
Identifying Conservation Introduction Sites for Endangered Birds through the Integration of Lidar-Based Habitat Suitability Models and Population Viability Analyses

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Lidar and Bird Data Acquisition
2.3. Lidar Data Processing
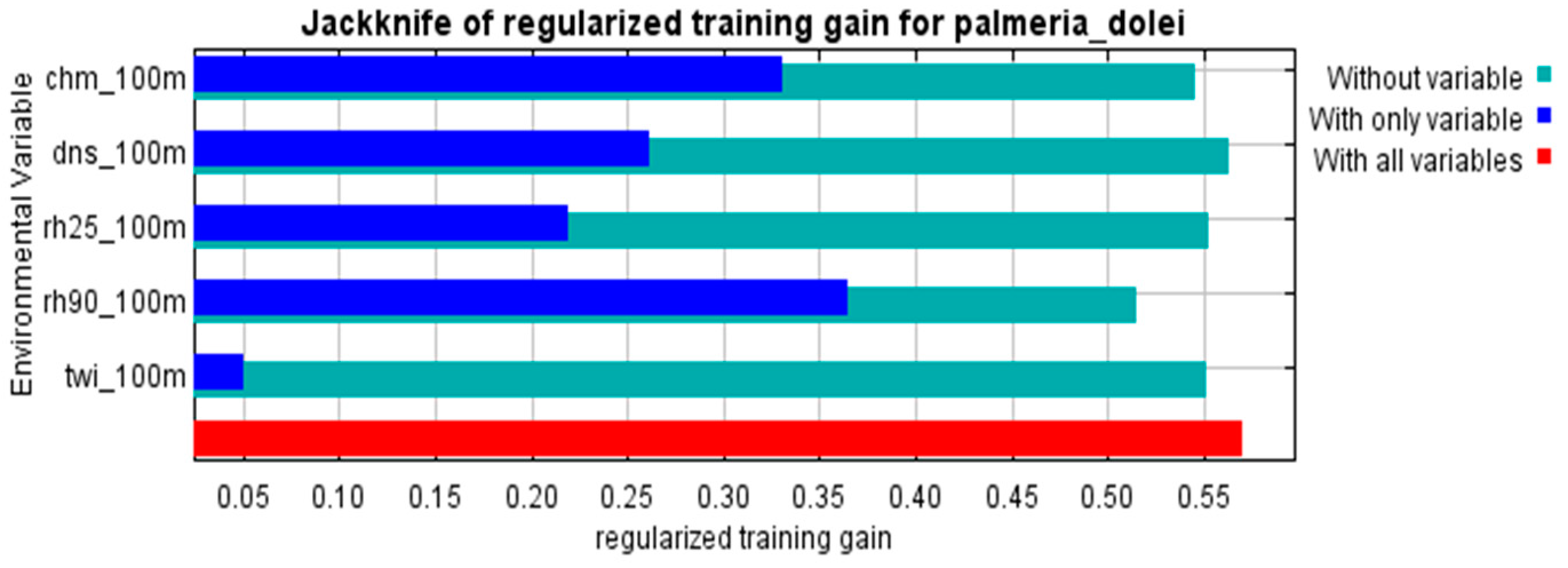
2.4. Habitat Suitability Models
2.5. Candidate Translocation Introduction Site Assessment
2.6. Population Viability Analysis
2.7. Web Application
3. Results
3.1. ‘Ākohekohe on the Island of Hawai’i
3.2. Candidate Site Evaluation
3.3. Population Viability Analysis
3.4. Web Application
4. Discussion
4.1. Lidar Sensor Correction
4.2. Habitat Preferences
4.3. Candidate Conservation Introduction Site Comparison
4.4. Release Site Selection
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Sensor Name | Beam Divergence (mrad) | Altitude (m AGL) | PRF (kHz) | Wavelength (nm) |
|---|---|---|---|---|---|
| GAO | Optech HA-500 | 0.25 | 2000 | 100–500 | 1064 |
| NSF | Optech Gemini | 0.25–0.8 | 1000 | 100 | 1064 |
| USGS | Leica SPL | 0.08 | 3200–4420 | 60 | 532 |
| FWS | Riegel LMS-Q680i | ≤0.5 | 500 | 80–400 | Near-infrared (700–1400) |
| Metrics | Hakalau (USGS~FWS) | Pu’u Maka’ala (USGS~NSF) | ||||
|---|---|---|---|---|---|---|
| Intercept | Slope | R2 | Intercept | Slope | R2 | |
| Canopy Height | −0.22 | 0.82 | 0.94 | 0.44 | 0.87 | 0.94 |
| Relative Height25 | 0.21 | 1.16 | 0.54 | −0.02 | 0.33 | 0.81 |
| Relative Height50 | 1.15 | 0.65 | 0.82 | 0.16 | 0.81 | 0.92 |
| Relative Height75 | 0.23 | 0.74 | 0.92 | 0.55 | 1.00 | 0.93 |
| Relative Height90 | −1.21 | 0.80 | 0.91 | 0.69 | 1.09 | 0.93 |
| Canopy Density | −3.20 | 0.89 | 0.85 | 6.51 | 1.02 | 0.89 |
| CHM | DNS | DTM | RH25 | RH50 | RH75 | RH90 | SLOPE | |
|---|---|---|---|---|---|---|---|---|
| CHM | 1.00 | |||||||
| DNS | 0.89 | 1.00 | ||||||
| DTM | −0.04 | −0.15 | 1.00 | |||||
| RH25 | 0.82 | 0.88 | −0.16 | 1.00 | ||||
| RH50 | 0.98 | 0.88 | −0.02 | 0.82 | 1.00 | |||
| RH75 | 0.98 | 0.84 | −0.01 | 0.75 | 0.97 | 1.00 | ||
| RH90 | 0.97 | 0.83 | −0.02 | 0.73 | 0.94 | 0.99 | 1.00 | |
| SLOPE | 0.40 | 0.31 | 0.23 | 0.19 | 0.41 | 0.46 | 0.47 | 1.00 |
| TWI | −0.14 | −0.09 | −0.27 | 0.05 | −0.17 | −0.21 | −0.21 | −0.67 |
| Variable | Percent Contribution | Permutation Importance |
|---|---|---|
| Canopy Height | 40.8 | 18.7 |
| RH90 | 30.9 | 37.8 |
| Canopy Density | 20.9 | 18.9 |
| TWI | 4.5 | 3.9 |
| RH25 | 2.9 | 20.7 |




