Insulin Controls Triacylglycerol Synthesis through Control of Glycerol Metabolism and Despite Increased Lipogenesis

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Rats and Sampling
2.2. WAT Adipocyte Isolation and Incubation Procedures
2.3. Processing of the Incubation Media: Label Distribution
2.4. Processing of Labelled Cell Components
2.5. Analysis of Metabolites in the Medium
2.6. Gene Expression Analysis
2.7. Statistical Procedures
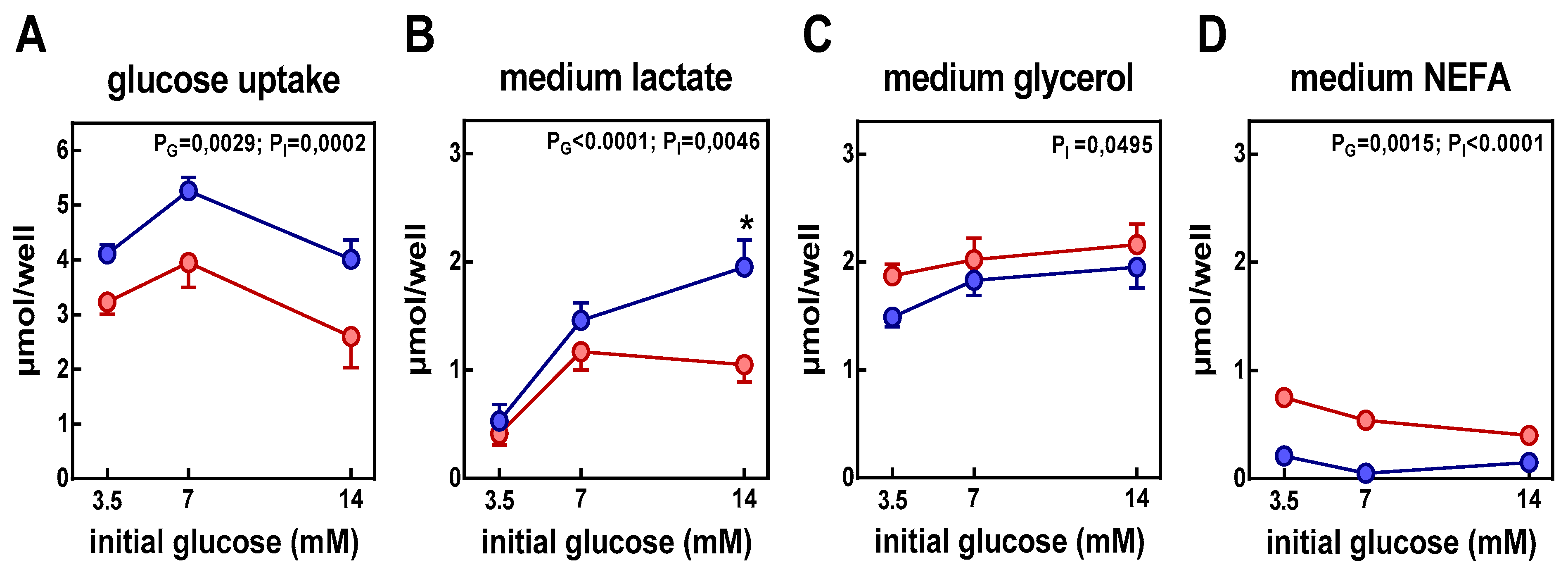
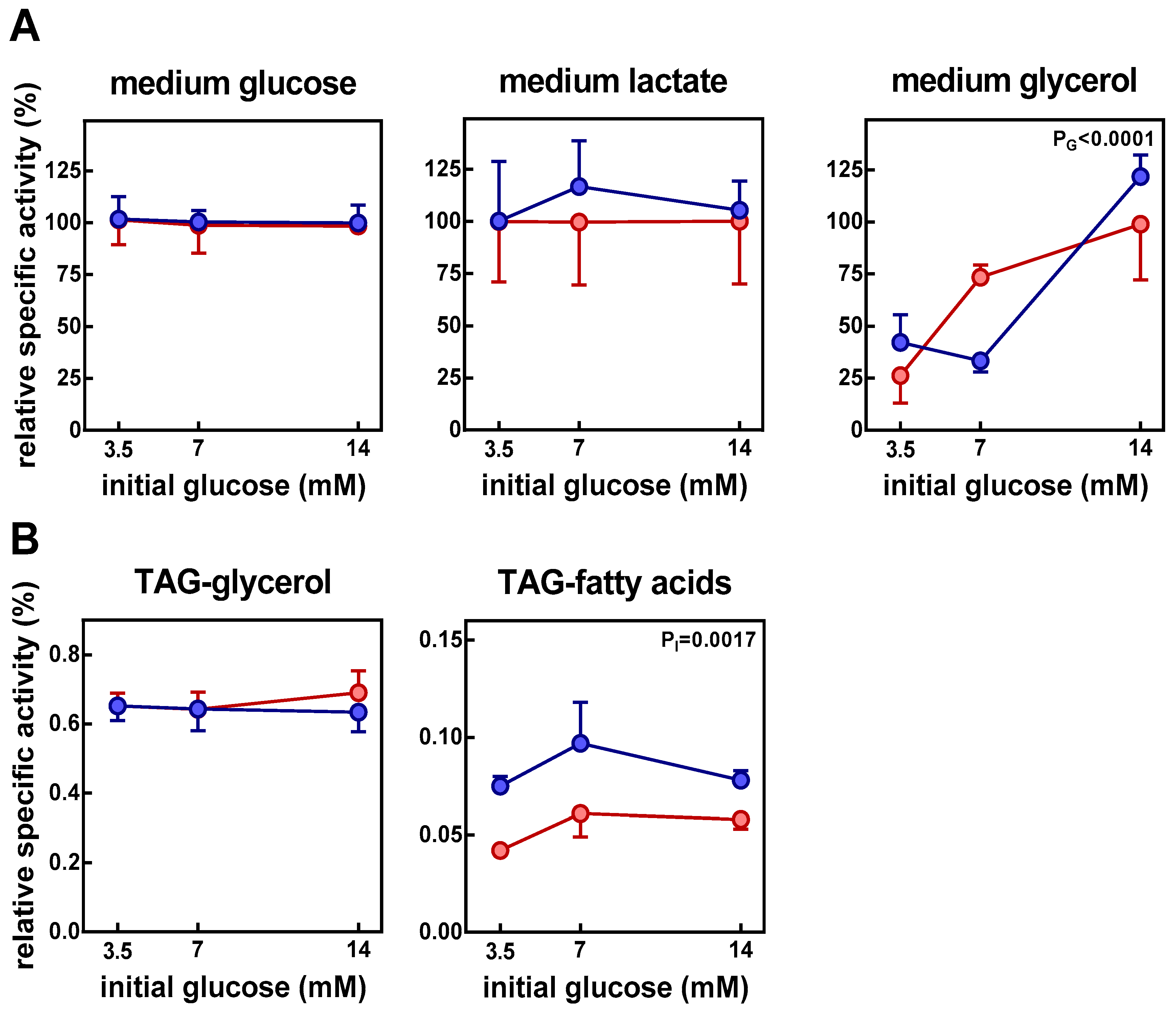
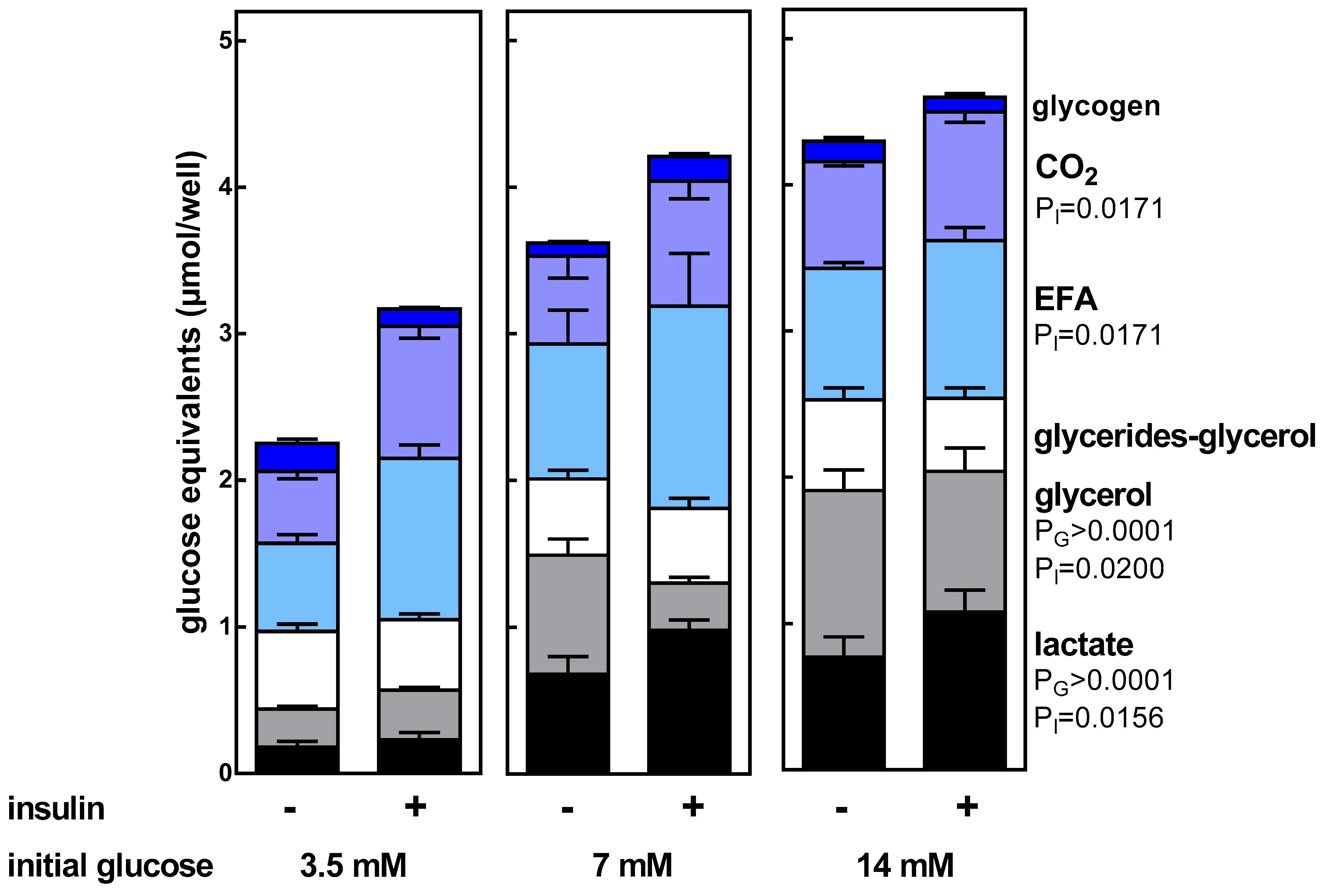
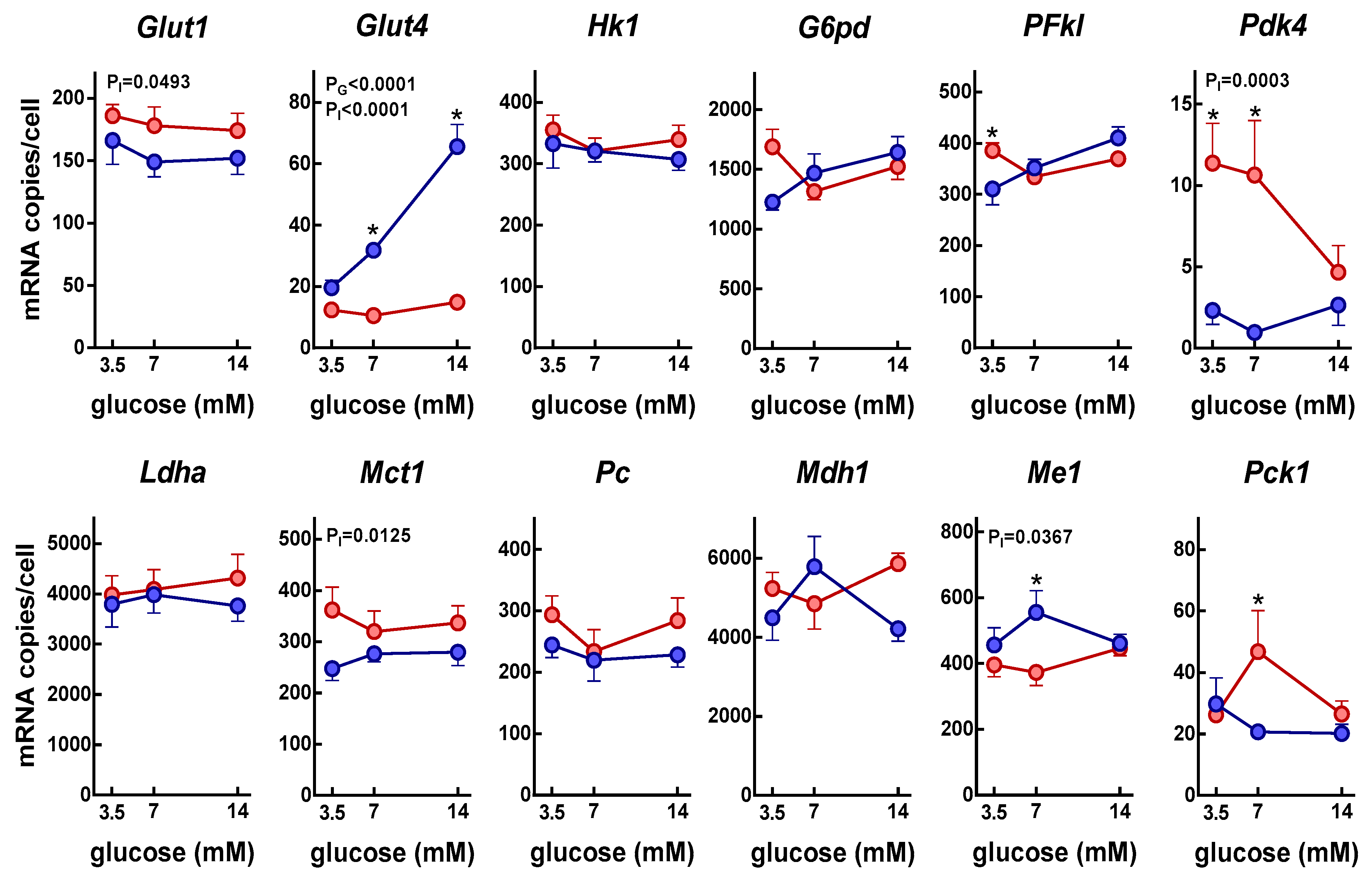
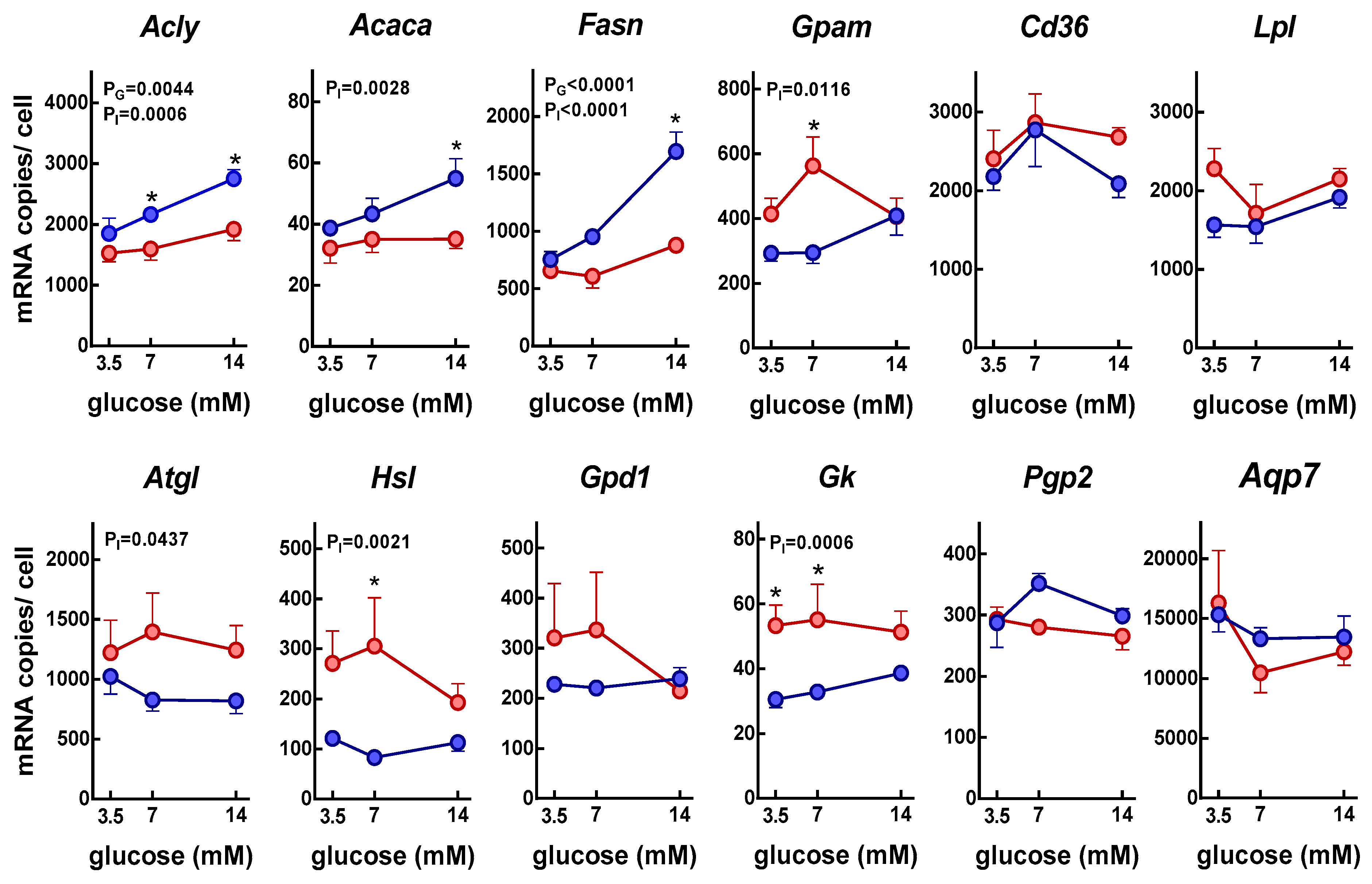
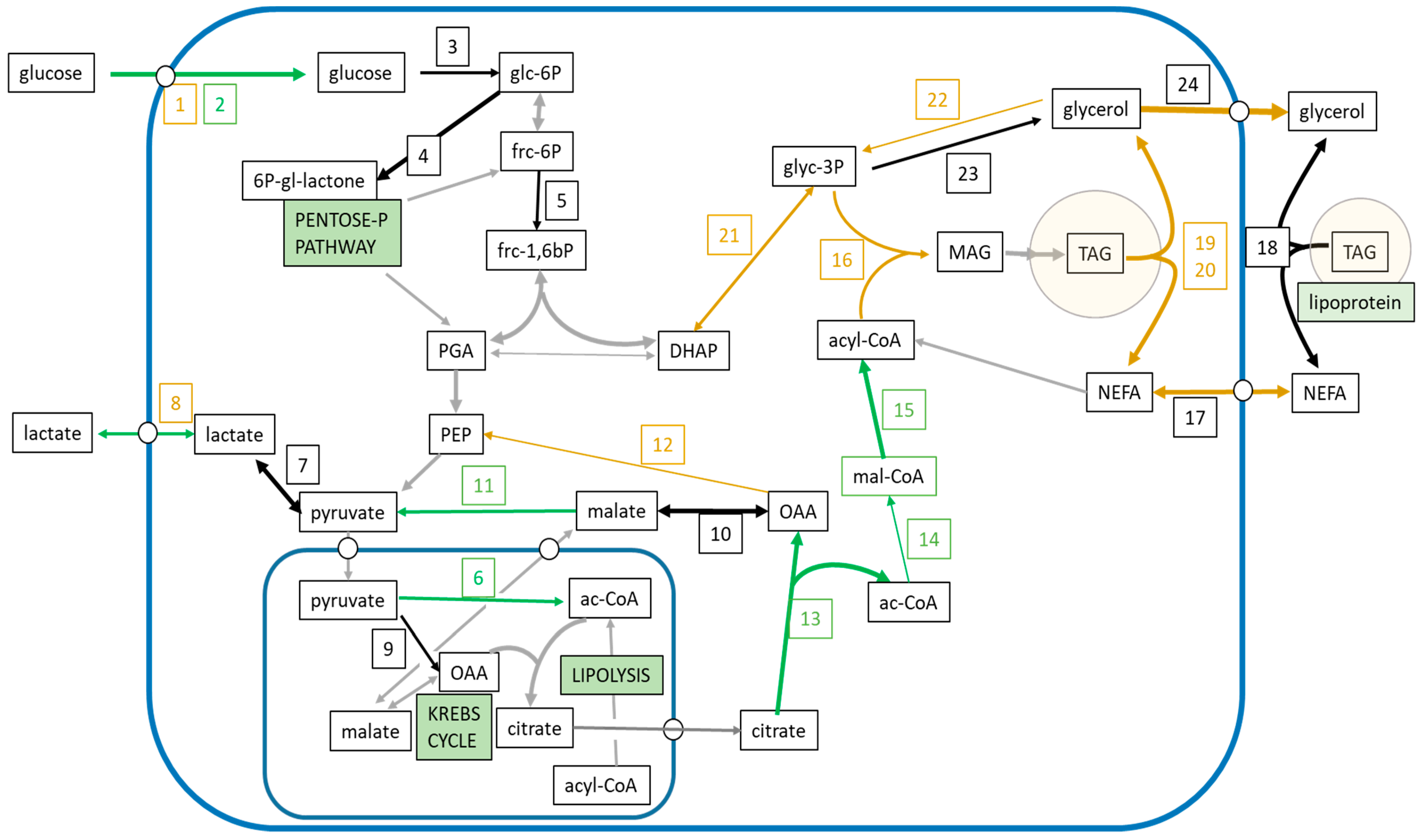
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Cinti, S. The adipose organ. Prostaglandins Leukot. Essent. Fat. Acids 2005, 73, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Romero, M.M.; Roy, S.; Pouillot, K.; Feito, M.; Esteve, M.; Grasa, M.M.; Fernández-López, J.A.; Alemany, M.; Remesar, X. Treatment of rats with a self-selected hyperlipidic diet, increases the lipid content of the main adipose tissue sites in a proportion similar to that of the lipids in the rest of organs and tissues. PLoS ONE 2014, 9, e90995. [Google Scholar] [CrossRef] [PubMed]
- Trayhurn, P. Endocrine and signalling role of adipose tissue: New perspectives on fat. Acta Physiol. Scand. 2005, 184, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Hauner, H. The new concept of adipose tissue function. Physiol. Behav. 2004, 83, 653–658. [Google Scholar] [CrossRef] [PubMed]
- Alemany, M.; Fernández-López, J.A. Adipose tissue: Something more than just adipocytes. Curr. Nutr. Food Sci. 2006, 2, 141–150. [Google Scholar] [CrossRef]
- Rosen, E.D.; Spiegelman, B.M. Adipocytes as regulators of energy balance and glucose homeostasis. Nature 2006, 444, 847–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arner, P.; Bernard, S.; Salehpour, M.; Possnert, G.; Liebl, J.; Steier, P.; Buchholz, B.A.; Eriksson, M.; Arner, E.; Hauner, H.; et al. Dynamics of human adipose lipid turnover in health and metabolic disease. Nature 2011, 478, 110–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arriarán, S.; Agnelli, S.; Remesar, X.; Fernández-López, J.A.; Alemany, M. Effects