Enhancing Omega-3 Long-Chain Polyunsaturated Fatty Acid Content of Dairy-Derived Foods for Human Consumption

 , , ,
, , ,
Abstract
:1. Introduction
2. Metabolic Pathways, Human Health Benefits and Recommended Intake of n-3 PUFA
2.1. Dietary n-3 PUFA Intake Recommendations
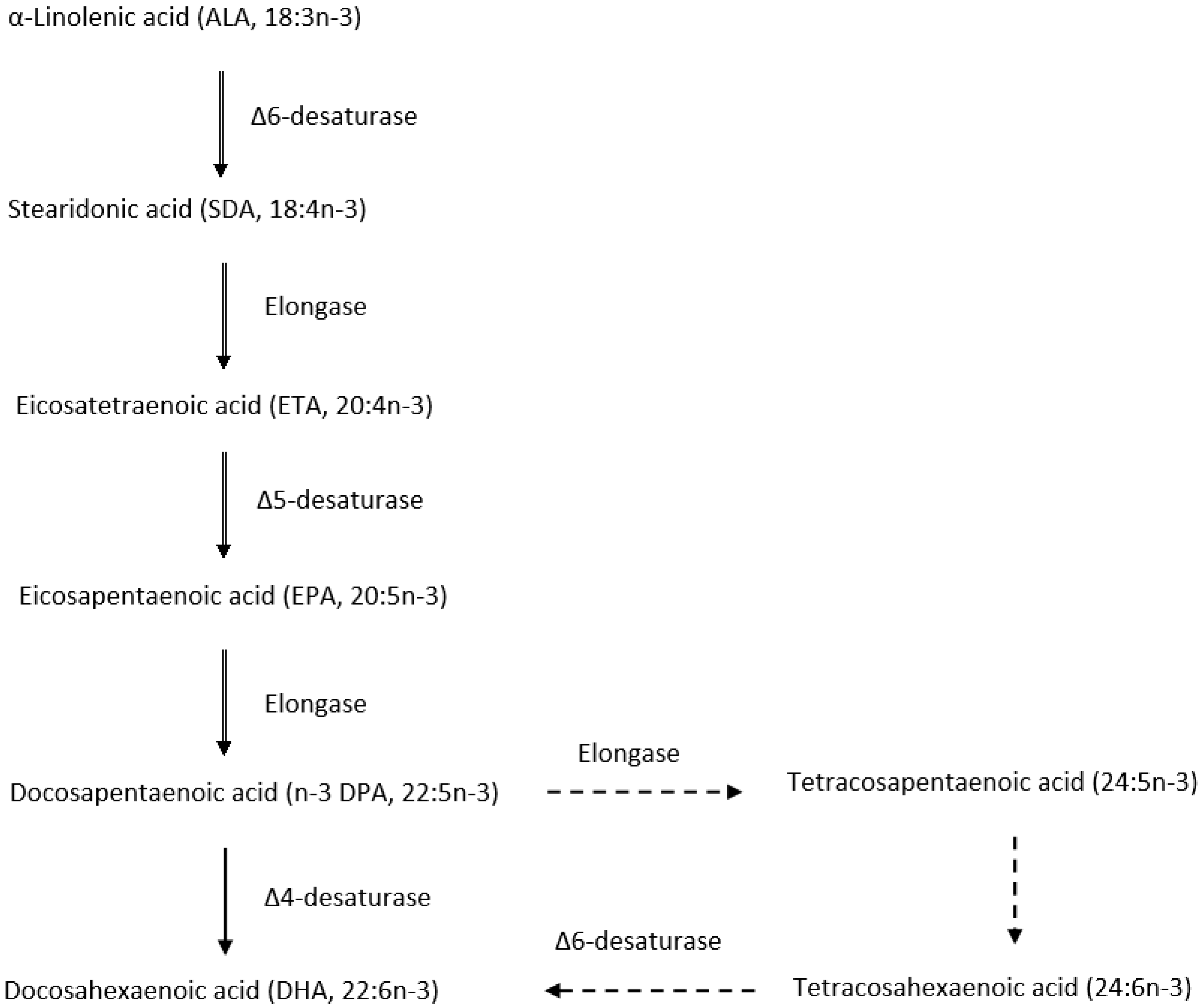
2.2. Metabolic Pathways for the Biosynthesis and Dietary Sources of n-3 PUFA
2.3. n-3 LC-PUFA Consumption and Chronic Diseases
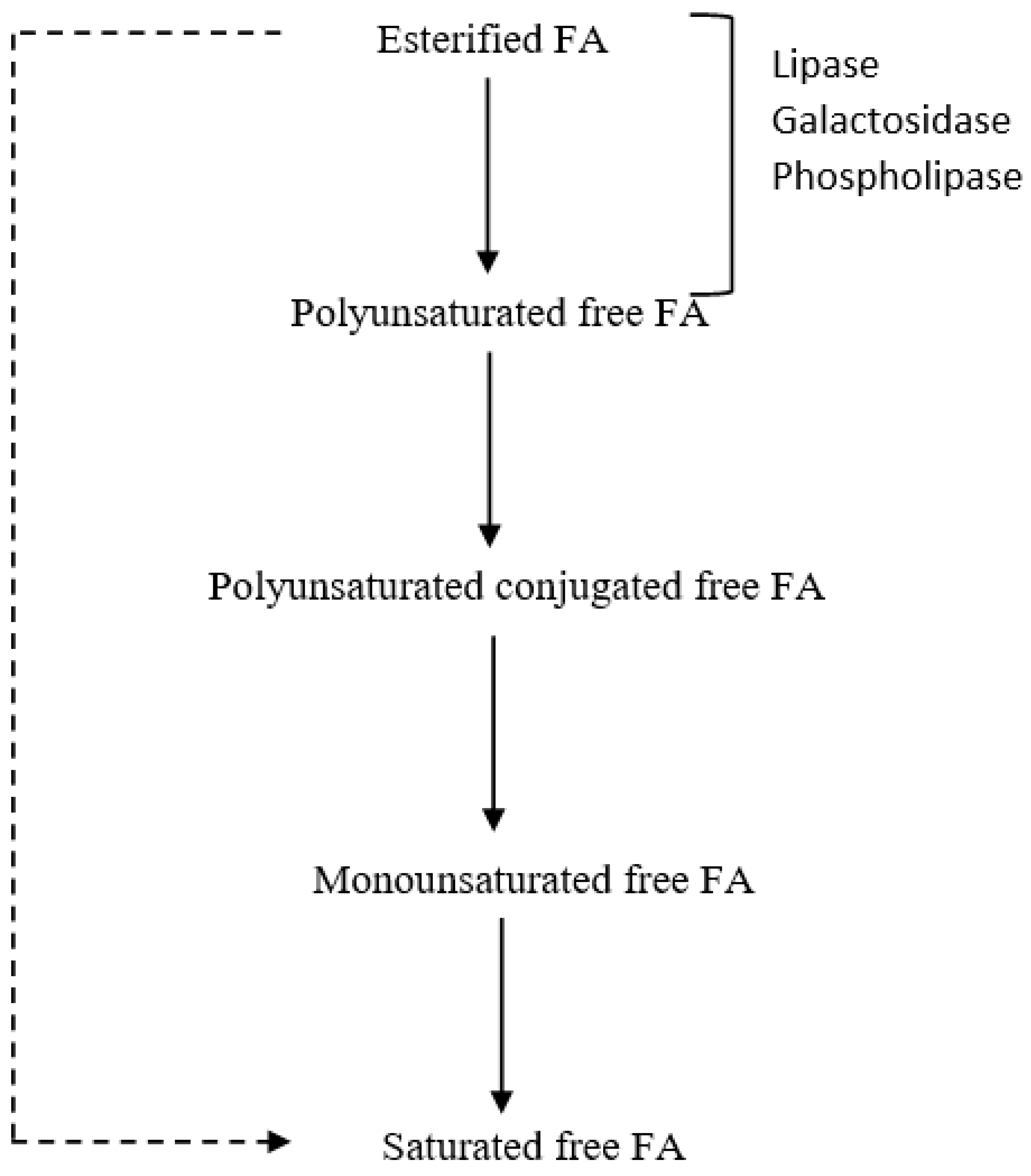
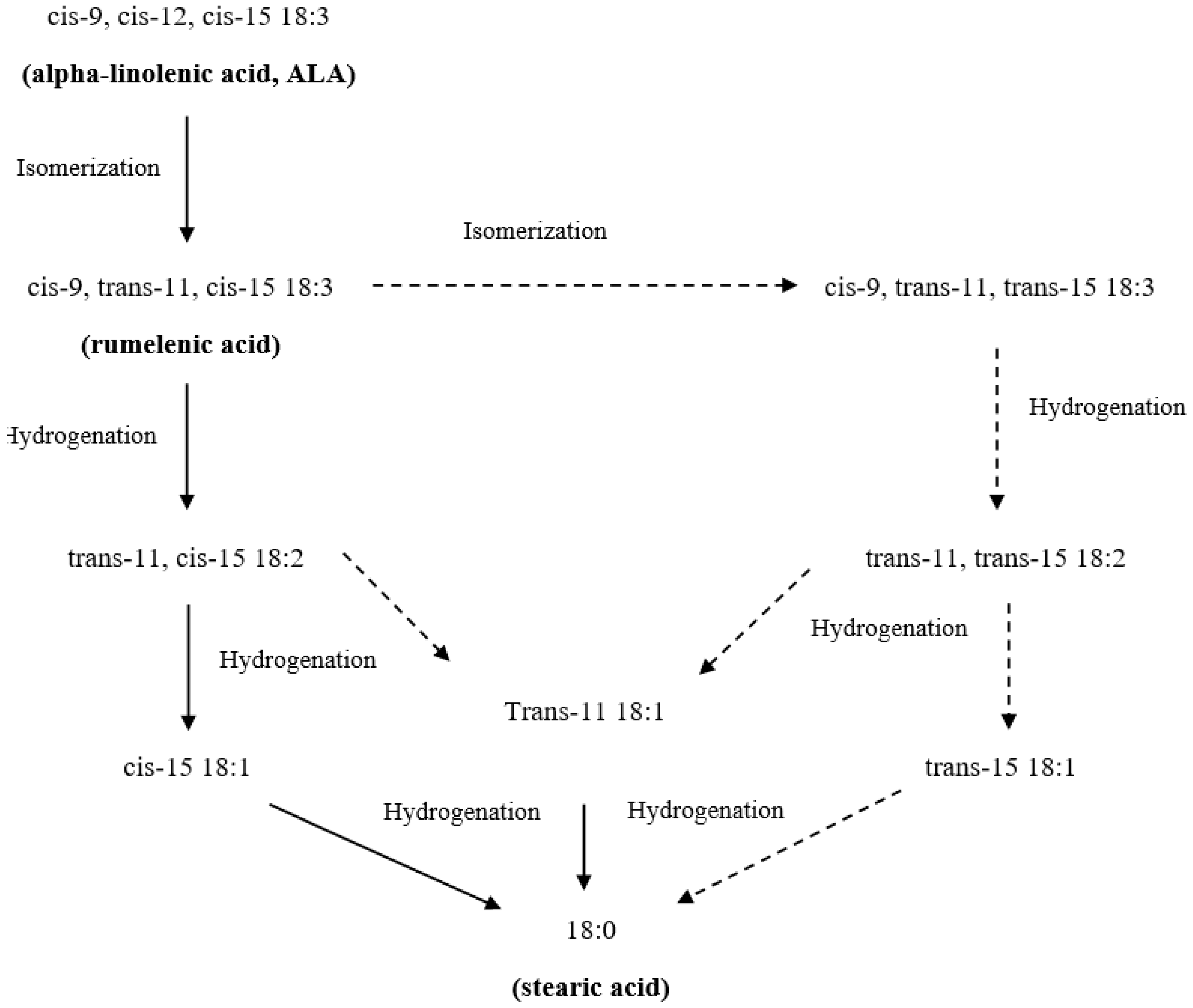
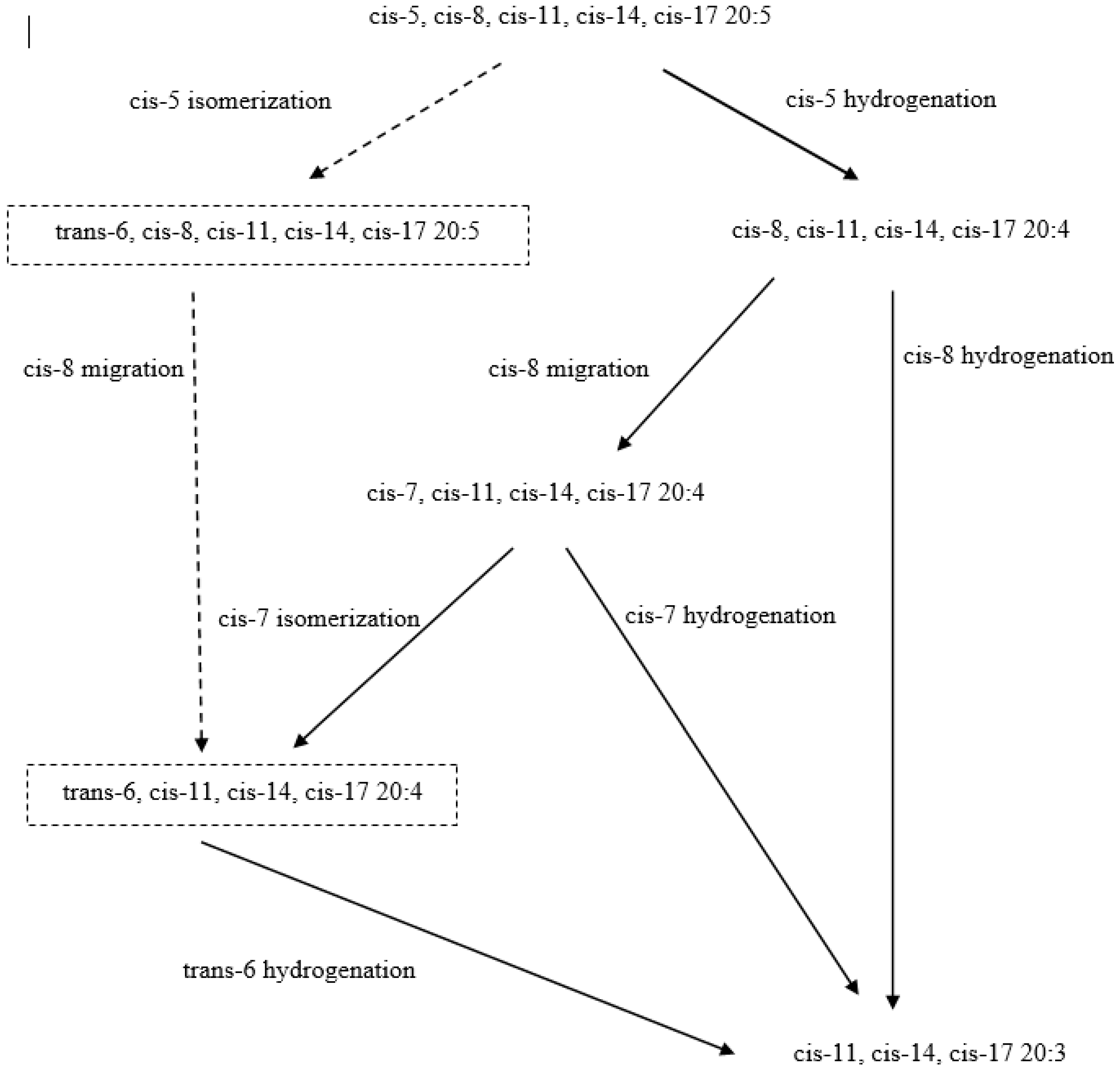
3. Lipid Metabolism in Ruminants: Obstacles to Enriching Milk Fat with n-3 PUFA
4. Recent Attempts to Increase n-3 PUFA Content In Dairy-Derived Products
4.1. Feeding Regime
4.2. Lipid Supplementation
4.2.1. Oil Seed and Vegetable Oil
4.2.2. Marine Lipid Sources
4.3. Genetic Manipulation as a Potential Tool for the Enrichment of Dairy Products with n-3 PUFA
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nichols, P.D.; Petrie, J.; Singh, S. Long-chain omega-3 oils-an update on sustainable sources. Nutrients 2010, 2, 572–585. [Google Scholar] [CrossRef]
- Bang, H.O.; Dyerberg, J.; Sinclair, H.M. The composition of the Eskimo food in north western Greenland. Am. J. Clin. Nutr. 1980, 33, 2657–2661. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Very long chain omega-3 (n-3) fatty acids and human health. Eur. J. Lipid. Sci. Technol. 2014, 116, 1280–1300. [Google Scholar] [CrossRef]
- McGuire, M.A.; McGuire, M.K. Conjugated linoleic acid (CLA): A ruminant fatty acid with beneficial effects on human health. J. Anim. Sci. 2000, 77, 1–8. [Google Scholar] [CrossRef]
- Calder, P.C. Long-chain fatty acids and inflammation. Proc. Nutr. Soc. 2012, 71, 284–289. [Google Scholar] [CrossRef]
- Calder, P.C. Omega-3 polyunsaturated fatty acids and inflammatory processes: Nutrition or pharmacology? Br. J. Clin. Pharmacol. 2013, 75, 645–662. [Google Scholar] [CrossRef]
- Belury, M.A. Inhibition of carcinogenesis by conjugated linoleic acid: Potential mechanisms of action. J. Nutr. 2002, 132, 2995–2998. [Google Scholar] [CrossRef]