Appendix B. Population Viability Analysis for ‘Ākohekohe (Palmeria dolei) Conservation Introductions on the Island of Hawai’i
Appendix B.1. Single Run and Stochastic Simulations
Appendix B.2. Sensitivity Analysis
Appendix B.3. R Code and Results
Appendix B.3.1. Defining Constants

Appendix B.3.2. Creating Model for Individual Release Population Projection


Appendix B.3.3. Running a Deterministic Individual Release Population Projection


Appendix B.3.4. Stochastic Release Population Projection

Appendix B.3.5. Explore the Success Rate by Starting Population







Appendix B.4. Sensitivity Analyses of PVA
Appendix B.4.1. CI Success Rate by Long-Term Mean Adult Survival Effect


Appendix B.4.2. Success Rate by Long-Term Mean Juvenile Survival Effect


Appendix B.4.3. Sensitivity Analysis of CI Success and First-Year Survival Effects on Adults


Appendix B.5. Discussion
References
- Dirzo, R.; Young, H.S.; Galetti, M.; Ceballos, G.; Isaac, N.J.B.; Collen, B. Defaunation in the Anthropocene. Science 2014, 345, 401–406. [Google Scholar] [CrossRef]
- Bubac, C.M.; Johnson, A.C.; Fox, J.A.; Cullingham, C.I. Conservation Translocations and Post-Release Monitoring: Identifying Trends in Failures, Biases, and Challenges from around the World. Biol. Conserv. 2019, 238, 108239. [Google Scholar] [CrossRef]
- Seddon, P.J. From Reintroduction to Assisted Colonization: Moving along the Conservation Translocation Spectrum. Restor. Ecol. 2010, 18, 796–802. [Google Scholar] [CrossRef]
- Berio Fortini, L.; Kaiser, L.R.; Lapointe, D.A. Fostering Real-Time Climate Adaptation: Analyzing Past, Current, and Forecast Temperature to Understand the Dynamic Risk to Hawaiian Honeycreepers from Avian Malaria. Glob. Ecol. Conserv. 2020, 23, e01069. [Google Scholar] [CrossRef]
- Paxton, E.H.; Laut, M.; Enomoto, S.; Bogardus, M. Hawaiian Forest Bird Conservation Strategies for Minimizing the Risk of Extinction: Biological and Biocultural Considerations; Hawaii Cooperative Studies Unit Technical Report 103; USGS: Reston, VA, USA, 2022.
- Lapointe, D.A.; Atkinson, C.T.; Samuel, M.D. Ecology and Conservation Biology of Avian Malaria. Ann. N. Y. Acad. Sci. 2012, 1249, 211–226. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, C.T.; Utzurrum, R.B.; Lapointe, D.A.; Camp, R.J.; Crampton, L.H.; Foster, J.T.; Giambelluca, T.W. Changing Climate and the Altitudinal Range of Avian Malaria in the Hawaiian Islands—An Ongoing Conservation Crisis on the Island of Kaua’i. Glob. Chang. Biol. 2014, 20, 2426–2436. [Google Scholar] [CrossRef] [PubMed]
- Fortini, L.B.; Vorsino, A.E.; Amidon, F.A.; Paxton, E.H.; Jacobi, J.D. Large-Scale Range Collapse of Hawaiian Forest Birds under Climate Change and the Need 21st Century Conservation Options. PLoS ONE 2015, 10, e0140389. [Google Scholar] [CrossRef]
- Liao, W.; Atkinson, C.T.; LaPointe, D.A.; Samuel, M.D. Mitigating Future Avian Malaria Threats to Hawaiian Forest Birds from Climate Change. PLoS ONE 2017, 12, e0168880. [Google Scholar] [CrossRef]
- Benning, T.L.; LaPointe, D.; Atkinson, C.T.; Vitousek, P.M. Interactions of Climate Change with Biological Invasions and Land Use in the Hawaiian Islands: Modeling the Fate of Endemic Birds Using a Geographic Information System. Proc. Natl. Acad. Sci. USA 2002, 99, 14246–14249. [Google Scholar] [CrossRef]
- Perkins, R.C.L. Vertebrata; Fauna Hawaiiensis; University Press: Cambridge, UK, 1903. [Google Scholar]
