Vegetarian Diet during Pregnancy Is Not Associated with Poorer Cognitive Performance in Children at Age 6–7 Years

 , , and
, , and
Abstract
:1. Introduction
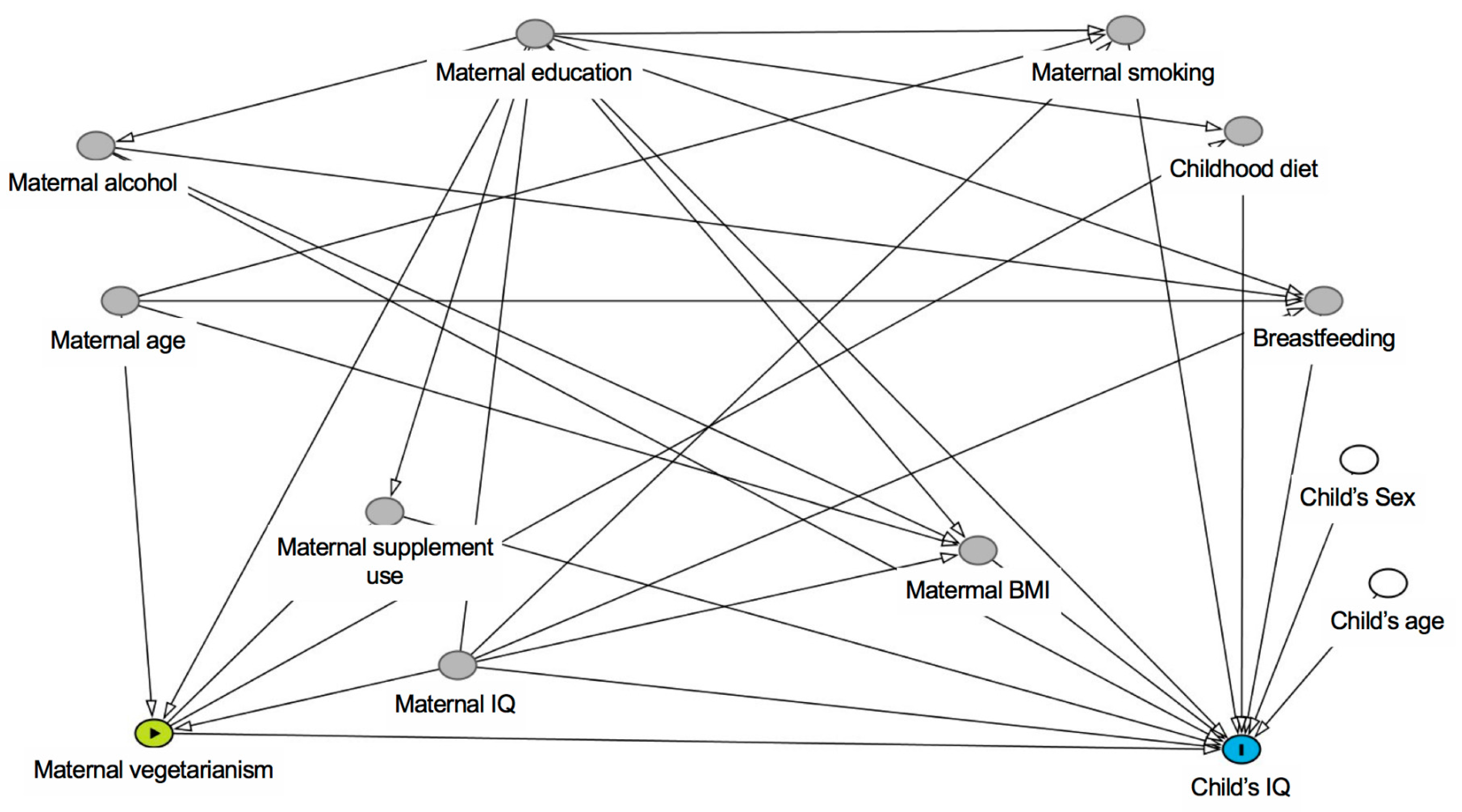
2. Materials and Methods
2.1. Ethical Statement
2.2. Study Sample
2.3. Maternal Blood Sample Collection and Analysis of Nutrient Status
2.4. Assessment of Cognitive Function in Children
2.5. Statistical Analysis
3. Results
3.1. Participant Characteristics
3.2. Serum Phosphatidylcholine Fatty Acid Concentrations in Pregnant Women
3.3. The Concentrations of Micronutrients in Serum from Vegetarian and Omnivorous Women
3.4. The Relationship between Maternal Dietary Choice and Cognitive Function in Their Children
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| ARA | Arachidonic acid |
| BMI | Body-mass-index |
| CANTAB® | Cambridge Neuropsychological Test Automated Battery |
| DAG | Direct acyclic graph |
| DHA | Docosahexaenoic acid |
| DMS | Delayed matching to sample |
| FAME | Fatty acid methyl esters |
| FFQ | Food frequency questionnaire |
| IED | Intra/extra-dimensional shift |
| IST | Information Sampling Task |
| PC | Phosphatidylcholine |
| SSP | Spatial Span |
| WASI | Wechsler Abbreviated Scale of Intelligence |
| WPPSI | Wechsler Preschool and Primary Scale of Intelligence. |
References
- Nyaradi, A.; Li, J.; Hickling, S.; Foster, J.; Oddy, W.H. The role of nutrition in children’s neurocognitive development, from pregnancy through childhood. Front. Hum. Neurosci. 2013, 7, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thatcher, R.W. Maturation of the Human Frontal Lobes—Physiological Evidence for Staging. Dev. Neuropsychol. 1991, 7, 397–419. [Google Scholar] [CrossRef]
- Connor, J.R.; Menzies, S.L. Relationship of iron to oligodendrocytes and myelination. Glia 1996, 17, 83–93. [Google Scholar] [CrossRef]
- Youdim, M.B.; Yehuda, S. The neurochemical basis of cognitive deficits induced by brain iron deficiency: Involvement of dopamine-opiate system. Cell. Mol. Biol. (Noisy-le-grand) 2000, 46, 491–500. [Google Scholar]
- Todorich, B.; Pasquini, J.M.; Garcia, C.I.; Paez, P.M.; Connor, J.R. Oligodendrocytes and Myelination: The Role of Iron. Glia 2009, 57, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, E.; Pasquini, J.M.; Thompson, K.; Felt, B.; Butkus, G.; Beard, J.; Connor, J.R. Effect of manipulation of iron storage, transport, or availability on myelin composition and brain iron content in three different animal models. J. Neurosci. Res. 2004, 77, 681–689. [Google Scholar] [CrossRef]
- McCann, J.C.; Ames, B.N. An overview of evidence for a causal relation between iron deficiency during development and deficits in cognitive or behavioral function. Am. J. Clin. Nutr. 2007, 85, 931–945. [Google Scholar] [CrossRef] [Green Version]
- Grantham-McGregor, S.; Ani, C. A review of studies on the effect of iron deficiency on cognitive development in children. J. Nutr. 2001, 131, 649S–666S. [Google Scholar] [CrossRef]
- Siddappa, A.M.; Georgieff, M.K.; Wewerka, S.; Worwa, C.; Nelson, C.A.; DeRegnier, R.A. Iron deficiency alters auditory recognition memory in newborn infants of diabetic mothers. Ped. Res. 2004, 55, 1034–1041. [Google Scholar] [CrossRef] [Green Version]
