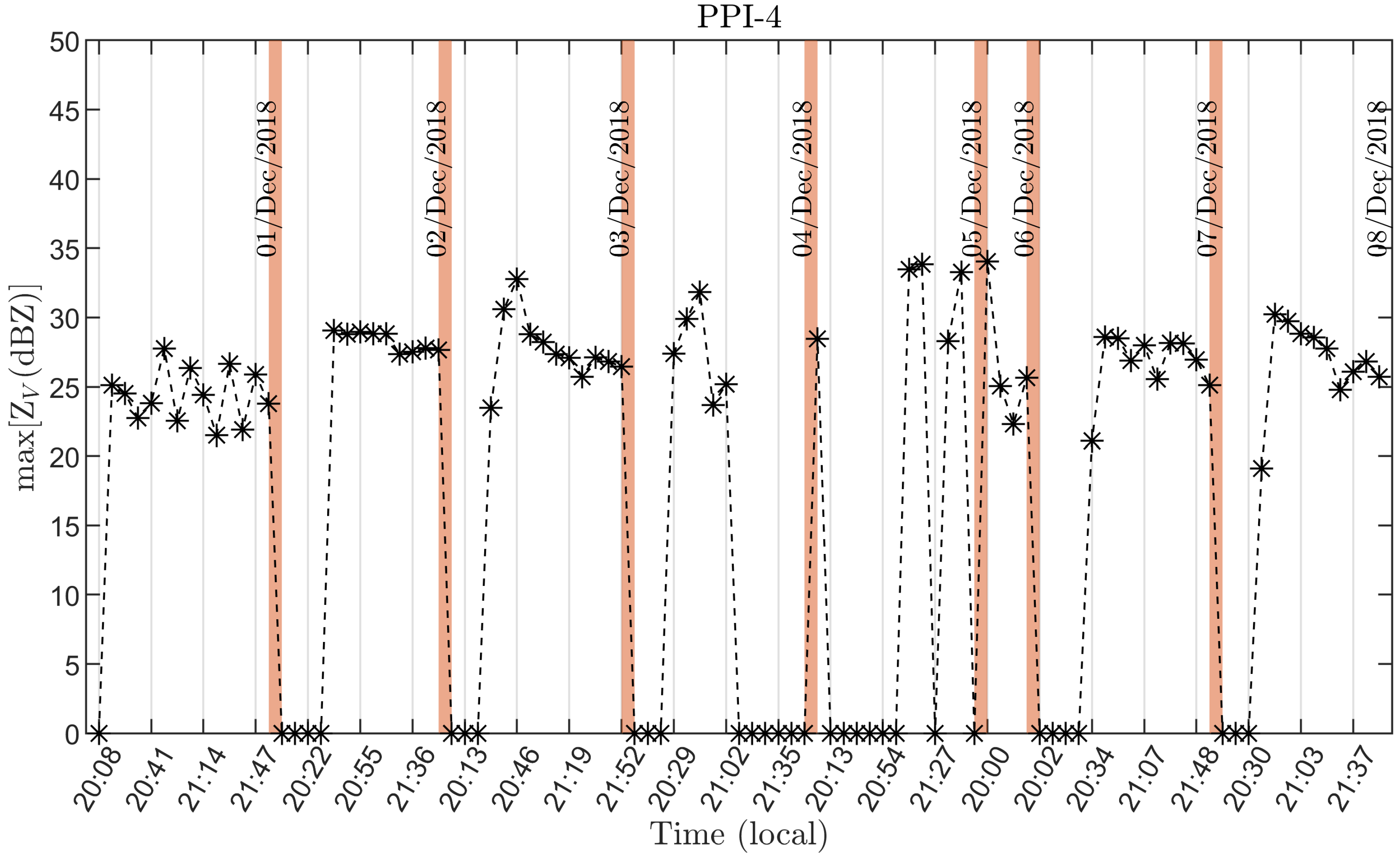
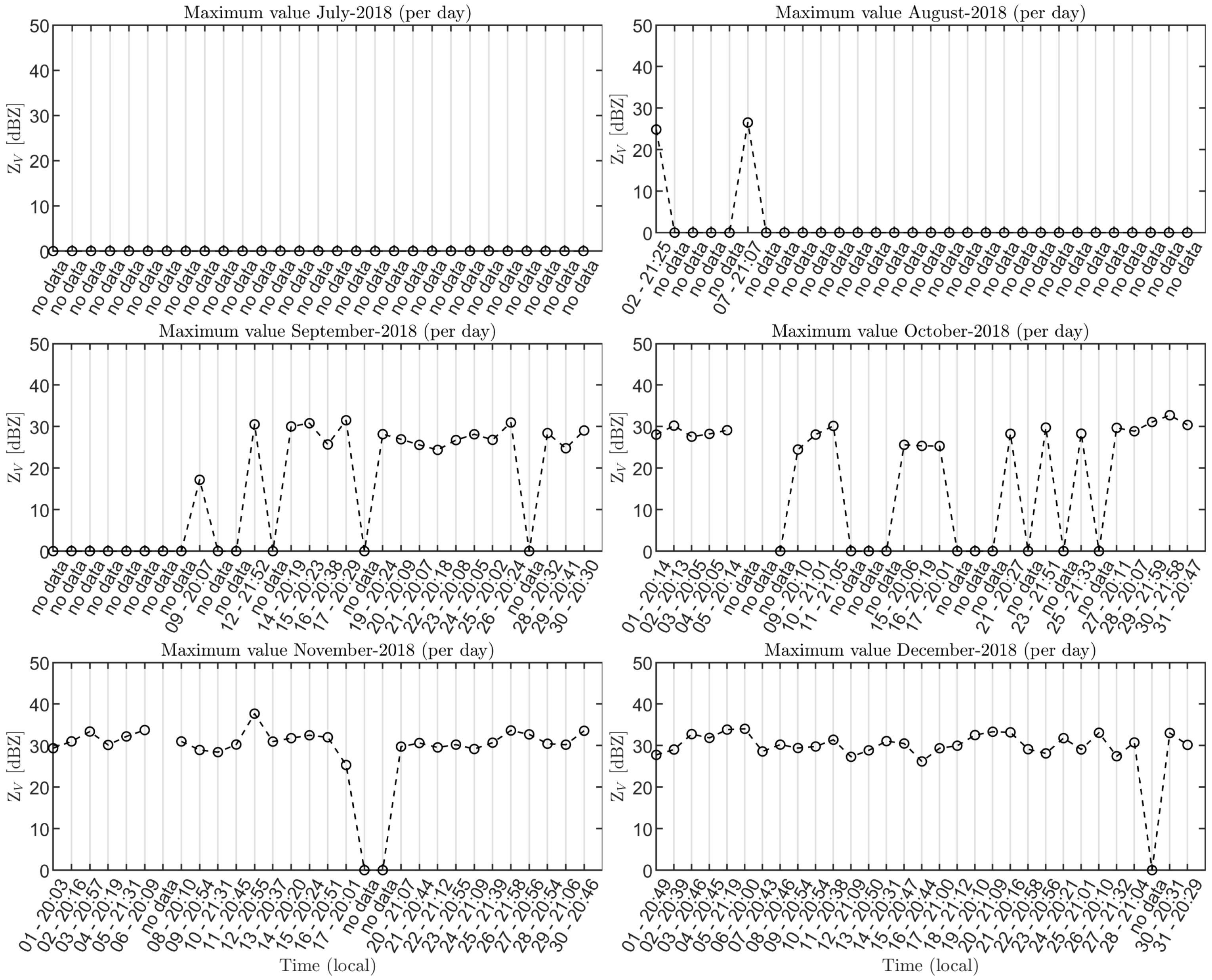
3.2. Radar Bat Detection
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Moore, F.; Gauthreaux, S.A.J.; Kerlinger, P.; Simons, T.R. Habitat requirements during migration: Important link in conservation. In Ecology and Management of Neotropical Migratory Birds; a Synthesis and Review of Critical Issues; Martin, T.E., Finch, D.M., Eds.; Oxford University Press: New York, NY, USA, 1995; pp. 121–144. [Google Scholar]
- Chilson, P.B.; Frick, W.F.; Kelly, J.F.; Howard, K.W.; Larkin, R.P.; Diehl, R.H.; Westbrook, J.K.; Kelly, T.A.; Kunz, T.H. Partly cloudy with a chance of migration: weather, radars, and aeroecology. Bull. Ame. Meteorol. Soc. 2012, 93, 669–686. [Google Scholar] [CrossRef]
- Loss, S.R.; Will, T.; Marra, P.P. Estimates of bird collision mortality at wind facilities in the contiguous United States. Biol. Conserv. 2013, 168, 201–209. [Google Scholar] [CrossRef]
- Kunz, T.H.; de Torrez, E.B.; Bauer, D.; Lobova, T.; Fleming, T.H. Ecosystem services provided by bats. Ann. N. Y. Acad. Sci. 2011, 1223, 1–38. [Google Scholar] [CrossRef] [PubMed]
- Boyles, J.G.; Cryan, P.M.; McCracken, G.F.; Kunz, T.H. Economic importance of bats in agriculture. Science 2011, 332, 41–42. [Google Scholar] [CrossRef] [PubMed]
- Voigt, C.C.; Kingston, T. Bats in the Anthropocene. In Bats in the Anthropocene: Conservation of Bats in a Changing World; Voigt, C.C., Kingston, T., Eds.; Springer: Cham, Switzerland, 2016; pp. 1–9. [Google Scholar]
- Jones, G.; Jacobs, D.S.; Kunz, T.H.; Willig, M.R.; Racey, P.A. Carpe noctem: the importance of bats as bioindicators. Endanger. Species Res. 2009, 8, 93–115. [Google Scholar] [CrossRef] [Green Version]
- Aguirre, L.F.; Medellín, R.A.; Rodríguez-Herrera, B. From threat to opportunity. In Tropical Conservation: Perspectives on Local and Global Priorities, 1st ed.; Aguirre, A.A., Sukumar, R., Eds.; Oxford University Press: New York, NY, USA, 2016; pp. 140–153. [Google Scholar]
- O’shea, T.J.; Cryan, P.M.; Hayman, D.T.; Plowright, R.K.; Streicker, D.G. Multiple mortality events in bats: A global review. Mammal Rev. 2016, 46, 175–190. [Google Scholar] [CrossRef] [PubMed]
- Schneeberger, K.; Voigt, C.C. Zoonotic viruses and conservation of bats. In Bats in The Anthropocene: Conservation of Bats in a Changing World; Springer: Cham, Switzerland, 2016; pp. 263–292. [Google Scholar]
- Simmons, N.B. An Eocene big bang for bats. Science 2005, 307, 527–528. [Google Scholar] [CrossRef]
