Cod Residual Protein Prevented Blood Pressure Increase in Zucker fa/fa Rats, Possibly by Inhibiting Activities of Angiotensin-Converting Enzyme and Renin

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Design
2.3. Preparation of Fish Meals
2.4. Diets
2.5. Analyses of Diets
2.6. In Vitro Inhibition of Angiotensin Converting Enzyme (ACE) and Renin
2.7. Analyses in Serum and Urine
2.8. Statistical Analyses
3. Results
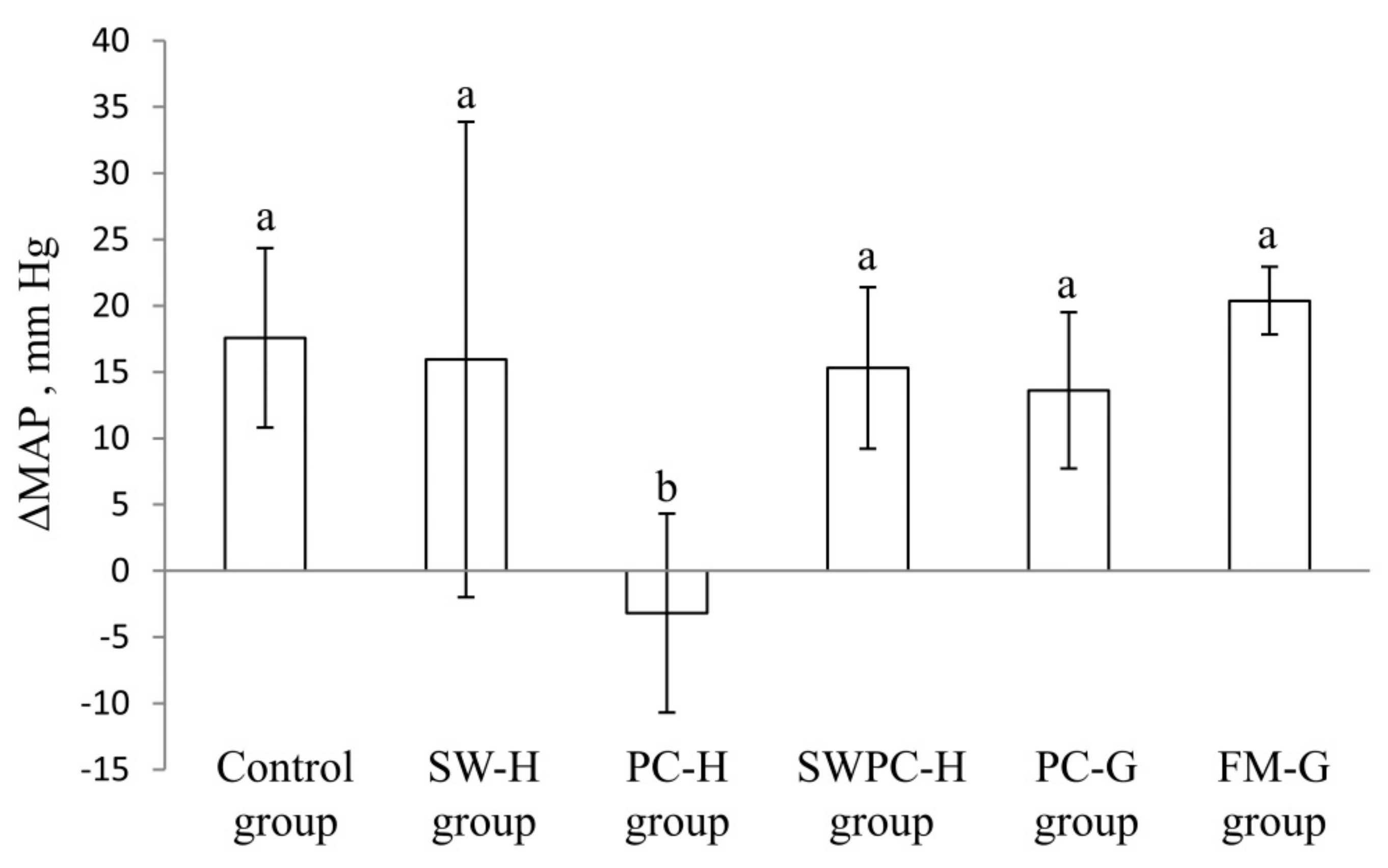
3.1. Measurements of Blood Pressure
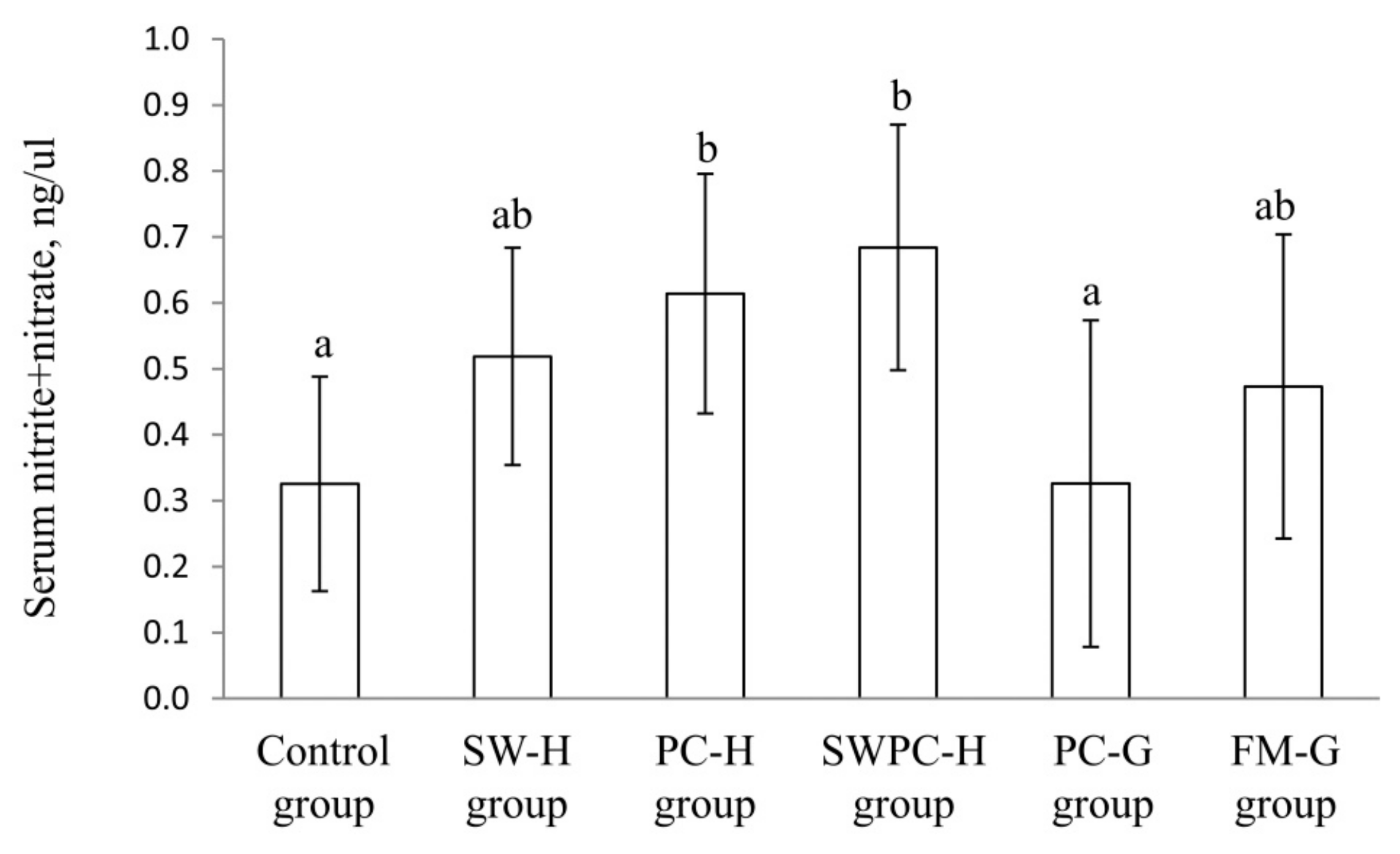
3.2. Circulating Concentration of Nitrite + Nitrate
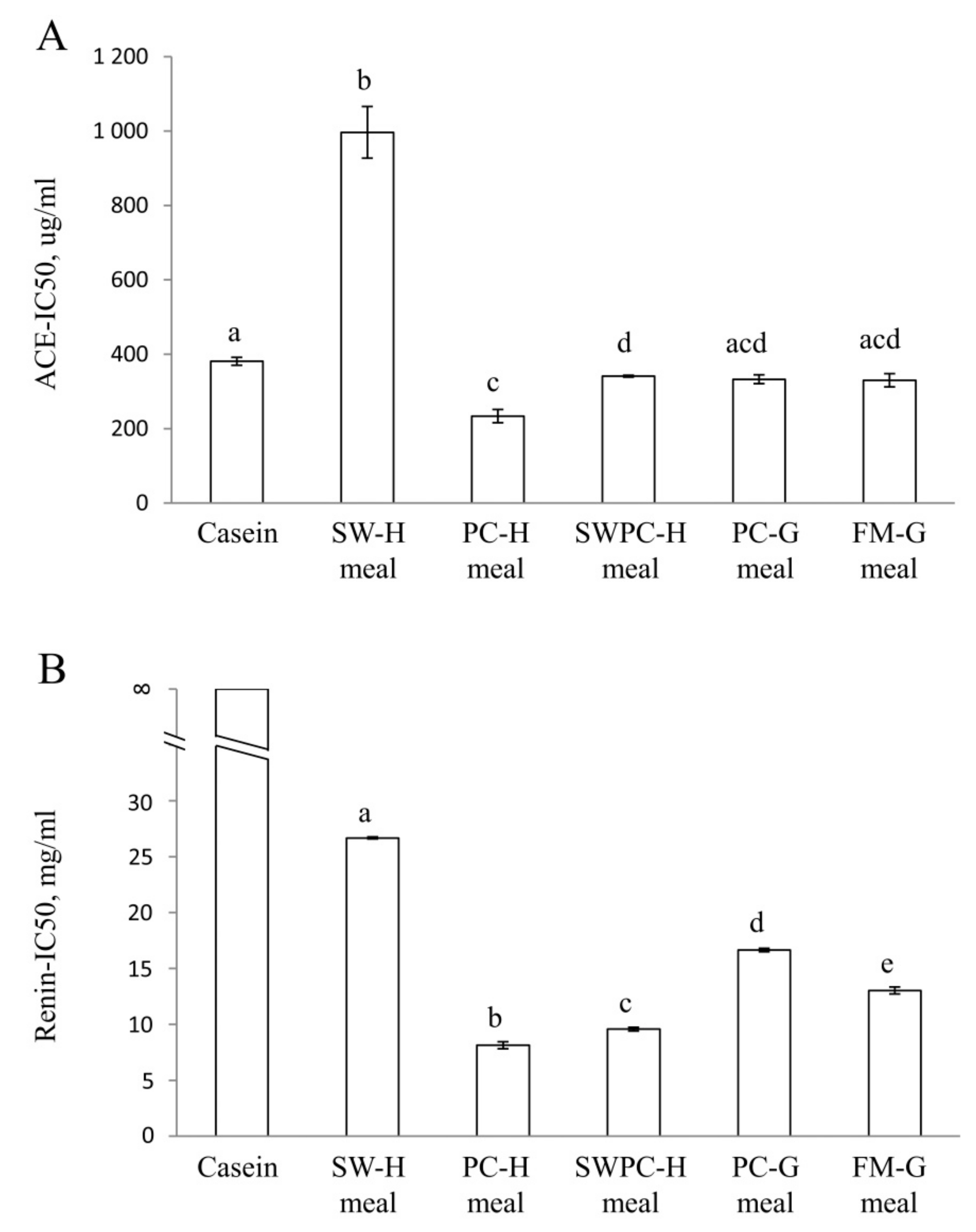
3.3. In Vitro Inhibition of ACE and Renin Activities
3.4. Growth and Dietary Intake
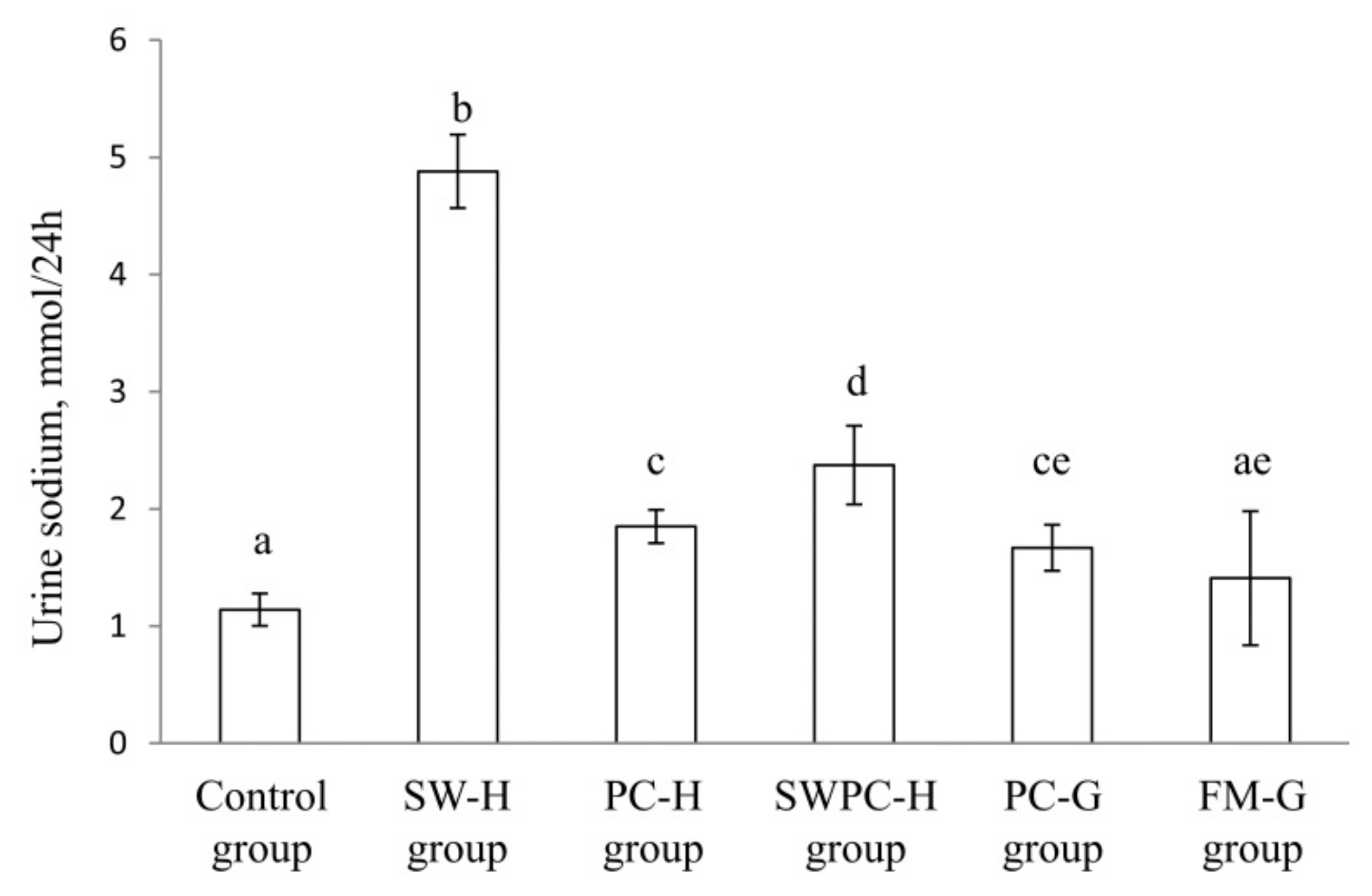
3.5. Urine Sodium Excretion
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kearney, P.M.; Whelton, M.; Reynolds, K.; Muntner, P.; Whelton, P.K.; He, J. Global burden of hypertension: Analysis of worldwide data. Lancet 2005, 365, 217–223. [Google Scholar] [CrossRef]
- Danaei, G.; Lu, Y.; Singh, G.; Stevens, G.; Cowan, M.; Farzadfar, F.; Lin, J.; Finucane, M.; Rao, M.; Khang, Y.; et al. Cardiovascular disease, chronic kidney disease, and diabetes mortality burden of cardiometabolic risk factors from 1980 to 2010: A comparative risk assessment. Lancet 2014, 2, 634–647. [Google Scholar]
- Olsen, M.H.; Angell, S.Y.; Asma, S.; Boutouyrie, P.; Burger, D.; Chirinos, J.A.; Damasceno, A.; Delles, C.; Gimenez-Roqueplo, A.P.; Hering, D.; et al. A call to action and a lifecourse strategy to address the global burden of raised blood pressure on current and future generations: The Lancet Commission on hypertension. Lancet 2016, 388, 2665–2712. [Google Scholar] [CrossRef]
- Kannel, W.B. Blood pressure as a cardiovascular risk factor: Prevention and treatment. JAMA 1996, 275, 1571–1576. [Google Scholar] [CrossRef] [PubMed]
- The World Health Organization. A Global Brief on Hypertension: Silent Killer, Global Public Health Crisis; WHO: Geneva, Switzerland, 2013. [Google Scholar]
- Whelton, P.K.; Carey, R.M.; Aronow, W.S.; Casey, D.E., Jr.; Collins, K.J.; Dennison Himmelfarb, C.; DePalma, S.M.; Gidding, S.; Jamerson, K.A.; Jones, D.W.; et al. 2017 ACC/AHA/AAPA/ABC/ACPM/AGS/APhA/ASH/ASPC/NMA/PCNA Guideline for the Prevention, Detection, Evaluation, and Management of High Blood Pressure in Adults: A Report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines. J. Am. Coll. Cardiol. 2018, 71, e127–e248. [Google Scholar] [PubMed]