- Calder, P.C. n-3 fatty acids and cardiovascular disease: Evidence explained and mechanisms explored. Clin. Sci. 2004, 107, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Calon, F.; Cole, G. Neuroprotective action of omega-3 polyunsaturated fatty acids against neurodegenerative diseases: Evidence from animal studies. Prostaglandins Leukot. Essent. Fatty Acids 2007, 77, 287–293. [Google Scholar] [CrossRef]
- Kalmijn, S.; Launer, L.J.; Ott, A.; Witteman, J.C.M.; Hofman, A.; Breteler, M.M.B. Dietary fat intake and the risk of incident dementia in the Rotterdam Study. Ann. Neurol. 1997, 42, 776–782. [Google Scholar] [CrossRef] [PubMed]
- Laugharne, J.D.E.; Mellor, J.E.; Peet, M. Fatty acids and schizophrenia. Lipids 1996, 31, 163–165. [Google Scholar] [CrossRef]
- Byelashov, O.A.; Sinclair, A.J.; Kaur, G. Dietary sources, current intakes, and nutritional role of omega-3 docosapentaenoic acid. Lipid Technol. 2015, 27, 79–82. [Google Scholar] [CrossRef] [PubMed]
- WHO. Non-Communicable Diseases Country Profiles 2018; Licence: CC BY-NC-SA 3.0 IGO; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Simopoulos, A.P. Evolutionary aspects of diet: The omega-6/omega-3 ratio and the brain. Mol. Neurobiol. 2011, 44, 203–215. [Google Scholar] [CrossRef]
- Cordain, L.; Eaton, S.B.; Sebastian, A.; Mann, N.; Lindeberg, S.; Watkins, B.A.; O’Keefe, J.H.; Brand-Miller, J. Origins and evolution of the Western diet: Health implications for the 21st century. Am. J. Clin. Nutr. 2005, 81, 341–354. [Google Scholar] [CrossRef]
- Kennedy, E.T.; Luo, H.; Ausman, L.M. Cost implications of alternative sources of (n-3) fatty acid consumption in the United States. J. Nutr. 2012, 142, 605S–609S. [Google Scholar] [CrossRef]
- Shingfield, K.J.; Bonnet, M.; Scollan, N.D. Recent developments in altering the fatty acid composition of ruminant-derived foods. Animal 2013, 7, 132–162. [Google Scholar] [CrossRef]
- Rozenberg, S.; Body, J.J.; Bruyere, O.; Bergmann, P.; Brandi, M.; Cooper, C.; Devogelaer, J.P.; Gielen, E.; Goemaere, S.; Kaufman, J.M.; et al. Effects of dairy products consumption on health: Benefits and beliefs-a commentary from the Belgian bone club and the European society for clinical and economic aspects of osteoporosis, osteoarthritis and musculoskeletal diseases. Calcif. Tissue Int. 2016, 98, 1–17. [Google Scholar] [CrossRef]
