
Assessing Drought Response in the Southwestern Amazon Forest by Remote Sensing and In Situ Measurements

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. RJA Climatology

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data | Equipament/Metodology | Period | Source |
|---|---|---|---|
| MODIS/MAIAC | MODIS | 2000–2017 | [50] |
| Evapotranspiration (mm) | MOD16A2 | 2001–2017 | [51] |
| Drought severity | Standard precipitation index (SPI-6) | 1983–2017 | [52,53] |
| Precipitation (mm) | Reanalysis | 1983–2017 | [54] |
| Air temperature (°C) | Sensor: 107 Campbell Scientific Inc. | 2004–2017 | LBA |
| Net radiation (W·m2) | CM-21, Kipp&Zonen | 2005–2017 | LBA |
| Soil moisture (%) | CS615 sensor, Campbell Scientific Inc., installed in 10 cm; 20 cm; 30 cm; 40 cm; 60 cm and 100 cm deep. | 2015–2017 | LBA |
| Water table depth (m) | Piezometer | May 2016–Dec 2017 | ** |
| Total Litter (Mg·ha−1) | Colectors (0.50 × 0.50 m) | May 2016–Jan 2018 | ** |
| Stem growth (cm) | Dendrometers ZN12-T-2IP; ZWEIFEL et al. (2005) metodology. | Sep. 2015–Ago 2017 | ** |
| P50 (MPa) | Pereira et al. (2016) metodology; Choat et al. (2012) limiar. | Oct. 2016 | ** |
| πTLP (Turgor Loss Point) (MPa) | Tyree e Hammel (1972) and Sack e Pasquet-Kok (2011) metodology; Bartlett et al. (2012) limiar | Jul. 2017 | ** |
| Isohydricity | PMS Instruments Co., Albany, NY, USA; Bartlett et al. (2012) and Choat et al. (2012) pattern. | Oct. 2016–Jul. 2017 | ** |
| Hydraulic safety margin (HSM) | Choat et al. (2012) pattern | Oct. 2016–Jul. 2017 | ** |
2.3. Study Plot and Plant Species
2.4. Field Measurements
2.5. Remote Sensing Products
3. Results
3.1. Seasonal Variability of Field Observations and Remote Sensing Data
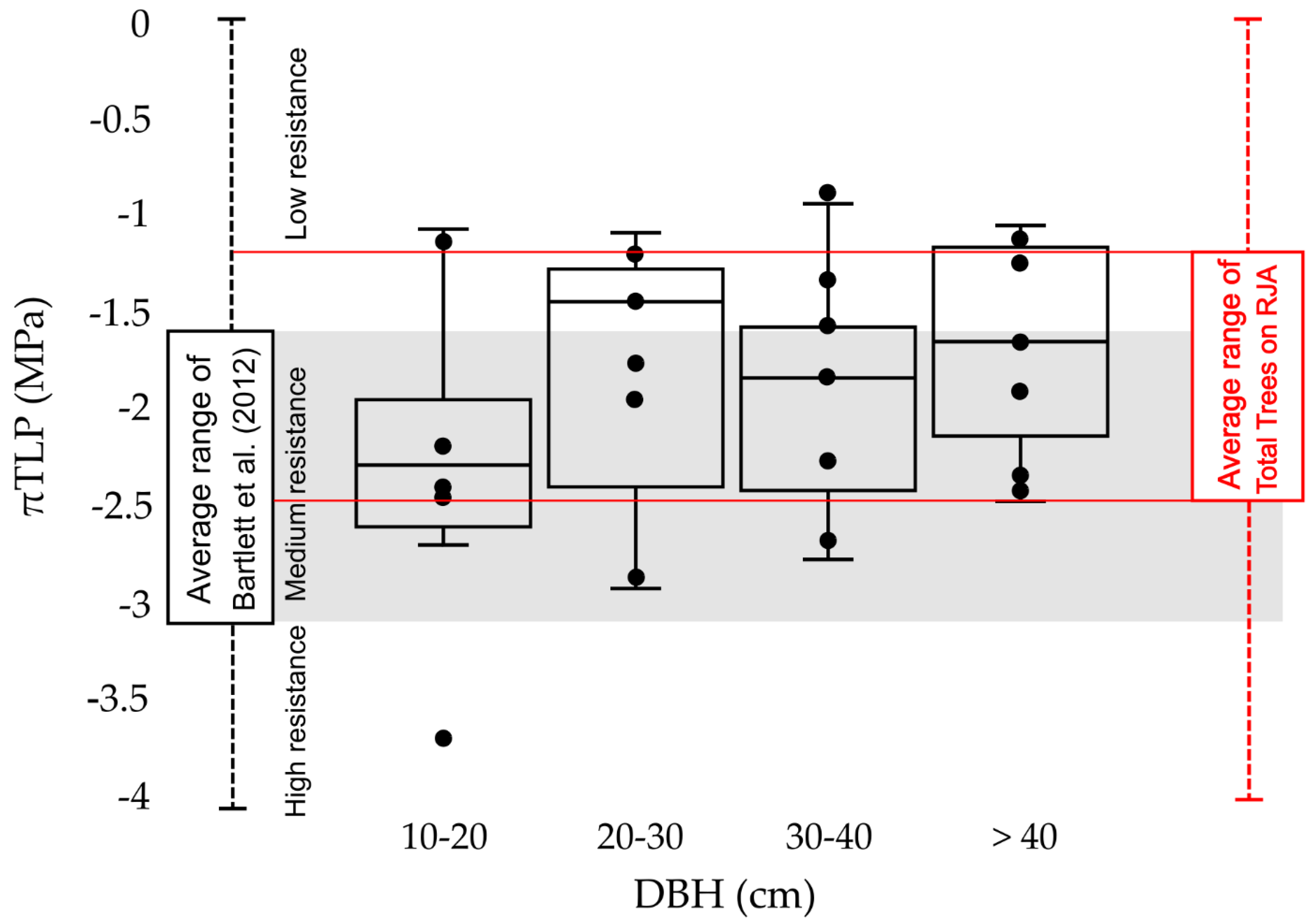
3.2. Drought Tolerance
4. Discussion
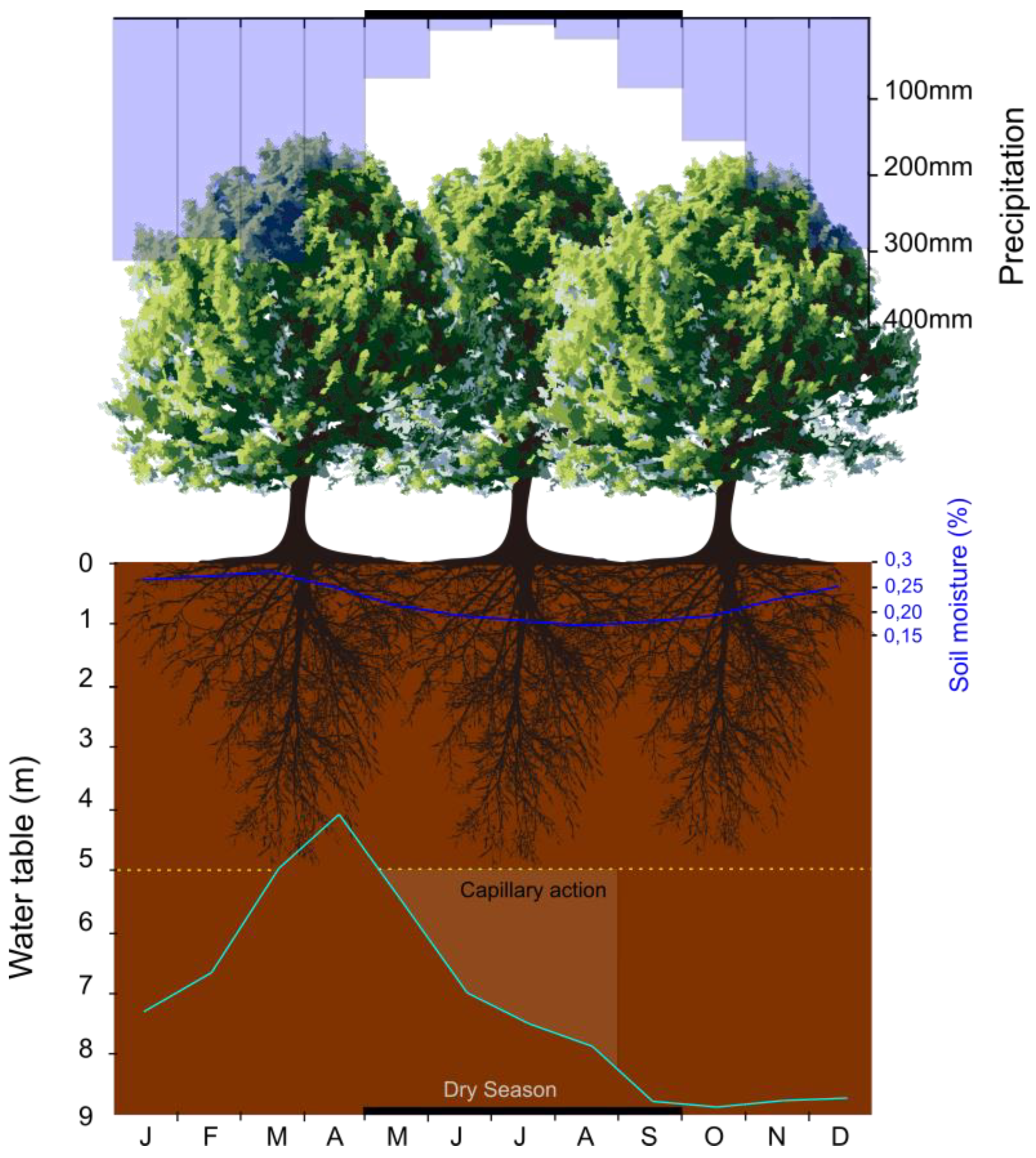
4.1. How Is the Seasonal Dynamics of the Forest in the Southwest Portion of the Amazon?
4.2. What Are The Forest Mechanisms to Resist Seasonal Drought?
4.3. Does Remote Sensing Have the Potential to Represent the Phenological and Physioloagical Changes of the Forest?
5. Conclusions
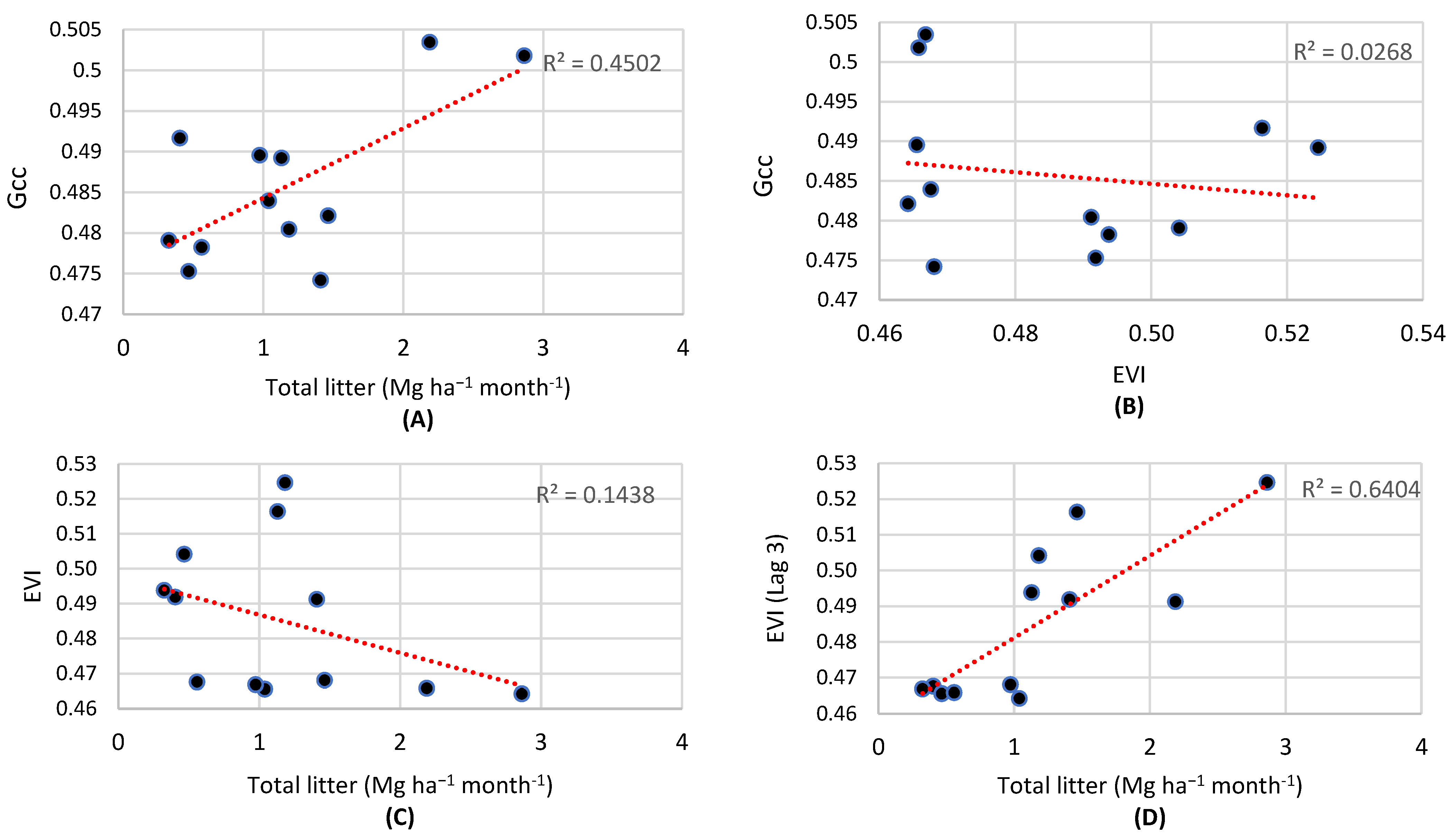
- EVI and Gcc are sensitive to different responses of the forest canopy structure. During the dry season, the forest reduces its photosynthetic activity even as it renews its leaves at the same time, a browning effect. During the wet season, photosynthetic activity increases, with a greening effect, revealing the relationship of this response with leaf maturity.
- The pattern of vulnerability to drought does not follow that of other Amazonian sites. Trees with larger stem diameters are more resistant than trees with thinner stem diameters.
- Negative effects on stem growth post-El-Niño 2015/2016 were observed, suggesting that the persistence of negative rainfall anomalies may be very critical for the forest, regardless of whether extreme drought is influenced by changes in ocean temperature.
- It is expected that precipitation is climatic forcing with the greatest influence on both the stem growth and the increase in photosynthetic activity of the canopy.
