Carotenoid Content in Human Colostrum is Associated to Preterm/Full-Term Birth Condition

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Measures
2.2.1. Milk and Blood Sample Collection
2.2.2. Extraction of the Carotenoid Fraction
2.2.3. Identification and Quantification of Carotenoids in the Carotenoid Extracts
2.2.4. Determination of the Lipid Content
2.3. Ethics Approval
2.4. Statistical Analysis
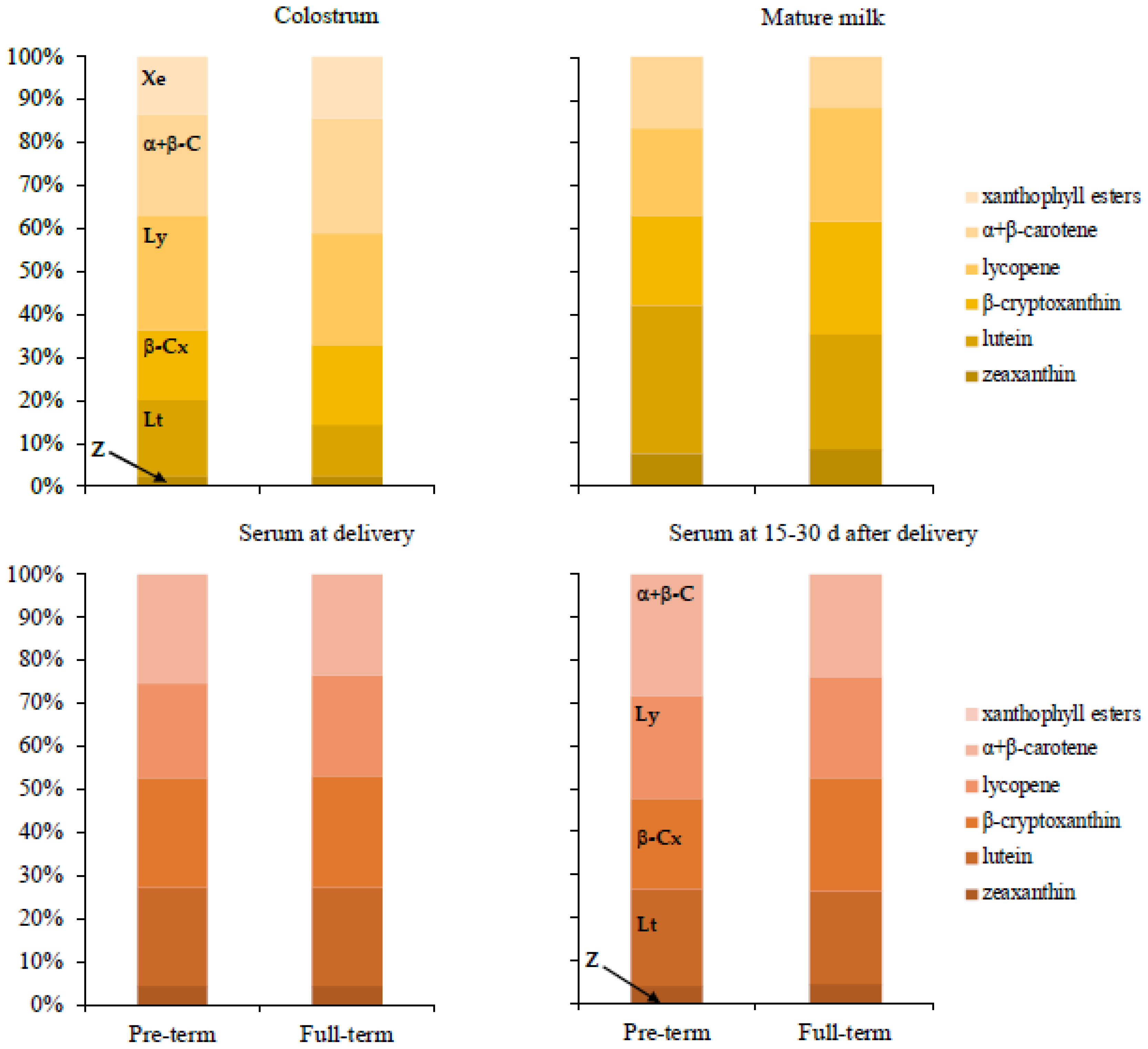
3. Results
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| PT | preterm |
| FT | full-term |
| HPLC | high-performance liquid chromatography |
| APCI | atmospheric pressure chemical ionization |
| UHR | ultra-high resolution |
| TOF | time of flight |
References
- Leon-Cava, N.; Lutter, C.; Ross, J.; Martin, L. Quantifying the Benefits of Breastfeeding: A Summary of the Evidence; Pan American Health Organization: Washington, DC, USA, 2002; pp. 1–168. ISBN 9275123977. [Google Scholar]
- Betran, A.P.; de Onis, M.; Lauer, J.A.; Villar, J. Ecological study of effect of breast feeding on infant mortality in Latin America. BMJ 2001, 323, 303–306. [Google Scholar] [CrossRef] [PubMed]
- WHO Collaborative Study Team on the Role of Breastfeeding on the Prevention of Infant Mortality. Effect of breastfeeding on infant and child mortality due to infectious diseases in less developed countries: A pooled analysis. Lancet 2000, 355, 451–455. [Google Scholar] [CrossRef]
- WHO Global Strategy for Infant and Young Child Feeding; World Health Organization: Geneva, Switzerland, 2003; pp. 1–30. ISBN 9241562218.
- Picciano, M.F. Nutrient composition of human milk. Pediatr. Clin. North Am. 2001, 48, 53–67. [Google Scholar] [CrossRef]
- Ballard, O.; Morrow, A.L. Human milk composition: Nutrients and bioactive factors. Pediatr. Clin. North Am. 2013, 60, 49–74. [Google Scholar] [CrossRef] [PubMed]
- Liaaen-Jensen, S. Basic carotenoid chemistry. In Carotenoids in Health and Disease; Krinsky, N.I., Mayne, S.T., Sies, H., Eds.; Marcel Dekker: New York, NY, USA, 2004; pp. 1–30. ISBN 0203026640. [Google Scholar]
- Stoltzfus, R.J.; Hakima, M.; Miller, K.W.; Rasmussen, K.M.; Dawiesah, S.I.; Habicht, J.P.; Dibley, M.J. High dose vitamin A supplementation of breast-feeding Indonesian mothers: Effects on the vitamin A status of mother and infant. J. Nutr. 1993, 123, 666–675. [Google Scholar] [CrossRef] [PubMed]
- Bendich, A. Non-provitamin A activity of carotenoids: Immunoenhancement. Trends Food Sci. Tech. 1991, 2, 127–130. [Google Scholar] [CrossRef]
- Stahl, W.; Sies, H. Bioactivity and protective effects of natural carotenoids. Biochim. Biophys. Acta 2005, 1740, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Provis, J.; Diaz, C.; Dreher, B. Ontogeny of the primate fovea: A central issue in retinal development. Prog. Neurobiol. 1998, 54, 549–581. [Google Scholar] [CrossRef]