- Burgess, K. Milk and Dairy Products in Human Nutrition; Muehlhoff, E., Bennett, A., McMahon, D., Eds.; Food and Agriculture Organisation of the United Nations (FAO): Rome, Italy, 2013; Volume 67, p. 303. ISBN 978-92-5-107864-8. [Google Scholar]
- Kardas, M.; Grochowska-Niedworok, E.; Calyniuk, B.; Kolasa, I.; Grajek, M.; Bielaszka, A.; Kiciak, A.; Muc-Wierzgon, M. Consumption of milk and milk products in the population of the Upper Silesian agglomeration inhabitants. Food Nutr. Res. 2016, 60, 28976. [Google Scholar] [CrossRef]
- OECD/FAO. OECD-FAO Agricultural Outlook 2017–2026; OECD Publishing: Paris, France, 2017. [Google Scholar] [CrossRef]
- Nguyen, D.V.; Malau-Aduli, B.S.; Cavalieri, J.; Nichols, P.D.; Malau-Aduli, A.E.O. Supplementation with plant-derived oils rich in omega-3 polyunsaturated fatty acids for lamb production. Vet. Anim. Sci. 2018, 6, 29–40. [Google Scholar] [CrossRef]
- NHMRC (National Health and Medical Research Council). Nutrient Reference Values for Australia and New Zealand including Recommended Dietary Intakes; Commonwealth Department of Health and Ageing: Canberra, Australia, 2006.
- FAO/WHO 2008. Interim Summary of Conclusions and Dietary Recommendations on Total Fat & Fatty Acids; Joint FAO/WHO Expert Consultation on Fats and Fatty Acids in Human Nutrition. WHO: Geneva, Switzerland. Available online: https://www.who.int/nutrition/topics/FFA_summary_rec_conclusion.pdf?ua=1 (accessed on 25 December 2018).
- Miller, M.; Stone, N.J.; Ballantyne, C.; Bittner, V.; Criqui, M.H.; Ginsberg, H.N.; Goldberg, A.C.; Howard, W.J.; Jacobson, M.S.; Kris-Etherton, P.M.; et al. Triglycerides and cardiovascular disease: A scientific statement from the American Heart Association. Circulation 2011, 123, 2292–2333. [Google Scholar] [CrossRef]
- Lee, J.M.; Lee, H.; Kang, S.; Park, W.J. Fatty acid desaturases, polyunsaturated fatty acid regulation, and biotechnological advances. Nutrients 2016, 8, 23. [Google Scholar] [CrossRef]
- Baker, E.J.; Miles, E.A.; Burdge, G.C.; Yaqoob, P.; Calder, P.C. Metabolism and functional effects of plant-derived omega-3 fatty acids in humans. Prog. Lipid Res. 2016, 64, 30–56. [Google Scholar] [CrossRef] [PubMed]
- Office of Dietary Supplements, National Institute of Health (NIH). Omega-3 Fatty Acids. Available online: https://ods.od.nih.gov/factsheets/Omega3FattyAcids-HealthProfessional/#en77 (accessed on 21 January 2019).