- In situ ecophysiological data are essential for this type of analysis; however, they are scarce, difficult to collect and have limited temporal resolution. Satellites provide a practical method to assess forest dynamics, at appropriate scales, to which this research sought to contribute.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, M.; Running, S.W. Drought-induced reduction in global terrestrial net primary production from 2000 through 2009. Science 2010, 329, 940–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malhi, Y.; Wood, D.; Baker, T.R.; Wright, J.; Phillips, O.L.; Cochrane, T.; Meir, P.; Chave, J.; Almeida, S.; Arroyo, L.; et al. The regional variation of aboveground live biomass in old-growth Amazonian forests. Glob. Chang. Biol. 2006, 12, 1107–1138. [Google Scholar] [CrossRef]
- Wagner, F.H.; Rossi, V.; Aubry-Kientz, M.; Bonal, D.; Dalitz, H.; Gliniars, R.; Stahl, C.; Trabucco, A.; Hérault, B. Pan-tropical analysis of climate effects on seasonal tree growth. PLoS ONE 2014, 9, e92337. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.H.; Foley, J.A. Combined effects of deforestation and doubled atmospheric CO2 concentrations on the climate of Amazonia. J. Clim. 2000, 13, 18–34. [Google Scholar] [CrossRef]
- Li, W.; Fu, R.; Négron-Juárez, R.I.; Fernandes, K. Observed change of the standardized precipitation index, its potential cause and implications to future climate change in the Amazon region. Biol. Sci. 2008, 363, 1767–1772. [Google Scholar] [CrossRef] [PubMed]
- Gloor, M.; Barichivich, J.; Ziv, G.; Brienen, R.; Schöngart, J.; Peylin, P.; Cintra, B.B.L.; Feldpausch, T.; Phillips, O.; Baker, J. Recent Amazon climate as background for possible ongoing and future changes of Amazon humid forests. Glob. Biogeochem. Cycles 2015, 29, 1384–1399. [Google Scholar] [CrossRef]
- Moura, V.; Souza, R.D.A.D.; Mercante, E.; Richetti, J.; Johann, J.A. Three Decades after: Landscape Dynamics in Different Colonisation Models Implemented in the Brazilian Legal Amazon. Remote Sens. 2021, 13, 4581. [Google Scholar] [CrossRef]
- Jiménez-Muñoz, J.C.; Mattar, C.; Barichivich, J.; Santamaría-Artigas, A.; Takahashi, K.; Malhi, Y.; Sobrino, J.A.; van der Schrier, G. Record-breaking warming and extreme drought in the Amazon rainforest during the course of El Niño 2015–2016. Sci. Rep. 2016, 6, 33130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, S.L.; Brando, P.M.; Phillips, O.L.; Van Der Heijden, G.M.F.; Nepstad, D. The 2010 Amazon drought. Science 2011, 331, 554. [Google Scholar] [CrossRef]
- Marengo, J.A.; Espinoza, J.C. Extreme seasonal droughts and floods in Amazonia: Causes, trends and impacts. Int. J. Climatol. 2016, 36, 1033–1050. [Google Scholar] [CrossRef]
- Marengo, J.A.; Nobre, C.A.; Tomasella, J.; Oyama, M.D.; Oliveira, G.S.; Oliveira, R.; Camargo, H.; Alves, L.M.; Brown, F. The drought of Amazonia in 2005. J. Clim. 2008, 21, 495–516. [Google Scholar] [CrossRef] [Green Version]
- Erfanian, A.; Wang, G.; Fomenko, L. Unprecedented drought over tropical South America in 2016: Significantly under-predicted by tropical SST. Sci. Rep. 2017, 7, 5811. [Google Scholar] [CrossRef] [Green Version]
- Cox, P.M.; Betts, R.A.; Collins, M.; Harris, P.P.; Huntingford, C.; Jones, C.D. Amazonian forest dieback under climate-carbon cycle projections for the 21st century. Theor. Appl. Climatol. 2004, 78, 137–156. [Google Scholar] [CrossRef]
- Fearnside, P.M.A. Vulnerabilidade da floresta amazônica perante as mudanças climáticas. Oecologia Bras. 2009, 13, 609–618. [Google Scholar] [CrossRef] [Green Version]