- Barquez, R.M.; Carbajal, M.N.; Failla, M.; Díaz, M.M. New distributional records for bats of the Argentine Patagonia and the southernmost known record for a molossid bat in the world. Mammalia 2013, 77, 119–126. [Google Scholar] [CrossRef] [Green Version]
- Cleveland, C.J.; Betke, M.; Federico, P.; Frank, J.D.; Hallam, T.G.; Horn, J.; Lopez, J.D., Jr.; McCracken, G.F.; Medellín, R.A.; Moreno-Valdez, A.; et al. Economic value of the pest control service provided by Brazilian free-tailed bats in south-central Texas. Front. Ecol. Environ. 2006, 4, 238–243. [Google Scholar] [CrossRef]
- Wilkins, K.T. Tadarida brasiliensis; Mamm. Species; 1989; pp. 1–10. [Google Scholar]
- Davis, R.B.; Herreid, C.F.; Short, H.L. Mexican free-tailed bats in Texas. Ecol. Monogr. 1962, 32, 311–346. [Google Scholar] [CrossRef]
- Fabian, M.E.; Marques, R. Aspectos do comportamento de Tadarida brasiliensis brasiliensis (I. Geoffroy, 1824) (Chiroptera, Molossidae) em ambiente urbano. Biociências 1996, 4, 65–86. [Google Scholar]
- Romano, M.C.; Maidagan, J.I.; Pire, E.F. Behavior and demography in an urban colony of Tadarida brasiliensis (Chiroptera: Molossidae) in Rosario, Argentina. Rev. Biol. Trop. 1999, 47, 1121–1127. [Google Scholar]
- McCracken, G. Estimates of population sizes in summer colonies of Brazilian free-tailed bats (Tadarida brasiliensis). In Monitoring Trends in Bat Populations of the US and Territories: Problems and Prospects; Biological Resources Discipline, Information and Technology Report, USGS/BRD/ITR-2003-003; O’Shea, T.J., Bogan, M.A., Eds.; United States Geological Survey: Reston, VA, USA, 2003; pp. 21–30. [Google Scholar]
- Gamboa Alurralde, S.; Barquez, R.M.; Díaz, M.M. New records of bats (Mammalia: Chiroptera) for a southern locality of the Argentine Yungas. Check List 2017, 13, 2105. [Google Scholar] [CrossRef] [Green Version]
- Botto Nuñez, G.; Genta, M.; Díaz, M.; Rodales, A.L.; González, E.M. Circannual sex distribution of the Brazilian free-tailed bat, Tadarida brasiliensis (Chiroptera: Molosssidae), suggests migration in colonies from Uruguay. Mastozool. Neotrop. 2018, 25, 213–219. [Google Scholar] [CrossRef]
- Russell, A.L.; McCracken, G.F. Population genetic structuring of very large populations: The Brazilian free-tailed bat Tadarida brasiliensis. In Functional and Evolutionary Ecology of Bats; Akbar, Z., McCracken, G.F., Kunz, T.H., Eds.; University Press: New York, NY, USA, 2006; pp. 227–247. [Google Scholar]