- Landrum, J.; Bone, R. Lutein, zeaxanthin, and the macular pigment. Arch. Biochem. Biophys. 2001, 385, 28–40. [Google Scholar] [CrossRef] [PubMed]
- Pintea, A.; Socaciu, C.; Rugina, D.O.; Pop, R.; Bunea, A. Xanthophylls protect against induced oxidation in cultured human retinal pigment epithelial cells. J. Food Compost. Anal. 2011, 26, 830–836. [Google Scholar] [CrossRef]
- Junghans, A.; Sies, H.; Stahl, W. Macular pigments lutein and zeaxanthin as blue light filters studied in liposomes. Arch. Biochem. Biophys. 2001, 391, 160–164. [Google Scholar] [CrossRef] [PubMed]
- Leung, I.; Sandstrom, M.M.; Zucker, C.; Neuringer, M.; Snodderly, D. Nutritional manipulation of primate retinas, II: Effects of age, n-3 fatty acids, lutein, and zeaxanthin on retinal pigment epithelium. Invest. Ophthalmol. Vis. Sci. 2004, 45, 3244–3256. [Google Scholar] [CrossRef] [PubMed]
- Loughman, J.; Nolan, J.M.; Howard, A.N.; Connolly, E.; Meagher, K.; Beatty, S. The impact of macular pigment augmentation on visual performance using different carotenoid formulations. Invest. Ophthalmol. Vis. Sci. 2012, 53, 7871–7880. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D.; Coen, R.F.; Akuffo, K.O.; Beatty, S.; Dennison, J.; Moran, R.; Stack, J.; Howard, A.N.; Mulcahy, R.; Nolan, J.M. Cognitive function and its relationship with macular pigment optical density and serum concentrations of its constituent carotenoids. J. Alzheimers. Dis. 2015, 48, 261–277. [Google Scholar] [CrossRef] [PubMed]
- Vishwanathan, R.; Iannaccone, A.; Scott, T.M.; Kritchevsky, S.B.; Jennings, B.J.; Carboni, G.; Forma, G.; Satterfield, S.; Harris, T.; Johnson, K.C.; et al. Macular pigment optical density is related to cognitive function in older people. Age Ageing 2014, 43, 271–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vishwanathan, R.; Neuringer, M.; Snodderly, D.M.; Schalch, W.; Johnson, E.J. Macular lutein and zeaxanthin are related to brain lutein and zeaxanthin in primates. Nutr. Neurosci. 2013, 16, 21–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craft, N.E.; Haitema, T.B.; Garnett, K.M.; Fitch, K.A.; Dorey, C.K. Carotenoid, tocopherol, and retinol concentrations in elderly human brain. J. Nutr. Health Aging 2004, 8, 156–162. [Google Scholar] [PubMed]
- Johnson, E.J.; Vishwanathan, R.; Johnson, M.A.; Hausman, D.B.; Davey, A.; Scott, T.M.; Green, R.C.; Miller, S.; Gearing, M.; Woodard, J.; et al. Relationship between serum and brain carotenoids, α-tocopherol, and retinol concentrations and cognitive performance in the oldest old from the Georgia Centenarian Study. J. Aging Res. 2013, 2013, 951786. [Google Scholar] [CrossRef] [PubMed]
