Effect of Rosmarinic Acid on the Serum Parameters of Glucose and Lipid Metabolism and Oxidative Stress in Estrogen-Deficient Rats


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Determination of Serum Concentrations of Estradiol and Progesterone
2.3. Determination of Serum Concentrations of Glucose, Insulin and Fructosamine, and HOMA-IR Index
2.4. Determination of Serum Concentrations of the Total Cholesterol, Low-Density Lipoprotein Cholesterol, High-Density Lipoprotein Cholesterol and Triglycerides
2.5. Determination of Serum Concentrations of Reduced and Oxidized Glutathione, and Total Antioxidant Capacity
2.6. Determination of Serum Activities of Superoxide Dismutase and Catalase
2.7. Determination of Serum Concentrations of Thiobarbituric Acid Reactive Substances, Advanced Products of Protein Oxidation and Protein Carbonyl Groups
2.8. Determination of Serum Concentrations of Interleukin 18, Uric Acid, Urea and Creatinine, and Activities of Aspartate Aminotransferase and Alanine Aminotransferase
2.9. Statistical Analysis
3. Results
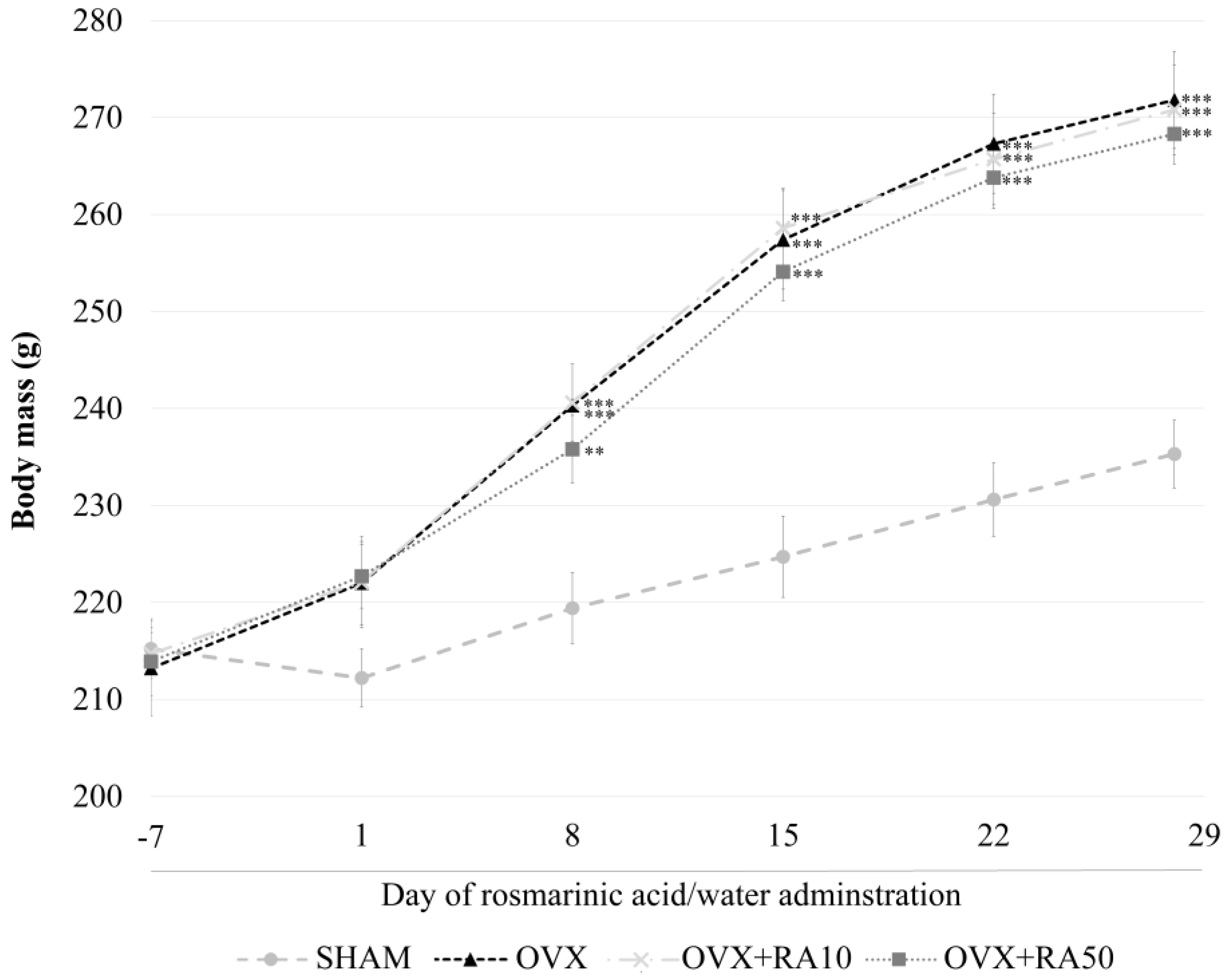
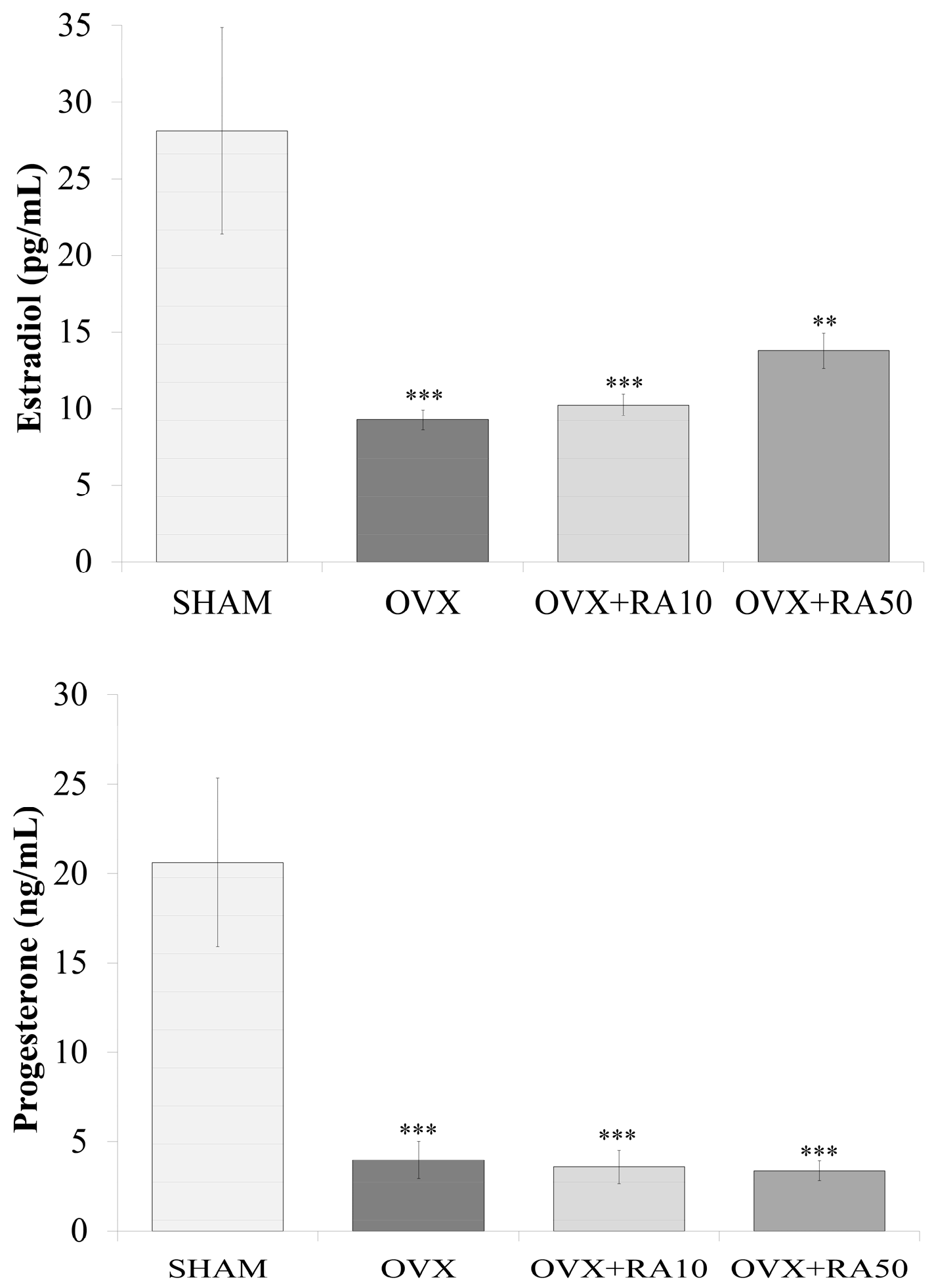
3.1. Effect of Rosmarinic Acid on the Body Mass, Body Mass Gain, Mass of Selected Organs and Serum Concentrations of Sex Hormones
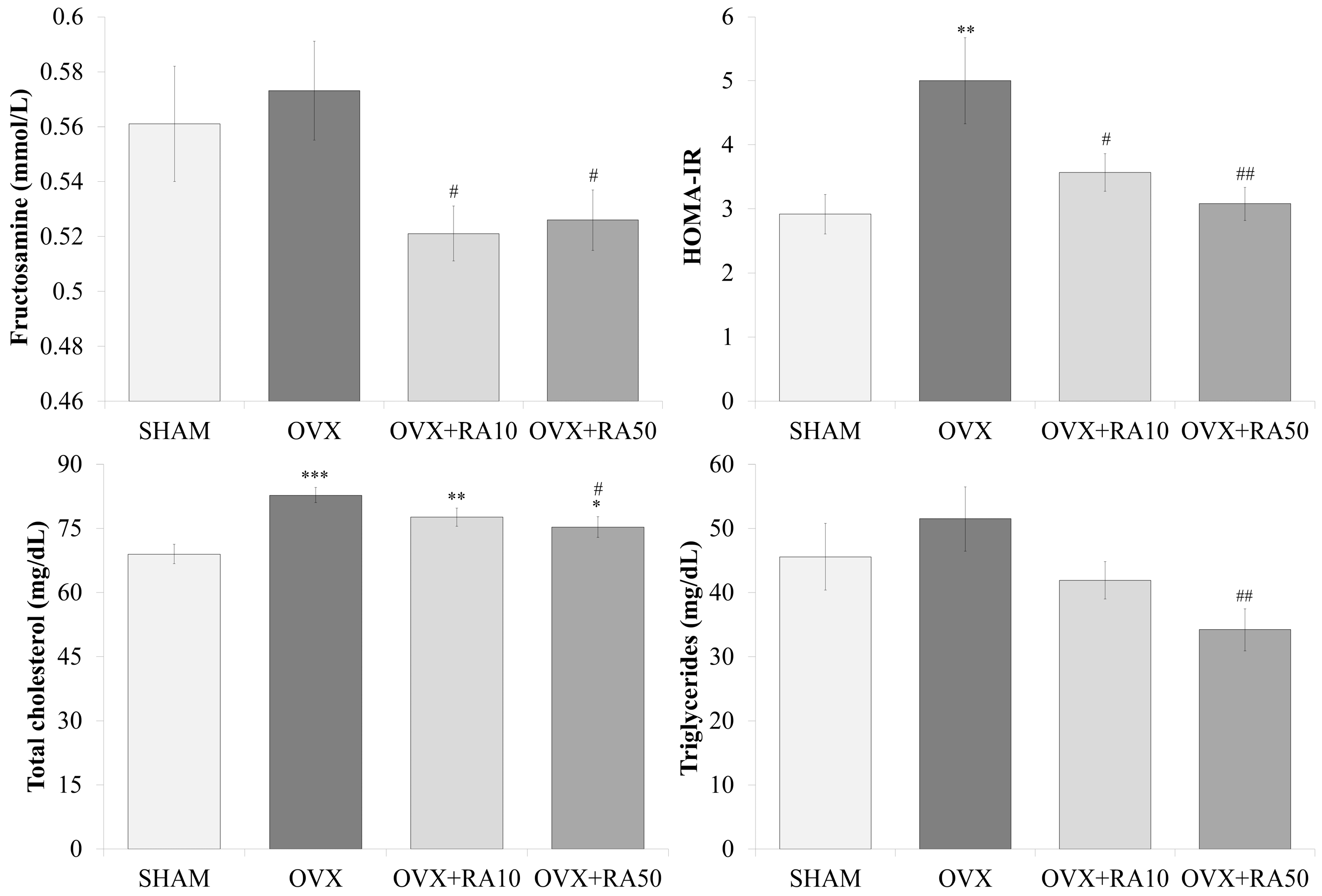
3.2. Effect of Rosmarinic Acid on the Serum Concentrations of Glucose, Insulin and Fructosamine, and the HOMA-IR Index
3.3. Effect of Rosmarinic Acid on the Serum Concentrations of Total Cholesterol, LDL-C, HDL-C and Triglycerides
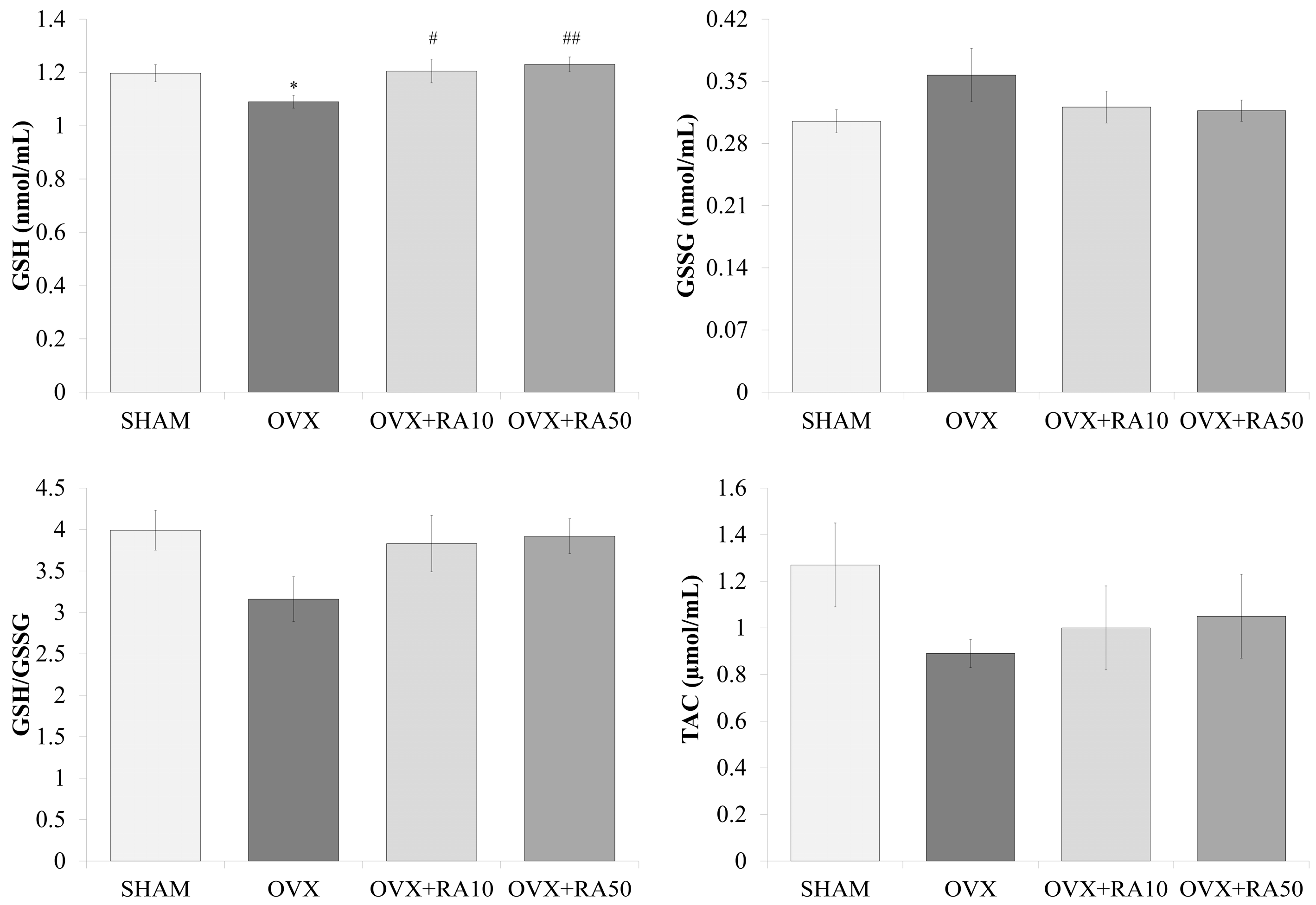
3.4. Effect of Rosmarinic Acid on the Serum Concentrations of GSH, GSSG and TAC
3.5. The Effect of Rosmarinic Acid on the Serum Activities of SOD and CAT, and Concentrations of TBARS, AOPP and PCG
3.6. Effect of Rosmarinic Acid on the Serum Concentrations of Interleukin 18, Uric Acid, Urea and Creatinine, and the Activities of AST and ALT
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Coyoy, A.; Guerra-Araiza, C.; Camacho-Arroyo, I. Metabolism regulation by estrogens and their receptors in the central nervous system before and after menopause. Horm. Metab. Res. 2016, 48, 489–496. [Google Scholar] [CrossRef]
- Sharma, G.; Mauvais-Jarvis, F.; Prossnitz, E.R. Roles of G protein-coupled estrogen receptor GPER in metabolic regulation. J. Steroid Biochem. Mol. Biol. 2018, 176, 31–37. [Google Scholar] [CrossRef]
- Townsend, N.; Nichols, M.; Scarborough, P.; Rayner, M. Cardiovascular disease in Europe—Epidemiological update 2015. Eur. Heart J. 2015, 36, 2696–2705. [Google Scholar] [CrossRef] [PubMed]