- Judge, S.W.; Warren, C.C.; Camp, R.J.; Berthold, L.K.; Mounce, H.L.; Hart, P.J.; Monello, R.J. Population Estimates and Trends of Three Maui Island-Endemic Hawaiian Honeycreepers. J. Field Ornithol. 2021, 92, 115–126. [Google Scholar] [CrossRef]
- Berlin, K.E.; Simon, J.C.; Pratt, T.K.; Kowalsy, J.R.; Hatfield, J.S. Akohekohe Response to Flower Availability: Seasonal Abundance, Foraging, Breeding and Molt. Stud. Avian Biol. 2001, 22, 202–212. [Google Scholar]
- Wang, A.X.; Paxton, E.H.; Mounce, H.L.; Hart, P.J. Divergent Movement Patterns of Adult and Juvenile ‘Akohekohe, an Endangered Hawaiian Honeycreeper. J. Field Ornithol. 2020, 91, 346–353. [Google Scholar] [CrossRef]
- Paxton, E.H.; Camp, R.J.; Gorresen, P.M.; Crampton, L.H.; Leonard, D.L.; VanderWerf, E.A. Collapsing Avian Community on a Hawaiian Island. Sci. Adv. 2016, 2, e1600029. [Google Scholar] [CrossRef]
- White, T.H.; de Melo Barros, Y.; Develey, P.F.; Llerandi-Román, I.C.; Monsegur-Rivera, O.A.; Trujillo-Pinto, A.M. Improving Reintroduction Planning and Implementation through Quantitative SWOT Analysis. J. Nat. Conserv. 2015, 28, 149–159. [Google Scholar] [CrossRef]
- Rounds, R. Hawaiian Forest Bird SWOT Workshop Report, 2022; Unpublished report to the U.S. Fish and Wildlife Service. Honolulu, Hawaii.
- Stamps, J.A.; Swaisgood, R.R. Someplace like Home: Experience, Habitat Selection and Conservation Biology. Appl. Anim. Behav. Sci. 2007, 102, 392–409. [Google Scholar] [CrossRef]
- Bilby, J.; Moseby, K. Review of Hyperdispersal in Wildlife Translocations. Conserv. Biol. 2023, 38, e14083. [Google Scholar] [CrossRef] [PubMed]
- Becker, D.; Massey, G.; Groombridge, J.; Hammond, R. Moving I’iwi (Vestiaria Coccinea) as a Surrogate for Future Translocations of Endangered ’Akohekohe (Palmeria Dolei). Pacific Coop. Stud. Unit Unv. Hawai’i Manoa 2010, 172, 1–13. [Google Scholar]
- Dayananda, S.K.; Mammides, C.; Liang, D.; Kotagama, S.W.; Goodale, E. A Review of Avian Experimental Translocations That Measure Movement through Human-Modified Landscapes. Glob. Ecol. Conserv. 2021, 31, e01876. [Google Scholar] [CrossRef]
- Gallerani, E.M.; Burgett, J.; Vaughn, N.; Berio Fortini, L.; Fricker, G.A.; Mounce, H.; Gillespie, T.W.; Crampton, L.; Knapp, D.; Hite, J.M.; et al. High Resolution Lidar Data Shed Light on Inter-Island Translocation of Endangered Bird Species in the Hawaiian Islands. Ecol. Appl. 2023, 33, e2889. [Google Scholar] [CrossRef] [PubMed]
- Fricker, G.A.; Crampton, L.H.; Gallerani, E.M.; Hite, J.M.; Inman, R.; Gillespie, T.W. Application of Lidar for Critical Endangered Bird Species Conservation on the Island of Kauai, Hawaii. Ecosphere 2021, 12, e03554. [Google Scholar] [CrossRef]
- Scott, J.M.; Mountainspring, S.; Ramsey, F.L.; Kepler, C.B. Forest Bird Communities of the Hawaiian Islands: Their Dynamics, Ecology, and Conservation. J. Wildl. Manag. 1986, 53, 860. [Google Scholar] [CrossRef]
- Warren, C.C.; Motyka, P.J.; Mounce, H.L. Home Range Sizes of Two Hawaiian Honeycreepers: Implications for Proposed Translocation Efforts. J. Field Ornithol. 2015, 86, 305–316. [Google Scholar] [CrossRef]
- Price, J.P.; Jacobi, J.D.; Gon, S.M., III; Matsuwaki, D.; Mehrhoff, L.; Wagner, W.; Lucas, M.; Rowe, B. Mapping Plant Species Ranges in the Hawaiian Islands: Developing a Methodology and Associated GIS Layers; U.S. Geological Survey Open-File Report: Reston, VA, USA, 2012.