- Armony-Sivan, R.; Jacobson, J.L.; Jacobson, S.W.; Molteno, C.D.; Carter, R.C.; Marais, A.S.; Viljoen, D.L. Does iron deficiency anemia mediate the relation of fetal alcohol exposure to postnatal growth? Alcohol. Clin. Exp. Res. 2004, 28, 44A. [Google Scholar]
- Lukowski, A.F.; Koss, M.; Burden, M.J.; Jonides, J.; Nelson, C.A.; Kaciroti, N.; Jimenez, E.; Lozoff, B. Iron deficiency in infancy and neurocognitive functioning at 19 years: Evidence of long-term deficits in executive function and recognition memory. Nutr. Neurosci. 2010, 13, 54–70. [Google Scholar] [CrossRef] [PubMed]
- Lovblad, K.O.; Ramelli, G.; Remonda, L.; Nirkko, A.C.; Ozdoba, C.; Schroth, G. Retardation of myelination due to dietary vitamin B-12 deficiency: Cranial MRI findings. Pediatr. Radiol. 1997, 27, 155–158. [Google Scholar] [CrossRef] [PubMed]
- Wighton, M.C.; Manson, J.I.; Speed, I.; Robertson, E.; Chapman, E. Brain damage in infancy and dietary vitamin B12 deficiency. Med. J. Aust. 1979, 2, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Black, M.M. Effects of vitamin B12 and folate deficiency on brain development in children. Food Nutr. Bull. 2008, 29, S126–S131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heyward, F.D.; Sweatt, J.D. DNA Methylation in Memory Formation: Emerging Insights. Neuroscientist 2015, 21, 475–489. [Google Scholar] [CrossRef] [Green Version]
- Lai, J.S.; Na’im Mohamad Ayob, M.; Cai, S.; Quah, P.L.; Gluckman, P.D.; Shek, L.P.; Yap, F.; Tan, K.H.; Chong, Y.S.; Godfrey, K.M.; et al. Maternal plasma vitamin B12 concentrations during pregnancy and infant cognitive outcomes at 2 years of age. Br. J. Nutr. 2019, 121, 1303–1312. [Google Scholar] [CrossRef] [Green Version]
- Pearce, E.N. Monitoring and effects of iodine deficiency in pregnancy: Still an unsolved problem? Eur. J. Clin. Nutr. 2013, 67, 481–484. [Google Scholar] [CrossRef]
- Korevaar, T.I.M.; Medici, M.; Visser, T.J.; Peeters, R.P. Thyroid disease in pregnancy: New insights in diagnosis and clinical management. Nat. Rev. Endocrinol. 2017, 13, 610–622. [Google Scholar] [CrossRef]
- Robinson, S.M.; Crozier, S.R.; Miles, E.A.; Gale, C.R.; Calder, P.C.; Cooper, C.; Inskip, H.M.; Godfrey, K.M. Preconception Maternal Iodine Status Is Positively Associated with IQ but Not with Measures of Executive Function in Childhood. J. Nutr. 2018, 148, 959–966. [Google Scholar] [CrossRef] [Green Version]
- Kuipers, R.S.; Luxwolda, M.F.; Offringa, P.J.; Boersma, E.R.; Dijck-Brouwer, D.A.; Muskiet, F.A. Fetal intrauterine whole body linoleic, arachidonic and docosahexaenoic acid contents and accretion rates. Prostaglandins Leukot. Essent. Fatty Acids 2012, 86, 13–20. [Google Scholar] [CrossRef]
- Salem, N., Jr.; Pawlosky, R.J. Docosahexaenoic acid is an essential nutrient in the nervous system. J. Nutr. Sci. Vitaminol. 1992, 38, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Neuringer, M.; Reisbick, S.; Janowsky, J. The role of n-3 fatty acids in visual and cognitive development: Current evidence and methods of assessment. J. Pediatr. 1994, 125, S39–S47. [Google Scholar] [CrossRef]
