
Diversity Effects on Canopy Structure Change throughout a Growing Season in Experimental Grassland Communities

 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Study Site and Trait-Based Experiment
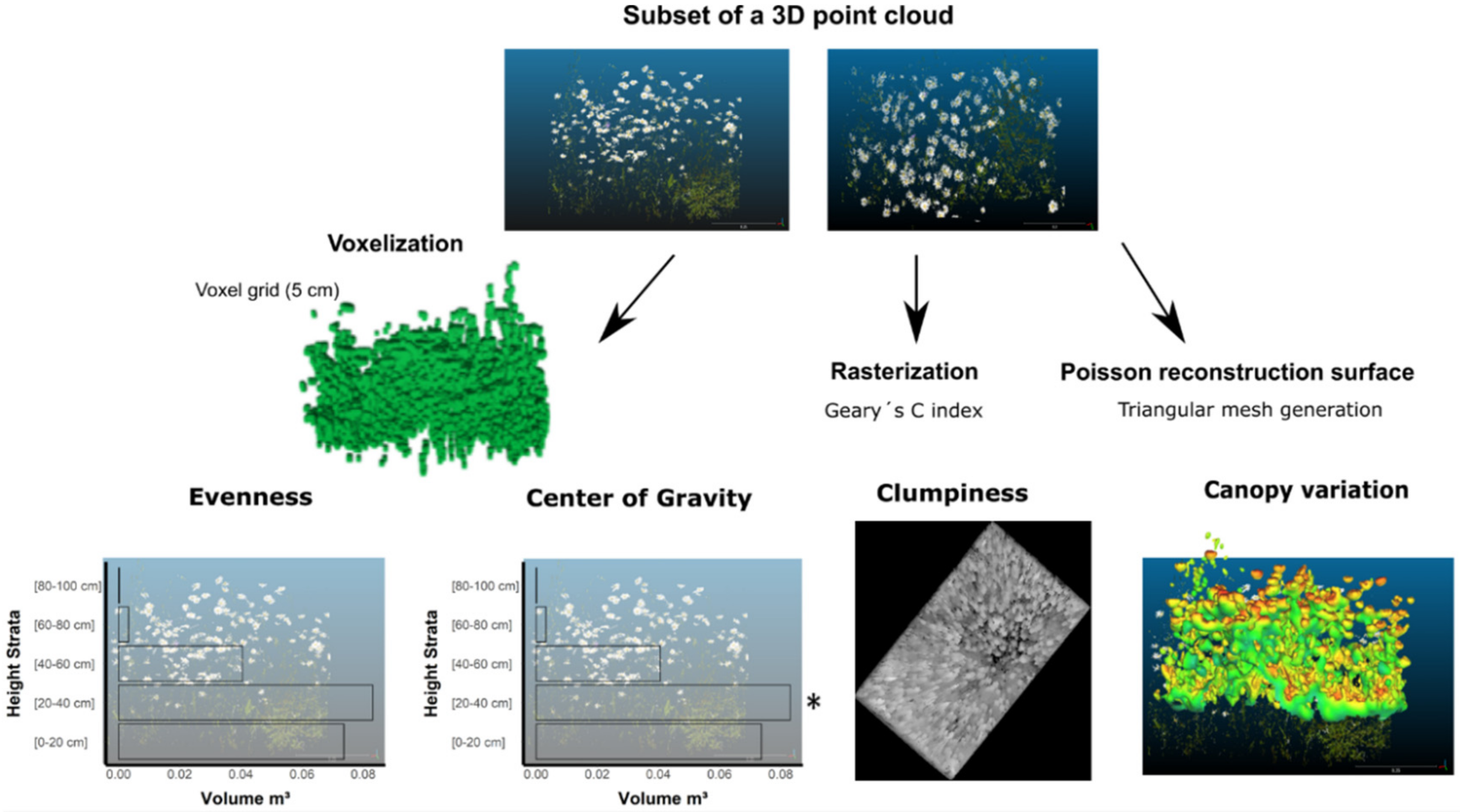
2.2. Terrestrial Laser Scanning: Data Acquisition and Processing
2.2.1. Canopy Structural Components
2.3. Data Analyses
Intra-Annual Diversity Effects on Plant Communities Canopy Structure
3. Results
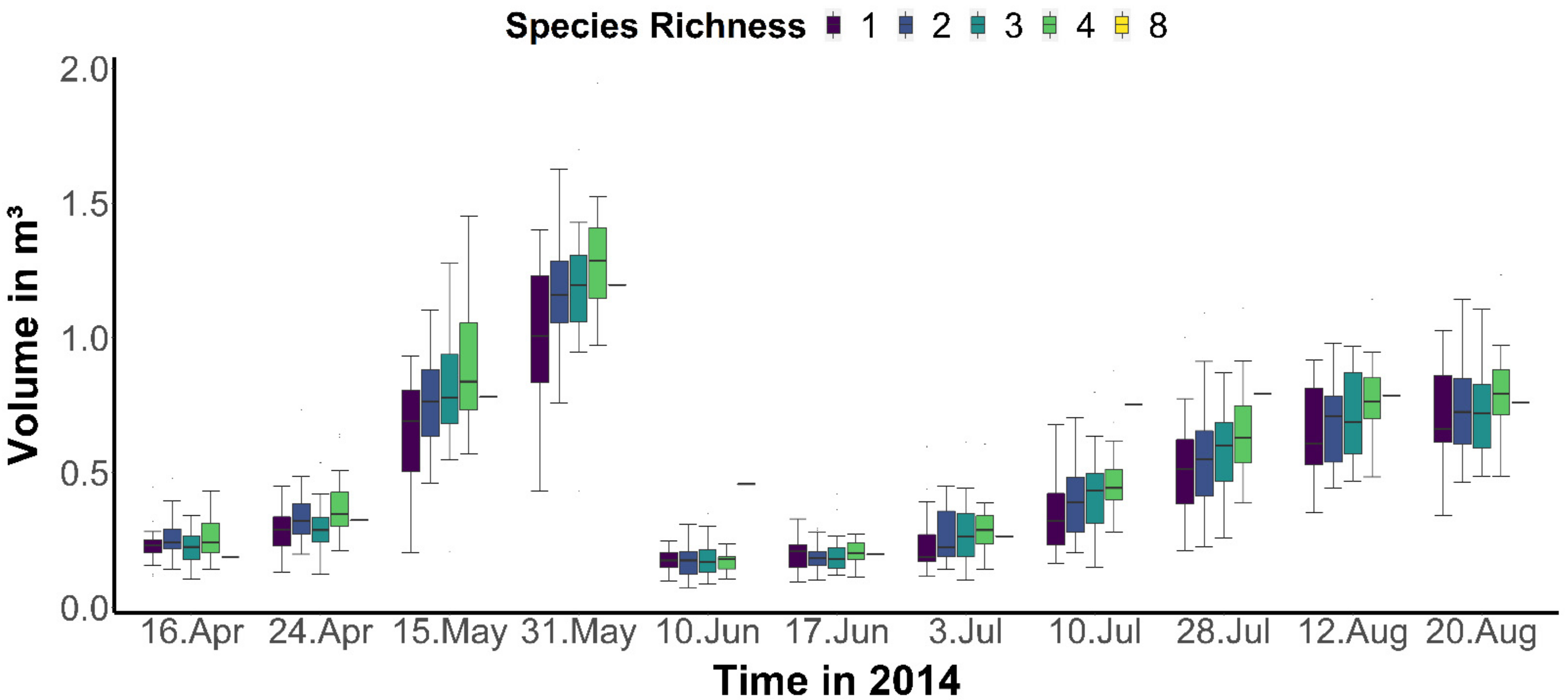
3.1. Plant Diversity Effects on Volume Distribution across the Season
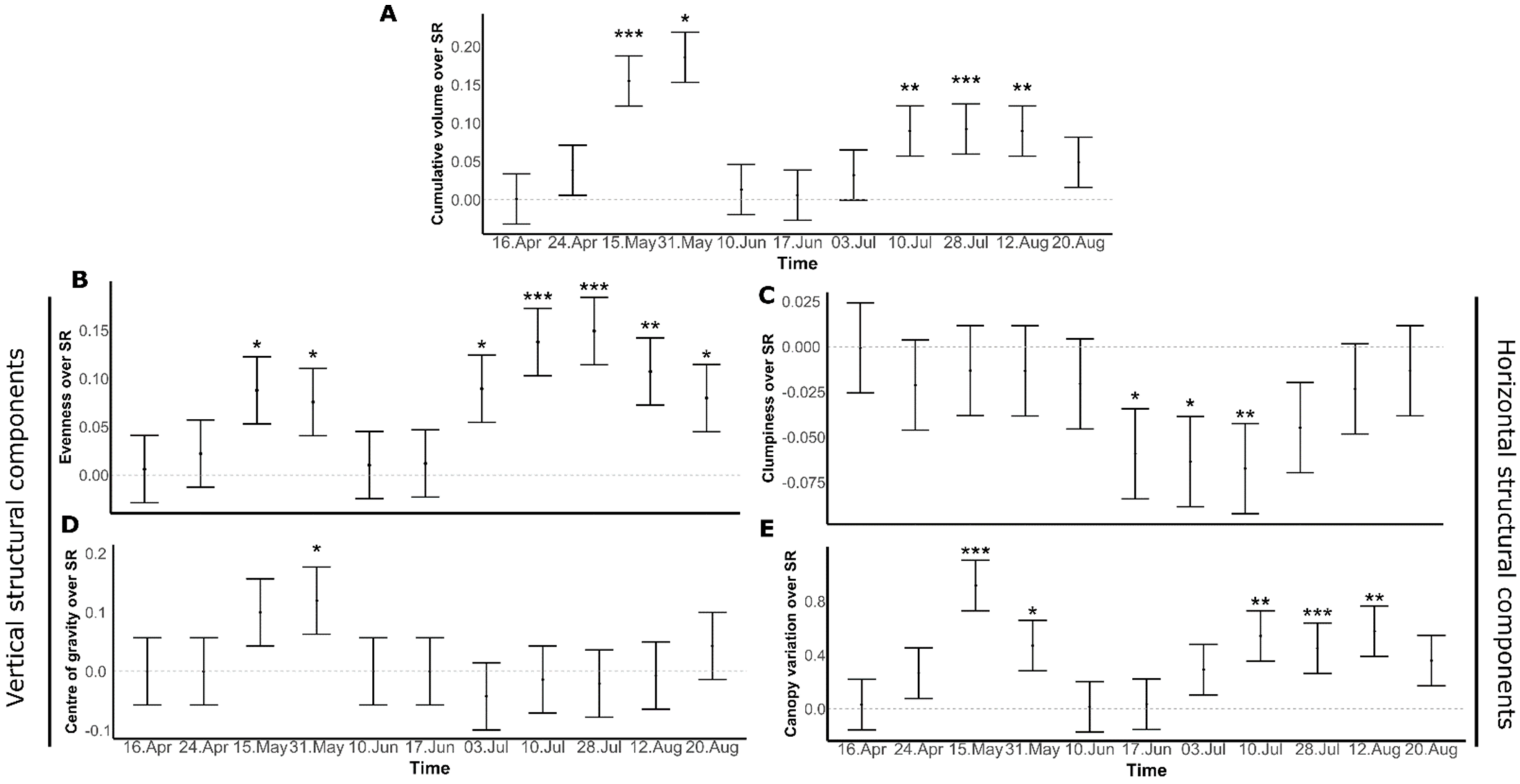
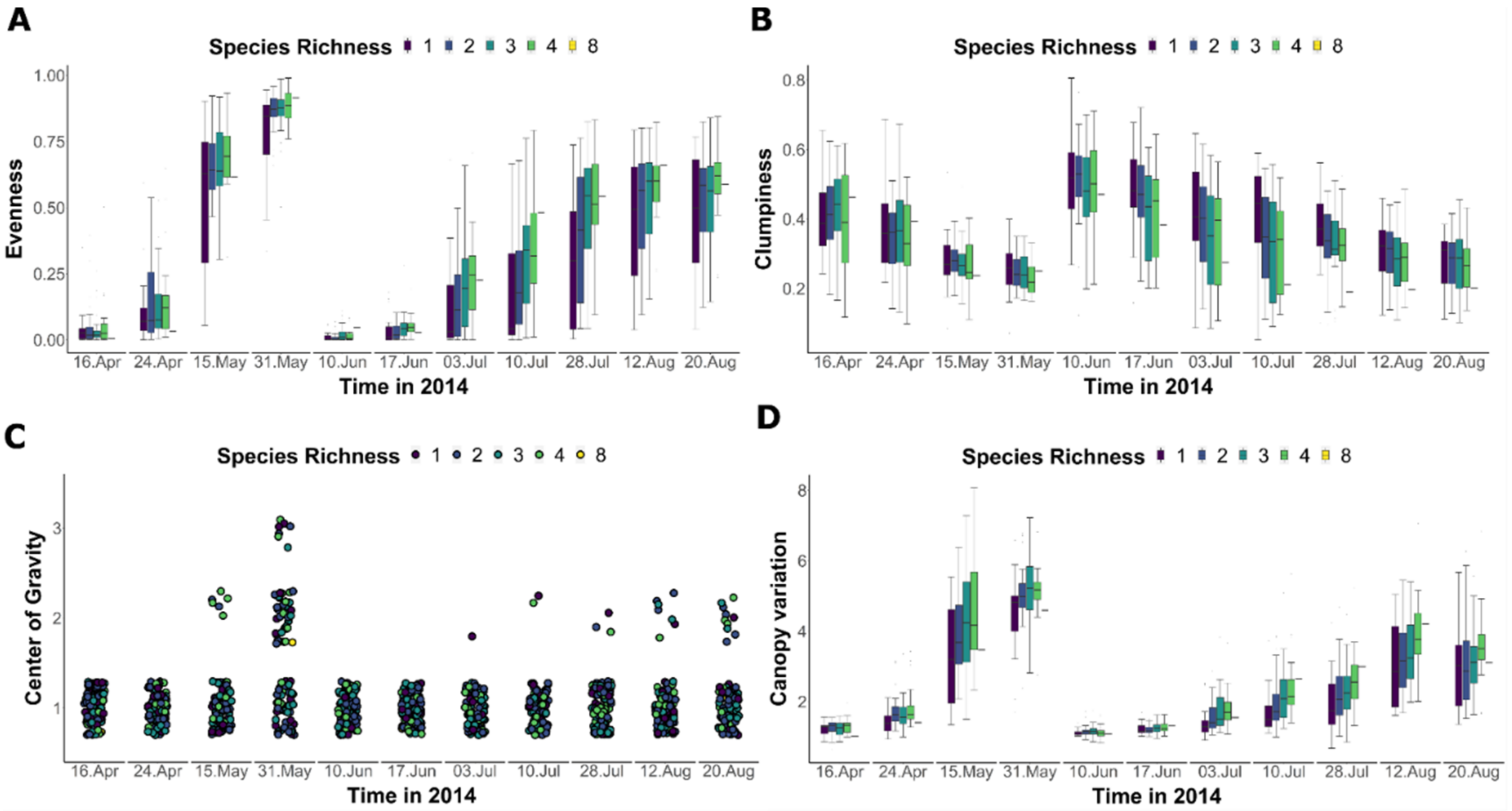
3.2. Seasonal Diversity Effects on Canopy Structural Components
4. Discussion
4.1. Diversity Effects on Vertical Metrics of Canopy Structural Components
4.2. Diversity Effects on Horizontal Components of Canopy Structure
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barry, K.E.; Weigelt, A.; van Ruijven, J.; de Kroon, H.; Ebeling, A.; Eisenhauer, N.; Gessler, A.; Ravenek, J.M.; Scherer-Lorenzen, M.; Oram, N.J.; et al. Above- and Belowground Overyielding Are Related at the Community and Species Level in a Grassland Biodiversity Experiment. In Advances in Ecological Research; Elsevier: Amsterdam, The Netherlands, 2019; Volume 61, pp. 55–89. ISBN 978-0-08-102912-1. [Google Scholar]
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; Mace, G.M.; Tilman, D.; Wardle, D.A. Biodiversity Loss and Its Impact on Humanity. Nature 2012, 486, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Lange, M.; Roth, V.; Eisenhauer, N.; Roscher, C.; Dittmar, T.; Fischer-Bedtke, C.; González Macé, O.; Hildebrandt, A.; Milcu, A.; Mommer, L.; et al. Plant Diversity Enhances Production and Downward Transport of Biodegradable Dissolved Organic Matter. J. Ecol. 2021, 109, 1284–1297. [Google Scholar] [CrossRef]
- Reich, P.B.; Tilman, D.; Isbell, F.; Mueller, K.; Hobbie, S.E.; Flynn, D.F.; Eisenhauer, N. Impacts of Biodiversity Loss Escalate through Time as Redundancy Fades. Science 2012, 336, 589–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.; Eisenhauer, N.; Ferlian, O.; Zhang, J.; Zhou, G.; Lu, X.; Liu, C.; Zhang, D. Species Richness Promotes Ecosystem Carbon Storage: Evidence from Biodiversity-Ecosystem Functioning Experiments. Proc. R. Soc. B. 2020, 287, 20202063. [Google Scholar] [CrossRef]
