1. Introduction
Climate change modifies species composition and extended drought increases fire risk and pests/diseases attacks [
1,
2]. Climate change is a significant factor contributing to the increase of forest fires [
3] and tree species being unable to adapt to the severity and frequency of droughts during the summer period. The possibility of pest attacks and tree diseases gets higher because trees are weakened by the extreme weather conditions [
4]. According to Shoukri and Zachariadis [
5], in comparison to other European regions, the Mediterranean region will be more affected by climate change [
5]. Cyprus is the third largest island of the Mediterranean sea [
6] and it lies in its north-eastern end (33
east of Greenwich and 35
north of the Equator) [
7]. The island has a diversity of microclimates [
6] and diverse landscapes [
7] that are sufficiently isolated to support a high number of species leading to the formation of endemic biota [
7]. According to ancient statements written by historic authors including Eratosthenes (275-195 B.C), Cyprus was rich in forests. The forests in plains were too dense for inhabitation [
8,
9]. According to a statement provided by A. K. Bovill and D.E Hutchins (1909), the demarcated forests in Cyprus were 10.7% and 19% if scrub-forests were also included [
9]. According to Delipetrou et al. [
7] (2008), 18.7% of the island is forested. A recent quantitative study showed that 65.62% participants, Cypriot residents, noticed a moderate to very large degradation of Cypriot coniferous forests [
10]. The Department of Forests, Cyprus, (2018) also claimed that Cypriot forests are negatively influenced by prolonged droughts occurrences due to climate change. The consequences of the prolonged droughts include high temperatures and a lack of soil moisture leading to intense forest species stress in Cypriot forest ecosystems (Department of Forests, 2018). The impacts of climate change may include alternations in the distribution of forest species, increased forest mortality events, higher fire risk and species extinction [
5].
Another potential threat that Cypriot and Mediterranean forests may face due to climate change is the increased attacks from pests. The pine processionary (
Thaumetopoea pityocampa) eats the needles of Scots pines (
Pinus sylvestris L.) and it is responsible for seasonal defoliation. According to Hódar et al. [
11], the attacks of pine processionary (
Thaumetopoea pityocampa) to Scots pines (
Pinus sylvestris L.) has increased in the Mediterranean region due to climate change. This causes a significant reduction of pine growth as well as some deaths [
11]. For example, Toffolo et al. [
12] reported significant intense defoliation events and an expansion of the action of the pine processionary in Northern Italy [
12]. If there are no extreme weather conditions, the infected pine trees produce new needles and survive [
13]. When the pest builds nests on the same tree for 3–4 continuous years, then the tree is negatively influenced, and tree mortality may occur [
13]. Pine processionary did not exist in higher elevation areas of the Troodos mountain range, but it is expanding, and its action is strengthened by the persistent drought and warmer weather conditions [
13], posing a potential threat to forest species that exist at higher elevations.
According to the “report on the future climate change impact, vulnerability and adaptation assessment for the case of Cyprus”, published in 2016, there are two important climate change threats for Cypriot forests: fires and “dieback of tree species, insect attacks and diseases” [
14]. Forest fires are usually the priority of the Department of Forests since they have an immediate effect on the forests. Nevertheless, according to Lemesios et al. [
14], both threats have the same vulnerability scores: a very high sensitivity, very high exposure and moderate adaptive capacity. It is, therefore, of crucial importance to study, monitor, model forest health and its resilience, as well as to introduce policies using scientific evidence for preserving forests and a healthy ecosystem. According to Stenlid et al. [
15], many anthropogenic factors strengthen forest diseases in Europe, but there is no adequate legislation to limit those factors [
15]. For that reason, there is a need to create forecasting models and derive concrete measurements that will add on and improve current knowledge of our forests. These models and the concrete results could be used as scientific evidence for promoting environmentally friendly policies aiming to mitigate the effects of climate change, preserve a healthy ecosystem and consequently maintain social stability and stimulate economic growth.
Phenology is “the study of the timing of recurring biological events, the causes of their timing with regard to biotic and abiotic forces and the interactions among phases of the same or different species”, as defined by the US committee on phenology [
16,
17]. Phenology observes both plants and animals, as well as seasonal characteristic of natural phenomena [
17]. For example, Gittings et al. [
18] assessed the phenological indices of phytoplankton blooms in relation to regional ocean warming and showed that warmer conditions were associated with significantly weaker phytoplankton blooms during the winter season [
18]. Similar work exists on observing the phenology of various plants and it was shown that climate change conferred shifts to plants’ blooming time [
19]. There are also efforts to predict how plants react to warmer conditions and it was shown that plants with higher temperature sensitivity bloomed earlier, but overall, the phenological responses to climate changes were unpredicted [
20]. Recently, Wolf et al. [
21] showed that a reduction of plant biodiversity caused the shifting of flowering time, thus demonstrating the significance of biotic interactions.
Satellite sensors have been widely used for observing the phenological stages of plants. Gupta et al. [
22] showed that apple growth was highly correlated with the Normalized Difference Snow Index (NDSI). Aragones et al. [
23] proved that pine species could be classified using phenology-derived classes using the Normalised Difference Vegetation Index (NDVI) in Mediterranean forest. Touhami et al. [
24] compared time series NDVI data with precipitation data and showed that land surface phenology was mostly affected by climate parameters during autumn and spring. Frison et al. [
25] investigated the potentials of Sentinel-1 synthetic aperture radar (SAR) data for monitoring forest phenology and claimed that phenology could be estimated with a higher accuracy using SAR than optical data.
