Alterations in IL-6/STAT3 Signaling by Korean Mistletoe Lectin Regulate the Self-Renewal Activity of Placenta-Derived Mesenchymal Stem Cells

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Line and Cell Culture
2.2. Cell Growth Assays
2.3. Differentiation into Mesodermal Lineages
2.4. Differentiation into Endodermal Lineages
2.5. MTT Assay
2.6. Neutralization of IL-6
2.7. RT-PCR and qRT-PCR
2.8. FACS Analysis
2.9. Genomic DNA Extraction and Sodium Bisulfite DNA Modification
2.10. qMSP
2.11. Western Blot Analysis
2.12. ELISA Analysis
2.13. Immunofluorescence
2.14. Statistical Analysis
3. Results
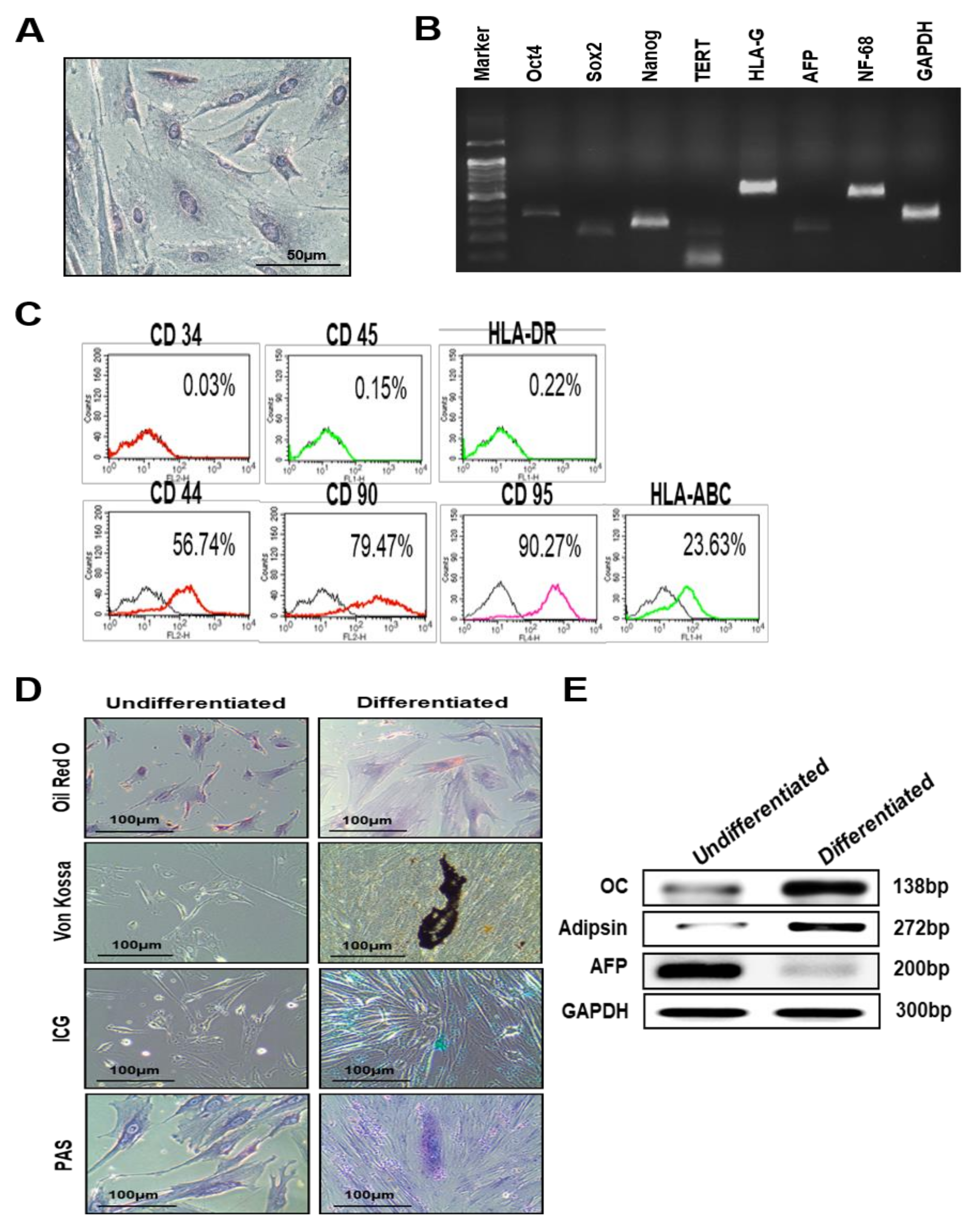
3.1. Characterization of PD-MSCs Isolated from Normal Term Placenta
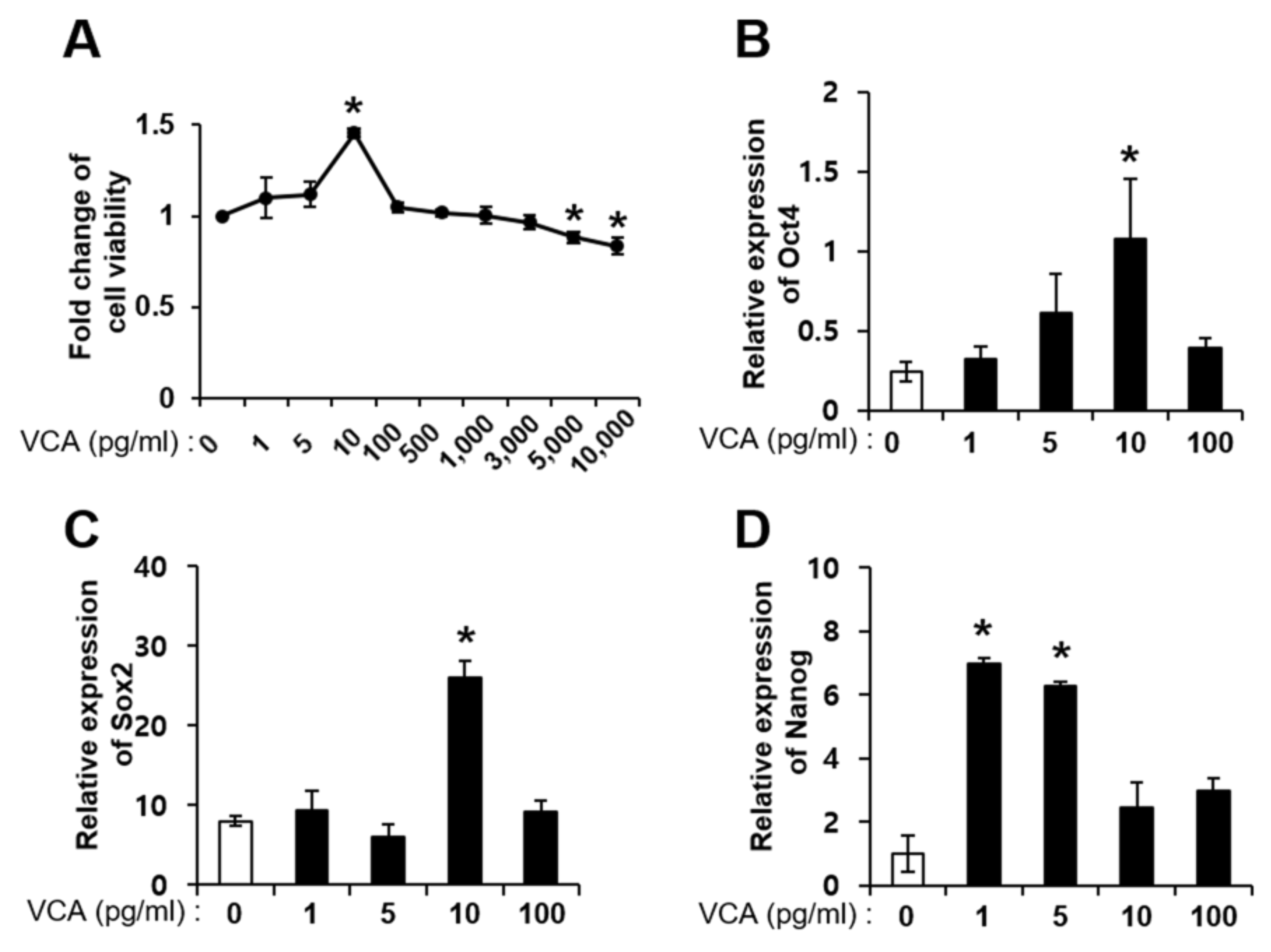
3.2. Effect of VCA on PD-MSC Self-Renewal
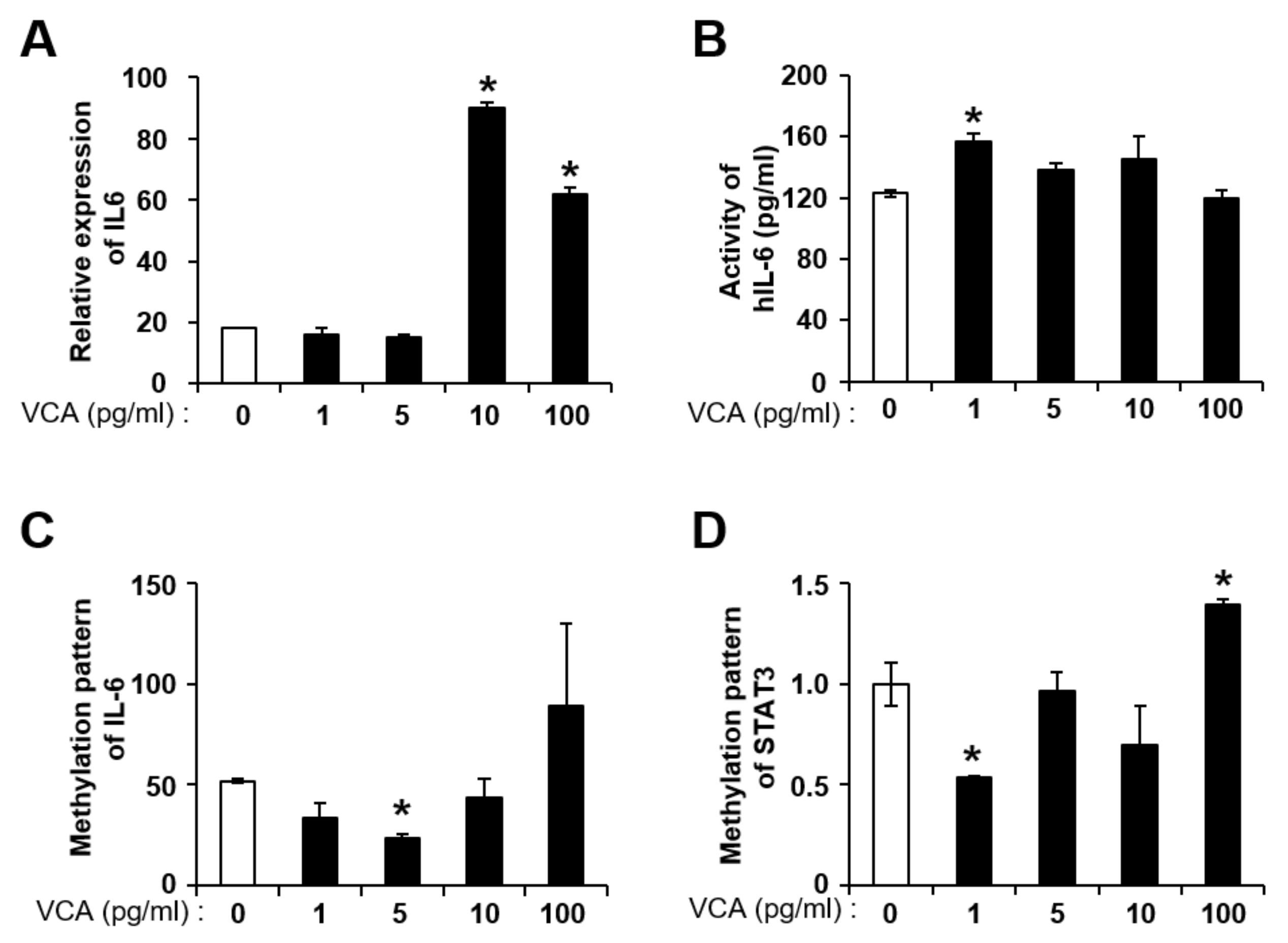
3.3. VCA Enhances the Self-Renewal of PD-MSCs by IL-6 Production
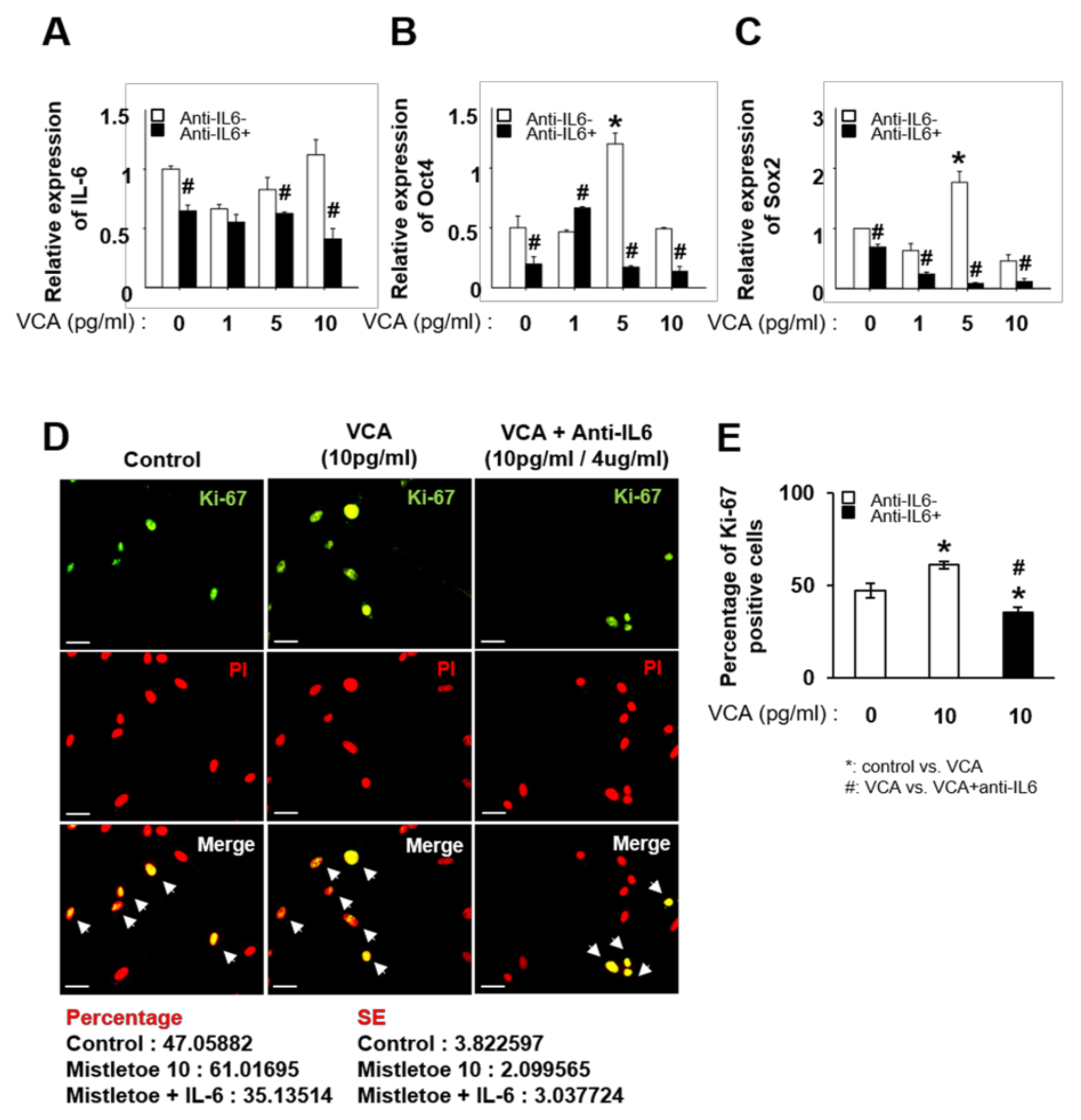
3.4. Inhibition of IL-6 Decreases the Proliferation of PD-MSCs Stimulated with VCA by Regulating Stemness Markers
3.5. VCA Enhances the Self-Renewal of PD-MSCs through IL-6-Mediated STAT3 Signaling
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, J.M.; Youd, M.E.; Lodie, T.A. Immunomodulatory activity of mesenchymal stem cells. Curr. Stem Cell Res. Ther. 2011, 6, 297–316. [Google Scholar] [CrossRef] [PubMed]
- De Miguel, M.P.; Fuentes-Julian, S.; Blazquez-Martinez, A.; Pascual, C.Y.; Aller, M.A.; Arias, J.; Arnalich-Montiel, F. Immunosuppressive properties of mesenchymal stem cells: Advances and applications. Curr. Mol. Med. 2012, 12, 574–591. [Google Scholar] [CrossRef] [PubMed]
- Karp, J.M.; Leng Teo, G.S. Mesenchymal stem cell homing: The devil is in the details. Cell Stem Cell 2009, 4, 206–216. [Google Scholar] [CrossRef]
- Selmani, Z.; Naji, A.; Zidi, I.; Favier, B.; Gaiffe, E.; Obert, L.; Borg, C.; Saas, P.; Tiberghien, P.; Rouas-Freiss, N.; et al. Human leukocyte antigen-G5 secretion by human mesenchymal stem cells is required to suppress T lymphocyte and natural killer function and to induce CD4+CD25highFOXP3+ regulatory T cells. Stem Cells 2008, 26, 212–222. [Google Scholar] [CrossRef]