- Chilson, P.B.; Frick, W.F.; Stepanian, P.M.; Shipley, J.R.; Kunz, T.H.; Kelly, J.F. Estimating animal densities in the aerosphere using weather radar: To Z or not to Z? Ecosphere 2012, 3, 1–19. [Google Scholar] [CrossRef]
- Chilson, P.B.; Bridge, E.; Frick, W.F.; Chapman, J.W.; Kelly, J.F. Radar Aeroecology: Exploring the Movements of Aerial Fauna through Radio-Wave Remote Sensing. Biol. Lett. 2012, 8, 698–701. [Google Scholar] [CrossRef]
- Bruderer, B. Three decades of tracking radar studies on bird migration in Europe and the Middle East. In Proceedings of the International Seminar on Birds and Flight Safety in the Middle East, Tel-Aviv, Israel, 25–29 April 1999; Leshem, Y., Mandelik, Y., Shamoun-Baranes, J., Eds.; Tel Aviv University: Tel Aviv, Israel, 1999; pp. 107–141. [Google Scholar]
- Gauthreaux, S.A., Jr.; Livingston, J.W.; Belser, C.G. Detection and discrimination of fauna in the aerosphere using Doppler weather surveillance radar. Integr. Comp. Biol. 2008, 48, 12–23. [Google Scholar] [CrossRef] [Green Version]
- Horn, J.W.; Kunz, T.H. Analyzing NEXRAD doppler radar images to assess nightly dispersal patterns and population trends in Brazilian free-tailed bats (Tadarida brasiliensis). Integr. Comp. Biol. 2008, 48, 24–39. [Google Scholar] [CrossRef] [Green Version]
- Bonter, D.N.; Gauthreaux, S.A., Jr.; Donovan, T.M. Characteristics of important stopover locations for migrating birds: Remote sensing with radar in the Great Lakes basin. Conserv. Biol. 2009, 23, 440–448. [Google Scholar] [CrossRef]
- Chapman, J.W.; Drake, V.A.; Reynolds, D.R. Recent insights from radar studies of insect flight. Annu. Rev. Entomol. 2011, 56, 337–356. [Google Scholar] [CrossRef] [Green Version]
- Poffo, D.A.; Beccacece, H.M.; Caranti, G.M.; Comes, R.A.; Drewniak, M.E.; Martina, A.; Zapata, A.I.; Rodriguez, A.; Saffe, J.N. Migration monitoring of Ascia monuste (Lepidoptera) and Schistocerca cancellata (Orthoptera) in Argentina using RMA1 weather radar. ISPRS J. Photogramm. Remote Sens. 2018, 145, 340–348. [Google Scholar] [CrossRef]
- Stepanian, P.M.; Wainwright, C.E. Ongoing changes in migration phenology and winter residency at Bracken Bat Cave. Glob. Chang. Biol. 2018, 24, 3266–3275. [Google Scholar] [CrossRef]
- Hüppop, O.; Ciach, M.; Diehl, R.; Reynolds, D.R.; Stepanian, P.M.; Menz, M.H. Perspectives and challenges for the use of radar in biological conservation. Ecography 2019, 42, 912–930. [Google Scholar] [CrossRef]
- Williams, T.C.; Ireland, L.C.; Williams, J.M. High altitude flights of the free-tailed bat, Tadarida brasiliensis, observed with radar. J. Mammal. 1973, 54, 807–821. [Google Scholar] [CrossRef]