- Sprecher, H. The roles of anabolic and catabolic reactions in the synthesis and recycling of polyunsaturated fatty acids. Prostaglandins Leukot. Essent. Fatty Acids 2002, 67, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Park, W.J.; Kothapalli, K.S.; Lawrence, P.; Tyburczy, C.; Brenna, J.T. An alternate pathway to long-chain polyunsaturates: The FADS2 gene product Delta8-desaturates 20:2n-6 and 20:3n-3. J. Lipid Res. 2009, 50, 1195–1202. [Google Scholar] [CrossRef]
- Park, H.G.; Park, W.J.; Kothapalli, K.S.; Brenna, J.T. The fatty acid desaturase 2 (FADS2) gene product catalyzes delta-4 desaturation to yield n-3 docosahexaenoic acid and n-6 docosapentaenoic acid in human cells. FASEB J. 2015, 29, 3911–3919. [Google Scholar] [CrossRef] [PubMed]
- Burdge, G.C.; Calder, P.C. Dietary alpha-linolenic acid and health-related outcomes: A metabolic perspective. Nutr. Res. Rev. 2006, 19, 26–52. [Google Scholar] [CrossRef]
- Brenner, R.R. Regulatory function of delta6 desaturase—Key enzyme of polyunsaturated fatty acid synthesis. Adv. Exp. Med. Biol. 1977, 83, 85–101. [Google Scholar] [PubMed]
- Narce, M.; Poisson, J.P.; Belleville, J.; Chanussot, B. Time-course effects of protein malnutrition on hepatic fatty acids Δ6 and Δ5 desaturation in the growing rat. Br. J. Nutr. 1988, 60, 389–402. [Google Scholar] [CrossRef]
- Johnson, S.B.; Kramer, T.R.; Briskeanderson, M.; Holman, R.T. Fatty acid pattern of tissue phospholipids in copper and iron deficiencies. Lipids 1989, 24, 141–145. [Google Scholar] [CrossRef]
- Simopoulos, A.P. An increase in the omega-6/omega-3 fatty acid ratio increases the risk for obesity. Nutrients 2016, 8, 128. [Google Scholar] [CrossRef]
- Tur, J.A.; Bibiloni, M.M.; Sureda, A.; Pons, A. Dietary sources of omega 3 fatty acids: Public health risks and benefits. Br. J. Nutr. 2012, 107, S23–S52. [Google Scholar] [CrossRef] [PubMed]
- Nichols, P.D.; Glencross, B.; Petrie, J.R.; Singh, S.P. Readily available sources of long-chain omega-3 oils: Is farmed Australian seafood a better source of the good oil than wild-caught seafood? Nutrients 2014, 6, 1063–1079. [Google Scholar] [CrossRef] [PubMed]
- Garcia, P.T.; Pensel, N.A.; Sancho, A.M.; Latimori, N.J.; Kloster, A.M.; Amigone, M.A.; Casal, J.J. Beef lipids in relation to animal breed and nutrition in Argentina. Meat Sci. 2008, 79, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Konieczka, P.; Czauderna, M.; Smulikowska, S. The enrichment of chicken meat with omega-3 fatty acids by dietary fish oil or its mixture with rapeseed or flaxseed—Effect of feeding duration. Anim. Feed Sci. Technol. 2017, 223, 42–52. [Google Scholar] [CrossRef]
- Dugan, M.E.; Vahmani, P.; Turner, T.D.; Mapiye, C.; Juarez, M.; Prieto, N.; Beaulieu, A.D.; Zijlstra, R.T.; Patience, J.F.; Aalhus, J.L. Pork as a source of omega-3 (n-3) fatty acids. J. Clin. Med. 2015, 4, 1999–2011. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.V.; Flakemore, A.R.; Otto, J.R.; Ives, S.W.; Smith, R.W.; Nichols, P.D.; Malau-Aduli, A.E.O. Nutritional value and sensory characteristics of meat eating quality of Australian prime lambs supplemented with pelleted canola and flaxseed oils: Fatty acid profiles of muscle and adipose tissues. Internal Med. Rev. 2017, 3, 1–21. [Google Scholar]
- Le, V.H.; Nguyen, D.V.; Nguyen, Q.V.; Malau-Aduli, B.S.; Nichols, P.D.; Malau-Aduli, A.E.O. Fatty acid profiles of muscle, liver, heart and kidney of Australian prime lambs fed different polyunsaturated fatty acids enriched pellets in a feedlot system. Sci. Rep. 2019, 9, 1238. [Google Scholar] [PubMed]
- Le, H.V.; Nguyen, Q.V.; Nguyen, D.V.; Otto, J.R.; Malau-Aduli, B.S.; Nichols, P.D.; Malau-Aduli, A.E.O. Enhanced omega-3 polyunsaturated fatty acid contents in muscle and edible organs of Australian prime lambs grazing lucerne and cocksfoot pastures. Nutrients 2018, 10, 1985. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Q.V.; Le, V.H.; Nguyen, D.V.; Malau-Aduli, B.S.; Nichols, P.D.; Malau-Aduli, A.E.O. Supplementing grazing dairy ewes with plant-derived oil and rumen-protected EPA plus DHA pellets enhances health-beneficial n-3 long-chain polyunsaturated fatty acids in sheep milk. Eur. J. Lipid Sci. Technol. 2018, 120, 256. [Google Scholar] [CrossRef]
- Nguyen, Q.V.; Van Le, H.; Nguyen, D.V.; Malau-Aduli, B.S.; Nichols, P.D.; Malau-Aduli, A.E.O. Enhancement of dairy sheep cheese eating quality with increased n-3 long-chain polyunsaturated fatty acids. J. Dairy Sci. 2019, 102, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Benbrook, C.M.; Butler, G.; Latif, M.A.; Leifert, C.; Davis, D.R. Organic production enhances milk nutritional quality by shifting fatty acid composition: A United States-wide, 18-month study. PLoS ONE 2013, 8, e82429. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Hu, X.J. Fish and its multiple human health effects in times of threat to sustainability and affordability: Are there alternatives? Asia Pac. J. Clin. Nutr. 2009, 18, 553–563. [Google Scholar] [PubMed]
- Surette, M.E. The science behind dietary omega-3 fatty acids. CMAJ 2008, 178, 177–180. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. n-3 polyunsaturated fatty acids, inflammation, and inflammatory diseases. Am. J. Clin. Nutr. 2006, 83, 1505–1519. [Google Scholar] [CrossRef]
- Kunnumakkara, A.B.; Sailo, B.L.; Banik, K.; Harsha, C.; Prasad, S.; Gupta, S.C.; Bharti, A.C.; Aggarwal, B.B. Chronic diseases, inflammation, and spices: How are they linked? J. Transl. Med. 2018, 16, 14. [Google Scholar] [CrossRef] [PubMed]
- Surette, M.E.; Koumenis, L.L.; Edens, M.B.; Tramposch, K.M.; Chilton, F.H. Inhibition of leukotriene synthesis, pharmacokinetics, and tolerability of a novel dietary fatty acid formulation in healthy adult subjects. Clin. Ther. 2003, 25, 948–971. [Google Scholar] [CrossRef]
- Zuijdgeest-Van Leeuwen, S.D.; Dagnelie, P.C.; Rietveld, T.; van den Berg, J.W.O.; Wilson, J.H.P. Incorporation and washout of orally administered n-3 fatty acid ethyl esters in different plasma lipid fractions. Br. J. Nutr. 1999, 82, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Alexander, D.D.; Miller, P.E.; Van Elswyk, M.E.; Kuratko, C.N.; Bylsma, L.C. A meta-analysis of randomized controlled trials and prospective cohort studies of eicosapentaenoic and docosahexaenoic long-chain omega-3 fatty acids and coronary heart disease risk. Mayo Clin. Proc. 2017, 92, 15–29. [Google Scholar] [CrossRef]