- Cooper, S.; Roy, D.; Schaaf, C.; Paynter, I. Examination of the Potential of Terrestrial Laser Scanning and Structure-from-Motion Photogrammetry for Rapid Nondestructive Field Measurement of Grass Biomass. Remote Sens. 2017, 9, 531. [Google Scholar] [CrossRef] [Green Version]
- Schulze-Brüninghoff, D.; Hensgen, F.; Wachendorf, M.; Astor, T. Methods for LiDAR-Based Estimation of Extensive Grassland Biomass. Comput. Electron. Agric. 2019, 156, 693–699. [Google Scholar] [CrossRef]
- Wallace, L.; Hillman, S.; Reinke, K.; Hally, B. Non-destructive Estimation of Above-ground Surface and Near-surface Biomass Using 3D Terrestrial Remote Sensing Techniques. Methods Ecol. Evol. 2017, 8, 1607–1616. [Google Scholar] [CrossRef] [Green Version]
- Guimarães-Steinicke, C.; Weigelt, A.; Ebeling, A.; Eisenhauer, N.; Duque-Lazo, J.; Reu, B.; Roscher, C.; Schumacher, J.; Wagg, C.; Wirth, C. Terrestrial Laser Scanning Reveals Temporal Changes in Biodiversity Mechanisms Driving Grassland Productivity. In Advances in Ecological Research; Elsevier: Amsterdam, The Netherlands, 2019; Volume 61, pp. 133–161. ISBN 978-0-08-102912-1. [Google Scholar]
- Schuldt, A.; Ebeling, A.; Kunz, M.; Staab, M.; Guimarães-Steinicke, C.; Bachmann, D.; Buchmann, N.; Durka, W.; Fichtner, A.; Fornoff, F.; et al. Multiple Plant Diversity Components Drive Consumer Communities across Ecosystems. Nat. Commun. 2019, 10, 1460. [Google Scholar] [CrossRef] [Green Version]
- Bachmann, D.; Roscher, C.; Buchmann, N. How Do Leaf Trait Values Change Spatially and Temporally with Light Availability in a Grassland Diversity Experiment? Oikos 2018, 127, 935–948. [Google Scholar] [CrossRef]
- Wang, R.; Gamon, J.; Montgomery, R.; Townsend, P.; Zygielbaum, A.; Bitan, K.; Tilman, D.; Cavender-Bares, J. Seasonal Variation in the NDVI–Species Richness Relationship in a Prairie Grassland Experiment (Cedar Creek). Remote Sens. 2016, 8, 128. [Google Scholar] [CrossRef] [Green Version]
- Spehn, E.M.; Joshi, J.; Schmid, B.; Diemer, M.; Korner, C. Above-Ground Resource Use Increases with Plant Species Richness in Experimental Grassland Ecosystems. Funct. Ecol. 2000, 14, 326–337. [Google Scholar]
- Anten, N.P.R.; Hirose, T. Interspecific Differences in Above-Ground Growth Patterns Result in Spatial and Temporal Partitioning of Light among Species in a Tall-Grass Meadow. J. Ecol. 1999, 87, 583–597. [Google Scholar] [CrossRef]