- Mirkovic, D.; Stepanian, P.M.; Kelly, J.F.; Chilson, P.B. Electromagnetic model reliably predicts radar scattering characteristics of airborne organisms. Sci. Rep. 2016, 6, 35637. [Google Scholar] [CrossRef] [PubMed]
- McCracken, G.F.; Gillam, E.H.; Westbrook, J.K.; Lee, Y.F.; Jensen, M.L.; Balsley, B.B. Brazilian free-tailed bats (Tadarida brasiliensis: Molossidae, Chiroptera) at high altitude: links to migratory insect populations. Integr. Comp. Biol. 2008, 48, 107–118. [Google Scholar] [CrossRef] [Green Version]
- Wiederholt, R.; López-Hoffman, L.; Svancara, C.; McCracken, G.; Thogmartin, W.; Diffendorfer, J.E.; Mattson, B.; Bagstad, K.; Cryan, P.; Russell, A.; et al. Optimizing conservation strategies for Mexican free-tailed bats: A population viability and ecosystem services approach. Biodivers. Conserv. 2015, 24, 63–82. [Google Scholar] [CrossRef]
- Burkart, R.; Bárbaro, O.; Sanchez, R.; Gómez, D. Ecorregiones de la Argentina; Administración de Parques Nacionales y Secretaria de Recursos Naturales y Desarrollo Sustentable: Buenos Aires, Argentina, 1999; pp. 1–43. [Google Scholar]
- Barquez, R.M.; Díaz, M.M. Los Murciélagos de Argentina: Clave de Identificación; Publicación Especial 1 PCMA (Programa de Conservación de Murciélagos de Argentina), Ediciones Magna: Tucumán, Argentina, 2009; pp. 1–84. [Google Scholar]
- Lee, Y.F.; McCracken, G.F. Timing and variation in the emergence and return of Mexican free-tailed bats, Tadarida brasiliensis mexicana (Molossidae). Zool. Stud. 2001, 9, 309–316. [Google Scholar]
- Rauber, R.M.; Nesbitt, S.W. Radar Meteorology: A First Course, 1st ed.; Wiley Blackwell: Hoboken, NY, USA, 2018; pp. 1–488. [Google Scholar]
- Poffo, D.A.; Caranti, G.M.; Comes, R.A.; Rodriguez, A. A new ash concentration estimation method using polarimetric data: The RMA observation of the 2015 Calbuco eruption. Remote Sens. Appl. Soc. Environ. 2019, 13, 224–233. [Google Scholar] [CrossRef]
- Damino, V. Orden Chiroptera. In Mamíferos de Córdoba y su Estado de Conservación, 1st ed.; Torres, R., Tamburini, D., Eds.; Editorial de la UNC: Córdoba, Argentina, 2018; pp. 226–270. [Google Scholar]
- Díaz, M.M.; Carbajal, M.; Lipps, E.; Lutz, M.A.; Rosenfeld, S.; Barquez, R.M. El estado de conservación de los murciélagos de Argentina. In Manejo de Fauna Silvestre en la ArgentinaArgentina, Programas de Conservación de Especies Amenazadas; Porini, G., Ramadori, D., Eds.; Dirección de Fauna Silvestre, Secretaría de Ambiente y Desarrollo Sustentable de la Nación: Buenos Aires, Argentina, 2013; pp. 273–281. [Google Scholar]