- Reisbick, S.; Neuringer, M.; Hasnain, R.; Connor, W.E. Home cage behavior of rhesus monkeys with long-term deficiency of omega-3 fatty acids. Physiol. Behav. 1994, 55, 231–239. [Google Scholar] [CrossRef]
- Ghys, A.; Bakker, E.; Hornstra, G.; van den Hout, M. Red blood cell and plasma phospholipid arachidonic and docosahexaenoic acid levels at birth and cognitive development at 4 years of age. Early Hum. Dev. 2002, 69, 83–90. [Google Scholar] [CrossRef]
- Bakker, E.C.; Ghys, A.J.; Kester, A.D.; Vles, J.S.; Dubas, J.S.; Blanco, C.E.; Hornstra, G. Long-chain polyunsaturated fatty acids at birth and cognitive function at 7 y of age. Eur. J. Clin. Nutr. 2003, 57, 89–95. [Google Scholar] [CrossRef] [Green Version]
- Crozier, S.R.; Sibbons, C.M.; Fisk, H.L.; Godfrey, K.M.; Calder, P.C.; Gale, C.R.; Robinson, S.M.; Inskip, H.M.; Baird, J.; Harvey, N.C.; et al. Arachidonic acid and DHA status in pregnant women is not associated with cognitive performance of their children at 4 or 6–7 years. Br. J. Nutr. 2018, 119, 1400–1407. [Google Scholar] [CrossRef] [Green Version]
- Sebastiani, G.; Herranz Barbero, A.; Borras-Novell, C.; Alsina Casanova, M.; Aldecoa-Bilbao, V.; Andreu-Fernandez, V.; Pascual Tutusaus, M.; Ferrero Martinez, S.; Gomez Roig, M.D.; Garcia-Algar, O. The Effects of Vegetarian and Vegan Diet during Pregnancy on the Health of Mothers and Offspring. Nutrients 2019, 11, 557. [Google Scholar] [CrossRef] [Green Version]
- Pawlak, R.; Lester, S.E.; Babatunde, T. The prevalence of cobalamin deficiency among vegetarians assessed by serum vitamin B12: A review of literature. Eur. J. Clin. Nutr. 2014, 68, 541–548. [Google Scholar] [CrossRef] [Green Version]
- Dagnelie, P.C.; van Staveren, W.A.; Vergote, F.J.; Dingjan, P.G.; van den Berg, H.; Hautvast, J.G. Increased risk of vitamin B-12 and iron deficiency in infants on macrobiotic diets. Am. J. Clin. Nutr. 1989, 50, 818–824. [Google Scholar] [CrossRef]
- Haider, L.M.; Schwingshackl, L.; Hoffmann, G.; Ekmekcioglu, C. The effect of vegetarian diets on iron status in adults: A systematic review and meta-analysis. Crit. Rev. Food Sci. Nutr. 2018, 58, 1359–1374. [Google Scholar] [CrossRef]
- Alwan, N.A.; Greenwood, D.C.; Simpson, N.A.; McArdle, H.J.; Godfrey, K.M.; Cade, J.E. Dietary iron intake during early pregnancy and birth outcomes in a cohort of British women. Hum. Reprod. 2011, 26, 911–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawlak, R.; Bell, K. Iron Status of Vegetarian Children: A Review of Literature. Ann. Nutr. Metab. 2017, 70, 88–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burdge, G.C.; Tan, S.Y.; Henry, C.J. Long-chain n-3 PUFA in vegetarian women: A metabolic perspective. J. Nutr. Sci. 2017, 6, e58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanders, T.A.; Ellis, F.R.; Dickerson, J.W. Studies of vegans: The fatty acid composition of plasma choline phosphoglycerides, erythrocytes, adipose tissue, and breast milk, and some indicators of susceptibility to ischemic heart disease in vegans and omnivore controls. Am. J. Clin. Nutr. 1978, 31, 805–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanders, T.A.; Reddy, S. The influence of a vegetarian diet on the fatty acid composition of human milk and the essential fatty acid status of the infant. J. Pediatr. 1992, 120, S71–S77. [Google Scholar] [CrossRef]