- Hirose, T.; Werger, M.J.A. Canopy Structure and Photon Flux Partitioning Among Species in a Herbaceous Plant Community. Ecology 1995, 76, 466–474. [Google Scholar] [CrossRef]
- Wacker, L.; Baudois, O.; Eichenberger-Glinz, S.; Schmid, B. Environmental Heterogeneity Increases Complementarity in Experimental Grassland Communities. Basic Appl. Ecol. 2008, 9, 467–474. [Google Scholar] [CrossRef] [Green Version]
- Williams, L.J.; Paquette, A.; Cavender-Bares, J.; Messier, C.; Reich, P.B. Spatial Complementarity in Tree Crowns Explains Overyielding in Species Mixtures. Nat. Ecol. Evol. 2017, 1, 0063. [Google Scholar] [CrossRef]
- Barry, K.E.; Ruijven, J.; Mommer, L.; Bai, Y.; Beierkuhnlein, C.; Buchmann, N.; Kroon, H.; Ebeling, A.; Eisenhauer, N.; Guimarães-Steinicke, C.; et al. Limited Evidence for Spatial Resource Partitioning across Temperate Grassland Biodiversity Experiments. Ecology 2020, 101, e02905. [Google Scholar] [CrossRef] [Green Version]
- Dimitrakopoulos, P.G.; Schmid, B. Biodiversity Effects Increase Linearly with Biotope Space: Diversity Effects Increase with Biotope Space. Ecol. Lett. 2004, 7, 574–583. [Google Scholar] [CrossRef]
- Lorentzen, S.; Roscher, C.; Schumacher, J.; Schulze, E.-D.; Schmid, B. Species Richness and Identity Affect the Use of Aboveground Space in Experimental Grasslands. Perspect. Plant Ecol. Evol. Syst. 2008, 10, 73–87. [Google Scholar] [CrossRef] [Green Version]
- Ratcliffe, S.; Liebergesell, M.; Ruiz-Benito, P.; Madrigal González, J.; Muñoz Castañeda, J.M.; Kändler, G.; Lehtonen, A.; Dahlgren, J.; Kattge, J.; Peñuelas, J.; et al. Modes of Functional Biodiversity Control on Tree Productivity across the European Continent: Functional Biodiversity Control on Tree Growth. Glob. Ecol. Biogeogr. 2016, 25, 251–262. [Google Scholar] [CrossRef] [Green Version]
- Walter, J.A.; Stovall, A.E.L.; Atkins, J.W. Vegetation Structural Complexity and Biodiversity in the Great Smoky Mountains. Ecosphere 2021, 12, e03390. [Google Scholar] [CrossRef]
- Marquard, E.; Weigelt, A.; Roscher, C.; Gubsch, M.; Lipowsky, A.; Schmid, B. Positive Biodiversity-Productivity Relationship Due to Increased Plant Density. J. Ecol. 2009, 97, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Hertzog, L.R.; Meyer, S.T.; Weisser, W.W.; Ebeling, A. Experimental Manipulation of Grassland Plant Diversity Induces Complex Shifts in Aboveground Arthropod Diversity. PLoS ONE 2016, 11, e0148768. [Google Scholar] [CrossRef]