- Vishwanathan, R.; Kuchan, M.; Sen, S.; Johnson, E.J. Lutein and preterm infants with decreased concentration of brain carotenoids. J. Pediatr. Gastroenterol. Nutr. 2014, 59, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Hammond, B.R. Jr. Possible role for dietary lutein and zeaxanthin in visual development. Nutr. Rev. 2008, 66, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.J. Role of lutein and zeaxanthin in visual and cognitive function throughout the lifespan. Nutr. Rev. 2014, 72, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Henriksen, B.S.; Chan, G.; Hoffman, R.O.; Sharifzadeh, M.; Ermakov, I.V.; Gellermann, W.; Bernstein, P.S. Interrelationships between maternal carotenoid status and newborn infant macular pigment optical density and carotenoid status. Invest. Ophthalmol. Vis. Sci. 2013, 54, 5568–5578. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, P.S.; Sharifzadeh, M.; Liu, A.; Ermakiv, I.; Nelson, K.; Sheng, X.; Panish, C.; Carlstrom, B.; Hoffman, R.O.; Gellermann, W. Blue-light reflectance imaging of macular pigment in infants and children. Invest. Ophthalmol. Vis. Sci. 2013, 54, 4034–4040. [Google Scholar] [CrossRef] [PubMed]
- Bone, R.A.; Landrum, J.T.; Friedes, L.M.; Gomez, C.M.; Kilburn, M.D.; Menendez, E.; Vidal, I.; Wang, W. Distribution of lutein and zeaxanthin stereoisomers in the human retina. Exp. Eye Res. 1997, 64, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Giuliano, A.R.; Neilson, E.M.; Yap, H.; Baier, M.; Canfield, L.M. Carotenoids of mature human milk: Inter/intraindividual variability. J. Nutr. Biochem. 1994, 5, 551–556. [Google Scholar] [CrossRef]
- Canfield, L.M.; Clandinin, M.T.; Davies, D.P.; Fernandez, M.C.; Jackson, J.; Hawkes, J.; Goldman, W.J.; Pramuk, K.; Reyes, H.; Sablan, B.; et al. Multinational study of major breast milk carotenoids of healthy mothers. Eur. J. Nutr. 2003, 42, 133–141. [Google Scholar] [PubMed]
- Patton, S.; Canfield, L.M.; Huston, G.E.; Ferris, A.M.; Jensen, R.G. Carotenoids of human colostrum. Lipids 1990, 25, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Khachik, F.; Spangler, C.J.; Smith, J.C.; Canfield, L.M.; Pfander, H.; Speck, A. Identification, quantification, and relative concentrations of carotenoids, and their metabolites in human milk and serum. Anal. Chem. 1997, 69, 1873–1881. [Google Scholar] [CrossRef] [PubMed]
- Gossage, C.P.; Deyhim, M.; Yamini, S.; Douglas, L.W.; Moser-Veillon, P.B. Carotenoid composition of human milk during the first month postpartum and the response to β-carotene supplementation. Am. J. Clin. Nutr. 2002, 76, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Lipkie, T.E.; Morrow, A.L.; Jouni, Z.E.; McMahon, R.J.; Ferruzzi, M.G. Longitudinal survey of carotenoids in human milk from urban cohorts in China, Mexico, and the USA. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Galan, P.; Viteri, F.E.; Bertrais, S.; Czernichow, S.; Faure, H.; Arnaud, J.; Ruffieux, D.; Chenal, S.; Arnault, N.; Favier, A.; et al. Serum concentrations of beta-carotene, vitamins C and E, zinc and selenium are influenced by sex, age, diet, smoking status, alcohol consumption and corpulence in a general French adult population. Eur. J. Clin. Nutr. 2005, 59, 1181–1190. [Google Scholar] [CrossRef] [PubMed]
- Forman, M.R.; Beecher, G.R.; Lanza, E.; Reichman, M.E.; Graubard, B.I.; Campbell, W.S.; Marr, T.; Yong, L.C.; Judd, J.T.; Taylor, P.R. Effect of alcohol consumption on plasma carotenoid concentrations in premenopausal women, a controlled dietary study. Am. J. Clin. Nutr. 1995, 62, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Schweigert, F.J.; Bathe, K.; Chen, F.; Büscher, U.; Dudenhausen, J.W. Effect of the stage of lactation in humans on carotenoid levels in milk, blood plasma and plasma lipoprotein fractions. Eur. J. Nutr. 2004, 43, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Ríos, J.J.; Xavier, A.A.O.; Díaz-Salido, E.; Arenilla-Vélez, I.; Jarén-Galán, M.; Garrido-Fernández, J.; Aguayo-Maldonado, J.; Pérez-Gálvez, A. Xanthophyll esters are found in human colostrum. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Bokor, S.; Koletzko, B.; Decsi, T. Systematic review of fatty acid composition of human milk from mothers of preterm compared to full-term infants. Ann. Nutr. Metab. 2007, 51, 550–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bobiński, R.; Mikulska, M.; Mojska, H.; Simon, M. Comparison of the fatty acid composition of transitional and mature milk of mothers who delivered healthy full-term babies, preterm babies and full-term small for gestational age infants. Eur. J. Clin. Nutr. 2013, 67, 966–971. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Gálvez, A.; Martin, H.D.; Sies, H.; Stahl, W. Incorporation of carotenoids from paprika oleoresin into human chylomicrons. Br. J. Nutr. 2003, 89, 787–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breithaupt, D.E.; Wirt, U.; Bamedi, A. Differentiation between lutein monoester regioisomers and detection of lutein diesters from marigold flowers (Tagetes erecta L.) and several fruits by liquid chromatography-mass spectrometry. J. Agric. Food Chem. 2002, 50, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Liaaen-Jensen, S. Combined approach, identification and structure elucidation of carotenoids. In Carotenoids, Volume 1B, Spectroscopy; Britton, G., Liaaen-Jensen, S., Pfander, H., Eds.; Birkhäuser Verlag: Basel, Switzerland, 1995; pp. 343–354. ISBN 0817629092. [Google Scholar]