- Kornsteiner, M.; Singer, I.; Elmadfa, I. Very low n-3 long-chain polyunsaturated fatty acid status in Austrian vegetarians and vegans. Ann. Nutr. Metab. 2008, 52, 37–47. [Google Scholar] [CrossRef]
- Phinney, S.D.; Odin, R.S.; Johnson, S.B.; Holman, R.T. Reduced arachidonate in serum phospholipids and cholesteryl esters associated with vegetarian diets in humans. Am. J. Clin. Nutr. 1990, 51, 385–392. [Google Scholar] [CrossRef]
- Rosell, M.S.; Lloyd-Wright, Z.; Appleby, P.N.; Sanders, T.A.; Allen, N.E.; Key, T.J. Long-chain n-3 polyunsaturated fatty acids in plasma in British meat-eating, vegetarian, and vegan men. Am. J. Clin. Nutr. 2005, 82, 327–334. [Google Scholar] [CrossRef]
- Lakin, V.; Haggarty, P.; Abramovich, D.R.; Ashton, J.; Moffat, C.F.; McNeill, G.; Danielian, P.J.; Grubb, D. Dietary intake and tissue concentration of fatty acids in omnivore, vegetarian and diabetic pregnancy. Prostaglandins Leukot. Essent. Fatty Acids 1998, 59, 209–220. [Google Scholar] [CrossRef]
- Inskip, H.M.; Godfrey, K.M.; Robinson, S.M.; Law, C.M.; Barker, D.J.; Cooper, C.; Group, S.W.S.S. Cohort profile: The Southampton Women’s Survey. Int. J. Epidemiol. 2006, 35, 42–48. [Google Scholar] [CrossRef] [Green Version]
- Robinson, S.; Godfrey, K.; Osmond, C.; Cox, V.; Barker, D. Evaluation of a food frequency questionnaire used to assess nutrient intakes in pregnant women. Eur. J. Clin. Nutr. 1996, 50, 302–308. [Google Scholar]
- Davey, G.K.; Spencer, E.A.; Appleby, P.N.; Allen, N.E.; Knox, K.H.; Key, T.J. EPIC-Oxford: Lifestyle characteristics and nutrient intakes in a cohort of 33,883 meat-eaters and 31,546 non meat-eaters in the UK. Public Health Nutr. 2003, 6, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Burdge, G.C.; Wright, P.; Jones, A.E.; Wootton, S.A. A method for separation of phosphatidylcholine, triacylglycerol, non-esterified fatty acids and cholesterol esters from plasma by solid-phase extraction. Br. J. Nutr. 2000, 84, 781–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folch, J.; Lees, M.; Sloane-Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Caesar, P.A.; Wilson, S.J.; Normand, I.C.S.; Postle, A.D. A Comparison of the Specificity of Phosphatidylcholine Synthesis by Human-Fetal Lung Maintained in Either Organ or Organotypic Culture. Biochem. J. 1988, 253, 451–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thurnham, D.I.; Smith, E.; Flora, P.S. Concurrent liquid-chromatographic assay of retinol, alpha-tocopherol, beta-carotene, alpha-carotene, lycopene, and beta-cryptoxanthin in plasma, with tocopherol acetate as internal standard. Clin. Chem. 1988, 34, 377–381. [Google Scholar] [PubMed]
- Adaikalakoteswari, A.; Webster, C.; Goljan, I.; Saravanan, P. Simultaneous detection of five one-carbon metabolites in plasma using stable isotope dilution liquid chromatography tandem mass spectrometry. J. Chromatogr. B. 2016, 1012, 186–192. [Google Scholar] [CrossRef]