- Eisenhauer, N.; Reich, P.B.; Scheu, S. Increasing Plant Diversity Effects on Productivity with Time Due to Delayed Soil Biota Effects on Plants. Basic Appl. Ecol. 2012, 13, 571–578. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Yates, S.; Mooney, H.A. Long-term data reveal complex dynamics in grassland in relation to climate and disturbance. Ecol. Monogr. 2007, 77, 545–568. [Google Scholar] [CrossRef]
- Jones, S.K.; Ripplinger, J.; Collins, S.L. Species Reordering, Not Changes in Richness, Drives Long-term Dynamics in Grassland Communities. Ecol. Lett. 2017, 20, 1556–1565. [Google Scholar] [CrossRef]
- Moorsel, S.J.; Hahl, T.; Petchey, O.L.; Ebeling, A.; Eisenhauer, N.; Schmid, B.; Wagg, C. Co-occurrence History Increases Ecosystem Stability and Resilience in Experimental Plant Communities. Ecology 2021, 102. [Google Scholar] [CrossRef]
- Weisser, W.W.; Roscher, C.; Meyer, S.T.; Ebeling, A.; Luo, G.; Allan, E.; Beßler, H.; Barnard, R.L.; Buchmann, N.; Buscot, F.; et al. Biodiversity Effects on Ecosystem Functioning in a 15-Year Grassland Experiment: Patterns, Mechanisms, and Open Questions. Basic Appl. Ecol. 2017, 23, 1–73. [Google Scholar] [CrossRef]
- Zhang, Z.; Cao, L.; She, G. Estimating Forest Structural Parameters Using Canopy Metrics Derived from Airborne LiDAR Data in Subtropical Forests. Remote Sens. 2017, 9, 940. [Google Scholar] [CrossRef] [Green Version]
- Liira, J.; Zobel, K.; Mägi, R.; Molenberghs, G. Vertical Structure of Herbaceous Canopies: The Importance of Plant Growth-Form and Species-Specific Traits. Plant Ecol. 2002, 163, 123–134. [Google Scholar] [CrossRef]
- Spehn, E.M.; Hector, A.; Joshi, J.; Scherer-Lorenzen, M.; Schmid, B.; Bazeley-White, E.; Beierkuhnlein, C.; Caldeira, M.C.; Diemer, M.; Dimitrakopoulos, P.G.; et al. Ecosystem Effects of Biodiversity Manipulations in European Grasslands. Ecol. Monogr. 2005, 75, 37–63. [Google Scholar]
- Leger, E.A.; Espeland, E.K. The Shifting Balance of Facilitation and Competition Affects the Outcome of Intra- and Interspecific Interactions over the Life History of California Grassland Annuals. Plant Ecol. 2010, 208, 333–345. [Google Scholar] [CrossRef]