- Bu, J.; Dou, Y.; Tian, X.; Wang, Z.; Chen, G. The role of omega-3 polyunsaturated fatty acids in stroke. Oxid. Med. Cell. Longev. 2016, 2016, 6906712. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Benjamin, E.J.; Go, A.S.; Arnett, D.K.; Blaha, M.J.; Cushman, M.; Das, S.R.; de Ferranti, S.; Despres, J.P.; Fullerton, H.J.; et al. Heart disease and stroke statistics-2016 update: A report from the American Heart Association. Circulation 2016, 133, e38–e360. [Google Scholar] [CrossRef] [PubMed]
- Colussi, G.; Catena, C.; Novello, M.; Bertin, N.; Sechi, L.A. Impact of omega-3 polyunsaturated fatty acids on vascular function and blood pressure: Relevance for cardiovascular outcomes. Nutr. Metab. Cardiovasc. Dis. 2017, 27, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Frostegard, J. Immunity, atherosclerosis and cardiovascular disease. BMC Med. 2013, 11, 117. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.J.; Senanayake, V.K.; Pu, S.; Jenkins, D.J.; Connelly, P.W.; Lamarche, B.; Couture, P.; Charest, A.; Baril-Gravel, L.; West, S.G.; et al. DHA-enriched high-oleic acid canola oil improves lipid profile and lowers predicted cardiovascular disease risk in the canola oil multicenter randomized controlled trial. Am. J. Clin. Nutr. 2014, 100, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Shirley, R.; Ord, E.N.; Work, L.M. Oxidative stress and the use of antioxidants in stroke. Antioxidants 2014, 3, 472–501. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Tang, H.; Yang, X.D.; Luo, X.Q.; Wang, X.Y.; Shao, C.; He, J.Q. Fish consumption and stroke risk: A meta-Analysis of prospective cohort studies. J. Stroke Cerebrovasc. Dis. 2019, 28, 604–611. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Ambigaipalan, P. Omega-3 polyunsaturated fatty acids and their health benefits. Annu. Rev. Food Sci. Technol. 2018, 9, 345–381. [Google Scholar] [CrossRef] [PubMed]
- Kato, I.; Akhmedkhanov, A.; Koenig, K.; Toniolo, P.G.; Shore, R.E.; Riboli, E. Prospective study of diet and female colorectal cancer: The New York University Women’s Health Study. Nutr. Cancer 1997, 28, 276–281. [Google Scholar] [CrossRef]
- Terry, P.; Lichtenstein, P.; Feychting, M.; Ahlbom, A.; Wolk, A. Fatty fish consumption and risk of prostate cancer. Lancet 2001, 357, 1764–1766. [Google Scholar] [CrossRef]
- Takezaki, T.; Inoue, M.; Kataoka, H.; Ikeda, S.; Yoshida, M.; Ohashi, Y.; Tajima, K.; Tominaga, S. Diet and lung cancer risk from a 14-year population-based prospective study in Japan: With special reference to fish consumption. Nutr. Cancer 2003, 45, 160–167. [Google Scholar] [CrossRef]
- Van Blarigan, E.; Fuchs, C.S.; Niedzwiecki, D.; Ye, X.; Zhang, S.; Song, M.Y. Long-chain omega-3 fatty acid and fish intake after colon cancer diagnosis and disease-free, recurrence-free, and overall survival in CALGB 89803 (Alliance). J. Clin. Oncol. 2017, 35, 1227–1230. [Google Scholar] [CrossRef]
- Makarem, N.; Chandran, U.; Bandera, E.V.; Parekh, N. Dietary Fat in Breast Cancer Survival. Annu. Rev. Nutr. 2013, 33, 319–348. [Google Scholar] [CrossRef] [PubMed]
- Sczaniecka, A.K.; Brasky, T.M.; Lampe, J.W.; Patterson, R.E.; White, E. Dietary intake of specific fatty acids and breast cancer risk among postmenopausal women in the VITAL cohort. Nutr. Cancer 2012, 64, 1131–1142. [Google Scholar] [CrossRef] [PubMed]
- Pereira, P.; Reis, A.D.; Diniz, R.R.; Lima, F.A.; Leite, R.D.; da Silva, M.C.P.; Guerra, R.N.M.; Vieira, E.B.D.; Garcia, J.B.S. Dietary supplements and fatigue in patients with breast cancer: A systematic review. Breast Cancer Res. Treat. 2018, 171, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Rizos, E.C.; Ntzani, E.E.; Bika, E.; Kostapanos, M.S.; Elisaf, M.S. Association between omega-3 fatty acid supplementation and risk of major cardiovascular disease events: A systematic review and meta-analysis. JAMA 2012, 308, 1024–1033. [Google Scholar] [CrossRef]
- Rhee, J.J.; Kim, E.; Buring, J.E.; Kurth, T. Fish consumption, omega-3 fatty acids, and risk of cardiovascular disease. Am. J. Prev. Med. 2017, 52, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Holmes, M.D.; Colditz, G.A.; Hunter, D.J.; Hankinson, S.E.; Rosner, B.; Speizer, F.E.; Willett, W.C. Meat, fish and egg intake and risk of breast cancer. Int. J. Cancer 2003, 104, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Park, M.K.; Kyung, W.Q.; Li, T.; Qureshi, A.A.; Cho, E. Prospective Evaluation of Fat Intake and Risk of Skin Cancer. FASEB J. 2017, 31, 168–173. [Google Scholar]
- Salem, N.; Litman, B.; Kim, H.Y.; Gawrisch, K. Mechanisms of action of docosahexaenoic acid in the nervous system. Lipids 2001, 36, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Dyall, S.C. Long-chain omega-3 fatty acids and the brain: A review of the independent and shared effects of EPA, DPA and DHA. Front. Aging Neurosci. 2015, 7, 52. [Google Scholar] [CrossRef] [PubMed]
- Cole, G.M.; Ma, Q.L.; Frautschy, S.A. Omega-3 fatty acids and dementia. Prostaglandins Leukot. Essent. Fatty Acids 2009, 81, 213–221. [Google Scholar] [CrossRef]
- Da Silva, T.M.; Munhoz, R.P.; Alvarez, C.; Naliwaiko, K.; Kiss, A.; Andreatini, R.; Ferraz, A.C. Depression in Parkinson’s disease: A double-blind, randomized, placebo-controlled pilot study of omega-3 fatty-acid supplementation. J. Affect. Disord. 2008, 111, 351–359. [Google Scholar] [CrossRef]
- Abdulrazaq, M.; Innes, J.K.; Calder, P.C. Effect of omega-3 polyunsaturated fatty acids on arthritic pain: A systematic review. Nutrition 2017, 39–40, 57–66. [Google Scholar] [CrossRef]
- Tsitouras, P.D.; Gucciardo, F.; Salbe, A.D.; Heward, C.; Harman, S.M. High omega-3 fat intake improves insulin sensitivity and reduces CRP and IL6, but does not affect other endocrine axes in healthy older adults. Horm. Metab. Res. 2008, 40, 199–205. [Google Scholar] [CrossRef]
- Wang, L.; Folsom, A.R.; Zheng, Z.J.; Pankow, J.S.; Eckfeldt, J.H.; Investigators, A.S. Plasma fatty acid composition and incidence of diabetes in middle-aged adults: The atherosclerosis risk in communities (ARIC) study. Am. J. Clin. Nutr. 2003, 78, 91–98. [Google Scholar]