- Giambelluca, T.W.; Chen, Q.; Frazier, A.G.; Price, J.P.; Chen, Y.L.; Chu, P.S.; Eischeid, J.K.; Delparte, D.M. Online Rainfall Atlas of Hawai’i. Bull. Am. Meteorol. Soc. 2013, 94, 313–316. [Google Scholar] [CrossRef]
- Camp, R.J. Landbirds Vital Signs Monitoring Protocol- Pacific Island Network; U.S. Department of the Interior, National Park Service, Natural Resource Center: Reston, VA, USA, 2011.
- Gopalakrishnan, R.; Thomas, V.A.; Coulston, J.W.; Wynne, R.H. Prediction of Canopy Heights over a Large Region Using Heterogeneous Lidar Datasets: Efficacy and Challenges. Remote Sens. 2015, 7, 11036–11060. [Google Scholar] [CrossRef]
- U.S. Geological Survey USGS Lidar Point Cloud HI_Hawaii_Island_Lidar_NOAA_2017. Available online: https://www.usgs.gov/the-national-map-data-delivery (accessed on 30 May 2023).
- U.S. Fish and Wildlife Service Lidar Data Collected by Survey Hawai’i for the USFWS Hakalau Forest National Wildlife Refuge 2021. Non-Publicly Shared Data Used for Internal Planning. (accessed on 30 May 2023).
- NEON (National Ecological Observatory Network) Discrete Return LiDAR Point Cloud (DPI.30003.001). Available online: https://data.neonscience.org/data-products/DP1.30003.001/RELEASE-2023 (accessed on 30 May 2023).
- Roussel, J.-R.; Auty, D.; Coops, N.C.; Tompalski, P.; Goodbody, T.R.H.; Meador, A.S.; Bourdon, J.-F.; de Boissieu, F.; Achim, A. LidR: An R Package for Analysis of Airborne Laser Scanning (ALS) Data. Remote Sens. Environ. 2020, 251, 112061. [Google Scholar] [CrossRef]
- Evans, J.S.; Hudak, A.T. A Multiscale Curvature Algorithm for Classifying Discrete Return LiDAR in Forested Environments. IEEE Trans. Geosci. Remote Sens. 2007, 45, 1029–1038. [Google Scholar] [CrossRef]
- Hijmans, R.J. Raster: Geographic Data Analysis and Modeling. Available online: https://rspatial.org/raster (accessed on 1 June 2023).
- Fricker, G.A.; Wolf, J.A.; Saatchi, S.S.; Gillespie, T.W. Predicting Spatial Variations of Tree Species Richness in Tropical Forests from High-Resolution Remote Sensing. Ecol. Appl. 2015, 25, 1776–1789. [Google Scholar] [CrossRef]
- Phillips, S.J. A Brief Tutorial on Maxent. Available online: http://biodiversityinformatics.amnh.org/open_source/maxent/ (accessed on 1 June 2023).
- Duque-Lazo, J.; van Gils, H.; Groen, T.A.; Navarro-Cerrillo, R.M. Transferability of Species Distribution Models: The Case of Phytophthora Cinnamomi in Southwest Spain and Southwest Australia. Ecol. Modell. 2016, 320, 62–70. [Google Scholar] [CrossRef]
- Bakx, T.R.M.; Koma, Z.; Seijmonsbergen, A.C.; Kissling, W.D. Use and Categorization of Light Detection and Ranging Vegetation Metrics in Avian Diversity and Species Distribution Research. Divers. Distrib. 2019, 25, 1045–1059. [Google Scholar] [CrossRef]
- Draper, D.; Marques, I.; Iriondo, J.M. Species Distribution Models with Field Validation, a Key Approach for Successful Selection of Receptor Sites in Conservation Translocations. Glob. Ecol. Conserv. 2019, 19, e00653. [Google Scholar] [CrossRef]
- Radosavljevic, A.; Anderson, R.P. Making Better Maxent Models of Species Distributions: Complexity, Overfitting and Evaluation. J. Biogeogr. 2014, 41, 629–643. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. Global Forest Resources Assessment. FRA Working Paper Series 188. 2020. Available online: https://www.fao.org/3/I8661EN/i8661en.pdf (accessed on 30 May 2023).