- Haarbo, J.; Marslew, U.; Gotfredsen, A.; Christiansen, C. Postmenopausal hormone replacement therapy prevents central distribution of body fat after menopause. Metabolism 1991, 40, 1323–1326. [Google Scholar] [CrossRef]
- Punnonen, R.H.; Jokela, H.A.; Dastidar, P.S.; Nevala, M.; Laippala, P.J. Combined estrogen-progestin replacement therapy prevents atherosclerosis in postmenopausal women. Maturitas 1995, 21, 179–187. [Google Scholar] [CrossRef]
- Rossouw, J.E.; Cushman, M.; Greenland, P.; Lloyd-Jones, D.M.; Bray, P.; Kooperberg, C.; Pettinger, M.; Robinson, J.; Hendrix, S.; Hsia, J. Inflammatory, lipid, thrombotic, and genetic markers of coronary heart disease risk in the Women’s Health Initiative trials of hormone therapy. Arch. Intern. Med. 2008, 168, 2245–2253. [Google Scholar] [CrossRef]
- Asgary, S.; Rastqar, A.; Keshvari, M. Functional food and cardiovascular disease prevention and treatment: A review. J. Am. Coll. Nutr. 2018, 37, 429–455. [Google Scholar] [CrossRef]
- Andrew, R.; Izzo, A.A. Principles of pharmacological research of nutraceuticals. Br. J. Pharmacol. 2017, 174, 1177–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakhtiary, A.; Yassin, Z.; Hanachi, P.; Rahmat, A.; Ahmad, Z.; Jalali, F. Effects of soy on metabolic biomarkers of cardiovascular disease in elderly women with metabolic syndrome. Arch. Iran. Med. 2012, 15, 462–468. [Google Scholar]
- Alam, M.A.; Subhan, N.; Hossain, H.; Hossain, M.; Reza, H.M.; Rahman, M.M.; Ullah, M.O. Hydroxycinnamic acid derivatives: A potential class of natural compounds for the management of lipid metabolism and obesity. Nutr. Metab. 2016, 13, 1–13. [Google Scholar] [CrossRef]
- Vinayagam, R.; Jayachandran, M.; Xu, B. Antidiabetic effects of simple phenolic acids: A comprehensive review. Phyther. Res. 2016, 30, 184–199. [Google Scholar] [CrossRef] [PubMed]
- Sroka, Z.; Cisowski, W. Hydrogen peroxide scavenging, antioxidant and anti-radical activity of some phenolic acids. Food Chem. Toxicol. 2003, 41, 753–758. [Google Scholar] [CrossRef]
- Jain, P.G.; Surana, S.J. Isolation, characterization and hypolipidemic activity of ferulic acid in high-fat-diet-induced hyperlipidemia in laboratory rats. EXCLI J. 2016, 15, 599–613. [Google Scholar] [PubMed]
- Cho, A.-S.; Jeon, S.-M.; Kim, M.-J.; Yeo, J.; Seo, K.-I.; Choi, M.-S.; Lee, M.-K. Chlorogenic acid exhibits anti-obesity property and improves lipid metabolism in high-fat diet-induced-obese mice. Food Chem. Toxicol. 2010, 48, 937–943. [Google Scholar] [CrossRef] [PubMed]