- Enzell, C.R.; Back, S. Mass spectrometry. In Carotenoids, Volume 1B, Spectroscopy; Britton, G., Liaaen-Jensen, S., Pfander, H., Eds.; Birkhäuser Verlag: Basel, Switzerland, 1995; pp. 261–320. ISBN 0817629092. [Google Scholar]
- Britton, G.; Liaaen-Jensen, S.; Pfander, H. Carotenoids Handbook; Birkhäuser Verlag: Basel, Switzerland, 2004; ISBN 3034878362. [Google Scholar]
- Rodriguez-Amaya, D.B. Food Carotenoids, Chemistry, Biology, and Technology; Wiley Blackwell: Singapore, 2016. [Google Scholar]
- Rivera, S.M.; Christou, P.; Canela-Garayoa, R. Identification of carotenoids using mass spectrometry. Mass Spectrom. Rev. 2014, 33, 353–372. [Google Scholar] [CrossRef] [PubMed]
- De Rosso, V.V.; Mercadante, A.Z. Identification and quantification of carotenoids, by HPLC-PDA-MS/MS, from Amazonian fruits. J. Agric. Food Chem. 2007, 55, 5062–5072. [Google Scholar] [CrossRef] [PubMed]
- Weller, P.; Breithaupt, D.E. Identification and quantification of zeaxanthin esters in plants using liquid chromatography-mass spectrometry. J. Agric. Food Chem. 2003, 51, 7044–7049. [Google Scholar] [CrossRef] [PubMed]
- Jensen, R.G.; Lammi-Keefe, C.J.; Koletzko, B. Representative sampling of human milk and the extraction of fat for analysis of environmental lipophilic contaminants. Toxicol. Environ. Chem. 1997, 62, 229–247. [Google Scholar] [CrossRef]
- Gebre-Medhin, M.; Vahlquist, A.; Hofvander, Y.; Uppsall, L.; Vahlquist, B. Breast milk composition in Ethiopean and Swedish mothers. I. Vitamin A and beta-carotene. Am. J. Clin. Nutr. 1976, 29, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Sommerburg, O.; Meissner, K.; Nelle, M.; Lenhartz, H.; Leichsenring, M. Carotenoid supply in breast-fed neonates. Eur. J. Pediatr. 2000, 159, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Macias, C.; Schweigert, F.J. Changes in the concentration of carotenoids, vitamin A, alpha-tocopherol and total lipids in human milk throughout early lactation. Ann. Nutr. Metab. 2001, 45, 82–85. [Google Scholar] [CrossRef] [PubMed]
- Jewell, V.C.; Northrop-Clewes, C.A.; Tubman, R.; Thurnham, D.I. Nutritional factors and visual function in premature infants. Proc. Nutr. Soc. 2001, 60, 171–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olmedilla, B.; Granado, F.; Southon, S.; Wright, A.J.A.; Blanco, I.; Gil-Martínez, E.; van den Berg, H.; Corridan, B.; Roussel, A.M.; Chopra, M.; et al. Serum concentrations of carotenoids and vitamins A, E, and C in control subjects from five European countries. Br. J. Nutr. 2001, 85, 227–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thurnham, D.I.; Northrop-Clewes, C.A.; Chopra, M. Biomarkers of vegetable and fruit intakes. Am. J. Clin. Nutr. 1998, 68, 756–757. [Google Scholar] [CrossRef] [PubMed]