- Zaman, M.A.; Oparil, S.; Calhoun, D.A. Drugs targeting the renin-angiotensin-aldosterone system. Nat. Rev. Drug Discov. 2002, 1, 621–636. [Google Scholar] [CrossRef] [PubMed]
- Palmer, R.M.; Ashton, D.S.; Moncada, S. Vascular endothelial cells synthesize nitric oxide from L-arginine. Nature 1988, 333, 664–666. [Google Scholar] [CrossRef] [PubMed]
- Blaustein, M.P.; Leenen, F.H.; Chen, L.; Golovina, V.A.; Hamlyn, J.M.; Pallone, T.L.; Van Huysse, J.W.; Zhang, J.; Wier, W.G. How NaCl raises blood pressure: A new paradigm for the pathogenesis of salt-dependent hypertension. Am. J. Physiol. Heart Circ. Physiol. 2012, 302, H1031–H1049. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.O.; Freeman, R.H. Mechanisms regulating renin release. Physiol. Rev. 1976, 56, 1–56. [Google Scholar] [CrossRef] [PubMed]
- Panagiotakos, D.B.; Zeimbekis, A.; Boutziouka, V.; Economou, M.; Kourlaba, G.; Toutouzas, P.; Polychronopoulos, E. Long-term fish intake is associated with better lipid profile, arterial blood pressure, and blood glucose levels in elderly people from Mediterranean islands (MEDIS epidemiological study). Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2007, 13, CR307–CR312. [Google Scholar]
- Ramel, A.; Martinez, J.A.; Kiely, M.; Bandarra, N.M.; Thorsdottir, I. Moderate consumption of fatty fish reduces diastolic blood pressure in overweight and obese European young adults during energy restriction. Nutrition 2010, 26, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Zaribaf, F.; Falahi, E.; Barak, F.; Heidari, M.; Keshteli, A.H.; Yazdannik, A.; Esmaillzadeh, A. Fish consumption is inversely associated with the metabolic syndrome. Eur. J. Clin. Nutr. 2014, 68, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Ke, L.; Ho, J.; Feng, J.; Mpofu, E.; Dibley, M.J.; Feng, X.; Van, F.; Leong, S.; Lau, W.; Lueng, P.; et al. Modifiable risk factors including sunlight exposure and fish consumption are associated with risk of hypertension in a large representative population from Macau. J. Steroid Biochem. Mol. Biol. 2014, 144, 152–155. [Google Scholar] [CrossRef] [PubMed]