- Luan, X.; Li, G.; Wang, G.; Wang, F.; Lin, Y. Human placenta-derived mesenchymal stem cells suppress T cell proliferation and support the culture expansion of cord blood CD34(+) cells: A comparison with human bone marrow-derived mesenchymal stem cells. Tissue Cell 2013, 45, 32–38. [Google Scholar] [CrossRef]
- Lee, J.M.; Jung, J.; Lee, H.J.; Jeong, S.J.; Cho, K.J.; Hwang, S.G.; Kim, G.J. Comparison of immunomodulatory effects of placenta mesenchymal stem cells with bone marrow and adipose mesenchymal stem cells. Int. Immunopharmacol. 2012, 13, 219–224. [Google Scholar] [CrossRef]
- Fazekasova, H.; Lechler, R.; Langford, K.; Lombardi, G. Placenta-derived MSCs are partially immunogenic and less immunomodulatory than bone marrow-derived MSCs. J. Tissue Eng. Regen. Med. 2011, 5, 684–694. [Google Scholar] [CrossRef]
- Pisciotta, A.; Riccio, M.; Carnevale, G.; Beretti, F.; Gibellini, L.; Maraldi, T.; Cavallini, G.M.; Ferrari, A.; Bruzzesi, G.; De Pol, A. Human serum promotes osteogenic differentiation of human dental pulp stem cells in vitro and in vivo. PLoS ONE 2012, 7, e50542. [Google Scholar] [CrossRef]
- Hong, L.; Zhang, G.; Sultana, H.; Yu, Y.; Wei, Z. The effects of 17-beta estradiol on enhancing proliferation of human bone marrow mesenchymal stromal cells in vitro. Stem Cells Dev. 2011, 20, 925–931. [Google Scholar] [CrossRef]
- Gharibi, B.; Hughes, F.J. Effects of medium supplements on proliferation, differentiation potential, and in vitro expansion of mesenchymal stem cells. Stem Cells Transl. Med. 2012, 1, 771–782. [Google Scholar] [CrossRef] [PubMed]
- Chieregato, K.; Castegnaro, S.; Madeo, D.; Astori, G.; Pegoraro, M.; Rodeghiero, F. Epidermal growth factor, basic fibroblast growth factor and platelet-derived growth factor-bb can substitute for fetal bovine serum and compete with human platelet-rich plasma in the ex vivo expansion of mesenchymal stromal cells derived from adipose tissue. Cytotherapy 2011, 13, 933–943. [Google Scholar] [CrossRef] [PubMed]