- Regidor, H.; Mosa, S.; Núñez, A. Confinamento de una colonia de Tadarida brasiliensis, una alternativa de manejo compatible con la conservación. Chiropt. Neotrop. 2003, 9, 157–162. [Google Scholar]
- Pacheco, S.M.; Sodré, M.; Gama, A.; Bredt, A.; Cavallini, E.M.; Marques, R.V.; Guimarães, M.; Bianconi, G. Morcegos urbanos: Status do conhecimento e plano de ação para a conservação no Brasil. Chiropt. Neotrop. 2010, 16, 629–647. [Google Scholar]
- Villa, B.R.; Cockrum, E.L. Migration in the guano bat Tadarida brasiliensis mexicana (Saussure). J. Mammal. 1962, 43, 43–64. [Google Scholar]
- Cockrum, E.L. Migration in the Guano Bat, Tadarida brasiliensis; University of Arizona Department of Biological Sciences: Tucson, AZ, USA, 1969. [Google Scholar]
- Glass, B.P. Seasonal movements of Mexican freetail bats Tadarida brasiliensis mexicana banded in the Great Plains. Southwest. Nat. 1982, 27, 127–133. [Google Scholar] [CrossRef]
- Kunz, T.H.; Robson, S.K. Postnatal growth and development in the Mexican free-tailed bat (Tadarida brasiliensis mexicana): Birth size, growth rates, and age estimation. J. Mammal. 1995, 76, 769–783. [Google Scholar] [CrossRef]
- Blanco, A.; Gutierrez, M.; de Henquin, C.G.; de Burgos, N.M. Testicular lactate dehydrogenase isozyme: Cyclic appearance in bats. Science 1969, 164, 835–836. [Google Scholar] [CrossRef] [PubMed]
- Krutzsch, P.H.; Fleming, T.H.; Crichton, E.G. Reproductive biology of male Mexican free-tailed bats (Tadarida brasiliensis mexicana). J. Mammal. 2002, 83, 489–500. [Google Scholar] [CrossRef]
- Gustin, M.K.; McCracken, G.F. Scent recognition between females and pups in the bat Tadarida brasiliensis mexicana. Anim. Behav. 1987, 35, 13–19. [Google Scholar] [CrossRef]
- Gelfand, D.L.; McCracken, G.F. Individual variation in the isolation calls of Mexican free-tailed bat pups (Tadarida brasiliensis mexicana). Anim. Behav. 1986, 34, 1078–1086. [Google Scholar] [CrossRef]
- Reichard, J.D.; Gonzalez, L.E.; Casey, C.M.; Allen, L.C.; Hristov, N.I.; Kunz, T.H. Evening emergence behavior and seasonal dynamics in large colonies of Brazilian free-tailed bats. J. Mammal. 2009, 90, 1478–1486. [Google Scholar] [CrossRef] [Green Version]
- Herreid, C.F.; Davis, R.B. Flight patterns of bats. J. Mammal. 1966, 47, 78–86. [Google Scholar] [CrossRef]