- Torris, C.; Molin, M.; Cvancarova, M.S. Lean fish consumption is associated with lower risk of metabolic syndrome: A Norwegian cross sectional study. BMC Public Health 2016, 16, 347. [Google Scholar] [CrossRef] [PubMed]
- Ait-Yahia, D.; Madani, S.; Savelli, J.L.; Prost, J.; Bouchenak, M.; Belleville, J. Dietary fish protein lowers blood pressure and alters tissue polyunsaturated fatty acid composition in spontaneously hypertensive rats. Nutrition 2003, 19, 342–346. [Google Scholar] [CrossRef]
- Vikoren, L.A.; Drotningsvik, A.; Mwakimonga, A.; Leh, S.; Mellgren, G.; Gudbrandsen, O.A. Diets containing salmon fillet delay development of high blood pressure and hyperfusion damage in kidneys in obese Zucker fa/fa rats. J. Am. Soc. Hypertens. 2018, 12, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Ness, A.R.; Whitley, E.; Burr, M.L.; Elwood, P.C.; Smith, G.D.; Ebrahim, S. The long-term effect of advice to eat more fish on blood pressure in men with coronary disease: Results from the diet and reinfarction trial. J. Hum. Hypertens. 1999, 13, 729–733. [Google Scholar] [CrossRef] [PubMed]
- Grieger, J.A.; Miller, M.D.; Cobiac, L. Investigation of the effects of a high fish diet on inflammatory cytokines, blood pressure, and lipids in healthy older Australians. Food Nutr. Res. 2014, 58, 20369. [Google Scholar] [CrossRef] [PubMed]
- von Houwelingen, R.; Nordoy, A.; van der Beek, E.; Houtsmuller, U.; de Metz, M.; Hornstra, G. Effect of a moderate fish intake on blood pressure, bleeding time, hematology, and clinical chemistry in healthy males. Am. J. Clin. Nutr. 1987, 46, 424–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngo, D.H.; Vo, T.S.; Ngo, D.N.; Wijesekara, I.; Kim, S.K. Biological activities and potential health benefits of bioactive peptides derived from marine organisms. Int. J. Biol. Macromol. 2012, 51, 378–383. [Google Scholar] [CrossRef] [PubMed]
- Vercruysse, L.; Van Camp, J.; Smagghe, G. ACE inhibitory peptides derived from enzymatic hydrolysates of animal muscle protein: A review. J. Agric. Food Chem. 2005, 53, 8106–8115. [Google Scholar] [CrossRef] [PubMed]
- Darewicz, M.; Borawska, J.; Vegarud, G.E.; Minkiewicz, P.; Iwaniak, A. Angiotensin I-converting enzyme (ACE) inhibitory activity and ACE inhibitory peptides of salmon (Salmo salar) protein hydrolysates obtained by human and porcine gastrointestinal enzymes. Int. J. Mol. Sci. 2014, 15, 14077–14101. [Google Scholar] [CrossRef] [PubMed]