of sex and site on amino acid metabolism enzyme gene expression and activity in rat white adipose tissue. PeerJ 2015, 3, e1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tchoukalova, Y.D.; Sarr, M.G.; Jensen, M.D. Measuring committed preadipocytes in human adipose tissue from severely obese patients by using adipocyte fatty acid binding protein. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 287, R1132–R1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, A.R.; Milner, J.J.; Makowski, L. The inflammation highway: Metabolism accelerates inflammatory traffic in obesity. Immunol. Rev. 2012, 249, 218–238. [Google Scholar] [CrossRef] [PubMed]
- Makki, K.; Froguel, P.; Wolowczuk, I. Adipose tissue in obesity-related inflammation and insulin resistance: Cells, cytokines, and chemokines. ISRN Inflamm. 2013, 139239. [Google Scholar] [CrossRef] [PubMed]
- Dandona, P.; Aljada, A.; Chaudhuri, A.; Mohanty, P.; Garg, R. Metabolic syndrome: A comprehensive perspective based on interactions between obesity, diabetes, and inflammation. Circulation 2005, 111, 1448–1454. [Google Scholar] [CrossRef] [PubMed]
- Ho-Palma, A.C.; Rotondo, F.; Romero, M.M.; Memmolo, S.; Remesar, X.; Fernández-López, J.A.; Alemany, M. A method for the measurement of lactate, glycerol and fatty acid production from 14C-glucose in primary cultures of rat epididymal adipocytes. Anal. Methods 2016, 8, 7873–7885. [Google Scholar] [CrossRef]
- Rotondo, F.; Ho-Palma, A.C.; Remesar, X.; Fernández-López, J.A.; Romero, M.M.; Alemany, M. Glycerol is synthesized and secreted by adipocytes to dispose of excess glucose, via glycerogenesis and increased acyl-glycerol turnover. Sci. Rep. 2017, 7, 8983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arriarán, S.; Agnelli, S.; Sabater, D.; Remesar, X.; Fernández-López, J.A.; Alemany, M. Evidences of Basal lactate production in the main white adipose tissue sites of rats. Effects of sex and a cafeteria diet. PLoS ONE 2015, 10, e0119572. [Google Scholar] [CrossRef] [PubMed]