- Wu, J.H.; Micha, R.; Imamura, F.; Pan, A.; Biggs, M.L.; Ajaz, O.; Djousse, L.; Hu, F.B.; Mozaffarian, D. Omega-3 fatty acids and incident type 2 diabetes: A systematic review and meta-analysis. Br. J. Nutr. 2012, 107, 214–227. [Google Scholar] [CrossRef] [PubMed]
- Albracht-Schulte, K.; Kalupahana, N.S.; Ramalingam, L.; Wang, S.; Rahman, S.M.; Robert-McComb, J.; Moustaid-Moussa, N. Omega-3 fatty acids in obesity and metabolic syndrome: A mechanistic update. J. Nutr. Biochem. 2018, 58, 1–16. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Rimm, E.B.; Merchant, A.; Rosner, B.A.; Stampfer, M.J.; Willett, W.C.; Ascherio, A. Fish consumption and risk of stroke in men. JAMA 2002, 288, 3130–3136. [Google Scholar] [CrossRef]
- Iso, H.; Rexrode, K.M.; Stampfer, M.J.; Manson, J.E.; Colditz, G.A.; Speizer, F.E.; Hennekens, C.H.; Willett, W.C. Intake of fish and omega-3 fatty acids and risk of stroke in women. JAMA 2001, 285, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Thota, R.N.; Ferguson, J.J.A.; Abbott, K.A.; Dias, C.B.; Garg, M.L. Science behind the cardio-metabolic benefits of omega-3 polyunsaturated fatty acids: Biochemical effects vs. clinical outcomes. Food Funct. 2018, 9, 3576–3596. [Google Scholar] [CrossRef]
- O’Connell, T.D.; Block, R.C.; Huang, S.P.; Shearer, G.C. ω3-Polyunsaturated fatty acids for heart failure: Effects of dose on efficacy and novel signaling through free fatty acid receptor 4. J. Mol. Cell Cardiol. 2017, 103, 74–92. [Google Scholar] [CrossRef]
- Chilliard, Y.; Glasser, F.; Ferlay, A.; Bernard, L.; Rouel, J.; Doreau, M. Diet, rumen biohydrogenation and nutritional quality of cow and goat milk fat. Eur. J. Lipid. Sci. Technol. 2007, 109, 828–855. [Google Scholar] [CrossRef]
- Buccioni, A.; Decandia, M.; Minieri, S.; Molle, G.; Cabiddu, A. Lipid metabolism in the rumen: New insights on lipolysis and biohydrogenation with an emphasis on the role of endogenous plant factors. Anim. Feed Sci. Technol. 2012, 174, 1–25. [Google Scholar] [CrossRef]
- Harfoot, C.G.; Hazlewood, G.P. Lipid metabolism in the rumen. In The Rumen Microbial Ecosystem; Hobson, P.N., Ed.; Elsevier Applied Science Publishers: London, UK, 1988; pp. 285–322. [Google Scholar]
- Bauman, D.E.; Lock, A.L. Concepts in lipid digestion and metabolism in dairy cows. In Proceedings of the Tri-State Dairy Nutrition Conference, Fort Wayne, IN, USA, 25–26 April 2006; Eastridge, M.L., Ed.; pp. 1–14. [Google Scholar]
- Shingfield, K.J.; Bernard, L.; Leroux, C.; Chilliard, Y. Role of trans fatty acids in the nutritional regulation of mammary lipogenesis in ruminants. Animal 2010, 4, 1140–1166. [Google Scholar] [CrossRef] [PubMed]
- Shingfield, K.J.; Kairenius, P.; Arola, A.; Paillard, D.; Muetzel, S.; Ahvenjarvi, S.; Vanhatalo, A.; Huhtanen, P.; Toivonen, V.; Griinari, J.M.; et al. Dietary fish oil supplements modify ruminal biohydrogenation, alter the flow of fatty acids at the omasum, and induce changes in the ruminal Butyrivibrio population in lactating cows. J. Nutr. 2012, 142, 1437–1448. [Google Scholar] [CrossRef] [PubMed]
- Kairenius, P.; Toivonen, V.; Shingfield, K.J. Identification and ruminal outflow of long-chain fatty acid biohydrogenation intermediates in cows fed diets containing fish oil. Lipids 2011, 46, 587–606. [Google Scholar] [CrossRef]
- Chilliard, Y.; Ferlay, A.; Mansbridge, R.M.; Doreau, M. Ruminant milk fat plasticity: Nutritional control of saturated, polyunsaturated, trans and conjugated fatty acids. Ann. Zootech. 2000, 49, 181–205. [Google Scholar] [CrossRef]
- Toral, P.G.; Hervas, G.; Leskinen, H.; Shingfield, K.J.; Frutos, P. In vitro ruminal biohydrogenation of eicosapentaenoic (EPA), docosapentaenoic (DPA), and docosahexaenoic acid (DHA) in cows and ewes: Intermediate metabolites and pathways. J. Dairy Sci. 2018, 101, 6109–6121. [Google Scholar] [CrossRef] [PubMed]
- Manso, T.; Gallardo, B.; Guerra-Rivas, C. Modifying milk and meat fat quality through feed changes. Small Ruminant Res. 2016, 142, 31–37. [Google Scholar] [CrossRef]
- Bisig, W.; Eberhard, P.; Collomb, M.; Rehberger, B. Influence of processing on the fatty acid composition and the content of conjugated linoleic acid in organic and conventional dairy products—A review. Lait 2007, 87, 1–19. [Google Scholar] [CrossRef]
- Collomb, M.; Schmid, A.; Sieber, R.; Wechsler, D.; Ryhanen, E.L. Conjugated linoleic acids in milk fat: Variation and physiological effects. Int. Dairy J. 2006, 16, 1347–1361. [Google Scholar] [CrossRef]
- Prandini, A.; Sigolo, S.; Piva, G. A comparative study of fatty acid composition and CLA concentration in commercial cheeses. J. Food Compos. Anal. 2011, 24, 55–61. [Google Scholar] [CrossRef]
- Dewhurst, R.J.; Shingfield, K.J.; Lee, M.R.F.; Scollan, N.D. Increasing the concentrations of beneficial polyunsaturated fatty acids in milk produced by dairy cows in high-forage systems. Anim. Feed Sci. Technol. 2006, 131, 168–206. [Google Scholar] [CrossRef]
- Woods, V.B.; Fearon, A.M. Dietary sources of unsaturated fatty acids for animals and their transfer into meat, milk and eggs: A review. Livest. Sci. 2009, 126, 1–20. [Google Scholar] [CrossRef]
- Gilliland, T.J.; Barrett, P.D.; Mann, R.L.; Agnew, R.E.; Fearon, A.M. Canopy morphology and nutritional quality traits as potential grazing value indicators for Lolium perenne varieties. J. Agric. Sci. 2002, 139, 257–273. [Google Scholar] [CrossRef]
- Elgersma, A.; Ellen, G.; van der Horst, H.; Muuse, B.G.; Boer, H.; Tamminga, S. Comparison of the fatty acid composition of fresh and ensiled perennial ryegrass (Lolium perenne L.), affected by cultivar and regrowth interval. Anim. Feed Sci. Technol. 2003, 108, 191–205. [Google Scholar] [CrossRef]
- Gomez-Cortes, P.; Frutos, P.; Mantecon, A.R.; Juarez, M.; de la Fuente, M.A.; Hervas, G. Effect of supplementation of grazing dairy ewes with a cereal concentrate on animal performance and milk fatty acid profile. J. Dairy Sci. 2009, 92, 3964–3972. [Google Scholar] [CrossRef]
- Leiber, F.; Kreuzer, M.; Nigg, D.; Wettstein, H.R.; Scheeder, M.R.L. A study on the causes for the elevated n-3 fatty acids in cows’ milk of alpine origin. Lipids 2005, 40, 191–202. [Google Scholar] [CrossRef]
- Mierlita, D. Fatty acid profile and health lipid indices in the raw milk of ewes grazing part-time and hemp seed supplementation of lactating ewes. S. Afr. J. Anim. Sci. 2016, 46, 237–246. [Google Scholar]