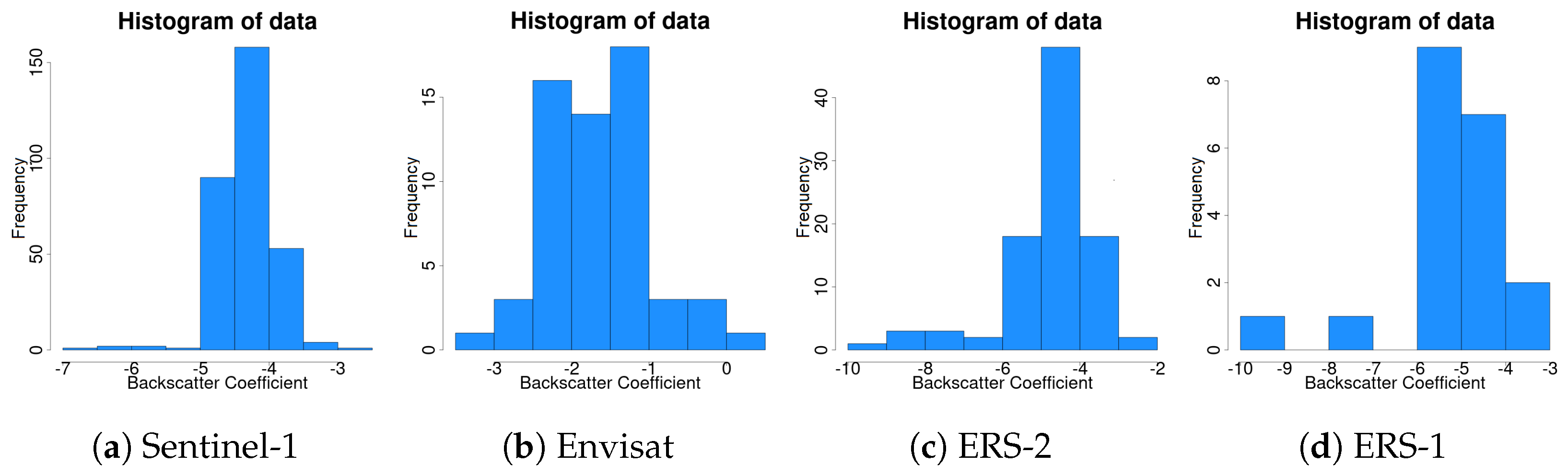
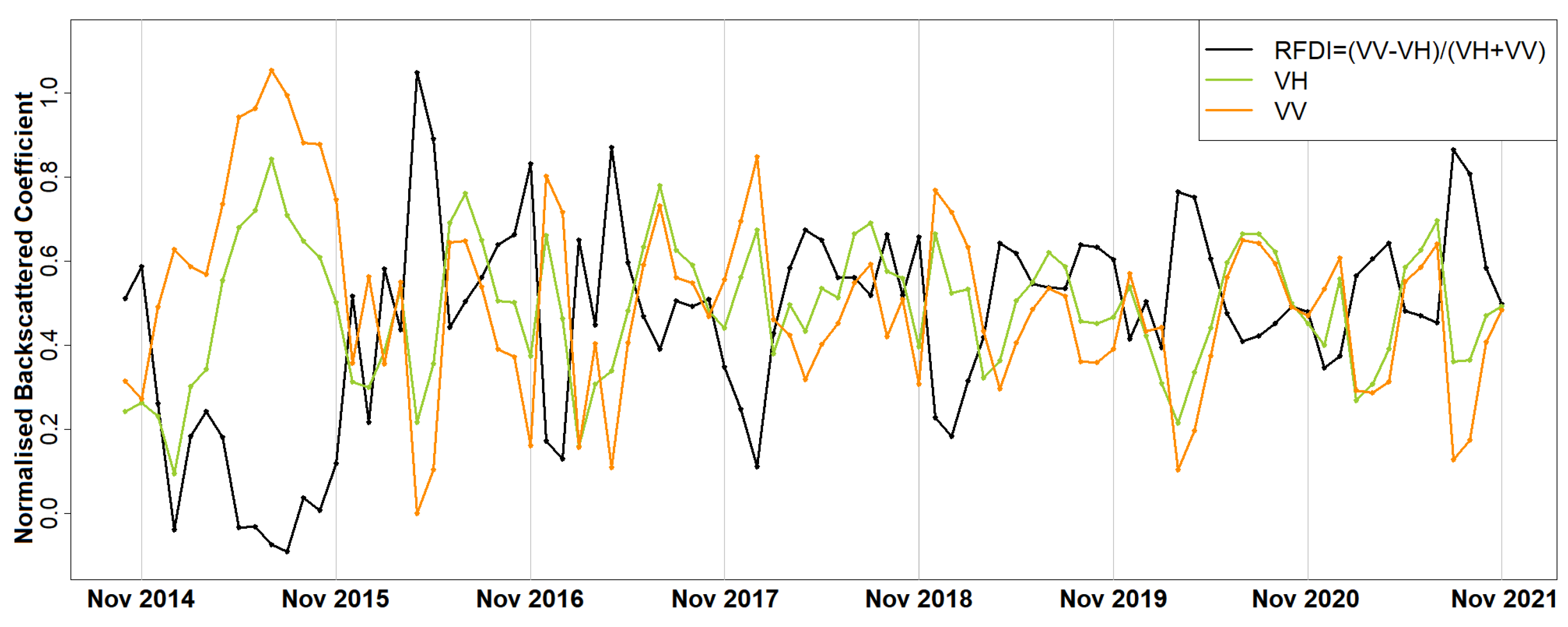
SAR sensors emit microwave energy and record the backscattered signal. Remote sensing is advancing, offering a higher and higher temporal resolution. The Sentinel-1 constellation offers a high temporal resolution of a 6-day repeat cycle making time series analysis of SAR data possible, while previous freely available data included ERS1/2 and Envisat with a repeat cycle of 35 days. Microwave remote sensing is important since microwave radiation can penetrate through objects and can record information, e.g., below a forest canopy, that cannot be acquired by traditional optical remote sensing sensors. The backscattered energy for a particular wavelength is proportional to structure and moisture [
26]. While NDVI provides information about the greenness of the plants [
26], the values of interferometry coherence can detect vegetation density [
27]. Due to the ability of SAR data to derive forest-density-related parameters, the C-band (used in this paper) has been exploited for biomass estimations [
28]. Seasonal changes (i.e., phenological changes) observed in evergreen forests using SAR data are hypothesised to be linked to tree foliage and the dielectric constant of the woody component of the trees [
29]. If the precipitation is removed or reduced to a minimum, then the tree foliage, water content of the trees and dielectric constant of the wood are the most likely information contained within the backscattering coefficient of the C-band. A time series of C-band SAR data could reveal how these parameters change seasonally and over time. Within a time series of SAR data, the recurrence of phases (i.e., phenology) is measurable as shown in this paper. SAR data were, therefore, selected for observing how the density and water content (e.g., foliage, needles regeneration, fruition) of the forest changes seasonally and over the years [
30].