- He, X.; H’Ng, S.C.; Leong, D.T.; Hutmacher, D.W.; Melendez, A.J. Sphingosine-1-phosphate mediates proliferation maintaining the multipotency of human adult bone marrow and adipose tissue-derived stem cells. J. Mol. Cell Biol 2010, 2, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.P.; Wang, T.T.; Chen, W.; Wang, C.Y.; Chen, D.F.; Shen, J.G. Characterization of chemical components in extracts from Si-wu decoction with proliferation-promoting effects on rat mesenchymal stem cells. Bioorgan. Med. Chem. 2008, 16, 5109–5114. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.H.; Zeng, H.P.; Chen, D.F.; Li, H.; Li, X.C.; Li, Y.W.; Du, S.H. Effect of extract components from Plastrum testudinis and their combination component on proliferation of rat mesenchymal stem cell in vitro. Zhong Yao Cai 2006, 29, 452–455. [Google Scholar] [PubMed]
- Li, X.; Zhou, J.; Li, H.; Du, S.; Li, Y.; Huang, L.; Chen, D. Study on proliferation effect of extracts of Piper longum on mesenchymal stem cells of rat bone marrow and the relationship to chemical functional groups. Zhong Yao Cai 2005, 28, 570–574. [Google Scholar] [PubMed]
- Potu, B.K.; Bhat, K.M.; Rao, M.S.; Nampurath, G.K.; Chamallamudi, M.R.; Nayak, S.R.; Muttigi, M.S. Petroleum ether extract of Cissus quadrangularis (Linn.) enhances bone marrow mesenchymal stem cell proliferation and facilitates osteoblastogenesis. Clinics 2009, 64, 993–998. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Kim, I.; Park, C.H.; Kim, J.B. Korean mistletoe lectin enhances natural killer cell cytotoxicity via upregulation of perforin expression. Asian Pac. J. Allergy Immunol. 2018, 36, 175–183. [Google Scholar] [CrossRef]
- Han, S.Y.; Hong, C.E.; Kim, H.G.; Lyu, S.Y. Anti-cancer effects of enteric-coated polymers containing mistletoe lectin in murine melanoma cells in vitro and in vivo. Mol. Cell. Biochem. 2015, 408, 73–87. [Google Scholar] [CrossRef]
- Khil, L.Y.; Kim, W.; Lyu, S.; Park, W.B.; Yoon, J.W.; Jun, H.S. Mechanisms involved in Korean mistletoe lectin-induced apoptosis of cancer cells. World J. Gastroenterol. 2007, 13, 2811–2818. [Google Scholar] [CrossRef]
- Kim, B.K.; Choi, M.J.; Park, K.Y.; Cho, E.J. Protective effects of Korean mistletoe lectin on radical-induced oxidative stress. Biol. Pharm. Bull. 2010, 33, 1152–1158. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Hong, J.H.; Kwon, H.J.; Kim, Y.; Lee, K.H.; Kim, J.B.; Song, S.K. TLR4-mediated activation of mouse macrophages by Korean mistletoe lectin-C (KML-C). Biochem. Biophys. Res. Commun. 2010, 396, 721–725. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Kim, J.K.; Kim, H.Y.; Park, S.M.; Lee, S.M. Immunomodulating effects of Korean mistletoe lectin in vitro and in vivo. Int. Immunopharmacol. 2009, 9, 1555–1561. [Google Scholar] [CrossRef] [PubMed]