- Ibitoye, O.B.; Ajiboye, T.O. Dietary phenolic acids reverse insulin resistance, hyperglycaemia, dyslipidaemia, inflammation and oxidative stress in high-fructose diet-induced metabolic syndrome rats. Arch. Physiol. Biochem. 2018, 124, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Cherng, Y.G.; Tsai, C.C.; Chung, H.H.; Lai, Y.W.; Kuo, S.C.; Cheng, J.T. Antihyperglycemic action of sinapic acid in diabetic rats. J. Agric. Food Chem. 2013, 61, 12053–12059. [Google Scholar] [CrossRef] [PubMed]
- Zych, M.; Folwarczna, J.; Trzeciak, H.I. Natural phenolic acids may increase serum estradiol level in ovariectomized rats. Acta Biochim. Pol. 2009, 56, 503–507. [Google Scholar]
- Zych, M.; Kaczmarczyk-Sedlak, I.; Wojnar, W.; Folwarczna, J. The effects of sinapic acid on the development of metabolic disorders induced by estrogen deficiency in rats. Oxid. Med. Cell. Longev. 2018, 2018, 9274246. [Google Scholar] [CrossRef]
- Amoah, S.K.S.; Sandjo, L.P.; Kratz, J.M.; Biavatti, M.W. Rosmarinic acid—Pharmaceutical and clinical aspects. Planta Med. 2016, 82, 388–406. [Google Scholar] [CrossRef]
- Alagawany, M.; Abd El-Hack, M.E.; Farag, M.R.; Gopi, M.; Karthik, K.; Malik, Y.S.; Dhama, K. Rosmarinic acid: Modes of action, medicinal values and health benefits. Anim. Health Res. Rev. 2017, 18, 167–176. [Google Scholar] [CrossRef]
- Nunes, S.; Madureira, A.R.; Campos, D.; Sarmento, B.; Gomes, A.M.; Pintado, M.; Reis, F. Therapeutic and nutraceutical potential of rosmarinic acid—Cytoprotective properties and pharmacokinetic profile. Crit. Rev. Food Sci. Nutr. 2017, 57, 1799–1806. [Google Scholar] [CrossRef]
- Jayanthy, G.; Subramanian, S.P. Rosmarinic acid modulates the antioxidant status and protects pancreatic tissues from glucolipotoxicity mediated oxidative stress in high-fat diet: Streptozotocin-induced diabetic rats. Mol. Cell. Biochem. 2015, 404, 143–159. [Google Scholar]
- Jayanthy, G.; Subramanian, S. RA abrogates hepatic gluconeogenesis and insulin resistance by enhancing IRS-1 and AMPK signalling in experimental type 2 diabetes. RSC Adv. 2015, 5, 44053–44067. [Google Scholar] [CrossRef]
- Jayanthy, G.; Devi, V.R.; Ilango, K.; Subramanian, S.P. Rosmarinic acid mediates mitochondrial biogenesis in insulin resistant skeletal muscle through activation of AMPK. J. Cell. Biochem. 2017, 118, 1839–1848. [Google Scholar] [PubMed]
- Runtuwene, J.; Cheng, K.C.; Asakawa, A.; Amitani, H.; Amitani, M.; Morinaga, A.; Takimoto, Y.; Kairupan, B.H.R.; Inui, A. Rosmarinic acid ameliorates hyperglycemia and insulin sensitivity in diabetic rats, potentially by modulating the expression of PEPCK and GLUT4. Drug Des. Devel. Ther. 2016, 10, 2193–2202. [Google Scholar] [PubMed] [Green Version]