- Hodson, L. Adipose tissue oxygenation: Effects on metabolic function. Adipocyte 2014, 3, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Goossens, G.H.; Bizzarri, A.; Venteclef, N.; Essers, Y.; Cleutjens, J.P.; Konings, E.; Jocken, J.W.; Cajlakovic, M.; Ribitsch, V.; Clément, K.; et al. Increased adipose tissue oxygen tension in obese compared with lean men is accompanied by insulin resistance, impaired adipose tissue capillarization, and inflammation. Circulation 2011, 124, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Hodson, L.; Humphreys, S.M.; Karpe, F.; Frayn, K.N. Metabolic signatures of human adipose tissue hypoxia in obesity. Diabetes 2013, 62, 1417–1425. [Google Scholar] [CrossRef] [PubMed]
- DiGirolamo, M.; Newby, F.D.; Lovejoy, J. Lactate production in adipose tissue: A regulated function with extra-adipose implications. FASEB J. 1992, 6, 2405–2412. [Google Scholar] [CrossRef] [PubMed]
- Rodbell, M. Metabolism of Isolated Fat Cells: I. Effects of hormones on glucose metabolism and lipolysis. J. Biol. Chem. 1964, 239, 375–380. [Google Scholar] [PubMed]
- Krebs, H.A.; Henseleit, K. Untersuchungen über die Harnstoffbildung im Tierkörper. Klin. Wochenschr. 1932, 11, 757–759. [Google Scholar] [CrossRef]
- Rotondo, F.; Romero, M.M.; Ho-Palma, A.C.; Remesar, X.; Fernández-López, J.A.; Alemany, M. Quantitative analysis of rat adipose tissue cell recovery, and non-fat cell volume, in primary cell cultures. PeerJ 2016, 4, e2725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabater, D.; Arriarán, S.; Romero, M.M.; Agnelli, S.; Remesar, X.; Fernández-López, J.A.; Alemany, M. Cultured 3T3L1 adipocytes dispose of excess medium glucose as lactate under abundant oxygen availability. Sci. Rep. 2014, 4, 3663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azain, M.J.; Kasser, T.R.; Baile, C.A. A method for the determination of glucose synthesis in isolated bovine hepatocytes. J. Nutr. Biochem. 1999, 10, 205–209. [Google Scholar] [CrossRef]
- Crespi, C.; Gastalver, G.; Palou, A.; Roca, P. Enzymatic determination of carbon (14C)-labeled glycerol in biological samples. J. Biochem. Biophys. Methods 1995, 30, 179–183. [Google Scholar] [CrossRef]