- Lyu, S.Y.; Choi, J.H.; Lee, H.J.; Park, W.B.; Kim, G.J. Korean mistletoe lectin promotes proliferation and invasion of trophoblast cells through regulation of Akt signaling. Reprod. Toxicol. 2013, 39, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Lyu, S.Y.; Lee, H.J.; Jung, J.; Park, W.B.; Kim, G.J. Korean mistletoe lectin regulates self-renewal of placenta-derived mesenchymal stem cells via autophagic mechanisms. Cell Prolif. 2012, 45, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Zon, L.I. Intrinsic and extrinsic control of haematopoietic stem-cell self-renewal. Nature 2008, 453, 306–313. [Google Scholar] [CrossRef]
- Wilson, A.; Murphy, M.J.; Oskarsson, T.; Kaloulis, K.; Bettess, M.D.; Oser, G.M.; Pasche, A.C.; Knabenhans, C.; Macdonald, H.R.; Trumpp, A. c-Myc controls the balance between hematopoietic stem cell self-renewal and differentiation. Genes Dev. 2004, 18, 2747–2763. [Google Scholar] [CrossRef] [Green Version]
- Boiani, M.; Scholer, H.R. Regulatory networks in embryo-derived pluripotent stem cells. Nat. Rev. Mol. Cell Biol. 2005, 6, 872–884. [Google Scholar] [CrossRef]
- Boyer, L.A.; Lee, T.I.; Cole, M.F.; Johnstone, S.E.; Levine, S.S.; Zucker, J.P.; Guenther, M.G.; Kumar, R.M.; Murray, H.L.; Jenner, R.G.; et al. Core transcriptional regulatory circuitry in human embryonic stem cells. Cell 2005, 122, 947–956. [Google Scholar] [CrossRef]
- Fan, Y.X.; Gu, C.H.; Zhang, Y.L.; Zhong, B.S.; Wang, L.Z.; Zhou, Z.R.; Wang, Z.Y.; Jia, R.X.; Wang, F. Oct4 and Sox2 overexpression improves the proliferation and differentiation of bone mesenchymal stem cells in Xiaomeishan porcine. Genet. Mol. Res. 2013, 12, 6067–6079. [Google Scholar] [CrossRef]