- Guimarães-Steinicke, C.; Weigelt, A.; Proulx, R.; Lanners, T.; Eisenhauer, N.; Duque-Lazo, J.; Reu, B.; Roscher, C.; Wagg, C.; Buchmann, N.; et al. Biodiversity Facets Affect Community Surface Temperature via 3D Canopy Structure in Grassland Communities. J. Ecol. 2021, 109, 1969–1985. [Google Scholar] [CrossRef]
- Atkins, J.W.; Bohrer, G.; Fahey, R.T.; Hardiman, B.S.; Morin, T.H.; Stovall, A.E.L.; Zimmerman, N.; Gough, C.M. Quantifying Vegetation and Canopy Structural Complexity from Terrestrial Li DAR Data Using the forestr r Package. Methods Ecol. Evol. 2018, 9, 2057–2066. [Google Scholar] [CrossRef]
- Ehbrecht, M.; Schall, P.; Ammer, C.; Seidel, D. Quantifying Stand Structural Complexity and Its Relationship with Forest Management, Tree Species Diversity and Microclimate. Agric. For. Meteorol. 2017, 242, 1–9. [Google Scholar] [CrossRef]
- Atkins, J.W.; Fahey, R.T.; Hardiman, B.H.; Gough, C.M. Forest Canopy Structural Complexity and Light Absorption Relationships at the Subcontinental Scale. J. Geophys. Res. Biogeosci. 2018, 123, 1387–1405. [Google Scholar] [CrossRef]
- Stark, S.C.; Enquist, B.J.; Saleska, S.R.; Leitold, V.; Schietti, J.; Longo, M.; Alves, L.F.; Camargo, P.B.; Oliveira, R.C. Linking Canopy Leaf Area and Light Environments with Tree Size Distributions to Explain Amazon Forest Demography. Ecol. Lett. 2015, 18, 636–645. [Google Scholar] [CrossRef]
- Hardiman, B.S.; Gough, C.M.; Halperin, A.; Hofmeister, K.L.; Nave, L.E.; Bohrer, G.; Curtis, P.S. Maintaining High Rates of Carbon Storage in Old Forests: A Mechanism Linking Canopy Structure to Forest Function. For. Ecol. Manag. 2013, 298, 111–119. [Google Scholar] [CrossRef]
- Ebeling, A.; Pompe, S.; Baade, J.; Eisenhauer, N.; Hillebrand, H.; Proulx, R.; Roscher, C.; Schmid, B.; Wirth, C.; Weisser, W.W. A Trait-Based Experimental Approach to Understand the Mechanisms Underlying Biodiversity–Ecosystem Functioning Relationships. Basic Appl. Ecol. 2014, 15, 229–240. [Google Scholar] [CrossRef] [Green Version]
- Roscher, C.; Schumacher, J.; Baade, J.; Wilcke, W.; Gleixner, G.; Weisser, W.W.; Schmid, B.; Schulze, E.-D. The Role of Biodiversity for Element Cycling and Trophic Interactions: An Experimental Approach in a Grassland Community. Basic Appl. Ecol. 2004, 5, 107–121. [Google Scholar] [CrossRef]