- López-Vidal, J.; Elizalde-Arellano, C.; Arroyo-Cabrales, J.; Medellín, R. Observaciones sobre movimientos y comportamiento de Tadarida brasiliensis mexicana en la cueva El Salitre, Meztitlán, Hidalgo, México. In Avances en el Estudio de los Mamíferos de México II, 1st ed.; Lorenzo, C., Espinoza, E., Ortega, J., Eds.; Sociación Mexicana de Mastozoología: Ciudad de México, México, 2008; pp. 615–634. [Google Scholar]
- Kunz, T.; Oftedal, O.; Robson, S.; Kretzmann, M.; Kirk, C. Changes in milk composition during lactation in three species of insectivorous bats. J. Comp. Physiol. B 1995, 164, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.F.; Kuo, Y.M. Predation on Mexican free-tailed bats by peregrine falcons and red-tailed hawks. J. Raptor Res. 2001, 35, 115–123. [Google Scholar]
- Rodríguez-San Pedro, A.; Allendes, J.L. Depredación del murciélago de cola libre Tadarida brasiliensis (I. Geoffroy Saint Hilaire, 1824) por el cernícalo Falco sparverius Linnaeus, 1758, en un sector urbano de Santiago, Región Metropolitana, Chile. Biodivers. Nat. History 2015, 1, 6–8. [Google Scholar]
- Medellin, R.A.; Wiederholt, R.; Lopez-Hoffman, L. Conservation relevance of bat caves for biodiversity and ecosystem services. Biol. Conserv. 2017, 211, 45–50. [Google Scholar] [CrossRef]
- Arnett, E.B.; Baerwald, E.F. Impacts of wind energy development on bats: Implications for conservation. In Bat Evolution, Ecology, and Conservation; Adams, R.A., Pedersen, S.C., Eds.; Springer: New York, NY, USA, 2013; pp. 435–456. [Google Scholar]
- Aguirre, L.F.; Barquez, R.M. Critical areas for bat conservation: Latin American conservationists build a grand strategy. Bats 2013, 31, 10–12. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Observation Inside of the Colony Shelter | Observation Outside of the Colony Shelter (Emergence). LT = Local Time | Observations in Other Tunnels (T). Species (n° Bats) |
|---|---|---|---|
| 21 January 2012 | Full colony | Onset time = 19:50 LT Sunset time = 20:22 LT End time = 20:35 LT | - |
| 06 July 2012 | Without bats | - | - |
| 23 July 2012 | 500–1000 bats | - | T3 = Without bats T5 = Desmodus rotundus (small colony) |
| 11 July 2013 | More than 500 bats | - | T5 = D. rotundus (10) |
| 15 August 2013 | Without bats | - | T5 = D. rotundus (15) |
| 13 September 2013 | Without bats | - | T3 = Histiotus sp. (9), Myotis dinellii (1) T4 = M. dinellii (2), Tadarida brasiliensis (1) T5 = D. rotundus (15) |
| 24 October 2013 | Full colony. Adults. Colony size estimation | - | - |
| 12 December 2013 | Full colony. Adults and newborns | Onset time = 20:35 LT Sunset time = 20:14 LT End time = 21:17 LT | - |