- Lee, J.; Go, Y.; Kang, I.; Han, Y.M.; Kim, J. Oct-4 controls cell-cycle progression of embryonic stem cells. Biochem. J. 2010, 426, 171–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coronado, D.; Godet, M.; Bourillot, P.Y.; Tapponnier, Y.; Bernat, A.; Petit, M.; Afanassieff, M.; Markossian, S.; Malashicheva, A.; Iacone, R.; et al. A short G1 phase is an intrinsic determinant of naive embryonic stem cell pluripotency. Stem Cell Res. 2013, 10, 118–131. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.E.; O’Keefe, R.A.; Grandis, J.R. Targeting the IL-6/JAK/STAT3 signalling axis in cancer. Nat. Rev. Clin. Oncol. 2018, 15, 234–248. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Huang, L.; Zhang, Y.; Qiao, M.; Yuan, Y. SiRNA-mediated PIAS1 silencing promotes inflammatory response and leads to injury of cerulein-stimulated pancreatic acinar cells via regulation of the P38MAPK signaling pathway. Int. J. Mol. Med. 2010, 26, 619–626. [Google Scholar] [CrossRef] [Green Version]
- Wehbe, H.; Henson, R.; Meng, F.; Mize-Berge, J.; Patel, T. Interleukin-6 contributes to growth in cholangiocarcinoma cells by aberrant promoter methylation and gene expression. Cancer Res. 2006, 66, 10517–10524. [Google Scholar] [CrossRef]
- Zhang, X.; Zeng, Y.; Qu, Q.; Zhu, J.; Liu, Z.; Ning, W.; Zeng, H.; Zhang, N.; Du, W.; Chen, C.; et al. PD-L1 induced by IFN-gamma from tumor-associated macrophages via the JAK/STAT3 and PI3K/AKT signaling pathways promoted progression of lung cancer. Int. J. Clin. Oncol. 2017, 22, 1026–1033. [Google Scholar] [CrossRef]
- Gao, S.P.; Mark, K.G.; Leslie, K.; Pao, W.; Motoi, N.; Gerald, W.L.; Travis, W.D.; Bornmann, W.; Veach, D.; Clarkson, B.; et al. Mutations in the EGFR kinase domain mediate STAT3 activation via IL-6 production in human lung adenocarcinomas. J. Clin. Investig. 2007, 117, 3846–3856. [Google Scholar] [CrossRef] [Green Version]
- Yeh, H.H.; Lai, W.W.; Chen, H.H.; Liu, H.S.; Su, W.C. Autocrine IL-6-induced Stat3 activation contributes to the pathogenesis of lung adenocarcinoma and malignant pleural effusion. Oncogene 2006, 25, 4300–4309. [Google Scholar] [CrossRef] [Green Version]
- Meng, F.; Yamagiwa, Y.; Ueno, Y.; Patel, T. Over-expression of interleukin-6 enhances cell survival and transformed cell growth in human malignant cholangiocytes. J. Hepatol. 2006, 44, 1055–1065. [Google Scholar] [CrossRef] [Green Version]