- Gale, C.R.; Martyn, C.N.; Marriott, L.D.; Limond, J.; Crozier, S.; Inskip, H.M.; Godfrey, K.M.; Law, C.M.; Cooper, C.; Robinson, S.M.; et al. Dietary patterns in infancy and cognitive and neuropsychological function in childhood. J. Child Psychol. Psychiatry 2009, 50, 816–823. [Google Scholar] [CrossRef] [Green Version]
- Lillycrop, K.A.; Costello, P.M.; Teh, A.L.; Murray, R.J.; Clarke-Harris, R.; Barton, S.J.; Garratt, E.S.; Ngo, S.; Sheppard, A.M.; Wong, J.; et al. Association between perinatal methylation of the neuronal differentiation regulator HES1 and later childhood neurocognitive function and behaviour. Int. J. Epidemiol. 2015, 44, 1263–1276. [Google Scholar] [CrossRef] [Green Version]
- Robbins, T.W.; James, M.; Owen, A.M.; Sahakian, B.J.; Lawrence, A.D.; McInnes, L.; Rabbitt, P.M. A study of performance on tests from the CANTAB battery sensitive to frontal lobe dysfunction in a large sample of normal volunteers: Implications for theories of executive functioning and cognitive aging. Cambridge Neuropsychological Test Automated Battery. J. Int. Neuropsychol. Soc. 1998, 4, 474–490. [Google Scholar]
- Armitage, P.B.B.; Berry, G. Statistical Methods in Medical Research, 3rd ed.; Blackwell: Oxford, UK, 1994. [Google Scholar]
- Greenland, S.; Pearl, J.; Robins, J.M. Causal diagrams for epidemiologic research. Epidemiology 1999, 10, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Abbassi-Ghanavati, M.; Greer, L.G.; Cunningham, F.G. Pregnancy and laboratory studies: A reference table for clinicians. Obstet. Gynecol. 2009, 114, 1326–1331. [Google Scholar] [CrossRef] [PubMed]
- Northstone, K.; Emmett, P.; Rogers, I. Dietary patterns in pregnancy and associations with socio-demographic and lifestyle factors. Eur. J. Clin. Nutr. 2008, 62, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Koebnick, C.; Heins, U. Vitamin B-12 during pregnancy. Ernahr. Umsch. 2001, 48, 92. [Google Scholar]
- Koebnick, C.; Heins, U.A.; Hoffmann, I.; Dagnelie, P.C.; Leitzmann, C. Folate status during pregnancy in women is improved by long-term high vegetable intake compared with the average Western diet. J. Nutr. 2001, 131, 733–739. [Google Scholar] [CrossRef] [Green Version]
- Varela-Moreiras, G.; Murphy, M.M.; Scott, J.M. Cobalamin, folic acid, and homocysteine. Nutr. Rev. 2009, 67, S69–S72. [Google Scholar] [CrossRef]
- Sable, P.; Randhir, K.; Kale, A.; Chavan-Gautam, P.; Joshi, S. Maternal micronutrients and brain global methylation patterns in the offspring. Nutr. Neurosci. 2015, 18, 30–36. [Google Scholar] [CrossRef]
- Sprecher, H.; Chen, Q.; Yin, F.Q. Regulation of the biosynthesis of 22:5n-6 and 22:6n-3: A complex intracellular process. Lipids 1999, 34, S153–S156. [Google Scholar] [CrossRef]
- Moriguchi, T.; Greiner, R.S.; Salem, N. Behavioral deficits associated with dietary induction of decreased brain docosahexaenoic acid concentration. J. Neurochem. 2000, 75, 2563–2573. [Google Scholar] [CrossRef]


{kind=link}
| Vegetarian | Omnivorous | p | |||
|---|---|---|---|---|---|
| n | Statistic | n | Statistic | ||
| Mother | Early pregnancy | ||||
| Age at child’s birth (y), mean (SD) | 78 | 30.4 (3.4) | 2142 | 30.5 (3.7) | 0.85 |
| IQ (measured when child 6–7 years old), mean (SD) | 18 | 113.7 (12.1) | 530 | 107.5 (12.7) | 0.04 |