- Ferraz, T.P.L.; Fiúza, M.C.; Dos Santos, M.L.A.; Pontes de Carvalho, L.; Soares, N.M. Comparison of six methods for the extraction of lipids from serum in terms of effectiveness and protein preservation. J. Biochem. Biophys. Methods 2004, 58, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Oliva, L.; Baron, C.; Fernández-López, J.A.; Remesar, X.; Alemany, M. Marked increase in rat red blood cell membrane protein glycosylation by one-month treatment with a cafeteria diet. PeerJ 2015, 3, e1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, M.M.; Grasa, M.M.; Esteve, M.; Fernández-López, J.A.; Alemany, M. Semiquantitative RT-PCR measurement of gene expression in rat tissues including a correction for varying cell size and number. Nutr. Metab. 2007, 4, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bamias, G.; Goukos, D.; Laoudi, E.; Balla, I.G.; Siakavellas, S.I.; Daikos, G.L.; Ladas, S.D. Comparative study of candidate housekeeping genes for quantification of target gene messenger RNA expression by real-time PCR in patients with inflammatory bowel disease. Inflamm. Bowel Dis. 2013, 19, 2840–2847. [Google Scholar] [CrossRef] [PubMed]
- Rotondo, F.; Ho-Palma, A.C.; Remesar, X.; Fernández-López, J.A.; Romero, M.M.; Alemany, M. Effect of sex on glucose handling by adipocytes isolated from rat subcutaneous, mesenteric and perigonadal adipose tissue. PeerJ 2018, 6, e5440. [Google Scholar] [CrossRef] [PubMed]
- Rotondo, F.; Ho-Palmaa, A.C.; Romero, M.M.; Remesar, X.; Fernández-López, J.A.; Alemany, M. Higher lactate production from glucose in cultured adipose nucleated stromal cells than for rat adipocytes. Adipocyte 2019. [Google Scholar] [CrossRef] [PubMed]
- Ross, B.D.; Hems, R.; Krebs, H.A. The rate of gluconeogenesis from various precursors in the perfused rat liver. Biochem. J. 1967, 102, 942–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katz, J.; Wals, P.A. Lipogenesis from lactate in rat adipose tissue. Biochim. Biophys. Acta 1974, 348, 344–356. [Google Scholar] [CrossRef]
- Alemany, M. Regulation of adipose tissue energy availability through blood flow control in the metabolic syndrome. Free Radic. Biol. Med. 2012, 52, 2108–2119. [Google Scholar] [CrossRef] [PubMed]
- Jansson, P.A.; Larsson, A.; Smith, U.; Lonnroth, P. Glycerol production in subcutaneous adipose tissue in lean and obese humans. J. Clin. Investig. 1992, 89, 1610–1617. [Google Scholar] [CrossRef] [PubMed]