- Okada, K.; Shimizu, Y.; Nambu, S.; Higuchi, K.; Watanabe, A. Interleukin-6 functions as an autocrine growth factor in a cholangiocarcinoma cell line. J. Gastroenterol. Hepatol. 1994, 9, 462–467. [Google Scholar] [CrossRef]
- Dandrea, M.; Donadelli, M.; Costanzo, C.; Scarpa, A.; Palmieri, M. MeCP2/H3meK9 are involved in IL-6 gene silencing in pancreatic adenocarcinoma cell lines. Nucleic Acids Res. 2009, 37, 6681–6690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armenante, F.; Merola, M.; Furia, A.; Palmieri, M. Repression of the IL-6 gene is associated with hypermethylation. Biochem. Biophys. Res. Commun. 1999, 258, 644–647. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.; Chun, E.; Kim, S.Y.; Kim, M.; Lee, K.Y.; Kim, S.J. Notch-induced hIL-6 production facilitates the maintenance of self-renewal of hCD34+ cord blood cells through the activation of Jak-PI3K-STAT3 pathway. Am. J. Pathol. 2012, 180, 351–364. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, E. Investigation of the proliferation, apoptosis/necrosis, and cell cycle phases in several human multiple myeloma cell lines. Comparison of Viscum album QuFrF extract with vincristine in an in vitro model. Sci. World J. 2010, 10, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.J.; Jung, J.; Na, K.H.; Moon, J.S.; Lee, H.J.; Kim, J.H.; Kim, G.I.; Kwon, S.W.; Hwang, S.G.; Kim, G.J. Anti-fibrotic effect of chorionic plate-derived mesenchymal stem cells isolated from human placenta in a rat model of CCl(4)-injured liver: Potential application to the treatment of hepatic diseases. J. Cell Biochem. 2010, 111, 1453–1463. [Google Scholar] [CrossRef]
- Lyu, S.Y.; Park, S.M.; Choung, B.Y.; Park, W.B. Comparative study of Korean (Viscum album var. coloratum) and European mistletoes (Viscum album). Arch. Pharm. Res. 2000, 23, 592–598. [Google Scholar] [CrossRef]
- Jaenisch, R.; Bird, A. Epigenetic regulation of gene expression: How the genome integrates intrinsic and environmental signals. Nat. Genet. 2003, 33, 245–254. [Google Scholar] [CrossRef]
- Nardelli, C.; Granata, I.; Iaffaldano, L.; D’Argenio, V.; Del Monaco, V.; Maruotti, G.M.; Omodei, D.; Del Vecchio, L.; Martinelli, P.; Salvatore, F.; et al. miR-138/miR-222 Overexpression Characterizes the miRNome of Amniotic Mesenchymal Stem Cells in Obesity. Stem Cells Dev. 2017, 26, 4–14. [Google Scholar] [CrossRef]