- Jensen, I.J.; Eysturskareth, J.; Madetoja, M.; Eilertsen, K.E. The potential of cod hydrolyzate to inhibit blood pressure in spontaneously hypertensive rats. Nutr. Res. 2014, 34, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Je, J.Y.; Park, P.J.; Byun, H.G.; Jung, W.K.; Kim, S.K. Angiotensin I converting enzyme (ACE) inhibitory peptide derived from the sauce of fermented blue mussel, Mytilus edulis. Bioresour. Technol. 2005, 96, 1624–1629. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.J.; Je, J.Y.; Kim, S.K. Antihypertensive effect of angiotensin i converting enzyme-inhibitory peptide from hydrolysates of Bigeye tuna dark muscle, Thunnus obesus. J. Agric. Food Chem. 2007, 55, 8398–8403. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, B.; Dong, S.; Liu, Z.; Zhao, X.; Wang, J.; Zeng, M. A novel ACE inhibitory peptide isolated from Acaudina molpadioidea hydrolysate. Peptides 2009, 30, 1028–1033. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization. FAO Fisheries Department: The State of World Fisheries and Aquaculture; Food and Agriculture Organization: Rome, Italy, 2014. [Google Scholar]
- de Artinano, A.A.; Castro, M.M. Experimental rat models to study the metabolic syndrome. Br. J. Nutr. 2009, 102, 1246–1253. [Google Scholar] [CrossRef] [PubMed]
- Franklin, S.S.; Gustin, W.t.; Wong, N.D.; Larson, M.G.; Weber, M.A.; Kannel, W.B.; Levy, D. Hemodynamic patterns of age-related changes in blood pressure. The Framingham Heart Study. Circulation 1997, 96, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, T.W.; Morris, R.C.; Pershadsingh, H.A. The Zucker fatty rat as a genetic model of obesity and hypertension. Hypertension 1989, 13, 896–901. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Wang, X.; Chen, C.; Wang, J.; Zou, X.; Li, C.; Xu, Z.; Yang, X.; Shi, W.; Zeng, C. Oxidative stress causes imbalance of renal renin angiotensin system (RAS) components and hypertension in obese Zucker rats. J. Am. Heart Assoc. 2015, 4, e001559. [Google Scholar] [CrossRef] [PubMed]
- Pinto, Y.M.; Paul, M.; Ganten, D. Lessons from rat models of hypertension: From Goldblatt to genetic engineering. Cardiovasc. Res. 1998, 39, 77–88. [Google Scholar] [CrossRef]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C., Jr. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef] [PubMed]