- Ou, J.; Huang, J.; Zhao, D.; Du, B.; Wang, M. Protective effect of rosmarinic acid and carnosic acid against streptozotocin-induced oxidation, glycation, inflammation and microbiota imbalance in diabetic rats. Food Funct. 2018, 9, 851–860. [Google Scholar] [CrossRef]
- Varlamov, O.; Bethea, C.L.; Roberts, C.T.J. Sex-specific differences in lipid and glucose metabolism. Front. Endocrinol. 2015, 5, 241. [Google Scholar] [CrossRef]
- Mauvais-Jarvis, F. Sex differences in metabolic homeostasis, diabetes, and obesity. Biol. Sex Differ. 2015, 6, 14. [Google Scholar] [CrossRef]
- Clayton, J.A.; Collins, F.S. Policy: NIH to balance sex in cell and animal studies. Nature 2014, 509, 282–283. [Google Scholar] [CrossRef] [Green Version]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Witko-Sarsat, V.; Friedlander, M.; Capeillère-Blandin, C.; Nguyen-Khoa, T.; Nguyen, A.T.; Zingraff, J.; Jungers, P.; Descamps-Latscha, B. Advanced oxidation protein products as a novel marker of oxidative stress in uremia. Kidney Int. 1996, 49, 1304–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cervellati, C.; Bergamini, C.M. Oxidative damage and the pathogenesis of menopause related disturbances and diseases. Clin. Chem. Lab. Med. 2016, 54, 739–753. [Google Scholar] [CrossRef]
- Cakir, T.; Goktas, B.; Mutlu, M.F.; Mutlu, I.; Bilgihan, A.; Erdem, M.; Erdem, A. Advanced oxidation protein products and malondialdehyde—The new biological markers of oxidative stress—Are elevated in postmenopausal women. Ginekol. Pol. 2016, 87, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Rasheed, O.F. Serum γ-glutamyltransferase as oxidative stress marker in pre-and postmenopausal Iraqi women. Oman Med. J. 2010, 25, 286–288. [Google Scholar] [CrossRef] [PubMed]
- Taleb-Belkadi, O.; Chaib, H.; Zemour, L.; Fatah, A.; Chafi, B.; Mekki, K. Lipid profile, inflammation, and oxidative status in peri- and postmenopausal women. Gynecol. Endocrinol. 2016, 32, 982–985. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, I.A.L.; Nogueira da Cruz, É.J.S.; Fontenele, A.M.M.; de Figueiredo Neto, J.A. Alterations in postmenopausal plasmatic lipidome. PLoS ONE 2018, 13, e0203027. [Google Scholar] [CrossRef] [PubMed]
- Parhizkar, S.; Latiff, L.A.; Rahman, S.A.; Dollah, M.A. Preventive effect of Nigella sativa on metabolic syndrome in menopause induced rats. J. Med. Plants Res. 2011, 5, 1478–1484. [Google Scholar]