- Bongers, F.J.; Schmid, B.; Bruelheide, H.; Bongers, F.; Li, S.; von Oheimb, G.; Li, Y.; Cheng, A.; Ma, K.; Liu, X. Functional Diversity Effects on Productivity Increase with Age in a Forest Biodiversity Experiment. Nat. Ecol. Evol. 2021, 5, 1594–1603. [Google Scholar] [CrossRef]
- Zuppinger-Dingley, D.; Schmid, B.; Petermann, J.S.; Yadav, V.; De Deyn, G.B.; Flynn, D.F. Selection for Niche Differentiation in Plant Communities Increases Biodiversity Effects. Nature 2014, 515, 108–111. [Google Scholar] [CrossRef]
- FARO Technologies Inc. FARO Laser Scaner Focus 3D Instruction Manual; FARO Technologies Inc.: Lake Mary, FL, USA, 2011. [Google Scholar]
- SCENE User Manual. 2021. Available online: https://knowledge.faro.com/Software/FARO_SCENE/SCENE/User_Manual_for_SCENE (accessed on 16 February 2022).
- CloudCompare Omnia. 2019. Available online: https://www.cloudcompare.org/release/notes/20200614/ (accessed on 16 February 2022).
- Attene, M.; Spagnuolo, M. Automatic Surface Reconstruction from Point Sets in Space. Comput. Graph. Forum 2000, 19, 457–465. [Google Scholar] [CrossRef]
- Kazhdan, M.; Hoppe, H. An Adaptive Multi-Grid Solver for Applications in Computer Graphics. Comput. Graph. Forum 2019, 38, 138–150. [Google Scholar] [CrossRef] [Green Version]
- Geary, R.C. The Contiguity Ratio and Statistical Mapping. Inc. Stat. 1954, 5, 115. [Google Scholar] [CrossRef]
- Pinheiro, J.C.; Bates, D.M. Mixed-Effects Models in Sand S-PLUS. In Statistics and Computing; Springer New York: New York, NY, USA, 2000; ISBN 978-1-4419-0317-4. [Google Scholar]
- Chi, E.M.; Reinsel, G.C. Models for Longitudinal Data with Random Effects and AR(1) Errors. J. Am. Stat. Assoc. 1989, 84, 452–459. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R. In Statistics for Biology and Health; Springer: New York, NY, USA, 2009; ISBN 978-0-387-87457-9. [Google Scholar]
- Milcu, A.; Roscher, C.; Gessler, A.; Bachmann, D.; Gockele, A.; Guderle, M.; Landais, D.; Piel, C.; Escape, C.; Devidal, S.; et al. Functional Diversity of Leaf Nitrogen Concentrations Drives Grassland Carbon Fluxes. Ecol. Lett. 2014, 17, 435–444. [Google Scholar] [CrossRef]
- Milcu, A.; Gessler, A.; Roscher, C.; Rose, L.; Kayler, Z.; Bachmann, D.; Pirhofer-Walzl, K.; Zavadlav, S.; Galiano, L.; Buchmann, T.; et al. Top Canopy Nitrogen Allocation Linked to Increased Grassland Carbon Uptake in Stands of Varying Species Richness. Sci. Rep. 2017, 7, 8392. [Google Scholar] [CrossRef]