- Nobre, C.A.; Sellers, P.J.; Shukla, J. Amazonian deforestation and global climate change. J. Clim. 1991, 4, 957–988. [Google Scholar] [CrossRef] [Green Version]
- Shuttleworth, W. Evaporation from Amazonian rainforest. Biol. Sci. 1988, 233, 321–346. [Google Scholar] [CrossRef]
- Von Randow, C.; Manzi, A.O.; Kruijt, B.; de Oliveira, P.J.; Zanchi, F.B.; Silva, R.L.; Hodnett, M.G.; Gash, J.H.C.; Elbers, J.A.; Waterloo, M.J.; et al. Comparative measurements and seasonal variations in energy and carbon exchange over forest and pasture in South West Amazonia. Theor. Appl. Climatol. 2004, 78, 5–26. [Google Scholar] [CrossRef]
- Da Rocha, H.R.; Manzi, A.O.; Cabral, O.M.; Miller, S.D.; Goulden, M.L.; Saleska, S.R.; R.-Coupe, N.; Wofsy, S.C.; Borma, L.S.; Artaxo, P.; et al. Patterns of water and heat flux across a biome gradient from tropical forest to savanna in Brazil. J. Geophys. Res. 2009, 114, 1–8. [Google Scholar] [CrossRef]
- Restrepo-Coupe, N.; da Rocha, H.R.; Hutyra, L.R.; Araujo, A.C.; Borma, L.S.; Christiffersen, B.; Cabral, O.M.R.; Camargo, P.B.; Cardoso, F.L.; Costa, A.C.L.; et al. What drives the seasonality of photosynthesis across the amazon basin? a cross-site analysis of eddy flux tower measurements from the Brasil flux network. Agric. For. Meteorol. 2013, 182, 128–144. [Google Scholar] [CrossRef] [Green Version]
- Saleska, S.R.; Didan, K.; Huete, A.R.; da Rocha, H.R. Amazon Forests Green-Up during 2005 Drought. Nature 2007, 318, 612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleska, S.R.; Wu, J.; Guan, K.; Araujo, A.C.; Huete, A.; Nobre, A.D.; Restrepo-Coupe, N. Dry-season greening of Amazon forests. Nature 2016, 531, E4–E5. [Google Scholar] [CrossRef] [PubMed]
- Asner, G.P.; Alencar, A. Drought impacts on the Amazon forest: The remote sensing perspective. New Phytol. 2010, 187, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Samanta, A.; Ganguly, S.; Hashimoto, H.; Devadiga, S.; Vermote, E.; Knyazikhin, Y.; Nemani, R.R.; Myneni, R.B. Amazon forests did not green-up during the 2005 drought. Geophys. Res. Lett. 2010, 37, 5. [Google Scholar] [CrossRef]
- Morton, D.C.; Nagol, J.; Carabajal, C.C.; Rosette, J.; Palace, M.; Cook, B.D.; Vermote, E.F.; Harding, D.J.; North, P.R.J. Amazon forests maintain consistent canopy structure and greenness during the dry season. Nature 2014, 506, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Restrepo-Coupe, N.; Levine, N.M.; Christoffersen, B.O.; Albert, L.P.; Wu, J.; Costa, M.H.; Galbraith, D.; Imbuzeiro, H.; Martins, G.; Araujo, A.G.; et al. Do dynamic global vegetation models capture the seasonality of carbon fluxes in the Amazon basin? A data-model intercomparison. Glob. Chang. Biol. 2016, 23, 191–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, O.L.; Aragao, L.E.O.C.; Lewis, S.L.; Fisher, J.B.; Lloyd, J.; López-González, G.; Malhi, Y.; Monteagudo, A.; Peacock, J.; Quesada, C.A.; et al. Drought sensitivity of the Amazon rainforest. Science 2009, 323, 1344–1347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, O.L.; van der Heijden, G.; Lewis, S.L.; López-González, G.; Aragao, L.E.O.C.; Lloyd, J.; Malhi, Y.; Monteagudo, A.; Almeida, S.; Dávila, E.A.; et al. Drought–mortality relationships for tropical forests. New Phytol. 2010, 187, 631–646. [Google Scholar] [CrossRef] [Green Version]