- Reeves, P.G. AIN-93 purified diets for the study of trace element metabolism in rodents. In Trace Elements in Laboratory Rodents; Watson, R.R., Ed.; CRC Press Inc.: Boca Raton, FL, USA, 1996; pp. 3–37. [Google Scholar]
- Cohen, S.A.; Michaud, D.P. Synthesis of a Fluorescent Derivatizing Reagent, 6-Aminoquinolyl-N-Hydroxysuccinimidyl Carbamate, and its Application for the Analysis of Hydrolysate Amino Acids via High-Performance Liquid Chromatography. Anal. Biochem. 1993, 211, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.L. Determination of the tryptophan content of feedingstuffs with particular reference to cereals. J. Sci. Food Agric. 1967, 18, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Bidlingmeyer, B.A.; Cohen, S.A.; Tarvin, T.L.; Frost, B. A new, rapid, high-sensitivity analysis of amino acids in food type samples. J. Assoc. Off. Anal. Chem. 1987, 70, 241–247. [Google Scholar] [PubMed]
- Oterhals, A.; Nygard, E. Reduction of persistent organic pollutants in fishmeal: A feasibility study. J. Agric. Food Chem. 2008, 56, 2012–2020. [Google Scholar] [CrossRef] [PubMed]
- International Organization for Standardization. Animal Feeding Stuffs—Determination of the Contents of Calcium, Copper, Iron, Magnesium, Manganese, Potassium, Sodium and Zinc—Method Using Atomic Absorption Spectrometry (ISO 6869:2000); International Organization for Standardization: Geneva, Switzerland, 2000. [Google Scholar]
- Association of Official Agricultural Chemists. AOAC Official Method 937.09 Salt (Chlorine as Sodium Chloride) in Seafood; Association of Official Agricultural Chemists: Arlington, VA, USA, 2013. [Google Scholar]
- Shalaby, S.M.; Zakora, M.; Otte, J. Performance of two commonly used angiotensin-converting enzyme inhibition assays using FA-PGG and HHL as substrates. J. Dairy Res. 2006, 73, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Drotningsvik, A.; Midttun, O.; McCann, A.; Ueland, P.M.; Hogoy, I.; Gudbrandsen, O.A. Dietary intake of cod protein beneficially affects concentrations of urinary markers of kidney function and results in lower urinary loss of amino acids in obese Zucker fa/fa rats. Br. J. Nutr. 2018, 120, 740–750. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Wang, B.; Li, Y.; Sun, F.; Li, P.; Xia, W.; Zhou, X.; Li, Q.; Wang, X.; Chen, J.; et al. Taurine Supplementation Lowers Blood Pressure and Improves Vascular Function in Prehypertension: Randomized, Double-Blind, Placebo-Controlled Study. Hypertension 2016, 67, 541–549. [Google Scholar] [CrossRef] [PubMed]