- Wacker, L.; Baudois, O.; Eichenberger-Glinz, S.; Schmid, B. Effects of Plant Species Richness on Stand Structure and Productivity. J. Plant Ecol. 2009, 2, 95–106. [Google Scholar] [CrossRef]
- Anten, N.P.R. Optimal Photosynthetic Characteristics of Individual Plants in Vegetation Stands and Implications for Species Coexistence. Ann. Bot. 2004, 95, 495–506. [Google Scholar] [CrossRef] [Green Version]
- Hikosaka, K.; Anten, N.P.R. An Evolutionary Game of Leaf Dynamics and Its Consequences for Canopy Structure. Funct Ecol 2012, 26, 1024–1032. [Google Scholar] [CrossRef]
- Fridley, J.D. The Influence of Species Diversity on Ecosystem Productivity: How, Where, and Why? Oikos 2001, 93, 514–526. [Google Scholar] [CrossRef]
- Peer, L.V.; Nijs, I.; Bogaert, J.; Verelst, I.; Reheul, D. Survival, Gap Formation, and Recovery Dynamics in Grassland Ecosystems Exposed to Heat Extremes: The Role of Species Richness. Ecosystems 2001, 4, 797–806. [Google Scholar] [CrossRef] [Green Version]
- Barry, K.E.; Mommer, L.; van Ruijven, J.; Wirth, C.; Wright, A.J.; Bai, Y.; Connolly, J.; De Deyn, G.B.; de Kroon, H.; Isbell, F.; et al. The Future of Complementarity: Disentangling Causes from Consequences. Trends Ecol. Evol. 2019, 34, 167–180. [Google Scholar] [CrossRef] [Green Version]
- De Boeck, H.J.; Nijs, I.; Lemmens, C.M.H.M.; Ceulemans, R. Underlying Effects of Spatial Aggregation (Clumping) in Relationships between Plant Diversity and Resource Uptake. Oikos 2006, 113, 269–278. [Google Scholar] [CrossRef]
- Tilman, D.; Lehman, C.; Thompson, K. Plant Diversity and Ecosystem Productivity: Theoretical Considerations. Proc. Natl. Acad. Sci. USA 1997, 94, 1857–1861. [Google Scholar] [CrossRef] [Green Version]
- Aarssen, L.W. High Productivity in Grassland Ecosystems: Effected by Species Diversity or Productive Species? Oikos 1997, 80, 183–184. [Google Scholar] [CrossRef]
- Fargione, J.; Tilman, D. Plant Species Traits and Capacity for Resource Reduction Predict Yield and Abundance under Competition in Nitrogen-Limited Grassland. Funct. Ecol. 2006, 20, 533–540. [Google Scholar] [CrossRef]
- Grime, J.P. Benefits of Plant Diversity to Ecosystems: Immediate, Filter and Founder Effects. J. Ecol. 1998, 86, 902–910. [Google Scholar] [CrossRef]