- Williamson, G.B.; Laurance, W.F.; Oliveira, A.A.; Delamônica, P.; Gascon, C.; Lovejoy, T.E.; Pohl, L. Amazonian tree mortality during the 1997 El Nino drought. Conserv. Biol. 2000, 14, 1538–1542. [Google Scholar] [CrossRef] [Green Version]
- Doughty, C.E.; Malhi, Y.; Araujo-Murakami, A.; Metcalfe, D.B.; Silva-Espejo, J.E.; Arroyo, L.; Heredia, J.P.; Pardo-Toledo, E.; Mendizabal, L.M.; Rojas-Landivar, V.D.; et al. Allocation trade-offs dominate the response of tropical forest growth to seasonal and interannual drought. Ecology 2014, 95, 1–6. [Google Scholar] [CrossRef]
- Janssen, T.; Fleischer, K.; Luyssaert, S.; Naudts, K.; Dolman, H. Drought resistance increases from the individual to the ecosystem level in highly diverse Neotropical rainforest: A meta-analysis of leaf, tree and ecosystem responses to drought. Biogeosciences 2020, 17, 2621–2645. [Google Scholar] [CrossRef]
- Janssen, T.; van der Velde, Y.; Hofhansl, F.; Luyssaert, S.; Naudts, K.; Driessen, B.; Fleischer, K.; Dolman, H. Drought effects on leaf fall, leaf flushing and stem growth in the Amazon forest: Reconciling remote sensing data and field observations. Biogeosciences 2021, 18, 4445–4472. [Google Scholar] [CrossRef]
- Naeem, S.; Bunker, D.E.; Hector, A.; Loreau, M.; Perrings, C. Biodiversity, Ecosystem Functioning, and Human Wellbeing: An Ecological and Economic Perspective; Oxford University Press: Oxford, UK, 2009; p. 388. [Google Scholar] [CrossRef] [Green Version]
- Hubau, W.; Lewis, S.L.; Phillips, O.L.; Affum-Baffoe, K.; Beeckman, H.; Cuní-Sanchez, A.; Daniels, A.K.; Ewango, C.E.N.; Fauset, S.; Mukinzi, M.; et al. Asynchronous carbon sink saturation in African and Amazonian tropical forests. Nature 2019, 579, 80–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacifici, M.; Foden, W.B.; Visconti, P.; Watson, J.E.; Butchart, S.H.; Kovacs, K.M.; Cheffers, B.R.; Hole, D.G.; Martin, T.G.; Akçakaya, H.R.; et al. Assessing species vulnerability to climate change. Nat. Clim. Chang. 2015, 5, 215–224. [Google Scholar] [CrossRef]
- Kissling, W.D.; Walls, R.; Bowser, A.; Jones, M.O.; Kattge, J.; Agosti, D.; Amengual, J.; Basset, A.; van Bodegom, P.M.; Cornelissen, J.H.C.; et al. Towards global data products of essential biodiversity variables on species traits. Nat. Ecol. Evol. 2018, 2, 1531–1540. [Google Scholar] [CrossRef] [Green Version]
- Marechaux, I.; Bartlett, M.K.; Sack, L.; Baraloto, C.; Engel, J.; Joetzjer, E.; Chave, J. Drought tolerance as predicted by leaf water potential at turgor loss point varies strongly across species within an Amazonian forest. Funct. Ecol. 2015, 29, 1268–1277. [Google Scholar] [CrossRef]
- Konings, A.; Gentine, P. Global variations in ecosystem-scale isohydricity. Glob. Chang. Biol. 2016, 23, 891–905. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, M.; Scoffon, C.; Sack, L. The determinants of leaf turgor loss point and prediction of drought tolerance of species and biomes: A global meta-analysis. Ecol. Lett. 2012, 15, 393–405. [Google Scholar] [CrossRef]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Field, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global convergence in the vulnerability of forests to drought. Nature 2012, 491, 752–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, M.L. Comparison of simulated hyperspectral HyspIRI and multispectral Landsat 8 and Sentinel-2 imagery for multi-seasonal, regional land-cover mapping. Remote Sens. Environ. 2017, 200, 311–325. [Google Scholar] [CrossRef]