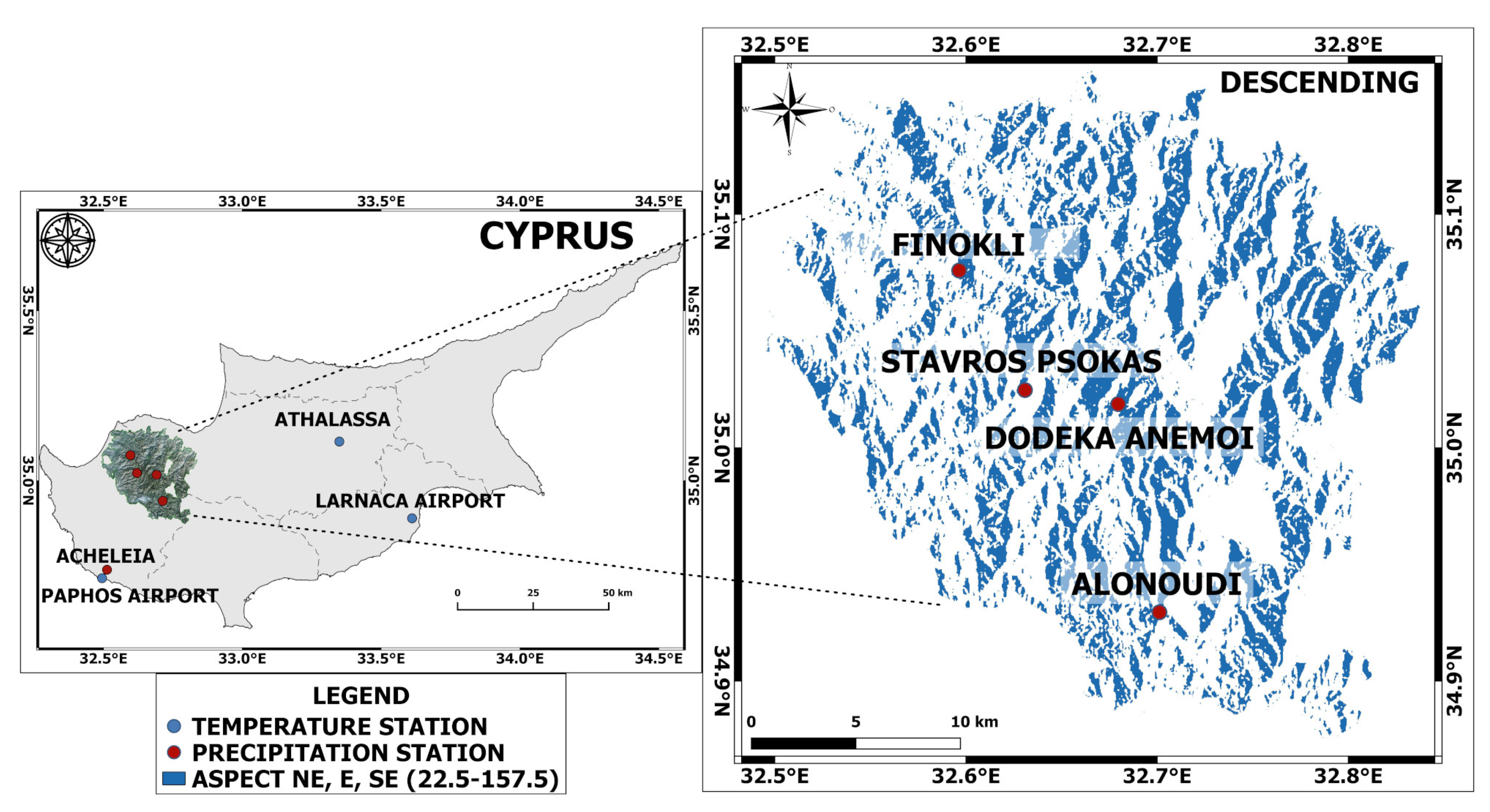
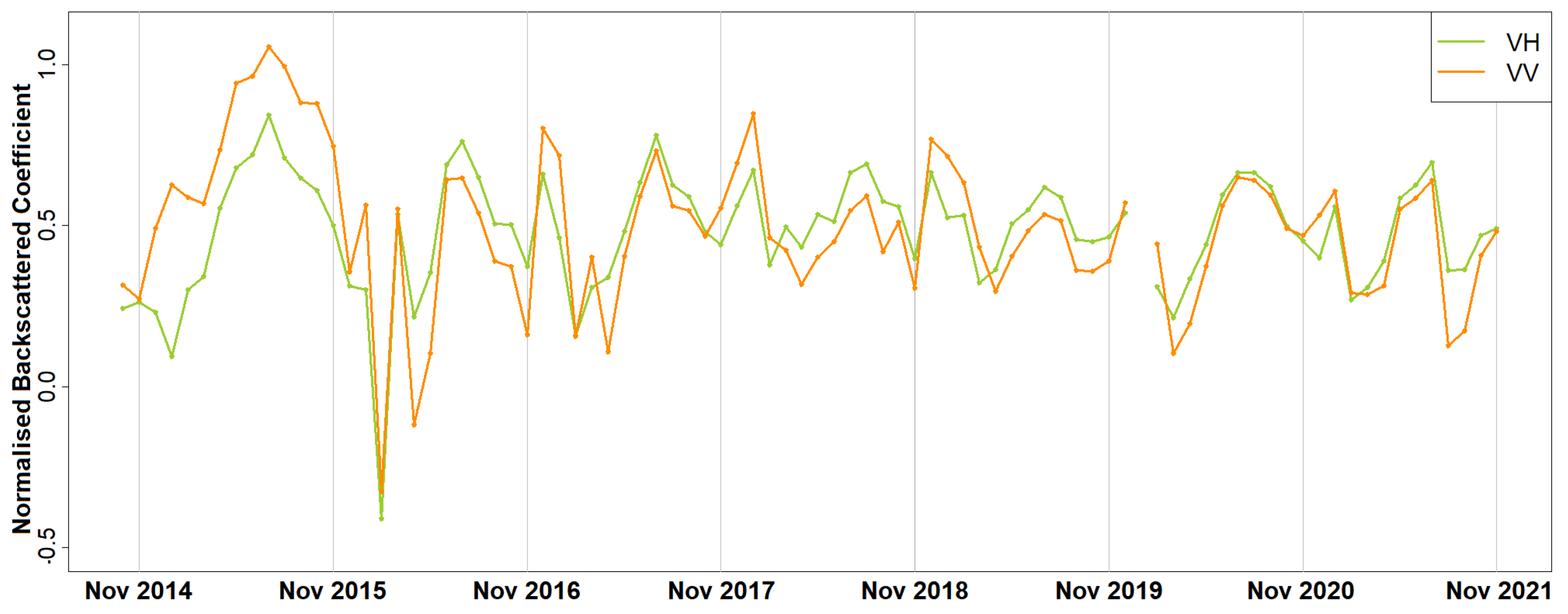
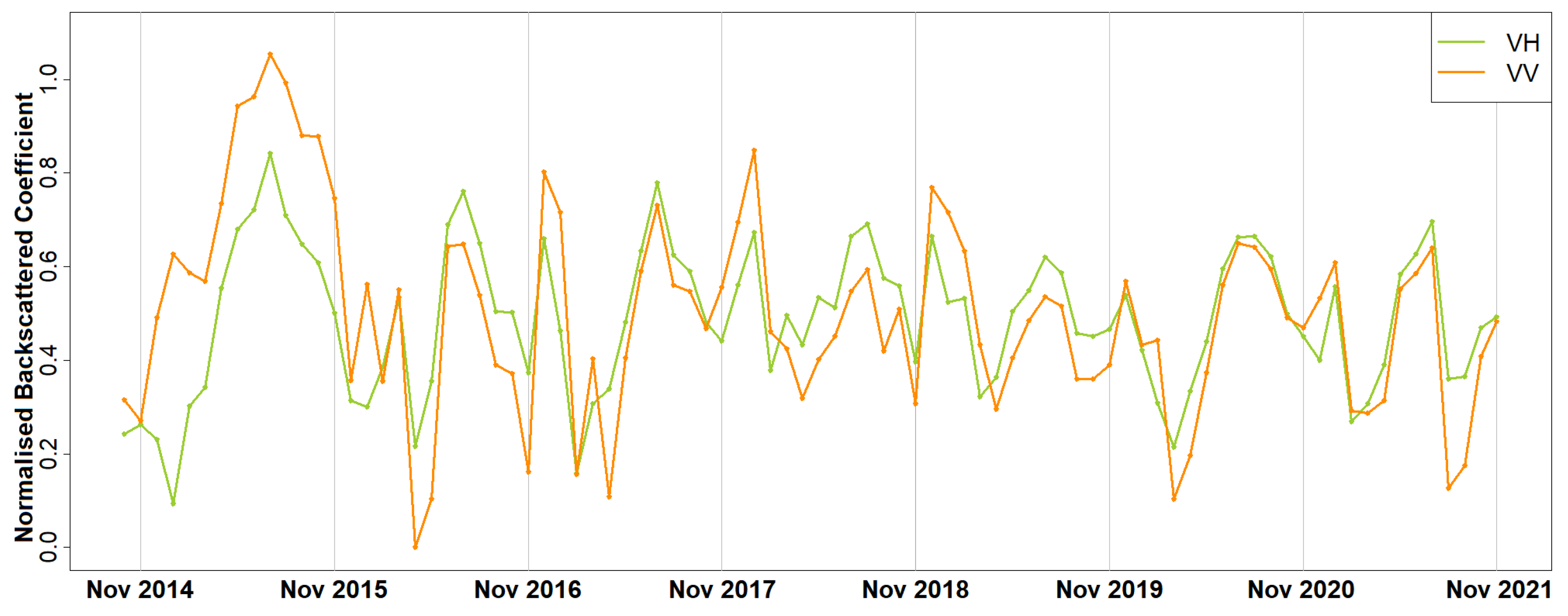
Furthermore, it is important to understand and predict the impacts of climate on forests since the acquired knowledge can be incorporated into decision making [
31]. SAR data have been available since 1992 and these data could be used to study climate change effects within a period of 30 years. This study interprets all the available SAR data for the study area but predominantly focuses on the period between November 2014 and October 2021. Within this time period, Sentinel-1 data are available. Sentinel-1 provides a higher temporal resolution [
32] while previous missions (ERS1/2, Envisat) present many gaps within the available datasets for the study area. This paper aimed to provide an in-depth understanding of the strengths and limitations of analysing time series SAR data for finding the drivers causing density-, foliage- and/or moisture-related phenological changes in Paphos forest, Cyprus. This was achieved by accomplishing the following objectives:
Study and understand the average annual phenological cycle of Paphos forest and how the time series diagrams could be improved by reducing the effect of precipitation.
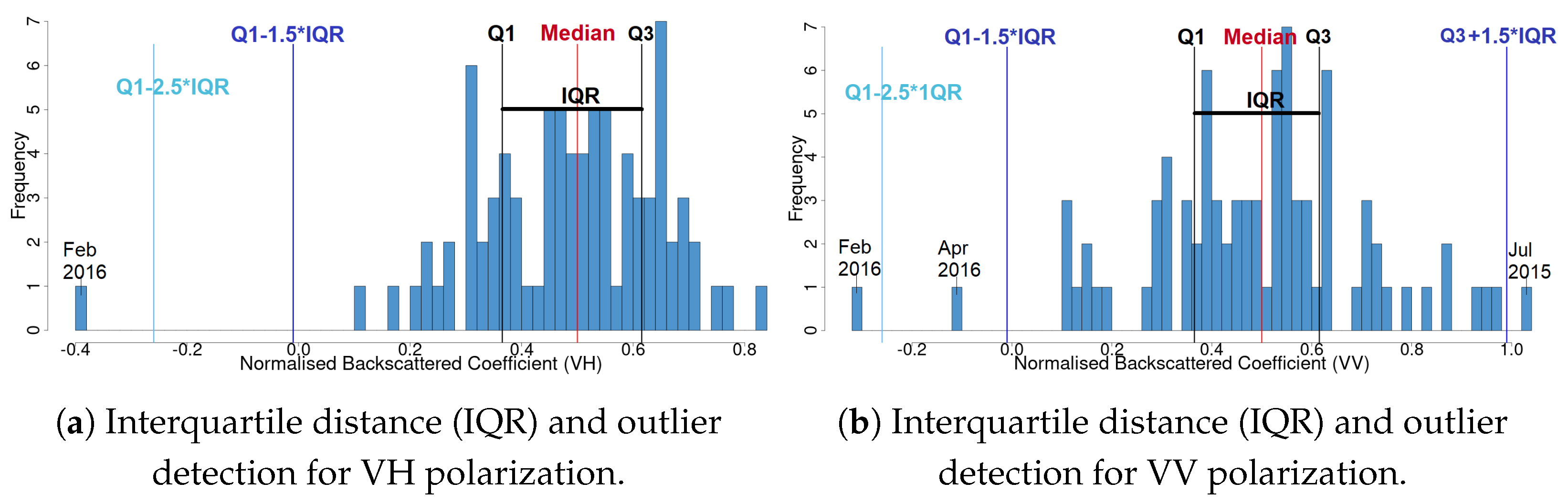
Detect outliers and justify their importance. Outliers may appear at unusual forest changes that may occur at important events (e.g., a pest attack). They need to be removed before creating predictive models.
Measure the initiation, duration and termination of detected peaks and link them with the relevant literature to understand the physical parameters that each peak may relate to.
Investigate the connections between unusual changes in the SAR time series and meteorological thermal and precipitation data.
Create and evaluate the prediction models and trend lines.
Investigate the applicability of existing SAR vegetation indexes for time series analysis of data.