- Mohammed, R.; Stanton, C.S.; Kennelly, J.J.; Kramer, J.K.; Mee, J.F.; Glimm, D.R.; O’Donovan, M.; Murphy, J.J. Grazing cows are more efficient than zero-grazed and grass silage-fed cows in milk rumenic acid production. J. Dairy Sci. 2009, 92, 3874–3893. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Cortes, P.; Tyburczy, C.; Brenna, J.T.; Juarez, M.; de la Fuente, M.A. Characterization of cis-9 trans-11 trans-15 C18:3 in milk fat by GC and covalent adduct chemical ionization tandem MS. J. Lipid Res. 2009, 50, 2412–2420. [Google Scholar] [CrossRef] [PubMed]
- Addis, M.; Cabiddu, A.; Pinna, G.; Decandia, M.; Piredda, G.; Pirisi, A.; Molle, G. Milk and cheese fatty acid composition in sheep fed Mediterranean forages with reference to conjugated linoleic acid cis-9, trans-11. J. Dairy Sci. 2005, 88, 3443–3454. [Google Scholar] [CrossRef]
- Bonanno, A.; Di Grigoli, A.; Mazza, F.; De Pasquale, C.; Giosue, C.; Vitale, F.; Alabiso, M. Effects of ewes grazing sulla or ryegrass pasture for different daily durations on forage intake, milk production and fatty acid composition of cheese. Animal 2016, 10, 2074–2082. [Google Scholar] [CrossRef] [PubMed]
- Guzatti, G.C.; Duchini, P.G.; Gama, M.A.S.; Ribeiro, H.M.N. Red clover silage improves milk fatty acid composition in dairy ewes. Can. J. Anim. Sci. 2018, 98, 787–793. [Google Scholar] [CrossRef]
- Cabiddu, A.; Molle, G.; Decandia, M.; Spada, S.; Fiori, M.; Piredda, G.; Addis, M. Responses to condensed tannins of flowering sulla (Hedysarum coronarium L.) grazed by dairy sheep Part 2: Effects on milk fatty acid profile. Livest. Sci. 2009, 123, 230–240. [Google Scholar] [CrossRef]
- Lourenco, M.; Ramos-Morales, E.; Wallace, R.J. The role of microbes in rumen lipolysis and biohydrogenation and their manipulation. Animal 2010, 4, 1008–1023. [Google Scholar] [CrossRef]
- Hristov, A.N.; Price, W.J.; Shafii, B. A meta-analysis examining the relationship among dietary factors, dry matter intake, and milk and milk protein yield in dairy cows. J. Dairy Sci. 2004, 87, 2184–2196. [Google Scholar] [CrossRef]
- Kennelly, J.J.; Bell, J.A.; Keating, A.F.; Doepel, L. Nutrition as a tool to alter milk composition. Adv. Dairy Tech. 2005, 17, 255–275. [Google Scholar]
- Dubois, V.; Breton, S.; Linder, M.; Fanni, J.; Parmentier, M. Fatty acid profiles of 80 vegetable oils with regard to their nutritional potential. Eur. J. Lipid Sci. Technol. 2007, 109, 710–732. [Google Scholar] [CrossRef]
- Petit, H.V. Review: Feed intake, milk production and milk composition of dairy cows fed flaxseed. Can. J. Anim. Sci. 2010, 90, 115–127. [Google Scholar] [CrossRef]
- Bernacchia, R.; Preti, R.; Vinci, G. Chemical composition and health benefits of flaxseed. Austin J. Nutr. Food Sci. 2014, 2, 1045–1053. [Google Scholar]
- Coppa, M.; Ferlay, A.; Monsallier, F.; Verdier-Metz, I.; Pradel, P.; Didienne, R.; Farruggia, A.; Montel, M.C.; Martin, B. Milk fatty acid composition and cheese texture and appearance from cows fed hay or different grazing systems on upland pastures. J. Dairy Sci. 2011, 94, 1132–1145. [Google Scholar] [CrossRef]
- Stergiadis, S.; Leifert, C.; Seal, C.J.; Eyre, M.D.; Steinshamn, H.; Butler, G. Improving the fatty acid profile of winter milk from housed cows with contrasting feeding regimes by oilseed supplementation. Food Chem. 2014, 164, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zhang, R.; Kang, R.; Meng, J.; Ao, C. Milk fatty acids profiles and milk production from dairy cows fed different forage quality diets. Anim. Nutr. 2016, 2, 329–333. [Google Scholar] [CrossRef]
- Mierlita, D.; Pop, I.M.; Lup, F.; Simeanu, D.; Vicas, S.I.; Simeanu, C. The fatty acid composition and health lipid indices in sheep raw milk under a pasture-based dairy system. Revista de Chimie 2018, 69, 160–165. [Google Scholar]
- Mohamed, B.; Sahnoun, A.; Bouderoua, K.; Homrani, A. Diet of ewes improve the fatty acid composition of milk. Adv. Biores. 2018, 9, 18–25. [Google Scholar]
- Khas, E.; Wang, J.Q.; Bu, D.P.; Wang, L.; Drackley, J.K.; Liu, Q.S.; Yang, G.; Wei, H.Y.; Zhou, L.Y. Short communication: Responses to increasing amounts of free alpha-linolenic acid infused into the duodenum of lactating dairy cows. J. Dairy Sci. 2010, 93, 1677–1684. [Google Scholar] [CrossRef] [PubMed]
- Kitessa, S.M.; Gulati, S.K.; Ashes, J.R.; Fleck, E.; Scott, T.W.; Nichols, P.D. Utilisation of fish oil in ruminants—I. Fish oil metabolism in sheep. Anim. Feed Sci. Technol. 2001, 89, 189–199. [Google Scholar] [CrossRef]
- Kitessa, S.M.; Gulati, S.K.; Simos, G.C.; Ashes, J.R.; Scott, T.W.; Fleck, E.; Wynn, P.C. Supplementation of grazing dairy cows with rumen-protected tuna oil enriches milk fat with n-3 fatty acids without affecting milk production or sensory characteristics. Br. J. Nutr. 2004, 91, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Kitessa, S.M.; Peake, D.; Bencini, R.; Williams, A.J. Fish oil metabolism in ruminants. Anim. Feed Sci. Technol. 2003, 108, 1–14. [Google Scholar] [CrossRef]
- Papadopoulos, G.; Goulas, C.; Apostolaki, E.; Abril, R. Effects of dietary supplements of algae, containing polyunsaturated fatty acids, on milk yield and the composition of milk products in dairy ewes. J. Dairy Res. 2002, 69, 357–365. [Google Scholar] [CrossRef]
- Boeckaert, C.; Vlaeminck, B.; Dijkstra, J.; Issa-Zacharia, A.; Van Nespen, T.; Van Straalen, W.; Fievez, V. Effect of dietary starch or micro algae supplementation on rumen fermentation and milk fatty acid composition of dairy cows. J. Dairy Sci. 2008, 91, 4714–4727. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.W.; Juarez, M.; Ramos, M.; Haenlein, G.F.W. Physico-chemical characteristics of goat and sheep milk. Small Ruminant Res. 2007, 68, 88–113. [Google Scholar] [CrossRef]
- Flakemore, A.R.; Malau-Aduli, B.S.; Nichols, P.D.; Malau-Aduli, A.E.O. Omega-3 fatty acids, nutrient retention values, and sensory meat eating quality in cooked and raw Australian lamb. Meat Sci. 2017, 123, 79–87. [Google Scholar] [CrossRef]
- FSANZ (Food Standards Australia New Zealand). Standard 1.2.8: Nutrition Information Requirements; Australia New Zealand Food Standards Code; FSANZ: Canberra, Australia, 2002; Volume 2.