- Wallis, C.I.; Homeier, J.; Peña, J.; Brandl, R.; Farwig, N.; Bendix, J. Modeling tropical montane forest biomass, productivity and canopy traits with multispectral remote sensing data. Remote Sens. Environ. 2019, 225, 77–92. [Google Scholar] [CrossRef]
- Aguirre-Gutiérrez, J.; Rifai, S.; Shenkin, A.; Oliveras, I.; Bentley, L.P.; Svátek, M.; Giradin, C.A.J.; Both, S.; Riutta, T.; Berenguer, E.; et al. Pantropical modelling of canopy functional traits using Sentinel-2 remote sensing data. Remote Sens. Environ. 2021, 252, 112122. [Google Scholar] [CrossRef]
- Herrmann, I.; Berger, K. Remote and Proximal Assessment of Plant Traits. Remote Sens. 2021, 13, 1893. [Google Scholar] [CrossRef]
- Ma, X.; Migliavacca, M.; Wirth, C.; Bohn, F.J.; Huth, A.; Richter, R.; Mahecha, M.D. Monitoring Plant Functional Diversity Using the Reflectance and Echo from Space. Remote Sens. 2020, 12, 1248. [Google Scholar] [CrossRef] [Green Version]
- IPCC-SREX, Intergovernmental Panel on Climate Change. Managing the Risks of Extreme Events and Disasters to Advance Climate Change Adaptation: Special Report of the Intergovernmental Panel on Climate Change. 2012. Available online: https://www.ipcc.ch/pdf/special-reports/srex/SREX_Full_Report.pdf (accessed on 10 April 2021).
- Barros, F.V. Hydraulic Functioning and Drought Vulnerability of Two Tropical Forests. Ph.D. Thesis, Universidade Estadual de Campinas, Campinas, Brazil, 2017; 146p. [Google Scholar]
- Michiles, A.A.S. Taxas de Armazenamento Térmico na Biomassa e Balanço de Energia em Superfície para Áreas de Floresta de Terra Firme na Amazônia. Ph.D. Thesis, Instituto Nacional de Pesquisas Espaciais, São José dos Campos, Brazil, 2009; 185p. [Google Scholar]
- Sombroek, W. Spatial and Temporal Patterns of Amazon Rainfall. AMBIO A J. Hum. Environ. 2001, 30, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Tian, H.; Pan, S.; Chen, G.; Zhang, B.; Dangal, S. Amazon droughts and forest responses: Largely reduced forest photosynthesis but slightly increased canopy greenness during the extreme drought of 2015/2016. Glob. Chang. Biol. 2018, 24, 1919–1934. [Google Scholar] [CrossRef] [PubMed]
- Dalagnol, R.; Wagner, F.H.; Galvao, L.S.; Nelson, B.W.; Aragao, L.E.O.C. Life cycle of bamboo in southwestern Amazon and its relation to fire events. Biogeosciences Discuss 2018, 15, 6087–6104. [Google Scholar] [CrossRef] [Green Version]
- USGS. MOD16A2v006 Description. 2021. Available online: https://lpdaac.usgs.gov/products/mod16a2v006/ (accessed on 11 October 2021).
- NASA(National Aeronautics and Space Administration). Standardized Precipitation Index (SPI). 2018. Available online: https://gmao.gsfc.nasa.gov/research/subseasonal/atlas/SPI-html/SPI-description.html (accessed on 10 September 2021).
- Mckee, T.B.; Doesken, N.J.; Kleist, J. The relationship of drought frequency and duration to time scales. In Proceedings of the Conference on Applied Climatology, Boston, MA, USA, 17–22 January 1993; pp. 179–184. [Google Scholar]
- IRI/LDEO Climate Data Library. NOAA NCEP CPC PRECL v1p0 deg0p5 Rain: Monthly Mean of Daily Rainfall Data. 2021. Available online: http://iridl.ldeo.columbia.edu/SOURCES/.NOAA/.NCEP/.CPC/.PRECL/.v1p0/.deg0p5/.rain/ (accessed on 10 September 2021).