| Educational attainment; qualifications ≥ A-level, n (%) | 78 | 61 (78.2) | 2138 | 1266 (59.2) | 0.001 |
| Smoked in pregnancy, n (%) | 78 | 5 (6.4) | 2114 | 335 (15.8) | 0.02 |
| BMI (kg/m2), median (IQR) | 78 | 23.9 (22.1, 25.4) | 2128 | 24.3 (21.9, 27.5) | 0.24 |
| Multiparous, n (%) | 78 | 35 (44.9) | 2142 | 1047 (48.9) | 0.49 |
| Duration breastfeeding (weeks), median (IQR) | 71 | 29.9 (23.5) | 1987 | 17.1 (21.1) | <0.001 |
| Supplement use, n (%) | 78 | 76 (97.4) | 2143 | 2047 (95.5) | 0.58 |
| Child | |||||
| Female, n (%) | 16 | 6 (37.5) | 479 | 237 (49.5) | 0.35 |
| Gestational age at birth (weeks), median (IQR) | 16 | 40.8 (40.1, 41.4) | 479 | 40.1 (39.0, 41.1) | 0.04 |
| BMI at 6–7 y (kg/m2), median (IQR) | 16 | 15.9 (14.7, 16.6) | 470 | 15.8 (14.9, 16.8) | 0.88 |
| Age at 6–7 y follow-up (y), mean (SD) | 16 | 7.1 (0.3) | 479 | 7.0 (0.2) | 0.12 |
| Mother | Late pregnancy | ||||
| Age at child’s birth (y), mean (SD) | 91 | 31.3 (3.4) | 2551 | 30.5 (3.8) | 0.04 |
| IQ (measured when child 6–7 years old), mean (SD) | 23 | 112.0 (11.6) | 583 | 107.4 (12.9) | 0.10 |
| Educational attainment; qualifications ≥ A-level, n (%) | 91 | 71 (78.0) | 2546 | 1464 (57.5) | <0.001 |
| Smoked in pregnancy, n (%) | 91 | 6 (6.6) | 2544 | 443 (17.4) | 0.007 |
| BMI (kg/m2), median (IQR) | 91 | 23.9 (22.1, 25.7) | 2531 | 24.2 (21.9, 27.5) | 0.50 |
| Multiparous, n (%) | 91 | 48 (52.7) | 2551 | 1326 (52.0) | 0.89 |
| Duration breastfeeding (weeks), median (IQR) | 85 | 31.3 (25.8) | 2373 | 17.3 (21.6) | <0.001 |
| Supplement use | 91 | 63 (69.2) | 2552 | 1226 (48.0) | <0.001 |
| Child | |||||
| Female, n (%) | 22 | 6 (27.3) | 524 | 255 (48.7) | 0.05 |
| Gestational age at birth (weeks), median (IQR) | 22 | 40.6 (40.0, 41.2) | 524 | 40.1 (39.2, 41.2) | 0.08 |
| BMI at 6–7 y (kg/m2), median (IQR) | 22 | 15.3 (14.7, 15.9) | 510 | 15.8 (14.9, 16.9) | 0.24 |
| Age at 6–7 y follow-up (y), mean (SD) | 22 | 7.1 (0.2) | 524 | 7.0 (0.2) | 0.10 |
| Fatty Acid Concentration (μg/mL) | ||||||
|---|---|---|---|---|---|---|
| Early Pregnancy | Late Pregnancy | |||||
| Vegetarians (n = 32) | Omnivores (n = 967) | p | Vegetarians (n = 59) | Omnivores (n = 1703) | p | |
| 14:0 | 4.8 (3.7, 7.5) | 6.0 (4.6, 7.8) | 0.07 | 3.2 (2.0, 4.9) | 3.2 (2.2, 4.5) | 0.40 |
| 16:0 | 489.5 (420.9, 594.0) | 536.8 (458.9, 639.0) | 0.06 | 462.5 (381.5, 569.4) | 514.8 (399.6, 631.8) | 0.04 |
| 16:1n-7 | 7.1 (5.6, 11.6) | 9.3 (6.9, 12.8) | 0.05 | 5.8 (3.3, 8.9) | 5.2 (3.3, 8.4) | 0.63 |
| 18:0 | 185.5 (161.1, 219.9) | 204.6 (171.9, 238.2) | 0.02 | 136.4 (97.5, 169.2) | 143.9 (111.2, 178.8) | 0.07 |
| 18:1n-9 | 158.9 (132.6, 194.1) | 177.9 (148.9, 212.6) | 0.08 | 148.9 (110.4, 192.1) | 170.2 (130.3, 210.3) | 0.01 |
| 18:1n-7 | 28.8 (24.5, 31.4) | 29.9 (24.2, 35.8) | 0.19 | 17.7 (12.7, 21.9) | 20.3 (13.9, 26.0) | 0.007 |
| 18:2n-6 | 367.2 (317.2, 428.9) | 368.5 (312.0, 435.5) | 0.97 | 328.3 (248.2, 387.0) | 333.6 (258.7, 408.3) | 0.60 |
| 18:3n-6 | 1.4 (1.0, 2.4) | 1.7 (1.3, 2.4) | 0.07 | 0.0 (0.0, 0.0) † | 0.0 (0.0, 0.0) † | 0.19 |
| 18:3n-3 | 5.9 (4.0, 8.9) | 6.1 (4.5, 8.1) | 0.53 | 3.7 (2.7, 5.2) | 4.0 (3.0, 5.6) | 0.21 |