- Morrone, M.D.S.; Schnorr, C.E.; Behr, G.A.; Gasparotto, J.; Bortolin, R.C.; Da Boit Martinello, K.; Saldanha Henkin, B.; Rabello, T.K.; Zanotto-Filho, A.; Gelain, D.P.; et al. Curcumin supplementation decreases intestinal adiposity accumulation, serum cholesterol alterations, and oxidative stress in ovariectomized rats. Oxid. Med. Cell. Longev. 2016, 2016, 5719291. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.-H.; Liu, Y.; Kim, H.-S. Hawthorn fruit extract elevates expression of Nrf2/HO-1 and improves lipid profiles in ovariectomized rats. Nutrients 2016, 8, 283. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, P. The laboratory rat: Relating its age with human’s. Int. J. Prev. Med. 2013, 4, 624–630. [Google Scholar] [PubMed]
- Folwarczna, J.; Pytlik, M.; Zych, M.; Cegieła, U.; Nowińska, B.; Kaczmarczyk-Sedlak, I.; Śliwiński, L.; Trzeciak, H.; Trzeciak, H.I. Effects of caffeic and chlorogenic acids on the rats skeletal system. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 682–693. [Google Scholar] [PubMed]
- Uesugi, T.U.; Toda, T.T.; Tsuji, K.T.; Ishida, H.I. Comparative study on reduction of bone loss and lipid metabolism abnormality in ovariectomized rats by soy isoflavones, daidzin, genistin, and glycitin. Biol. Pharm. Bull. 2001, 24, 368–372. [Google Scholar] [CrossRef] [PubMed]
- Tawfik, S.H.; Mahmoud, B.F.; Saad, M.I.; Shehata, M.; Kamel, M.A.; Helmy, M.H. Similar and additive effects of ovariectomy and diabetes on insulin resistance and lipid metabolism. Biochem. Res. Int. 2015, 2015, 567945. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.-O.; Lee, I.; Paik, S.-Y.-R.; Kim, D.E.; Lim, J.D.; Kang, W.-S.; Ko, S. Ultrafine Angelica gigas powder normalizes ovarian hormone levels and has antiosteoporosis properties in ovariectomized rats: Particle size effect. J. Med. Food 2012, 15, 863–872. [Google Scholar] [CrossRef] [PubMed]
- Sivasinprasasn, S.; Sa-nguanmoo, P.; Pratchayasakul, W.; Kumfu, S.; Chattipakorn, S.C.; Chattipakorn, N. Obese-insulin resistance accelerates and aggravates cardiometabolic disorders and cardiac mitochondrial dysfunction in estrogen-deprived female rats. Age 2015, 37, 28. [Google Scholar] [CrossRef] [PubMed]
- Rani, V.; Deep, G.; Singh, R.K.; Palle, K.; Yadav, U.C.S. Oxidative stress and metabolic disorders: Pathogenesis and therapeutic strategies. Life Sci. 2016, 148, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Provan, G.J.; Helliwell, K. Determination of rosmarinic acid and caffeic acid in aromatic herbs by HPLC. Food Chem. 2004, 87, 307–311. [Google Scholar] [CrossRef]
- Fecka, I.; Turek, S. Determination of water-soluble polyphenolic compounds in commercial herbal teas from Lamiaceae: Peppermint, melissa, and sage. J. Agric. Food Chem. 2007, 55, 10908–10917. [Google Scholar] [CrossRef] [PubMed]
- European Medicines Agency. EMA/HMPC/196746/2012 Assessment Report on Melissa officinalis L., Folium; European Medicines Agency: London, UK, 2013; pp. 1–20. [Google Scholar]