- Azevedo, G.B. Composição, Estrutura e Diversidade em Floresta Ombrófila na Reserva Biológica do Jaru-RO. Master’s Thesis, Universidade Federal de Mato Grosso, Cuiabá, Brazil, 2014. [Google Scholar]
- Dan, L.; Ferreira, B.G.A.; Siqueira, J.D.P.; Oliveira, M.M.; Ferreira, A.M. Floristic and phytosociology in dense “terra firme” rainforest in the Belo Monte Hydroelectric Plant influence area, Pará, Brazil. Braz. J. Biol. 2015, 75, 257–276. [Google Scholar] [CrossRef] [Green Version]
- Irvine, J.; Grace, J. Continuous measurement of water tensions in the xylem of trees based on the elastic properties of wood. Planta 1997, 202, 455–461. [Google Scholar] [CrossRef]
- Zweifel, R.; Item, H.; Häsler, R. Stem radius changes and their relation to stored water in stems of young norway spruce trees. Trees 2000, 15, 50–57. [Google Scholar] [CrossRef] [Green Version]
- Zweifel, R.; Zimmermann, L.; Newbery, D.M. Modelling tree water deficit from microclimate: An approach to quantifying drought stress. Tree Physiol. 2005, 25, 147–156. [Google Scholar] [CrossRef] [Green Version]
- Lyapustin, A.I.; Wang, Y.; Laszlo, I.; Hilker, T.; Hall, F.G.; Sellers, P.J.; Tucker, C.J.; Korkin, S.V. Multi-angle implementation of atmospheric correction for MODIS (MAIAC): 3. atmospheric correction. Remote Sens. Environ. 2012, 127, 385–393. [Google Scholar] [CrossRef]
- Huete, A.R.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Nijland, W.; Jong, R.; Jong, S.M.; Wulder, M.A.; Bater, C.W.; Coops, N.C. Monitoring plant condition and phenology using infrared sensitive consumer grade digital cameras. Agric. For. Meteorol. 2014, 184, 98–106. [Google Scholar] [CrossRef] [Green Version]
- Woebbecke, D.M.; Meyer, G.E.; Bargen, K.V.; Mortensen, D.A. Color indices for weed identification under various soil, residue, and lighting conditions. Trans. ASAE 1995, 38, 259–269. [Google Scholar] [CrossRef]
- Muler, R.A.S.; Moura, V.; Borma, L.S. Distribuição espaço-temporal da precipitação e SPI no estado de Rondônia (RO) por meio de técnicas de geoestatística. Rev. Geográfica Venez. 2018, 59, 246–260. [Google Scholar]
- Khanna, J.; Medvigy, D.; Fueglistaler, S.; Walko, R. Regional dry-season climate changes due to three decades of Amazonian deforestation. Nat. Clim. Chang. 2017, 7, 200–204. [Google Scholar] [CrossRef]
- Garzon, M.B.; Munoz, N.G.; Wigneron, J.P.; Moisy, C.; Fernandez-Manjarres, J.; Delzon, S. The legacy of water deficit on populations having experienced negative hydraulic safety margin. Glob. Ecol. Biogeogr. 2018, 27, 356. [Google Scholar] [CrossRef]
- Choat, B. Predicting thresholds of drought-induced mortality in woody plant species. Tree Physiol. 2013, 33, 669–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zweifel, R. Radial stem variations: A source of tree physiological information not fully exploited yet. Plant Cell Environ. 2016, 39, 231–232. [Google Scholar] [CrossRef]
- Wagner, F.H.; Hérault, B.; Bonal, D.; Stahl, C.; Anderson, L.O.; Baker, T.R.; Becker, G.S.; Beeckman, H.; Souza, D.B.; Botosso, P.C.; et al. Climate seasonality limits leaf carbon assimilation and wood productivity in tropical forests. Biogeosciences 2016, 13, 2537–2562. [Google Scholar] [CrossRef] [Green Version]
- Krepkowski, J.; Bräuning, A.; Gebrekirstos, A.; Strobl, S. Cambial growth dynamics and climatic control of different tree life forms in tropical mountain forest in Ethiopia. Trees 2011, 25, 59–70. [Google Scholar] [CrossRef]
- Wagner, F.H.; Rossi, V.; Stahl, C.; Bonal, D.; Hérault, B. Asynchronism in leaf and wood production in tropical forests: A study combining satellite and ground-based measurements. Biogeosciences 2013, 10, 7307–7321. [Google Scholar] [CrossRef] [Green Version]
- Lopes, A.P.; Nelson, B.W.; Wu, J.; Graça, P.M.L.A.; Tavares, J.V.; Prohaska, N.; Martins, G.A.; Saleska, S.R. Leaf flush drives dry season green-up of the Central Amazon. Remote Sens. Environ. 2016, 182, 90–98. [Google Scholar] [CrossRef]
- Wu, J.; Albert, L.P.; Lopes, A.P.; restrepo-Coupa, N.; Hayek, M.; Wiedemann, K.T.; Guan, K.; Stark, S.C.; Christoffersen, B.; Prohaska, N.; et al. Leaf development and demography explain photosynthetic seasonality in Amazon evergreen forests. For. Ecol. 2016, 351, 972–976. [Google Scholar] [CrossRef] [Green Version]
- Sevanto, S.; Mcdowell, N.G.; Dickman, L.T.; Pangle, R.; Pockman, W.T. How do trees die? A test of the hydraulic failure and carbon starvation hypotheses. Plant Cell Environ. 2013, 37, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, T.; Porporato, A. Strategies of a Bornean tropical rainforest water use as a function of rainfall regime: Isohydric or anisohydric? Plant Cell Environ. 2012, 35, 61–71. [Google Scholar] [CrossRef]