- Watson, R.T.; Kanzaki, M.; Pessin, J.E. Regulated membrane trafficking of the insulin-responsive glucose transporter 4 in adipocytes. Endocr. Rev. 2004, 25, 177–204. [Google Scholar] [CrossRef] [PubMed]
- Saltiel, A.R.; Kahn, C.R. Insulin signalling and the regulation of glucose and lipid metabolism. Nature 2001, 414, 799–806. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Czech, M.P. The GLUT4 glucose transporter. Cell Metab. 2007, 5, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Manolescu, A.R.; Witkowska, K.; Kinnaird, A.; Cessford, T.; Cheeseman, C. Facilitated hexose transporters: New perspectives on form and function. Physiology 2007, 22, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Kaestner, K.H.; Christy, R.J.; McLenithan, J.C.; Braiterman, L.T.; Cornelius, P.; Pekala, P.H.; Lane, M.D. Sequence, tissue distribution, and differential expression of mRNA for a putative insulin-responsive glucose transporter in mouse 3T3-L1 adipocytes. Proc. Natl. Acad. Sci. USA 1989, 86, 3150–3154. [Google Scholar] [CrossRef] [PubMed]
- Behrooz, A.; Ismail-Beigi, F. Dual control of glut1 glucose transporter gene expression by hypoxia and by inhibition of oxidative phosphorylation. J. Biol. Chem. 1997, 272, 5555–5562. [Google Scholar] [CrossRef] [PubMed]
- Fandos, C.; Sánchez-Feutrie, M.; Santalucía, T.; Viñals, F.; Cadefau, J.; Gumà, A.; Cussó, R.; Kaliman, P.; Canicio, J.; Palacín, M.; et al. GLUT1 glucose transporter gene transcription is repressed by Sp3. Evidence for a regulatory role of Sp3 during myogenesis. J. Mol. Biol. 1999, 294, 103–119. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.Y.; Ismail-Beigi, F. Control of Glut1 promoter activity under basal conditions and in response to hyperosmolarity: Role of Sp1. Am. J. Physiol. Cell Physiol. 2006, 290, C337–C344. [Google Scholar] [CrossRef] [PubMed]
- Leto, D.; Saltiel, A.R. Regulation of glucose transport by insulin: Traffic control of GLUT4. Nat. Rev. Mol. Cell Biol. 2012, 13, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Wood, I.S.; Wang, B.; Lorente-Cebrián, S.; Trayhurn, P. Hypoxia increases expression of selective facilitative glucose transporters (GLUT) and 2-deoxy-d-glucose uptake in human adipocytes. Biochem. Biophys. Res. Commun. 2007, 361, 468–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansour, T.E. Phosphofructokinase. Curr. Top. Cell. Regul. 1972, 5, 1–46. [Google Scholar] [PubMed]