- European Medicines Agency. EMEA/HMPC/193910/2007 Assessment Report on Mentha x piperita L., Folium; European Medicines Agency: London, UK, 2008; pp. 1–14. [Google Scholar]
- European Medicines Agency. EMA/HMPC/13631/2009 Assessment Report on Rosmarinus officinalis L., Aetheroleum and Rosmarinus officinalis L., Folium; European Medicines Agency: London, UK, 2010; Volume 44, pp. 1–31. [Google Scholar]
- Shin, J.; Seol, I.; Son, C. Interpretation of animal dose and human equivalent dose for drug development. J. Korean Orient. Med. 2010, 31, 1–7. [Google Scholar]
- Sotnikova, R.; Kaprinay, B.; Navarova, J. Rosmarinic acid mitigates signs of systemic oxidative stress in streptozotocin-induced diabetes in rats. Gen. Physiol. Biophys. 2015, 34, 449–452. [Google Scholar]
- Lu, C.; Zou, Y.; Liu, Y.; Niu, Y. Rosmarinic acid counteracts activation of hepatic stellate cells via inhibiting the ROS-dependent MMP-2 activity: Involvement of Nrf2 antioxidant system. Toxicol. Appl. Pharmacol. 2017, 318, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Tandogan, B.; Kuruüzüm-Uz, A.; Sengezer, C.; Güvenalp, Z.; Demirezer, L.Ö.; Nuray Ulusu, N. In vitro effects of rosmarinic acid on glutathione reductase and glucose 6-phosphate dehydrogenase. Pharm. Biol. 2011, 49, 587–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baba, S.; Osakabe, N.; Natsume, M.; Terao, J. Orally administered rosmarinic acid is present as the conjugated and/or methylated forms in plasma, and is degraded and metabolized to conjugated forms of caffeic acid, ferulic acid and m-coumaric acid. Life Sci. 2004, 75, 165–178. [Google Scholar] [CrossRef]
- Enns, G.M.; Cowan, T.M. Glutathione as a redox biomarker in mitochondrial disease—Implications for therapy. J. Clin. Med. 2017, 6, 50. [Google Scholar] [CrossRef] [PubMed]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef]
- Guo, Z.J.; Niu, H.X.; Hou, F.F.; Zhang, L.; Fu, N.; Nagai, R.; Lu, X.; Chen, B.H.; Shan, Y.X.; Tian, J.W.; et al. Advanced oxidation protein products activate vascular endothelial cells via a RAGE-mediated signaling pathway. Antioxid. Redox Signal. 2008, 10, 1699–1712. [Google Scholar] [CrossRef] [PubMed]
- Capeillère-Blandin, C.; Gausson, V.; Descamps-Latscha, B.; Witko-Sarsat, V. Biochemical and spectrophotometric significance of advanced oxidized protein products. Biochim. Biophys. Acta. 2004, 1689, 91–102. [Google Scholar] [CrossRef] [Green Version]
- Fonteles, A.A.; de Souza, C.M.; de Sousa Neves, J.C.; Menezes, A.P.F.; Santos do Carmo, M.R.; Fernandes, F.D.P.; de Araújo, P.R.; de Andrade, G.M. Rosmarinic acid prevents against memory deficits in ischemic mice. Behav. Brain Res. 2016, 297, 91–103. [Google Scholar] [CrossRef]