- Turnbull, L.A.; Coomes, D.A.; Purves, D.W.; Rees, M. How Spatial Structure Alters Population and Community Dynamics in a Natural Plant Community. J. Ecol. 2007, 95, 79–89. [Google Scholar] [CrossRef]
- Proulx, R.; Roca, I.T.; Cuadra, F.S.; Seiferling, I.; Wirth, C. A Novel Photographic Approach for Monitoring the Structural Heterogeneity and Diversity of Grassland Ecosystems. J. Plant Ecol. 2014, 7, 518–525. [Google Scholar] [CrossRef] [Green Version]
- Eisenhauer, N.; Herrmann, S.; Hines, J.; Buscot, F.; Siebert, J.; Thakur, M.P. The Dark Side of Animal Phenology. Trends Ecol. Evol. 2018, 33, 898–901. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Df | Chisq | Pr (>Chisq) | |
|---|---|---|---|
| Time | 11 | 10,122.029 | 2.2 × 10−16 |
| Time × SR | 11 | 69.421 | 1.574 × 10−7 |
| Evenness | Clumpiness | ||||||
|---|---|---|---|---|---|---|---|
| Df | Chisq | Pr (>Chisq) | Df | Chisq | Pr (>Chisq) | ||
| time | 11 | 9022.90 | <2.2 × 10−16 *** | time | 11 | 2419.695 | 2 × 10−16 *** |
| SR × time | 11 | 39.53 | 4.301 × 10−5 *** | SR × time | 11 | 14.373 | 0.213 |
| Center of Gravity | Canopy variation | ||||||
| Df | Chisq | Pr (>Chisq) | Df | Chisq | Pr (>Chisq) | ||
| time | 11 | 6038.5361 | <2 ×10−16 *** | time | 11 | 4740.987 | <2.2 × 10−16 *** |
| SR × time | 11 | 8.3985 | 0.6772 | SR × time | 11 | 47.521 | 1.738 × 10−6 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guimarães-Steinicke, C.; Weigelt, A.; Ebeling, A.; Eisenhauer, N.; Wirth, C. Diversity Effects on Canopy Structure Change throughout a Growing Season in Experimental Grassland Communities. Remote Sens. 2022, 14, 1557. https://0-doi-org.brum.beds.ac.uk/10.3390/rs14071557
Guimarães-Steinicke C, Weigelt A, Ebeling A, Eisenhauer N, Wirth C. Diversity Effects on Canopy Structure Change throughout a Growing Season in Experimental Grassland Communities. Remote Sensing. 2022; 14(7):1557. https://0-doi-org.brum.beds.ac.uk/10.3390/rs14071557
Chicago/Turabian StyleGuimarães-Steinicke, Claudia, Alexandra Weigelt, Anne Ebeling, Nico Eisenhauer, and Christian Wirth. 2022. "Diversity Effects on Canopy Structure Change throughout a Growing Season in Experimental Grassland Communities" Remote Sensing 14, no. 7: 1557. https://0-doi-org.brum.beds.ac.uk/10.3390/rs14071557