- Haralick, R.M.; Dinstein, I.; Shanmugam, K. Textural Features for Image Classification. IEEE Trans. Syst. Man Cybern. 1973, SMC-3, 610–621. [Google Scholar] [CrossRef]
- Chang, W.; Cheng, J.; Allaire, J.J.; Sievert, C.; Schloerke, B.; Xie, Y.; Allen, J.; McPherson, J.; Dipert, A.; Borges, B. Shiny: Web Application Framework for R. Available online: https://shiny.posit.co/ (accessed on 30 July 2023).
- Ussyshkin, V.; Theriault, L. Airborne Lidar: Advances in Discrete Return Technology for 3D Vegetation Mapping. Remote Sens. 2011, 3, 416–434. [Google Scholar] [CrossRef]
- Nilsson, M. Estimation of Tree Heights and Stand Volume Using an Airborne Lidar System. Remote Sens. Environ. 1996, 56, 1–7. [Google Scholar] [CrossRef]
- Hopkinson, C. The Influence of Flying Altitude, Beam Divergence, and Pulse Repetition Frequency on Laser Pulse Return Intensity and Canopy Frequency Distribution. Can. J. Remote Sens. 2007, 33, 312–324. [Google Scholar] [CrossRef]
- Wei, G.; Shalei, S.; Bo, Z.; Shuo, S.; Faquan, L.; Xuewu, C. Multi-Wavelength Canopy LiDAR for Remote Sensing of Vegetation: Design and System Performance. ISPRS J. Photogramm. Remote Sens. 2012, 69, 1–9. [Google Scholar] [CrossRef]
- Yang, Y.; Saatchi, S.S.; Xu, L.; Yu, Y.; Choi, S.; Phillips, N.; Kennedy, R.; Keller, M.; Knyazikhin, Y.; Myneni, R.B. Post-Drought Decline of the Amazon Carbon Sink. Nat. Commun. 2018, 9, 3172. [Google Scholar] [CrossRef]
- Ehlers, D.; Wang, C.; Coulston, J.; Zhang, Y.; Pavelsky, T.; Frankenberg, E.; Woodcock, C.; Song, C. Mapping Forest Aboveground Biomass Using Multisource Remotely Sensed Data. Remote Sens. 2022, 14, 1115. [Google Scholar] [CrossRef]
- Camp, R.J.; LaPointe, D.A.; Hart, P.J.; Sedgwick, D.E.; Canale, L.K. Large-Scale Tree Mortality from Rapid Ohia Death Negatively Influences Avifauna in Lower Puna, Hawaii Island, USA. Condor 2019, 121, duz007. [Google Scholar] [CrossRef]
- Berio Fortini, L.; Gallerani, E. Island of Hawai’i Lidar-Based Habitat Suitability for ‘ākohekohe (Palmeria Dolei) Conservation Introductions; U.S. Geological Survey Data Release: Reston, VA, USA, 2024. [CrossRef]
- Sæther, B.E.; Engen, S.; Lande, R.; Møller, A.P.; Bensch, S.; Hasselquist, D.; Beier, J.; Leisler, B. Time to Extinction in Relation to Mating System and Type of Density Regulation in Populations with Two Sexes. J. Anim. Ecol. 2004, 73, 925–934. [Google Scholar] [CrossRef]
- Caswell, H. Matrix Population Models: Construction, Analysis and Interpretation; Sinauer Associates Inc.: Sunderland, MA, USA, 2002. [Google Scholar]
- Simon, J.C.; Pratt, T.K.; Berlin, K.E.; Kowalsy, J.R. Reproductive Ecology and Demography of the Akohekohe. Condor 2001, 103, 736–745. [Google Scholar] [CrossRef]
- Woodworth, B.L.; Pratt, T.K. Life History and Demography. In Conservation Biology of Hawaiian Forest Birds: Implications for Island Avifauna; Pratt, T.K., Atkinson, C.T., Banko, P.C., Jacobi, J.D., Woodworth, B.L., Eds.; Yale University Press: New Haven, CT, USA, 2009; ISBN 9780300141085. [Google Scholar]
- Brook, B.W.; Grady, J.J.O.; Chapman, A.P.; Burgman, M.A.; Akc, H.R. Predictive Accuracy of Population Viability Analysis in Conservation Biology. Nature 2000, 404, 385–387. [Google Scholar] [CrossRef] [PubMed]
- Fancy, S.G.; Snetsinger, T.J.; Jacob, J.D. Translocation of the Palila, an Endangered Hawaiian Honeycreeper. Pac. Conserv. Biol. 1997, 3, 39–46. [Google Scholar] [CrossRef]