| 20:0 | 3.9 (2.5, 5.6) | 4.2 (3.1, 5.6) | 0.30 | 2.1 (0.0, 3.1) | 2.3 (0.0, 3.2) | 0.36 |
| 20:1n-9 | 3.7 (3.1, 5.0) | 3.8 (3.0, 4.8) | 0.91 | 1.4 (0.0, 2.3) | 2.0 (0.0, 2.7) | 0.007 |
| 20:2n-6 | 7.6 (5.3, 9.3) | 8.0 (6.6, 9.8) | 0.30 | 4.9 (3.7, 6.1) | 5.1 (3.9, 6.5) | 0.26 |
| 20:3n-6 | 58.2 (49.7, 79.2) | 67.7 (53.7, 83.9) | 0.13 | 52.4 (38.2, 64.8) | 53.6 (39.6, 68.6) | 0.26 |
| 20:4n-6 | 143.9 (134.8, 170.1) | 165.8 (135.5, 200.8) | 0.02 | 100.9 (72.9, 122.7) | 108.4 (83.8, 139.0) | 0.03 |
| 22:0 | 2.1 (0.9, 2.9) | 1.8 (1.1, 2.7) | 0.60 | 0.0 (0.0, 0.0) † | 0.0 (0.0, 0.0) † | 0.47 |
| 20:4n-3 | 3.1 (2.3, 5.5) | 4.2 (2.9, 5.9) | 0.04 | 0.0 (0.0, 2.3) | 1.8 (0.0, 2.9) | 0.02 |
| 20:5n-3 | 8.4 (6.1, 10.4) | 14.0 (10.4, 18.8) | <0.001 | 3.5 (2.0, 4.6) | 5.3 (3.7, 7.8) | <0.001 |
| 22:4n-6 | 7.4 (5.0, 9.2) | 6.9 (5.2, 8.6) | 0.51 | 3.4 (2.4, 4.4) | 3.3 (2.4, 4.3) | 0.74 |
| 22:5n-6 | 7.1 (6.0, 10.4) | 6.2 (4.5, 8.3) | 0.01 | 5.2 (3.9, 7.3) | 4.3 (3.2, 6.0) | 0.001 |
| 24:1n-9 | 4.2 (1.8, 5.3) | 3.2 (2.1, 5.1) | 0.63 | 0.0 (0.0, 0.0) † | 0.0 (0.0, 0.0) † | 0.18 |
| 22:5n-3 | 13.3 (10.6, 16.9) | 15.2 (12.3, 19.6) | 0.01 | 5.1 (3.6, 6.5) | 6.6 (4.8, 8.8) | <0.001 |
| 22:6n-3 | 57.8 (50.2, 62.9) | 82.6 (66.4, 102.7) | <0.001 | 38.0 (27.9, 48.9) | 54.3 (41.2, 72.3) | <0.001 |
| Early Pregnancy | Late Pregnancy | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| n | Vegetarians | n | Omnivores | p | n | Vegetarians | n | Omnivores | p | |
| β-carotene (μmol/L) | 29 | 0.4 (0.3, 0.7) | 876 | 0.4 (0.3, 0.6) | 0.52 | 48 | 0.3 (0.3, 0.5) | 1299 | 0.3 (0.2, 0.5) | 0.15 |
| Cobalamin (pg/mL) | 71 | 292.6 (211.3, 358.8) | 1910 | 368.7 (279.6, 464.1) | <0.001 | 78 | 131.5 (99.0, 182.0) | 1982 | 162.0 (128.0, 209.0) | <0.001 |
| Serum folate (μg/L) | 71 | ND | ND | 78 | 13.2 (8.9, 21.8) | 1940 | 9.9 (6.5, 17.4) | 0.001 | ||
| Erythrocyte folate (nmol/L) | 61 | 1190 (891, 1400) | 1624 | 1061 (802, 1381) | 0.06 | ND | ND | |||
| Homocysteine (μmol/L) | 71 | 13.3 (8.7, 19.8) | 1904 | 12.4 (7.8, 20.3) | 0.50 | 14 | 5.9 (5.3, 7.4) | 481 | 6.1 (5.2, 7.4) | 0.63 |
| Riboflavin (nmol/L) | ND | ND | 14 | 14.9 (10.8, 27.6) | 481 | 17.1 (11.9, 25.3) | 0.74 | |||
| Nicotinamide (nmol/L) | ND | ND | 14 | 118.9 (36.6, 209.9) | 481 | 140.2 (82.6, 213.6) | 0.24 | |||
| Haemoglobin (g/L) | 69 | 124.0 (119.0, 131.0) | 1851 | 127.0 (121.0, 133.0) | 0.09 | 87 | 112.0 (105.0, 119.0) | 2192 | 115.0 (109.0, 122.0) | <0.001 |
| Ferritin (ng/mL) | ND | ND | 15 | 46.8 (28.3, 78.1) | 484 | 48.7 (32.6, 75.4) | 0.58 | |||
| Unadjusted | Adjusted# | |||||
|---|---|---|---|---|---|---|
| n | β (95% CI) | p | n | β (95% CI) | p | |
| IQ (WASI) | ||||||
| Early pregnancy vegetarian, Y/N | 564 | 7.07 (−0.42, 14.55) | 0.06 | 537 | 3.41 (−3.46, 10.29) | 0.33 |
| Late pregnancy vegetarian, Y/N | 619 | 4.01 (−2.60, 10.62) | 0.23 | 594 | 1.34 (−4.75, 7.43) | 0.66 |
| CANTAB® DMS total correct (12 sec delay) | ||||||
| Early pregnancy vegetarian, Y/N | 494 | 0.53 (−0.07, 1.12) | 0.08 | 479 | 0.54 (−0.07, 1.14) | 0.08 |
| Late pregnancy vegetarian, Y/N | 546 | 0.38 (−0.13, 0.88) | 0.15 | 532 | 0.41 (−0.11, 0.93) | 0.12 |
| CANTAB® IED pre-ED errors (z-score) | ||||||