- van den Elsen, L.W.; Spijkers, L.J.; van den Akker, R.F.; van Winssen, A.M.; Balvers, M.; Wijesinghe, D.S.; Chalfant, C.E.; Garssen, J.; Willemsen, L.E.; Alewijnse, A.E.; et al. Dietary fish oil improves endothelial function and lowers blood pressure via suppression of sphingolipid-mediated contractions in spontaneously hypertensive rats. J. Hypertens. 2014, 32, 1050–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frenoux, J.M.; Prost, E.D.; Belleville, J.L.; Prost, J.L. A polyunsaturated fatty acid diet lowers blood pressure and improves antioxidant status in spontaneously hypertensive rats. J. Nutr. 2001, 131, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Coimbra, T.M.; Janssen, U.; Grone, H.J.; Ostendorf, T.; Kunter, U.; Schmidt, H.; Brabant, G.; Floege, J. Early events leading to renal injury in obese Zucker (fatty) rats with type II diabetes. Kidney Int. 2000, 57, 167–182. [Google Scholar] [CrossRef] [PubMed]
- Skeggs, L.T., Jr.; Kahn, J.R.; Lentz, K.; Shumway, N.P. The preparation, purification, and amino acid sequence of a polypeptide renin substrate. J. Exp. Med. 1957, 106, 439–453. [Google Scholar] [CrossRef] [PubMed]
- FitzGerald, R.J.; Murray, B.A.; Walsh, D.J. Hypotensive peptides from milk proteins. J. Nutr. 2004, 134, 980S–988S. [Google Scholar] [CrossRef] [PubMed]
- Girgih, A.T.; Nwachukwu, I.D.; Hasan, F.; Fagbemi, T.N.; Gill, T.; Aluko, R.E. Kinetics of the inhibition of renin and angiotensin I-converting enzyme by cod (Gadus morhua) protein hydrolysates and their antihypertensive effects in spontaneously hypertensive rats. Food Nutr. Res. 2015, 59, 29788. [Google Scholar] [CrossRef] [PubMed]
- McCoy, R.H. Dietary requirements of the rat. In The Rat in Laboratory Investigation; Griffith, J.Q., Farris, E.J., Eds.; J. B. Lippincott Company: Philadelphia, PA, USA, 1949; pp. 67–101. [Google Scholar]