| Variable Name | Description |
|---|---|
| Canopy Density (DNS) | Percentage of lidar points > 1.37 m (4.5 ft) above ground in the selected region |
| Canopy Height (CHM) | Height (m) of the modeled canopy surface above the ground |
| RH25 | 25th relative height above the ground for all lidar points in the selected region |
| RH90 | 90th relative height above the ground for all lidar points in the selected region |
| Topographic Wetness Index (TWI) | Relative wetness of the selected region within the landscape |
| Candidate Sites | Suitable Clusters | Average Area (km2) | Total Area of Clusters (km2) | Not Suitable Clusters |
|---|---|---|---|---|
| Hakalau | 21 | 0.15 | 3.14 | 1 |
| Ka’ū | 13 | 1.48 | 19.22 | 2 |
| Pu’u Maka’ala | 0 | NA | 0 | 11 |
| Pu’u Wa’awa’a | 3 | 0.11 | 0.32 | 0 |
| Kīpuka | 2 | 0.04 | 0.09 | 5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gallerani, E.M.; Berio Fortini, L.; Warren, C.C.; Paxton, E.H. Identifying Conservation Introduction Sites for Endangered Birds through the Integration of Lidar-Based Habitat Suitability Models and Population Viability Analyses. Remote Sens. 2024, 16, 680. https://0-doi-org.brum.beds.ac.uk/10.3390/rs16040680
Gallerani EM, Berio Fortini L, Warren CC, Paxton EH. Identifying Conservation Introduction Sites for Endangered Birds through the Integration of Lidar-Based Habitat Suitability Models and Population Viability Analyses. Remote Sensing. 2024; 16(4):680. https://0-doi-org.brum.beds.ac.uk/10.3390/rs16040680
Chicago/Turabian StyleGallerani, Erica Marie, Lucas Berio Fortini, Christopher C. Warren, and Eben H. Paxton. 2024. "Identifying Conservation Introduction Sites for Endangered Birds through the Integration of Lidar-Based Habitat Suitability Models and Population Viability Analyses" Remote Sensing 16, no. 4: 680. https://0-doi-org.brum.beds.ac.uk/10.3390/rs16040680