- Hilel, D. Soil-water and soil-energy balances in the field. In Soil in the Environment; Academic Press: Cambridge, MA, USA, 2008; pp. 121–133. [Google Scholar]
- Larcher, W. Ecofisiologia Vegetal; Rima Artes e Textos: São Carlos, Brazil, 2000. [Google Scholar]
- Andrade, N.L.R.; Aguiar, R.G.; Sanches, L.; Alves, E.C.R.F.; Nogueira, J.S. Partição do saldo de radiação em áreas de floresta amazônica e floresta de transição Amazônia-Cerrado. Rev. Bras. Meteorol. 2009, 24, 346–355. [Google Scholar] [CrossRef]
- Huete, A.R.; Didan, K.; Shimabukuro, Y.E.; Ratana, P.; Saleska, S.R.; Hutyra, L.R.; Yang, W.; Nermani, R.R.; Myneni, R. Amazon rainforests green-up with sunlight in dry season. Geophys. Res. Lett. 2006, 33, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Da Rocha, H.R.; Goulden, M.L.; Miller, S.D.; Menton, M.C.; Pinto, L.D.V.; Freitas, H.C.; Figueira, A.M.S. Seasonality of water and heat fluxes over a tropical Forest in eastern Amazônia. Ecol. Appl. 2004, 14, 22–32. [Google Scholar] [CrossRef]
- Fan, Y.; Miguez-Macho, G. Potential groundwater contribution to Amazon evapotranspiration. Hydrol. Earth Syst. Sci. 2010, 14, 2039–2056. [Google Scholar] [CrossRef] [Green Version]
- Nepstad, D.C.; Moutinho, P. The recovery of biomass, nutrient stocks, and deep soil function in secondary forests. In The Biogeochemistry of the Amazon Basin and Its Role in a Changing World; Oxford University Press: Oxford, UK, 2001; pp. 139–155. [Google Scholar]
- Oliveira, R.S.; Dawson, T.E.; Burgess, S.S.O.; Nepstad, D.C. Hydraulic redistribution in three Amazonian trees. Oecologia 2005, 145, 354–363. [Google Scholar] [CrossRef]
- Freire, G.A.P.; Ventura, D.J.; Fotopoulos, I.G.; Rosa, D.M.; Aguiar, R.A.; Araujo, A.C. Dinâmica sazonal de serapilheira em uma área de floresta de terra firme, Amazônia Ocidental. Nativa 2020, 8, 323–328. [Google Scholar] [CrossRef]
- Almeida, E.J.; Luizão, F.; Rodrigues, D.J. Produção de serrapilheira em florestas intactas e exploradas seletivamente no sul da Amazônia em função da área basal da vegetação e da densidade de plantas. Acta Amaz. 2015, 45, 157–166. [Google Scholar] [CrossRef] [Green Version]
- Rowland, L.; Costa, A.C.L.; Galbaith, D.R.; Oliveira, R.S.; Binks, O.J.; Oliveira, A.A.R.; Pullen, A.M.; Doughty, C.E. Metcalfe, D.B., Vasconcelos, S.S.; et al. Death from drought in tropical forests is triggered by hydraulics not carbon starvation. Nature 2015, 528, 119–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, K.; Pan, M.; Li, H.; Wolf, A.; Wu, J.; Medvigy, D.; Caylor, K.K.; Sheffield, J.; Wood, E.F.; Malhi, Y.; et al. Photosynthetic seasonality of global tropical forests constrained by hydroclimate. Nat. Geosci. 2015, 8, 284–289. [Google Scholar] [CrossRef]
- Bertani, G.; Wagner, F.H.; Anderson, L.O.; Aragao, L.E.O.C. Chlorophyll fluorescence data reveals climate-related photosynthesis seasonality in Amazonian forests. Remote Sens. 2017, 9, 1275. [Google Scholar] [CrossRef] [Green Version]
- Moura, Y.M.; Hilker, T.; Lyapustin, A.I.; Galvao, L.S.; Santos, J.R.; Anderson, L.O.; Sousa, C.H.R.; Arai, E. Seasonality and drought effects of Amazonian forests observed from multi-angle satellite data. Remote Sens. Environ. 2015, 171, 278–290. [Google Scholar] [CrossRef]
- Galvão, L.S.; Breunig, F.M.; Santos, J.R.; Moura, Y.M. View-illumination effects on hyperspectral vegetation indices in the Amazonian tropical forest. Int. J. Appl. Earth Obs. Geoinf. 2013, 21, 291–300. [Google Scholar] [CrossRef]
- Moura, Y.; Galvao, L.S.; Hilker, T.; Wu, J.; Saleska, S.; Amaral, C.H.; Nelson, B.W.; Lopes, A.P.; Wiedeman, K.K.; Prohaska, N.; et al. Spectral analysis of amazon canopy phenology during the dry season using a tower hyperspectral camera and MODIS observations. ISPRS J. Photogramm. Remote Sens. 2017, 131, 52–64. [Google Scholar] [CrossRef]
- Jensen, J.R. Sensoriamento Remoto Do Ambiente: Uma Perspectiva em Recursos Terrestres; Parêntese: São José dos Campos, Brazil, 2009; p. 598. [Google Scholar]
- Ponzoni, F.J.; Shimabukuro, Y.E.; Kuplich, T.M. Sensoriamento Remoto da Vegetação, 2nd ed.; Oficina de Textos: São Paulo, Brazil, 2012. [Google Scholar]
- Anderson, L.O.; Aragao, L.E.O.C.; Shimabikuro, Y.E.; Almeida, S.; Huete, A. Fraction images for monitoring intra-annual phenology of different vegetation physiognomies in Amazonian. Int. J. Remote Sens. 2011, 32, 387–408. [Google Scholar] [CrossRef]
- Maeda, E.E.; Heiskanen, J.; Aragao, L.E.O.C.; Rinne, J. Can MODIS EVI monitor ecosystem productivity in the Amazon rainforest? Geophys. Res. Lett. 2014, 41, 1–8. [Google Scholar] [CrossRef]