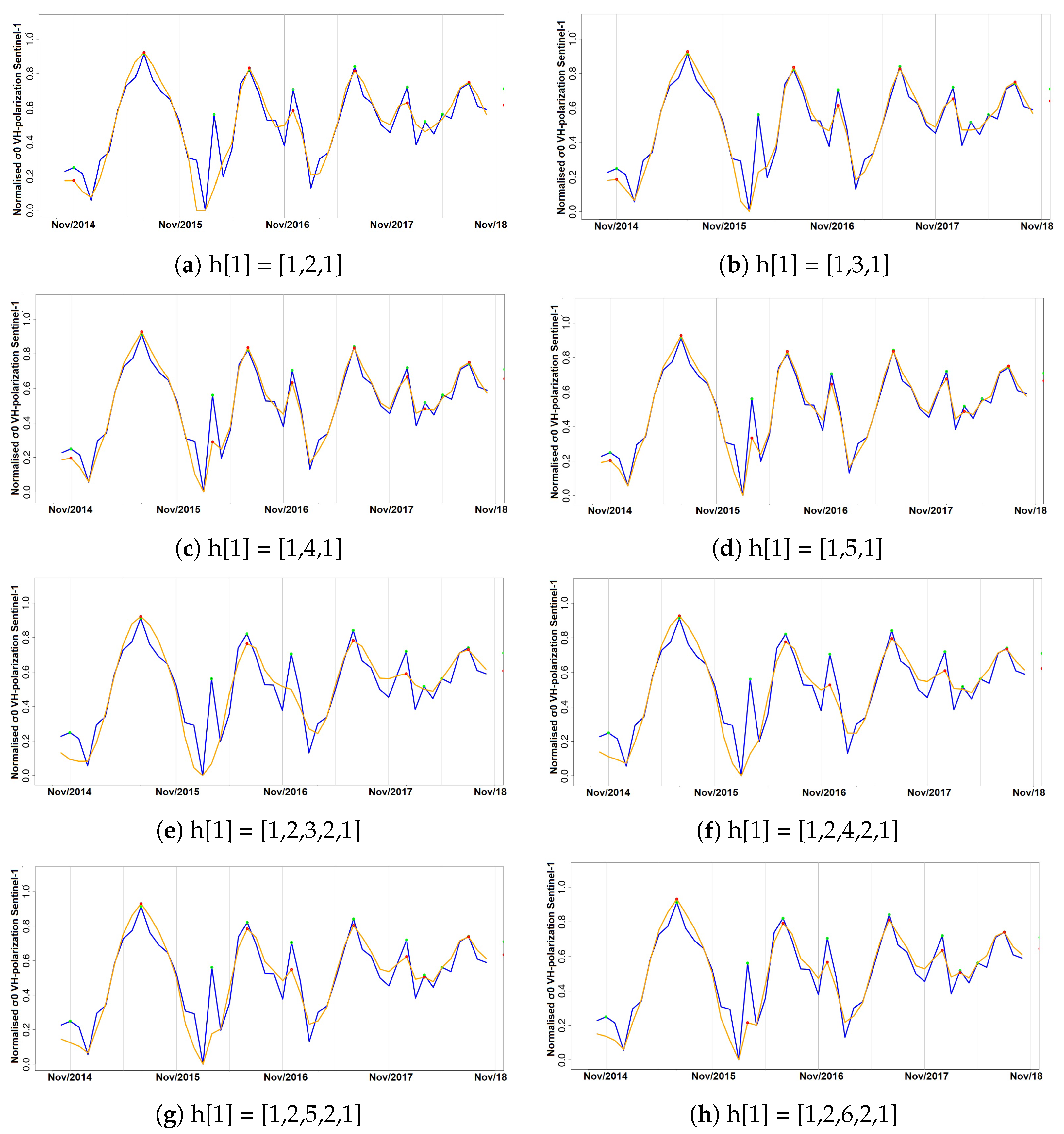
Experiment with filtering approaches and their applicability for removing noise in the SAR data time series diagrams.
7. Conclusions
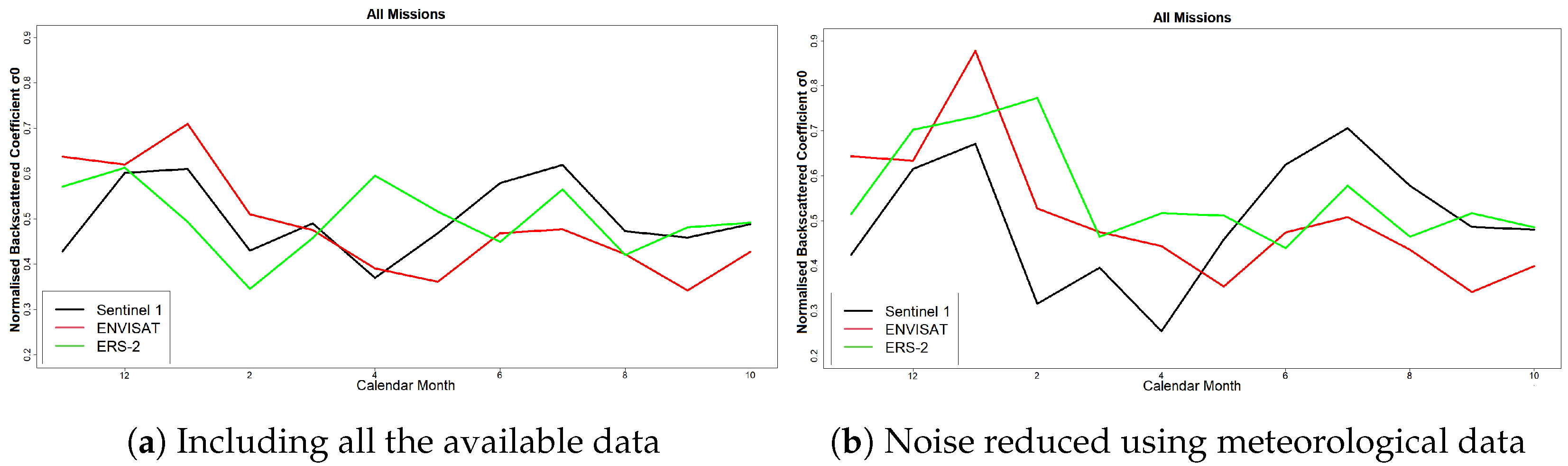
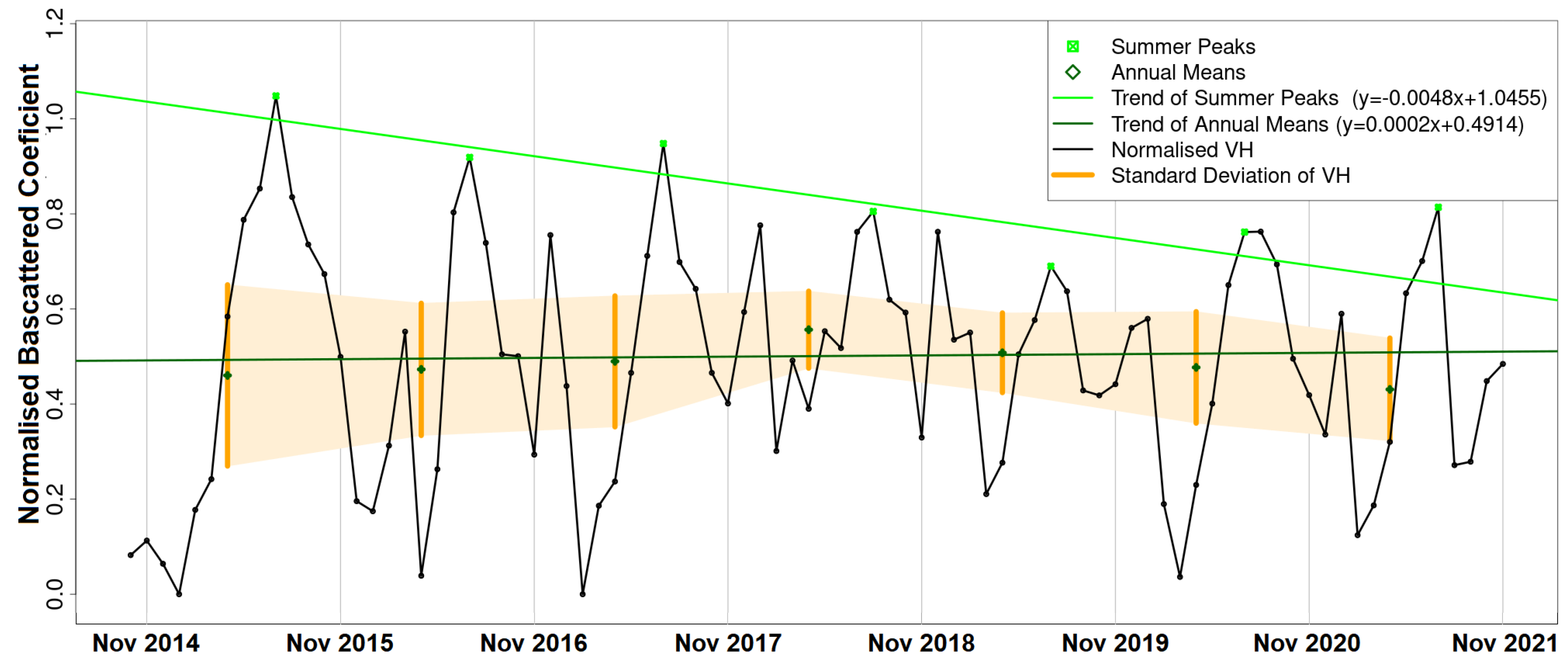
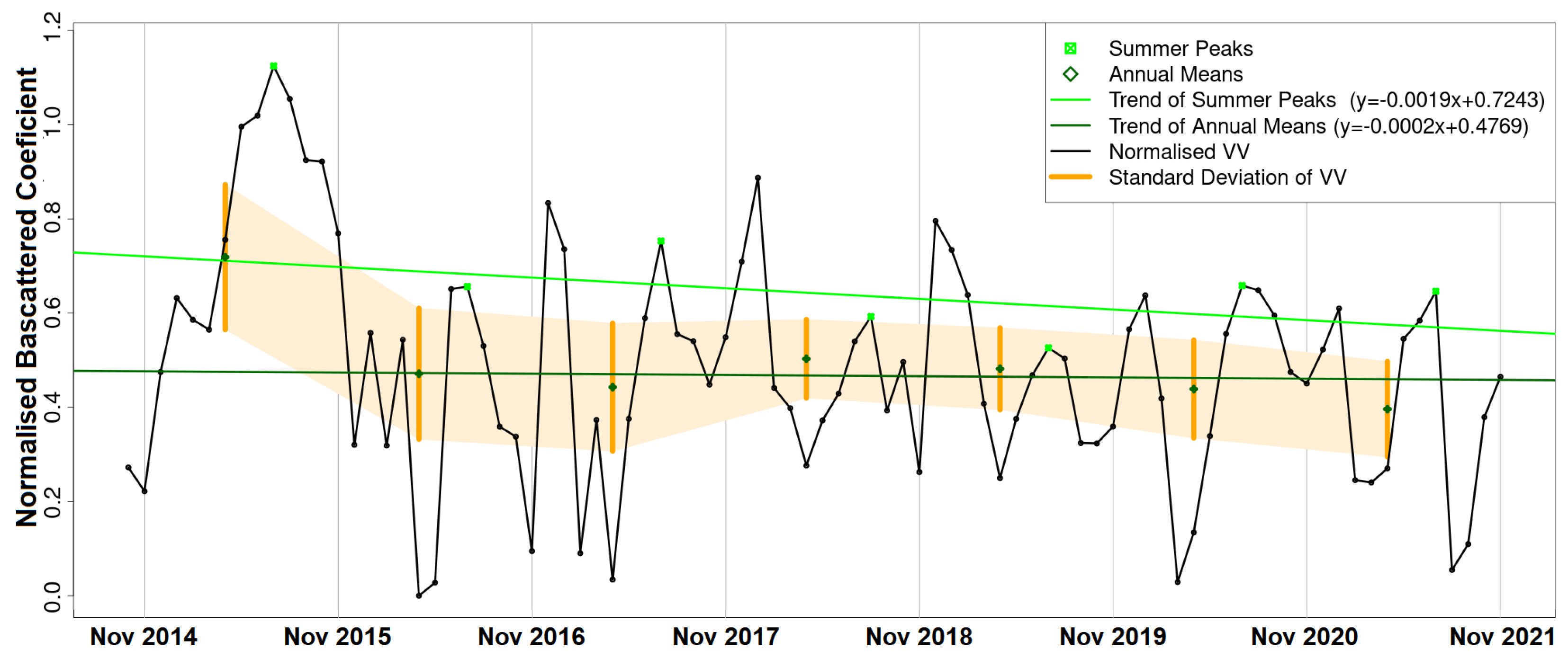
The results of this paper both agree with and adds to the existing literature. As in [
25], who claimed that phenology can be estimated with higher accuracy using SAR than optical data. It was shown that phenological diagrams derived with spaceborne SAR data of Paphos forest, Mediterranean sea, Cyprus, contained two major peaks instead of one identified with optical imagery [
34]. After a direct communication with experienced foresters from the Department of Forests, Cyprus, it was concluded that the most