| Early pregnancy vegetarian, Y/N | 495 | 0.08 (−0.39, 0.56) | 0.73 | 480 | 0.11 (−0.37, 0.59) | 0.66 |
| Late pregnancy vegetarian, Y/N | 545 | −0.05 (−0.46, 0.36) | 0.80 | 531 | −0.06 (−0.47, 0.36) | 0.79 |
| CANTAB® IED EDS errors | ||||||
| Early pregnancy vegetarian, Y/N | 495 | −0.51 (−1.00, −0.03) | 0.04 | 480 | −0.48 (−0.96, −0.00) | 0.05 |
| Late pregnancy vegetarian, Y/N | 545 | −0.39 (−0.81, 0.02) | 0.07 | 531 | −0.33 (−0.75, 0.09) | 0.12 |
| CANTAB® IED total errors (stage 1) in 5 groups | ||||||
| Early pregnancy vegetarian, Y/N | 492 | −0.39 (−0.95, 0.18) | 0.18 | 477 | −0.37 (−0.94, 0.19) | 0.20 |
| Late pregnancy vegetarian, Y/N | 540 | −0.21 (−0.70, 0.28) | 0.41 | 526 | −0.15 (−0.64, 0.35) | 0.56 |
| CANTAB® IED total errors (stage 8) in 5 groups | ||||||
| Early pregnancy vegetarian, Y/N | 492 | −0.73 (−1.43, −0.03) | 0.04 | 477 | −0.65 (−1.34, 0.04) | 0.07 |
| Late pregnancy vegetarian, Y/N | 540 | −0.61 (−1.20, −0.01) | 0.05 | 526 | −0.49 (−1.08, 0.11) | 0.11 |
| CANTAB® IED total errors (adjusted) | ||||||
| Early pregnancy vegetarian, Y/N | 495 | −0.58 (−1.06, −0.11) | 0.02 | 480 | −0.48 (−0.94, −0.01) | 0.04 |
| Late pregnancy vegetarian, Y/N | 545 | −0.46 (−0.88, −0.04) | 0.03 | 531 | −0.36 (−0.79, 0.06) | 0.09 |
| CANTAB® IED stages completed in 4 groups | ||||||
| Early pregnancy vegetarian, Y/N | 495 | 0.53 (0.08, 0.97) | 0.02 | 480 | 0.43 (−0.00, 0.86) | 0.05 |
| Late pregnancy vegetarian, Y/N | 545 | 0.46 (0.08, 0.84) | 0.02 | 531 | 0.34 (−0.04, 0.72) | 0.08 |
| CANTAB® SSP span length | ||||||
| Early pregnancy vegetarian, Y/N | 468 | −0.05 (−0.50, 0.40) | 0.83 | 453 | −0.12 (−0.56, 0.31) | 0.58 |
| Late pregnancy vegetarian, Y/N | 521 | −0.09 (−0.47, 0.28) | 0.63 | 507 | −0.19 (−0.56, 0.18) | 0.32 |
| CANTAB® IST mean prob. correct (win condition fixed) in 5 groups | ||||||
| Early pregnancy vegetarian, Y/N | 452 | 0.56 (−0.18, 1.30) | 0.14 | 438 | 0.52 (−0.22, 1.27) | 0.17 |
| Late pregnancy vegetarian, Y/N | 486 | 0.24 (−0.39, 0.86) | 0.46 | 473 | 0.20 (−0.43, 0.83) | 0.53 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crozier, S.R.; Godfrey, K.M.; Calder, P.C.; Robinson, S.M.; Inskip, H.M.; Baird, J.; Gale, C.R.; Cooper, C.; Sibbons, C.M.; Fisk, H.L.; et al. Vegetarian Diet during Pregnancy Is Not Associated with Poorer Cognitive Performance in Children at Age 6–7 Years. Nutrients 2019, 11, 3029. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11123029
Crozier SR, Godfrey KM, Calder PC, Robinson SM, Inskip HM, Baird J, Gale CR, Cooper C, Sibbons CM, Fisk HL, et al. Vegetarian Diet during Pregnancy Is Not Associated with Poorer Cognitive Performance in Children at Age 6–7 Years. Nutrients. 2019; 11(12):3029. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11123029
Chicago/Turabian StyleCrozier, Sarah R., Keith M. Godfrey, Philip C. Calder, Sian M. Robinson, Hazel M. Inskip, Janis Baird, Catharine R. Gale, Cyrus Cooper, Charlene M. Sibbons, Helena L. Fisk, and et al. 2019. "Vegetarian Diet during Pregnancy Is Not Associated with Poorer Cognitive Performance in Children at Age 6–7 Years" Nutrients 11, no. 12: 3029. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11123029