- Trachtman, H.; Del Pizzo, R.; Rao, P.; Rujikarn, N.; Sturman, J.A. Taurine lowers blood pressure in the spontaneously hypertensive rat by a catecholamine independent mechanism. Am. J. Hypertens. 1989, 2, 909–912. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.C.; Sacks, F.; Rosner, B. Does fish oil lower blood pressure? A meta-analysis of controlled trials. Circulation 1993, 88, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.A.; Woodman, R.J. The independent effects of eicosapentaenoic acid and docosahexaenoic acid on cardiovascular risk factors in humans. Curr. Opin. Clin. Nutr. Metab. Care 2006, 9, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Appel, L.J.; Brands, M.W.; Daniels, S.R.; Karanja, N.; Elmer, P.J.; Sacks, F.M.; American Heart, A. Dietary approaches to prevent and treat hypertension: A scientific statement from the American Heart Association. Hypertension 2006, 47, 296–308. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.; Whitesall, S.; Zhang, Y.; Beibel, M.; D’Alecy, L.; DiPetrillo, K. Validation of volume-pressure recording tail-cuff blood pressure measurements. Am. J. Hypertens. 2008, 21, 1288–1291. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| g/kg Diet | Control Diet | SW-H Diet | PC-H Diet | SWPC-H Diet | PC-G Diet | FM-G Diet |
|---|---|---|---|---|---|---|
| Casein * | 223.8 | 167.8 | 167.8 | 167.8 | 167.8 | 167.8 |
| Stickwater from Havstrand † | - | 82.2 | - | - | - | - |
| Presscake meal from Havstrand ‡ | - | - | 81.9 | - | - | - |
| Stickwater + Presscake mealfrom Havstrand § | - | - | - | 84.6 | - | - |
| Presscake meal from Granit || | - | - | - | - | 77.6 | - |
| Fillet meal from Granit # | - | - | - | - | - | 60.7 |
| Soybean Oil | 70.0 | 70.0 | 70.0 | 70.0 | 70.0 | 70.0 |
| Cornstarch | 504.1 | 477.8 | 478.2 | 475.5 | 482.5 | 499.3 |
| Sucrose | 90.0 | 90.0 | 90.0 | 90.0 | 90.0 | 90.0 |
| Cellulose | 50.0 | 50.0 | 50.0 | 50.0 | 50.0 | 50.0 |
| Tert-butylhydroquinone | 0.014 | 0.014 | 0.014 | 0.014 | 0.014 | 0.014 |
| Mineral Mix (AIN-93MX) | 35.0 | 35.0 | 35.0 | 35.0 | 35.0 | 35.0 |
| Vitamin Mix (AIN-93VX) | 10.0 | 10.0 | 10.0 | 10.0 | 10.0 | 10.0 |
| L-Methionine | 1.6 | 1.6 | 1.6 | 1.6 | 1.6 | 1.6 |
| L-Cystine | 3.0 | 3.0 | 3.0 | 3.0 | 3.0 | 3.0 |
| Choline Bitartrate ** | 2.5 | 2.5 | 2.5 | 2.5 | 2.5 | 2.5 |
| Growth and Maintenance Supplement †† | 10.0 | 10.0 | 10.0 | 10.0 | 10.0 | 10.0 |
| Control Group | SW-H Group | PC-H Group | SWPC Group | PC-G Group | FM-G Group | |
|---|---|---|---|---|---|---|