reasonable explanation for the summer peak was the annual regeneration of the needles and the drop of the old ones. Furthermore, similarly to [
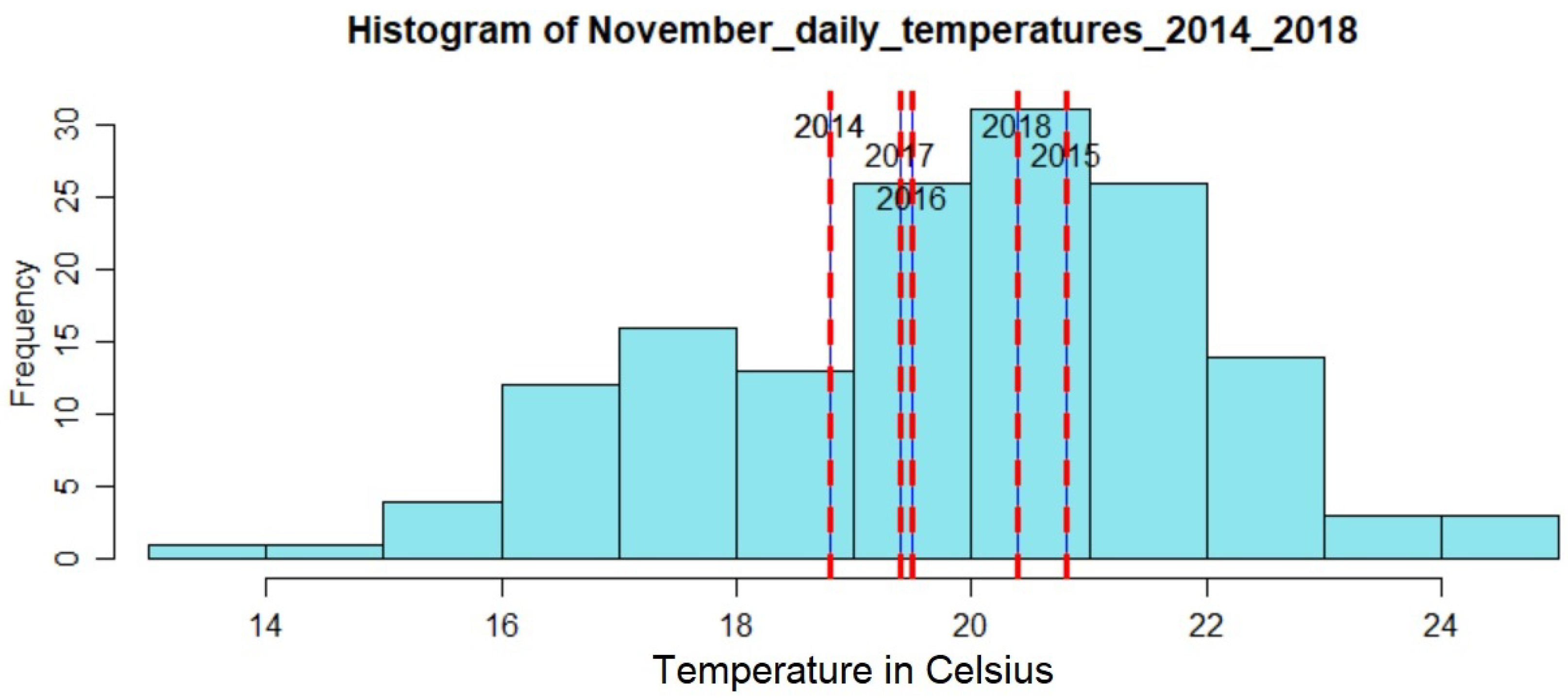
24], we showed that autumn and spring climatic conditions play a substantial role in changes presented in land surface phenology. Thus, if the temperature in May is high, then there may be a delay of the summer peak. Additionally, low temperatures in November may relate to a decreased action of the pine processionary (
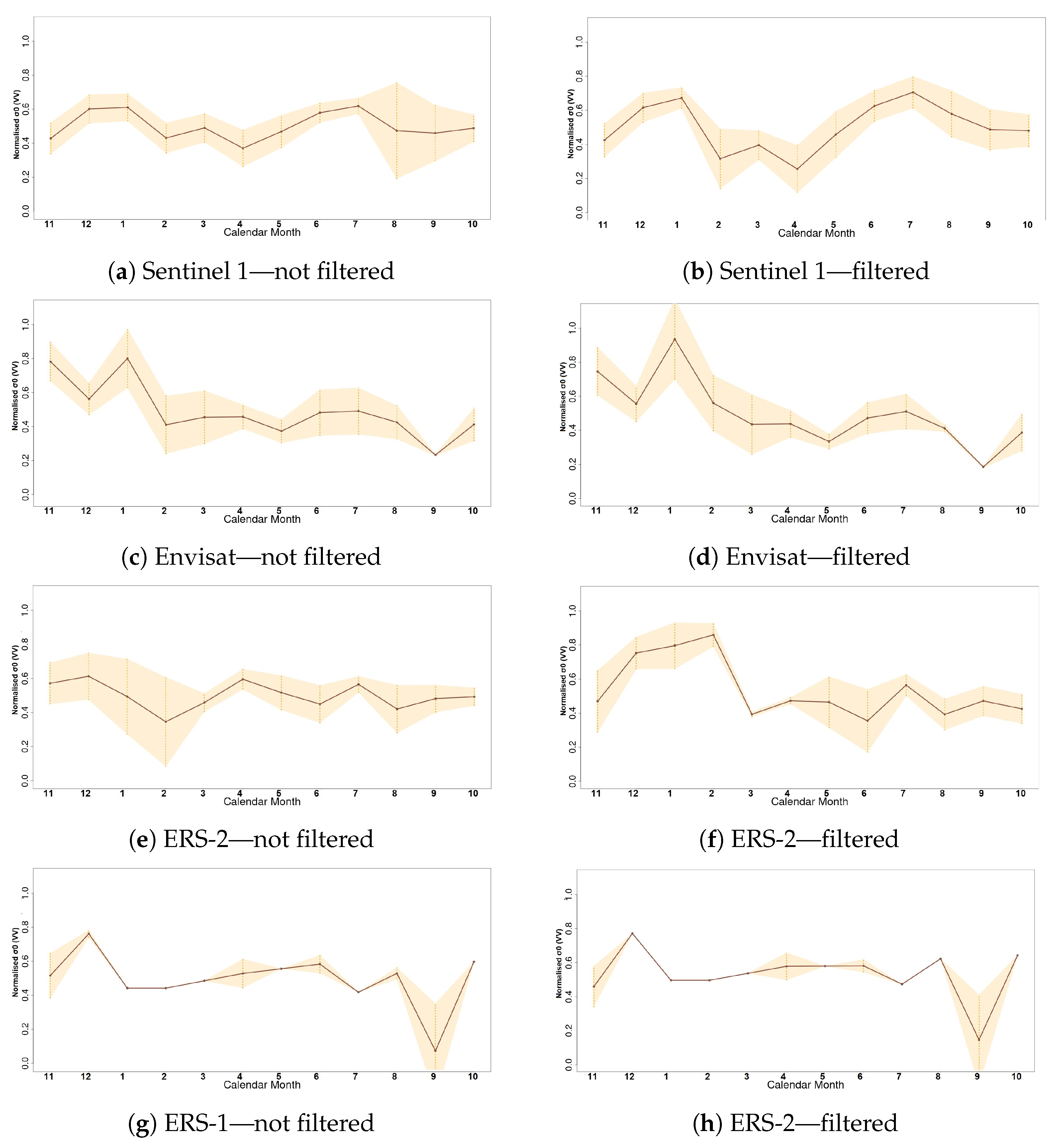
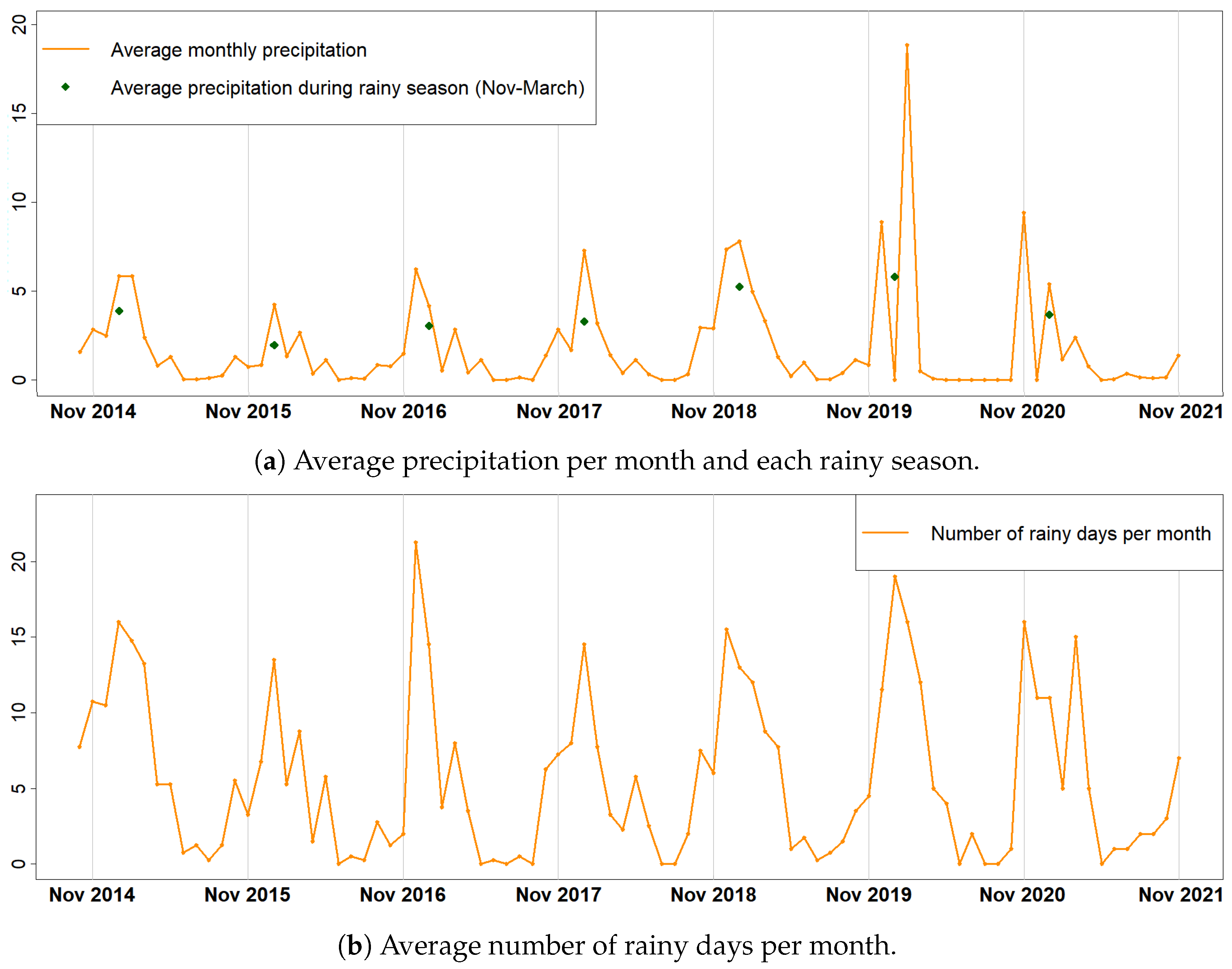
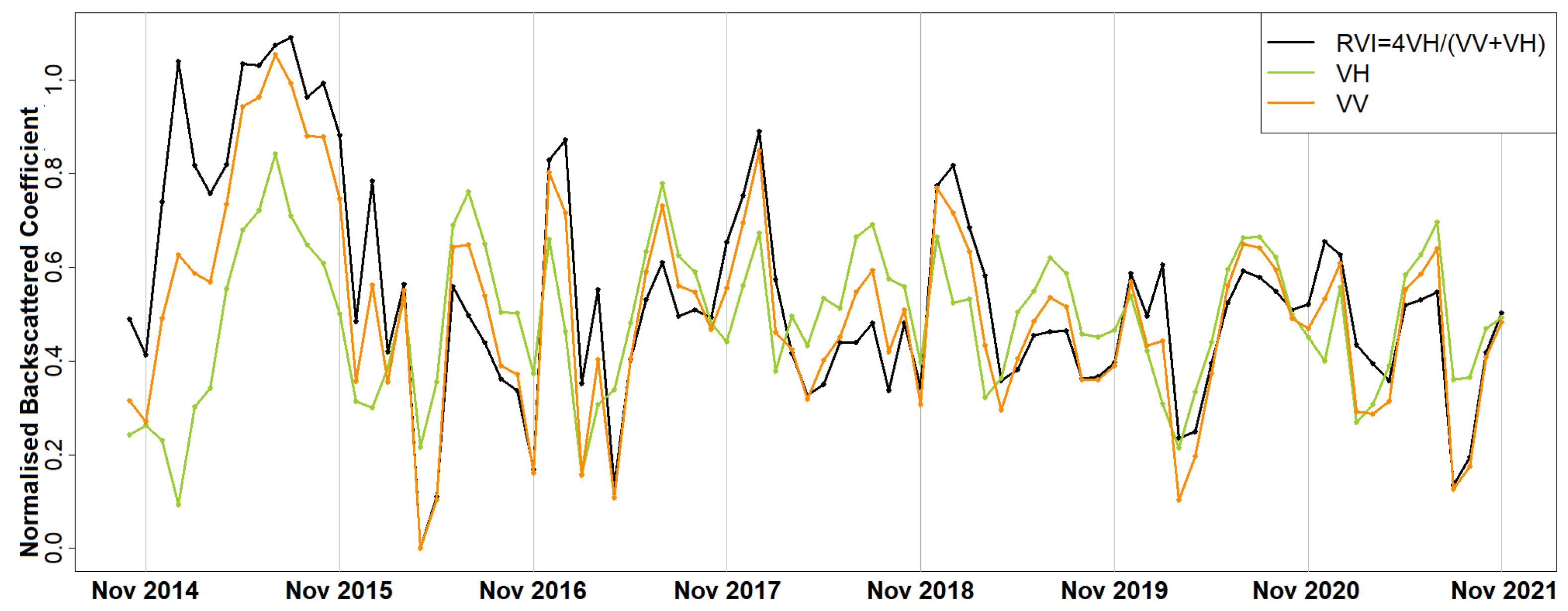
Thaumetopoea pityocampa) around February and consequently the appearance of one major peak, instead of two, over a phenological year (November-October). Equally important was the association of the rainy season of November 2015–March 2016 that drought (reduced average precipitation) existed with the outliers of February 2016 and April 2016. Secondary conclusions of this work include: a preprocessing that includes smoothing of signals can influence the quality of the results by dropping small peaks that may be important; the radar vegetation indexes (RVI and RFDI) were considered unreliable for time series phenological observations of forests, and in contrast to RVI and RFDI, both
and
returned high amplitudes and became comparable once normalised; finally, the detection of peak amplitude and mean backscattering coefficient was possible using trend lines but the time series was short and the trend was highly sensitive to other factors (e.g., precipitation) producing high rRMSE values. Overall, the launch of Sentinel-1 brought new research opportunities for observing the phenological changes of forests. After some more years of Sentinel-1 operation, when the period of investigation is longer and more data are available for the time series analysis, the use of advanced machine/deep learning techniques [
52] and signal processing approaches could improve prediction. Combining multisensor/multimodal data improves prediction [
50,
53]. Therefore, combining satellite multisensor, ground-truth and other spaceborne data in the time series analysis are soon expected to emerge for a better understanding and modelling of the drivers of phenological changes. This will further support climate-related research.


 ,
, 














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}