- Fan, Y.-T.; Yin, G.-J.; Xiao, W.-Q.; Qiu, L.; Yu, G.; Hu, Y.-L.; Xing, M.; Wu, D.-Q.; Cang, X.-F.; Wan, R.; et al. Rosmarinic acid attenuates sodium taurocholate-induced acute pancreatitis in rats by inhibiting nuclear factor-κB activation. Am. J. Chin. Med. 2015, 43, 1117–1135. [Google Scholar] [CrossRef]
- Ozturk, H.; Ozturk, H.; Terzi, E.H.; Ozgen, U.; Duran, A.; Uygun, I. Protective effects of rosmarinic acid against renal ischaemia/reperfusion injury in rats. J. Pak. Med. Assoc. 2014, 64, 260–265. [Google Scholar]
- Ou, J.; Huang, J.; Wang, M.; Ou, S. Effect of rosmarinic acid and carnosic acid on AGEs formation in vitro. Food Chem. 2017, 221, 1057–1061. [Google Scholar] [CrossRef] [PubMed]
- Vlavcheski, F.; Naimi, M.; Murphy, B.; Hudlicky, T.; Tsiani, E. Rosmarinic acid, a rosemary extract polyphenol, increases skeletal muscle cell glucose uptake and activates AMPK. Molecules 2017, 22, 1669. [Google Scholar] [CrossRef] [PubMed]
- Casanova, L.M.; Gu, W.; Costa, S.S.; Jeppesen, P.B. Phenolic substances from Ocimum species enhance glucose-stimulated insulin secretion and modulate the expression of key insulin regulatory genes in mice pancreatic islets. J. Nat. Prod. 2017, 80, 3267–3275. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, M.F.; Lima, C.F.; Fernandes-Ferreira, M.; Almeida, M.J.; Wilson, J.M.; Pereira-Wilson, C. Rosmarinic acid, major phenolic constituent of Greek sage herbal tea, modulates rat intestinal SGLT1 levels with effects on blood glucose. Mol. Nutr. Food Res. 2011, 55, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Wang, D.; Ye, L.; Li, P.; Hao, W.; Chen, X.; Ma, J.; Wang, B.; Shang, J.; Li, D.; et al. Rosmarinic acid protects against inflammation and cardiomyocyte apoptosis during myocardial ischemia/reperfusion injury by activating peroxisome proliferator-activated receptor gamma. Front. Pharmacol. 2017, 8, 456. [Google Scholar] [CrossRef]
- Yki-Järvinen, H. Thiazolidinediones. N. Engl. J. Med. 2004, 351, 1106–1118. [Google Scholar] [CrossRef] [PubMed]
- Javidanpour, S.; Dianat, M.; Badavi, M.; Mard, S.A. The inhibitory effect of rosmarinic acid on overexpression of NCX1 and stretch- induced arrhythmias after acute myocardial infarction in rats. Biomed. Pharmacother. 2018, 102, 884–893. [Google Scholar] [CrossRef] [PubMed]
- Barakat, R.; Oakley, O.; Kim, H.; Jin, J.; Ko, C.J. Extra-gonadal sites of estrogen biosynthesis and function. BMB Rep. 2016, 49, 488–496. [Google Scholar] [CrossRef] [Green Version]
- Le Bail, J.C.; Laroche, T.; Marre-Fournier, F.; Habrioux, G. Aromatase and 17β-hydroxysteroid dehydrogenase inhibition by favonoids. Cancer Lett. 1998, 133, 101–106. [Google Scholar] [CrossRef]