| MAP baseline, mm Hg | 111 ± 11 | 114 ± 5 | 120 ± 8 | 115 ± 10 | 110 ± 7 | 112 ± 8 |
| MAP Day 14, mm Hg | 126 ± 10 | 127 ± 7 | 115 ± 13 | 128 ± 8 | 127 ± 12 | 118 ± 8 |
| MAP endpoint, mm Hg | 128 ± 10 | 130 ± 18 | 117 ± 7 | 130 ± 13 | 124 ± 9 | 132 ± 10 |
| p for change baseline to Day 14 | 4.5 × 10−4 | 4.1 × 10−3 | 0.22 | 0.051 | 0.011 | 6.4 × 10−3 |
| p for change baseline to endpoint | 1.6 × 10−4 | 0.12 | 0.35 | 0.0016 | 2.4 × 10−3 | 6.5 × 10−6 |
| ΔMAP baseline to Day 14, mm Hg | 15 ± 7 ab | 12 ± 5 ab | −5 ± 9 c | 13 ± 12 ab | 17 ± 11 a | 6 ± 3 b |
| ΔMAP baseline to endpoint, mm Hg | 18 ± 7 a | 16 ± 18 a | −3 ± 8 b | 15 ± 6 a | 14 ± 6 a | 20 ± 3 a |
| Control Group | SW-H Group | PC-H Group | SWPC-H Group | PC-G Group | FM-G Group | p Anova | |
|---|---|---|---|---|---|---|---|
| Bodyweight at baseline, g | 339 ± 16 | 345 ± 15 | 340 ± 11 | 334 ± 14 | 335 ± 13 | 342 ± 16 | 0.81 |
| Body weight gain, % | 31 ± 6 | 37 ± 5 | 35 ± 3 | 37 ± 5 | 30 ± 4 | 34 ± 6 | 0.053 |
| Bodyweight-to-square body length without tail ratio, kg/m2 | 8.4 ± 0.5 | 8.3 ± 0.4 | 8.3 ± 0.4 | 8.2 ± 0.4 | 8.3 ± 0.3 | 8.4 ± 0.3 | 0.94 |
| Energy intake, kcal/kg bodyweight/24 h | 219 ± 31 ab | 238 ± 10 abc | 209 ± 14 a | 245 ± 20 bd | 212 ± 36 a | 248 ± 25 cd | 0.035 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vildmyren, I.; Drotningsvik, A.; Oterhals, Å.; Ween, O.; Halstensen, A.; Gudbrandsen, O.A. Cod Residual Protein Prevented Blood Pressure Increase in Zucker fa/fa Rats, Possibly by Inhibiting Activities of Angiotensin-Converting Enzyme and Renin. Nutrients 2018, 10, 1820. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10121820
Vildmyren I, Drotningsvik A, Oterhals Å, Ween O, Halstensen A, Gudbrandsen OA. Cod Residual Protein Prevented Blood Pressure Increase in Zucker fa/fa Rats, Possibly by Inhibiting Activities of Angiotensin-Converting Enzyme and Renin. Nutrients. 2018; 10(12):1820. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10121820
Chicago/Turabian StyleVildmyren, Iselin, Aslaug Drotningsvik, Åge Oterhals, Ola Ween, Alfred Halstensen, and Oddrun Anita Gudbrandsen. 2018. "Cod Residual Protein Prevented Blood Pressure Increase in Zucker fa/fa Rats, Possibly by Inhibiting Activities of Angiotensin-Converting Enzyme and Renin" Nutrients 10, no. 12: 1820. https://0-doi-org.brum.beds.ac.uk/10.3390/nu10121820