- Couillard, C.; Lemieux, S.; Vohl, M.C.; Couture, P.; Lamarche, B. Carotenoids as biomarkers of fruit and vegetable intake in men and women. Br. J. Nutr. 2016, 116, 1206–1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borel, P.; Lietz, G.; Goncalves, A.; Edelenyi, F.S.; Lecompte, S.; Curtis, P.; Goumidi, L.; Caslake, M.J.; Miles, E.A.; Packard, C.; et al. CD36 and SR-BI are involved in cellular uptake of provitamin A carotenoids by Caco-2 and HEK cells, and some of their genetic variants are associated with plasma concentrations of these micronutrients in humans. J. Nutr. 2013, 143, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Moussa, M.; Gouranton, E.; Gleize, B.; Yazidi, C.E.; Niot, I.; Besnard, P.; Borel, P.; Landrier, J.F. CD36 is involved in lycopene and lutein uptake by adipocytes and adipose tissue cultures. Mol. Nutr. Food Res. 2011, 55, 578–584. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Suzuki, R.; Kobayashi, M.; Itagaki, S.; Hirano, T.; Noda, T.; Mizuno, S.; Sugawara, M.; Iseki, K. Involvement of cholesterol membrane transporter Niemann-Pick C1-like 1 in the intestinal absorption of lutein. J. Pharm. Pharm. Sci. 2012, 15, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Landschulz, K.; Pathak, R.; Rigotti, A.; Krieger, M.; Hobbs, H. Regulation of scavenger receptor, class B, type I, a high density lipoprotein receptor, in liver and steroidogenic tissues of the rat. J. Clin. Invest. 1996, 98, 984–995. [Google Scholar] [CrossRef] [PubMed]
- Pussinen, P.J.; Karten, B.; Wintersperger, A.; Reicher, H.; McLean, M.; Malle, E.; Sattler, W. The human breast carcinoma cell line HBL-100 acquires exogenous cholesterol from high-density lipoprotein via CLA-1 (CD-36 and LIMPII analogous 1)-mediated selective cholesteryl ester uptake. Biochem. J. 2000, 349, 559–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bionaz, M.; Loor, J.J. Gene networks driving bovine milk fat synthesis during the lactation cycle. BMC Genomics 2008, 9, 366. [Google Scholar] [CrossRef] [PubMed]
- Macías, H.; Hinck, L. Mammary gland development. Wiley Interdiscip. Rev. Dev. Biol. 2012, 1, 533–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmer, J.P.; Hammond, B.R.Jr. Possible influences of lutein and zeaxanthin on the developing retina. Clin. Ophthalmol. 2007, 1, 25–35. [Google Scholar] [PubMed]


{kind=link}
| PT Mothers 1 | FT Mothers 1 | |||||
|---|---|---|---|---|---|---|
| 25th Percentile | Median | 75th Percentile | 25th Percentile | Median | 75th Percentile | |
| Colostrum 2 | ||||||
| Zeaxanthin (X) 3 | 35.9 | 63.2 | 112.6 | 73.7 | 106.4 | 141.4 |
| Lutein (X) | 231.6 | 432.8 a | 667.9 | 322.9 | 486.3 a | 745.8 |
| β-Cryptoxanthin (X) | 231.4 | 406.7 | 853.0 | 429.6 | 754.6 | 1486 |
| Lycopene (C) 4 | 388.1 | 669.9 | 931.6 | 483.0 | 1065 | 1846 |
| α+β-Carotene (C) | 331.0 | 594.6 | 1364 | 602.3 | 1103 | 2238 |