- Lyu, S.Y.; Park, W.B. Effects of Korean mistletoe lectin (Viscum album coloratum) on proliferation and cytokine expression in human peripheral blood mononuclear cells and T-lymphocytes. Arch. Pharm. Res. 2007, 30, 1252–1264. [Google Scholar] [CrossRef]
- Lyu, S.Y.; Park, W.B. Mistletoe lectin (Viscum album coloratum) modulates proliferation and cytokine expressions in murine splenocytes. J. Biochem. Mol. Biol. 2006, 39, 662–670. [Google Scholar] [CrossRef]
- Hajto, T.; Hostanska, K.; Frei, K.; Rordorf, C.; Gabius, H.J. Increased secretion of tumor necrosis factors alpha, interleukin 1, and interleukin 6 by human mononuclear cells exposed to beta-galactoside-specific lectin from clinically applied mistletoe extract. Cancer Res. 1990, 50, 3322–3326. [Google Scholar] [PubMed]
- Li, Y.; Wang, L.; Pappan, L.; Galliher-Beckley, A.; Shi, J. IL-1beta promotes stemness and invasiveness of colon cancer cells through Zeb1 activation. Mol. Cancer 2012, 11, 87. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, Z.; Li, Y.; Pappan, L.; Galliher-Beckley, A.; Shi, J. Pro-inflammatory cytokine interleukin-1beta promotes the development of intestinal stem cells. Inflamm. Res. 2012, 61, 1085–1092. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wang, G.Y.; Dong, B.H.; Zhang, Y.C.; Wang, Y.X.; Sun, B.C. Biological characteristics of human placental mesenchymal stem cells and their proliferative response to various cytokines. Cells Tissues Organs 2007, 186, 169–179. [Google Scholar] [CrossRef]
- Pogribny, I.P.; Beland, F.A. DNA methylome alterations in chemical carcinogenesis. Cancer Lett. 2013, 334, 39–45. [Google Scholar] [CrossRef]
- Poplutz, M.K.; Wessels, I.; Rink, L.; Uciechowski, P. Regulation of the Interleukin-6 gene expression during monocytic differentiation of HL-60 cells by chromatin remodeling and methylation. Immunobiology 2014, 219, 619–626. [Google Scholar] [CrossRef]
- Tekpli, X.; Landvik, N.E.; Anmarkud, K.H.; Skaug, V.; Haugen, A.; Zienolddiny, S. DNA methylation at promoter regions of interleukin 1B, interleukin 6, and interleukin 8 in non-small cell lung cancer. Cancer Immunol. Immunother. 2013, 62, 337–345. [Google Scholar] [CrossRef]