- Bi, J.; Knyazikhin, Y.; Choi, S.; Park, T.; Barichivich, J.; Ciais, P.; Fu, R.; Ganguly, S.; Hall, F.; Hilker, T.; et al. Sunlight mediated seasonality in canopy structure and photosynthetic activity of Amazonian rainforests. Environ. Res. Lett. 2015, 10, 64014. [Google Scholar] [CrossRef] [Green Version]
- Bi, J.; Myneni, R.; Lyapustin, A.; Wang, Y.; Park, T.; Chi, C.; Yan, K.; Knyazikhin, Y. Amazon Forests’ Response to Droughts: A Perspective from the MAIAC Product. Remote Sens. 2016, 8, 356. [Google Scholar] [CrossRef] [Green Version]
- Sousa, C.H.R.; Hilker, T.; Waring, R.; Moura, Y.M.; Lyapustin, A. Progress in Remote Sensing of Photosynthetic Activity over the Amazon Basin. Remote Sens. 2017, 9, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]









| ID | Species | Canopy Positions | Family | Leaf Phenology | Rd | G | IVI |
|---|---|---|---|---|---|---|---|
| 36 | Astronium lecointei Ducke | DC | Anacardiaceae | EG | 0.79 | 1.28 | 1.11 1 |
| 254 | Astronium lecointei Ducke | DC | EG | 1.35 | |||
| 152 | Protium tenuifolium (Engl.) Engl. | DC | Burseraceae | EG | 0.23 1 | 1.30 | 0.22 1 |
| 151 | Protium nitidifolium (Cuatrec.) Daly | DC | EG | 0.20 | 0.75 | 0.52 1 | |
| 168 | Tetragastris altíssima (Aubl.) Swarts | U | EG | 1.04 1 | 1.25 | 2.69 1 | |
| 288 | Licania hypoleuca Benth. | U | Chrysobalanaceae | EG | 0.40 | 0.37 | 0.32 |
| 498 | Licania sprucei (Hook.f.) Fritsch | DC | EG | 0.40 | 1.05 | 0.33 | |
| 284 | Licania sprucei (Hook.f.) Fritsch | U | 0.52 | ||||
| 382 | Anamalocalyx uleanus (Pax & K.Hoffm.) Ducke | U | Euphorbiaceae | EG | 12.9 | 0.43 | 4.35 1 |
| 380 | Sagotia brachysepala (Mull. Arg.) R. Secco | U | EG | 0.01 2 | 0.52 | 0.01 2 | |
| 379 | Sagotia brachysepala (Mull. Arg.) R. Secco | U | |||||
| 147 | Copaifera multijuga Hayne | DC | Fabaceae | EG | 2.96 | 1.86 | 3.78 1 |
| 31 | Copaifera multijuga Hayne | DC | EG | 0.85 | |||
| 24 | Dialium guianense (Aublet.) Sandwith | DC | EG | 1.19 | 1.66 | 5.03 1 | |
| 328 | Dipteryx magnifica Ducke | DC | SD | 0.20 | 1.62 | 0.41 | |
| 183 | Dipteryx odorata (Aublet) Willd. | DC | SD | 2.15 1 | 0.97 | 6.45 1 | |
| 20 | Macrolobium suaveolens Benth | DC | SD | 4.35 | 1.76 | 0.10 1 | |
| 29 | Tachigalichrysophylla (Poepp.) Zarucchi & Herend. | DC | EG | 3.36 | 0.67 | 2.76 | |
| 177 | Tachigali chrysophylla (Poepp.) Zarucchi & Herend. | DC | EG | 0.64 | |||
| 180 | Tachigali chrysophylla (Poepp.) Zarucchi & Herend. | DC | EG | 1.58 | |||
| 102 | Swartzia ingifolia Ducke | DC | SD | 0.20 | 1.53 | 0.16 | |
| 144 | Sterculia duckei E.L. Taylor ex J.A.C. Silva&M.F.Silva | DC | Malvaceae | SD | 0.20 | 1.24 | 0.29 |
| 182 | Lueheopsis rosea (Ducke) Burret | U | EG | 0.14 | 0.65 | 0.92 | |
| 286 | Cariniana decandra Ducke | U | Lecythidaceae | EG | 0.59 | 1.13 | 5.22 1 |
| 161 | Eschweilera coriacea (DC.) S.A.Mori | DC | 0.20 | 0.94 | 0.24 | ||
| 381 | Virola michelii Heckel | U | Myristicaceae | EG | 1.38 | 0.41 | 1.08 |
| 27 | Minquartia guianensis Aubl. | U | Olacaceae | EG | 1.58 | 1.17 | 2.06 |
| 167 | Minquartia guianensis Aubl. | U | 1.10 | ||||
| 03 | Minquartia guianensis Aubl. | U | 1.12 | ||||
| 137 | Pouteria egleri Eyma | U | Sapotaceae | EG | 1.00 | 0.65 | 1.17 |
| 32 | Pouteria durlandii (Standl.) Baehni | U | EG | 0.58 1 | 0.88 | 0.45 | |
| Total (n = 31) | 36.05 | 31.25 | 39.67 | ||||
| Diameter Class | πTLP (MPa) | P50 (MPa) | ||||
|---|---|---|---|---|---|---|
| Trees | Average | σ | Trees | Average | σ | |
| DBH 10–20 cm | sp = 5 | −2.29 | 0.63 | sp = 3 | −1.507 | 1.028 |
| DBH 20–30 cm | sp = 5 | −1.79 | 0.69 | sp = 2 | −2.780 | 0.537 |
| DBH 30–40 cm | sp = 6 | −1.93 | 0.55 | sp = 4 | −4.486 | 0.503 |
| DBH > 40 cm | sp = 6 | −1.67 | 0.53 | sp = 1 | −4.71 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Souza, R.D.A.D.; Moura, V.; Paloschi, R.A.; Aguiar, R.G.; Webler, A.D.; Borma, L.D.S. Assessing Drought Response in the Southwestern Amazon Forest by Remote Sensing and In Situ Measurements. Remote Sens. 2022, 14, 1733. https://0-doi-org.brum.beds.ac.uk/10.3390/rs14071733
Souza RDAD, Moura V, Paloschi RA, Aguiar RG, Webler AD, Borma LDS. Assessing Drought Response in the Southwestern Amazon Forest by Remote Sensing and In Situ Measurements. Remote Sensing. 2022; 14(7):1733. https://0-doi-org.brum.beds.ac.uk/10.3390/rs14071733
Chicago/Turabian StyleSouza, Ranieli Dos Anjos De, Valdir Moura, Rennan Andres Paloschi, Renata Gonçalves Aguiar, Alberto Dresch Webler, and Laura De Simone Borma. 2022. "Assessing Drought Response in the Southwestern Amazon Forest by Remote Sensing and In Situ Measurements" Remote Sensing 14, no. 7: 1733. https://0-doi-org.brum.beds.ac.uk/10.3390/rs14071733