| Xanthophyll esters (X) | 144.5 | 334.6 b | 733.4 | 251.5 | 598.7 b | 1211 |
| Total | 1810 | 2641 | 4498 | 2854 | 4961 | 7209 |
| Mature milk 5 | ||||||
| Zeaxanthin | 36.7 | 46.6 | 65.9 | 38.4 | 59.9 | 91.9 |
| Lutein | 170.0 | 217.3 | 283.0 | 150.3 | 195.9 | 270.8 |
| β-Cryptoxanthin | 53.0 | 135.1 | 224.6 | 87.4 | 190.6 | 353.9 |
| Lycopene | 89.4 | 125.8 | 173.6 | 118.8 | 192.7 | 221.2 |
| α+β-Carotene | 50.0 | 106.4 | 202.2 | 59.4 | 84.8 | 315.0 |
| Xanthophyll esters | 0 | 0 | 0 | 0 | 0 | 0 |
| Total | 508.7 | 693.5 | 934.0 | 512.3 | 768.0 | 1174 |
| PT Mothers | FT Mothers | |||||
|---|---|---|---|---|---|---|
| 25th Percentile | Median | 75th Percentile | 25th percentile | Median | 75th Percentile | |
| Serum samples collected at 3–5 day postpartum 1 | ||||||
| Zeaxanthin | 108.2 | 142.9 | 171.8 | 94.4 | 122.1 | 152.4 |
| Lutein | 658.3 | 744.8 | 911.9 | 552.4 | 656.0 | 834.7 |
| β-Cryptoxanthin | 419.8 | 814.1 | 964.7 | 617.8 | 737.3 | 1084 |
| Lycopene | 481.7 | 712.9 | 980.2 | 450.3 | 673.5 | 887.3 |
| α+β-Carotene | 519.6 | 816.8 | 1089 | 494.6 | 671.2 | 1547 |
| Xanthophyll esters | 0 | 0 | 0 | 0 | 0 | 0 |
| Total | 2984 | 3288 | 3751 | 2869 | 3222 | 3811 |
| Serum samples collected at 15–30 day postpartum 1 | ||||||
| Zeaxanthin | 105.3 | 136.1 | 187.4 | 96.0 | 135.5 | 158.6 |
| Lutein | 622.6 | 768.4 | 1010 | 579.0 | 673.9 | 809.1 |
| β-Cryptoxanthin | 329.9 | 729.3 | 1152 | 669.2 | 822.4 | 925.2 |
| Lycopene | 555.3 | 824.0 | 1007 | 473.9 | 730.4 | 905.7 |
| α+β-Carotene | 583.3 | 967.4 | 1594 | 549.2 | 743.8 | 1087 |
| Xanthophyll esters | 0 | 0 | 0 | 0 | 0 | 0 |
| Total | 3279 | 3636 | 3898 | 2938 | 3295 | 3711 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xavier, A.A.O.; Díaz-Salido, E.; Arenilla-Vélez, I.; Aguayo-Maldonado, J.; Garrido-Fernández, J.; Fontecha, J.; Sánchez-García, A.; Pérez-Gálvez, A. Carotenoid Content in Human Colostrum is Associated to Preterm/Full-Term Birth Condition. Nutrients 2018, 10, 1654. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10111654
Xavier AAO, Díaz-Salido E, Arenilla-Vélez I, Aguayo-Maldonado J, Garrido-Fernández J, Fontecha J, Sánchez-García A, Pérez-Gálvez A. Carotenoid Content in Human Colostrum is Associated to Preterm/Full-Term Birth Condition. Nutrients. 2018; 10(11):1654. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10111654
Chicago/Turabian StyleXavier, Ana A. O., Elena Díaz-Salido, Isabel Arenilla-Vélez, Josefa Aguayo-Maldonado, Juan Garrido-Fernández, Javier Fontecha, Alicia Sánchez-García, and Antonio Pérez-Gálvez. 2018. "Carotenoid Content in Human Colostrum is Associated to Preterm/Full-Term Birth Condition" Nutrients 10, no. 11: 1654. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10111654