- Zhang, Y.; Zeng, X.; Jin, S. Autophagy in adipose tissue biology. Pharm. Res. 2012, 66, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Trayhurn, P. Hypoxia and adipose tissue function and dysfunction in obesity. Physiol. Rev. 2013, 93, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.T.; Hsia, D.S.; Chacko, S.K.; Rodriguez, L.M.; Haymond, M.W. Increased gluconeogenesis in youth with newly diagnosed type 2 diabetes. Diabetologia 2015, 58, 596–603. [Google Scholar] [CrossRef] [PubMed]
- van der Merwe, M.T.; Crowther, N.J.; Schlaphoff, G.P.; Boyd, I.H.; Gray, I.P.; Joffe, B.I.; Lönnroth, P.N. Lactate and glycerol release from the subcutaneous adipose tissue of obese urban women from South Africa; important metabolic implications. J. Clin. Endocrinol. Metab. 1998, 83, 4084–4091. [Google Scholar] [CrossRef] [PubMed]
- Fredholm, B.B. Adenosine and lipolysis. Int. J. Obes. 1981, 5, 643–649. [Google Scholar] [PubMed]
- Romero, M.M.; Sabater, D.; Fernández-López, J.A.; Remesar, X.; Alemany, M. Glycerol production from glucose and fructose by 3T3-L1 Cells: A mechanism of adipocyte defense from excess substrate. PLoS ONE 2015, 10, e0139502. [Google Scholar] [CrossRef] [PubMed]
- Mugabo, Y.; Zhao, S.; Seifried, A.; Gezzar, S.; Al-Mass, A.; Zhang, D.; Lamontagne, J.; Attane, C.; Poursharifi, P.; Iglesias, J.; et al. Identification of a mammalian glycerol-3-phosphate phosphatase: Role in metabolism and signaling in pancreatic β-cells and hepatocytes. Proc. Natl. Acad. Sci. USA 2016, 113, E430–E439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Loh, K.; Song, Z.Y.; Yang, H.Q.; Zhang, Y.; Lin, S. Update on glycerol-3-phosphate acyltransferases: The roles in the development of insulin resistance. Nutr. Diabetes 2018, 8, 34. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.Y.; Repa, J.J. The Liver X Receptor (LXR) and hepatic lipogenesis. The carbohydrate-response element-binding protein is a target gene of LXR. J. Biol. Chem. 2007, 282, 743–751. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, K.; Reue, K. Biochemistry, physiology, and genetics of GPAT, AGPAT, and lipin enzymes in triglyceride synthesis. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E1195–E1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boden, G.; Salehi, S.; Cheung, P.; Homko, C.; Song, W.; Loveland-Jones, C.; Jayarajan, S. Comparison of in vivo effects of insulin on SREBP-1c activation and INSIG-1/2 in rat liver and human and rat adipose tissue. Obesity 2013, 21, 1208–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carneiro, L.; Asrih, M.; Repond, C.; Sempoux, C.; Stehle, J.C.; Leloup, C.; Jornayvaz, F.R.; Pellerin, L. AMPK activation caused by reduced liver lactate metabolism protects against hepatic steatosis in MCT1 haploinsufficient mice. Mol. Metab. 2017, 6, 1625–1633. [Google Scholar] [CrossRef] [PubMed]
- Rafecas, I.; Fernández-López, J.A.; Salinas, I.; Formiguera, X.; Remesar, X.; Foz, M.; Alemany, M. Insulin degradation by adipose tissue is increased in human obesity. J. Clin. Endocrinol. Metab. 1995, 80, 693–695. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | 5′ primer | 3′ primer | bp | |
|---|---|---|---|---|
| Glut1 | glucose transporter type 1 | GCTCGGGTATCGTCAACACG | ATGCCAGCCAGACCAATGAG | 97 |
| Glut4 | glucose transporter type 4 | CTTGATGACGGTGGCTCTGC | CACAATGAACCAGGGGATGG | 127 |
| Hk1 | hexokinase 1 | TGGATGGGACGCTCTACAAA | GACAGGAGGAAGGACACGGTA | 100 |
| G6pd | glucose-6-phosphate dehydrogenase | GACTGTGGGCAAGCTCCTCAA | GCTAGTGTGGCTATGGGCAGGT | 77 |
| PFkL | phospho-fructokinase, liver, b-type | CAGCCACCATCAGCAACAAT | TGCGGTCACAACTCTCCATT | 90 |
| Pdk4 | pyruvate dehydrogenase kinase 4 | CTGCTCCAACGCCTGTGAT | GCATCTGTCCCATAGCCTGA | 142 |
| Ldha | L-lactate dehydrogenase a | AAAGGCTGGGAGTTCATCCA | CGGCGACATTCACACCACT | 96 |
| Mct1 | monocarboxylate transporter 1 | CCCAGAGGTTCTCCAGTGCT | ACGCCACAAGCCCAGTATGT | 133 |
| Pc | pyruvate carboxylase | GCCAGAGGCAGGTGTTCTTTG | TTTGGCCCTTCACATCCTTCA | 120 |
| Mdh1 | malate dehydrogenase 1 | GCTGGCTCAAGGGAGAGTTC | TCTCATGTGGTCCGAGATGG | 116 |
| Me1 | NADP+-dependent malic enzyme | GGAGTTGCTCTTGGGGTAGTGG | CGGATGGTGTTCAAAGGAGGA | 143 |
| Pck1 | phosphoenolpyruvate carboxykinase 1 | CGGGTGGAAAGTTGAATGTG | AATGGCGTTCGGATTTGTCT | 142 |