- Way, T.-D.; Lee, H.-H.; Kao, M.-C.; Lin, J.-K. Black tea polyphenol theaflavins inhibit aromatase activity and attenuate tamoxifen resistance in HER2/neu -transfected human breast cancer cells through tyrosine kinase suppression. Eur. J. Cancer 2004, 40, 2165–2174. [Google Scholar] [CrossRef]
- Kang, S.J.; Choi, B.R.; Kim, S.H.; Yi, H.Y.; Park, H.R.; Kim, D.C.; Choi, S.H.; Han, C.H.; Park, S.J.; Song, C.H.; et al. Dried pomegranate potentiates anti-osteoporotic and anti-obesity activities of red clover dry extracts in ovariectomized rats. Nutrients 2015, 7, 2622–2647. [Google Scholar] [CrossRef] [PubMed]
- Das, A.S.; Das, D.; Mukherjee, M.; Mukherjee, S.; Mitra, C. Phytoestrogenic effects of black tea extract (Camellia sinensis) in an oophorectomized rat (Rattus norvegicus) model of osteoporosis. Life Sci. 2005, 77, 3049–3057. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter/Group | SHAM | OVX | OVX + RA10 | OVX + RA50 |
|---|---|---|---|---|
| SOD (U/mg of protein) | 5.22 ± 0.19 | 6.20 ± 0.34 * | 6.39 ± 0.35 ** | 6.21 ± 0.27 * |
| CAT (nmol/min/mg of protein) | 0.52 ± 0.08 | 0.74 ± 0.19 | 0.74 ± 0.14 | 0.91 ± 0.25 |
| TBARS (nmol/mL) | 21.5 ± 1.9 | 24.1 ± 1.9 | 19.1 ± 0.7 | 19.9 ± 1.1 |
| AOPP (nmol/mL) | 19.9 ± 1.2 | 25.0 ± 4.4 | 16.9 ± 2.0 # | 13.6 ± 2.2 ## |
| PCG (nmol/mL) | 5.52 ± 0.40 | 5.87 ± 0.85 | 5.16 ± 0.44 | 5.00 ± 0.95 |
| Parameter/Group | SHAM | OVX | OVX + RA10 | OVX + RA50 |
|---|---|---|---|---|
| Interleukin 18 (pg/mL) | 312.0 ± 13.7 | 235.0 ± 22.7 | 298.0 ± 27.3 | 315.9 ± 33.5 |
| AST (U/L) | 39.22 ± 3.38 | 34.72 ± 2.78 | 37.60 ± 3.57 | 41.61 ± 2.57 |
| ALT (U/L) | 23.68 ± 2.19 | 23.52 ± 1.38 | 25.20 ± 1.86 | 30.65 ± 3.84 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zych, M.; Kaczmarczyk-Sedlak, I.; Wojnar, W.; Folwarczna, J. Effect of Rosmarinic Acid on the Serum Parameters of Glucose and Lipid Metabolism and Oxidative Stress in Estrogen-Deficient Rats. Nutrients 2019, 11, 267. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11020267
Zych M, Kaczmarczyk-Sedlak I, Wojnar W, Folwarczna J. Effect of Rosmarinic Acid on the Serum Parameters of Glucose and Lipid Metabolism and Oxidative Stress in Estrogen-Deficient Rats. Nutrients. 2019; 11(2):267. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11020267
Chicago/Turabian StyleZych, Maria, Ilona Kaczmarczyk-Sedlak, Weronika Wojnar, and Joanna Folwarczna. 2019. "Effect of Rosmarinic Acid on the Serum Parameters of Glucose and Lipid Metabolism and Oxidative Stress in Estrogen-Deficient Rats" Nutrients 11, no. 2: 267. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11020267