- Nile, C.J.; Read, R.C.; Akil, M.; Duff, G.W.; Wilson, A.G. Methylation status of a single CpG site in the IL6 promoter is related to IL6 messenger RNA levels and rheumatoid arthritis. Arthritis Rheum. 2008, 58, 2686–2693. [Google Scholar] [CrossRef]
- Jung, J.; Moon, J.W.; Choi, J.H.; Lee, Y.W.; Park, S.H.; Kim, G.J. Epigenetic Alterations of IL-6/STAT3 Signaling by Placental Stem Cells Promote Hepatic Regeneration in a Rat Model with CCl4-induced Liver Injury. Int. J. Stem Cells 2015, 8, 79–89. [Google Scholar] [CrossRef]
- Burdon, T.; Smith, A.; Savatier, P. Signalling, cell cycle and pluripotency in embryonic stem cells. Trends Cell Biol. 2002, 12, 432–438. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.C.; Lin, J.H.; Hsu, T.W.; Su, K.; Li, A.F.; Hsu, H.S.; Hung, S.C. IL-6 enriched lung cancer stem-like cell population by inhibition of cell cycle regulators via DNMT1 upregulation. Int. J. Cancer 2015, 136, 547–559. [Google Scholar] [CrossRef] [PubMed]
- Won, C.; Lee, C.S.; Lee, J.K.; Kim, T.J.; Lee, K.H.; Yang, Y.M.; Kim, Y.N.; Ye, S.K.; Chung, M.H. CADPE suppresses cyclin D1 expression in hepatocellular carcinoma by blocking IL-6-induced STAT3 activation. Anticancer Res. 2010, 30, 481–488. [Google Scholar] [PubMed]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, G.D.; Choi, J.H.; Lim, S.M.; Jun, J.H.; Moon, J.W.; Kim, G.J. Alterations in IL-6/STAT3 Signaling by Korean Mistletoe Lectin Regulate the Self-Renewal Activity of Placenta-Derived Mesenchymal Stem Cells. Nutrients 2019, 11, 2604. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11112604
Kim GD, Choi JH, Lim SM, Jun JH, Moon JW, Kim GJ. Alterations in IL-6/STAT3 Signaling by Korean Mistletoe Lectin Regulate the Self-Renewal Activity of Placenta-Derived Mesenchymal Stem Cells. Nutrients. 2019; 11(11):2604. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11112604
Chicago/Turabian StyleKim, Gi Dae, Jong Ho Choi, Seung Mook Lim, Ji Hye Jun, Ji Wook Moon, and Gi Jin Kim. 2019. "Alterations in IL-6/STAT3 Signaling by Korean Mistletoe Lectin Regulate the Self-Renewal Activity of Placenta-Derived Mesenchymal Stem Cells" Nutrients 11, no. 11: 2604. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11112604