| Acly | ATP citrate lyase | TGTGCTGGGAAGGAGTATGG | GCTGCTGGCTCGGTTACAT | 137 |
| Acaca | acetyl-coA carboxylase alpha | AGGAAGATGGTGTCCGCTCTG | GGGGAGATGTGCTGGGTCAT | 145 |
| Fasn | fatty acid synthase | CCCGTTGGAGGTGTCTTCA | AAGGTTCAGGGTGCCATTGT | 117 |
| Gpam | glycerol-3P acyl-transferase | GGTGAGGAGCAGCGTGATT | GTGGACAAAGATGGCAGCAG | 129 |
| Cd36 | platelet glycoprotein 4 (fatty acid transporter) | TGGTCCCAGTCTCATTTAGCC | TTGGATGTGGAACCCATAACTG | 154 |
| Lpl | lipoprotein lipase | TGGCGTGGCAGGAAGTCT | CCGCATCATCAGGAGAAAGG | 116 |
| Atgl | triacylglycerol lipase (adipose tissue) | CACCAACACCAGCATCCAAT | CGAAGTCCATCTCGGTAGCC | 120 |
| Hsl | hormone-sensitive lipase | TCCTCTGCTTCTCCCTCTCG | ATGGTCCTCCGTCTCTGTCC | 108 |
| Gpd1 | glycerol-3P dehydrogenase (NAD+) | CTGGAGAAAGAGATGCTGAACG | GCGGTGAACAAGGGAAACTT | 113 |
| Gk | glycerol kinase | ACTTGGCAGAGACAAACCTGTG | ACCAGCGGATTACAGCACCA | 74 |
| Pgp2 | phosphoglycolate phosphatase (glycerophosphatase) | CCTGGACACAGACATCCTCCT | TTCCTGATTGCTCTTCACATCC | 100 |
| Aqp7 | aquaporin 7 | ACAGGTCCCAAATCCACTGC | CCGTGATGGCGAAGATACAC | 127 |
| Arbp | 0 S acidic ribosomal phosphoprotein PO (housekeeping gene) | CCTTCTCCTTCGGGCTGAT | CACATTGCGGACACCCTCTA | 122 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ho-Palma, A.C.; Toro, P.; Rotondo, F.; Romero, M.d.M.; Alemany, M.; Remesar, X.; Fernández-López, J.A. Insulin Controls Triacylglycerol Synthesis through Control of Glycerol Metabolism and Despite Increased Lipogenesis. Nutrients 2019, 11, 513. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11030513
Ho-Palma AC, Toro P, Rotondo F, Romero MdM, Alemany M, Remesar X, Fernández-López JA. Insulin Controls Triacylglycerol Synthesis through Control of Glycerol Metabolism and Despite Increased Lipogenesis. Nutrients. 2019; 11(3):513. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11030513
Chicago/Turabian StyleHo-Palma, Ana Cecilia, Pau Toro, Floriana Rotondo, María del Mar Romero, Marià Alemany, Xavier Remesar, and José Antonio Fernández-López. 2019. "Insulin Controls Triacylglycerol Synthesis through Control of Glycerol Metabolism and Despite Increased Lipogenesis" Nutrients 11, no. 3: 513. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11030513