| 27 January 2014 | Full colony. Adults and young | Onset time = 20:23 LT End time = 21:27 LT | - |
| 26 February 2014 | Full colony | - | T3 = Histiotus sp. (62), M. dinellii (2) |
| 13 March 2014 | Full colony | Onset time= 19:58 LT Sunset time = 19:35 LT End time = 20:40 LT | - |
| 26 March 2014 | Full colony | Onset time = 19:45 LT Sunset time = 19:19 LT End time = - | - |
| 15 April 2014 | Full colony | Onset time = 19:10 LT Sunset time = 18:55 LT End time = 20:00 LT | - |
| 30 April 2014 | Full colony | - | - |
| 13 May 2014 | Partial colony (less than 25%) | - | T3 = Histiotus sp. (2), M. dinellii (1) |
| 22 September 2014 | Full colony. Adults. | - | T3 = Histiotus sp. (1), M. dinellii (1) |
| 09 December 2014 | Full colony. Adults and young | Onset time = 20:30 LT Sunset time = 20:12 LT End time = - | - |
| 15 April 2015 | Partial colony (less than 50%) | - | - |
| 05 May 2015 | Partial colony (less than 25%) | - | - |
| 11 November 2016 | Full colony. Adults. | - | T3 = Histiotus sp. (40) |
| 02 February 2017 | Full colony. Adults and young | - | T3 = Histiotus sp. (2), M. dinellii (1) |
| 30 November 2017 | Full colony. Adults and newborns | - | T3 = Histiotus sp. (23), D. rotundus(1) |
| 07 December 2018 | - | Onset time = 20:30 LT Sunset time = 20:11 LT End time = 21:35 LT | - |
| 03 April 2019 | Full colony | - | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boero, L.; Poffo, D.; Damino, V.; Villalba, S.; Barquez, R.M.; Rodríguez, A.; Suárez, M.; Beccacece, H.M. Monitoring and Characterizing Temporal Patterns of a Large Colony of Tadarida brasiliensis (Chiroptera: Molossidae) in Argentina Using Field Observations and the Weather Radar RMA1. Remote Sens. 2020, 12, 210. https://0-doi-org.brum.beds.ac.uk/10.3390/rs12020210
Boero L, Poffo D, Damino V, Villalba S, Barquez RM, Rodríguez A, Suárez M, Beccacece HM. Monitoring and Characterizing Temporal Patterns of a Large Colony of Tadarida brasiliensis (Chiroptera: Molossidae) in Argentina Using Field Observations and the Weather Radar RMA1. Remote Sensing. 2020; 12(2):210. https://0-doi-org.brum.beds.ac.uk/10.3390/rs12020210
Chicago/Turabian StyleBoero, Lourdes, Denis Poffo, Verónica Damino, Sabrina Villalba, Rubén M. Barquez, Andrés Rodríguez, Matías Suárez, and Hernán Mario Beccacece. 2020. "Monitoring and Characterizing Temporal Patterns of a Large Colony of Tadarida brasiliensis (Chiroptera: Molossidae) in Argentina Using Field Observations and the Weather Radar RMA1" Remote Sensing 12, no. 2: 210. https://0-doi-org.brum.beds.ac.uk/10.3390/rs12020210