- Watkins, P.J.; Frank, D.; Singh, T.K.; Young, O.A.; Warner, R.D. Sheep meat flavor and the effect of different feeding systems: A review. J. Agric. Food Chem. 2013, 61, 3561–3579. [Google Scholar] [CrossRef] [PubMed]
- Caroprese, M.; Marzano, A.; Marino, R.; Gliatta, G.; Muscio, A.; Sevi, A. Flaxseed supplementation improves fatty acid profile of cow milk. J. Dairy Sci. 2010, 93, 2580–2588. [Google Scholar] [CrossRef]
- Dai, X.J.; Wang, C.; Zhu, Q. Milk performance of dairy cows supplemented with rapeseed oil, peanut oil and sunflower seed oil. Czech J. Anim. Sci. 2011, 56, 181–191. [Google Scholar] [CrossRef]
- Otto, J.R.; Malau-Aduli, B.S.; Nichols, P.D.; Malau-Aduli, A.E.O. Influence of supplementing pasture-based primiparous Holstein-Friesian dairy cows with crude degummed canola oil on milk fatty acid composition. J. Nutr. Ther. 2014, 3, 55–66. [Google Scholar]
- Cattani, M.; Mantovani, R.; Schiavon, S.; Bittante, G.; Bailoni, L. Recovery of n-3 polyunsaturated fatty acids and conjugated linoleic acids in ripened cheese obtained from milk of cows fed different levels of extruded flaxseed. J. Dairy Sci. 2014, 97, 123–135. [Google Scholar] [CrossRef]
- Li, R.; Beaudoin, F.; Ammah, A.A.; Bissonnette, N.; Benchaar, C.; Zhao, X.; Lei, C.Z.; Ibeagha-Awemu, E.M. Deep sequencing shows microRNA involvement in bovine mammary gland adaptation to diets supplemented with linseed oil or safflower oil. BMC Genom. 2015, 16, 884. [Google Scholar] [CrossRef] [PubMed]
- Welter, K.C.; Martins, C.M.; de Palma, A.S.; Martins, M.M.; Dos Reis, B.R.; Schmidt, B.L.; Saran Netto, A. Canola oil in lactating dairy cow diets reduces milk saturated fatty acids and improves its omega-3 and oleic fatty acid content. PLoS ONE 2016, 11, e0151876. [Google Scholar] [CrossRef]
- Vanbergue, E.; Peyraud, J.L.; Hurtaud, C. Effects of new n-3 fatty acid sources on milk fatty acid profile and milk fat properties in dairy cows. J. Dairy Res. 2018, 85, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Bodas, R.; Manso, T.; Mantecon, A.R.; Juarez, M.; De la Fuente, M.A.; Gomez-Cortes, P. Comparison of the fatty acid profiles in cheeses from ewes fed diets supplemented with different plant oils. J. Agric. Food Chem. 2010, 58, 10493–10502. [Google Scholar] [CrossRef]
- Mughetti, L.; Sinesio, F.; Acuti, G.; Antonini, C.; Moneta, E.; Peparaio, M.; Trabalza-Marinucci, M. Integration of extruded linseed into dairy sheep diets: Effects on milk composition and quality and sensorial properties of Pecorino cheese. Anim. Feed Sci. Technol. 2012, 178, 27–39. [Google Scholar] [CrossRef]
- Caroprese, M.; Ciliberti, M.G.; Marino, R.; Santillo, A.; Sevi, A.; Albenzio, M. Polyunsaturated fatty acid supplementation: Effects of seaweed ascophyllum nodosum and flaxseed on milk production and fatty acid profile of lactating ewes during summer. J. Dairy Res. 2016, 83, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Parentet, M.D.M.; Susin, I.; Nolli, C.P.; Ferreira, E.M.; Gentil, R.S.; Polizel, D.M.; Pires, A.V.; Alves, S.P.; Bessa, R.J.B. Effects of supplementation with vegetable oils, including castor oil, on milk production of ewes and on growth of their lambs. J. Anim. Sci. 2018, 96, 354–363. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, A.E.; Da Silva, A.S.; Biazus, A.H.; Richards, N.; Pellegrini, L.G.; Baldissera, M.D.; Macedo, V.P.; Da Silveira, A.L.F. Adding palm oil to the diet of sheep alters fatty acids profile on yogurt: Benefits to consumers. Anais da Academia Brasileira de Ciencias 2017, 89, 2471–2478. [Google Scholar] [CrossRef]
- Soyeurt, H.; Gillon, A.; Vanderick, S.; Mayeres, P.; Bertozzi, C.; Gengler, N. Estimation of heritability and genetic correlations for the major fatty acids in bovine milk. J. Dairy Sci. 2007, 90, 4435–4442. [Google Scholar] [CrossRef]
- Stoop, W.M.; van Arendonk, J.A.; Heck, J.M.; van Valenberg, H.J.; Bovenhuis, H. Genetic parameters for major milk fatty acids and milk production traits of Dutch Holstein-Friesians. J. Dairy Sci. 2008, 91, 385–394. [Google Scholar] [CrossRef]
- Bilal, G.; Cue, R.I.; Mustafa, A.F.; Hayes, J.F. Short communication: Genetic parameters of individual fatty acids in milk of Canadian Holsteins. J. Dairy Sci. 2014, 97, 1150–1156. [Google Scholar] [CrossRef] [PubMed]
- Pegolo, S.; Cecchinato, A.; Casellas, J.; Conte, G.; Mele, M.; Schiavon, S.; Bittante, G. Genetic and environmental relationships of detailed milk fatty acids profile determined by gas chromatography in Brown Swiss cows. J. Dairy Sci. 2016, 99, 1315–1330. [Google Scholar] [CrossRef]
- Correddu, F.; Cellesi, M.; Serdino, J.; Manca, M.G.; Contu, M.; Dimauro, C.; Ibba, I.; Macciotta, N.P.P. Genetic parameters of milk fatty acid profile in sheep: Comparison between gas chromatographic measurements and Fourier-transform IR spectroscopy predictions. Animal 2018, 13, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Boichard, D.; Govignon-gion, A.; Larroque, H.; Maroteau, C.; Palhière, I.; Tosser-Klopp, G.; Rupp, R.; Sanchez, M.-P.; Brochard, M. Déterminisme génétique de la composition en acides gras et protéines du lait des ruminants, et potentialités de sélection. INRA Prod. Anim. 2014, 27, 283–289. [Google Scholar]
- Maroteau, C.; Palhiere, I.; Larroque, H.; Clement, V.; Ferrand, M.; Tosser-Klopp, G.; Rupp, R. Genetic parameter estimation for major milk fatty acids in Alpine and Saanen primiparous goats. J. Dairy Sci. 2014, 97, 3142–3155. [Google Scholar] [CrossRef]
- Marquardt, A.; Stohr, H.; White, K.; Weber, B.H.F. cDNA cloning, genomic structure, and chromosomal localization of three members of the human fatty acid desaturase family. Genomics 2000, 66, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Malau-Aduli, A.E.O.; Bignell, C.W.; McCulloch, R.; Kijas, J.W.; Nichols, P.D. Genetic association of delta-six fatty acid desaturase (FADS) single nucleotide polymorphic molecular marker and muscle long chain omega-3 fatty acids in Australian lamb. In Global Challenges to Production, Processing and Consumption of Meat, Proceedings of the 57th International Congress of Meat Science and Technology, Ghent, Belgium, 7–12 August 2011; De Smet, S., Ed.; University of Ghent: Ghent, Belgium, 2011; Volume 57, p. 126. [Google Scholar]
- Malau-Aduli, A.E.O.; Otto, J.R.; Suybeng, B.; Kashani, A.; Lane, P.A.; Malau-Aduli, B.S.; Nichols, P.D. Gene expression profiles of Aralkylamine N-acetyltransferase, B-cell translocation gene-2 and Fatty Acid Synthase genes in pasture-based primiparous Holstein-Friesian dairy cows supplemented with crude degummed canola oil. Adv. Genet. Eng. 2015, 4, 1000123. [Google Scholar] [CrossRef]
- Malau-Aduli, A.E.O.; Kashani, A. Molecular genetics-nutrition interactions in the expression of AANAT, ADRB3, BTG2 and FASN genes in the heart, kidney and liver of Australian lambs supplemented with Spirulina (Arthrospira platensis). Genes Genom. 2015, 37, 633–644. [Google Scholar] [CrossRef]
- Malau-Aduli, A.E.O.; Holman, B.W.B.; Kashani, A.; Nichols, P.D. Sire breed and sex effects on the fatty acid composition and content of heart, kidney, liver, adipose and muscle tissues of purebred and first-cross prime lambs. Anim. Prod. Sci. 2016, 56, 2122–2132. [Google Scholar] [CrossRef]
- Malau-Aduli, A.E.O.; Nguyen, D.V.; Le, V.H.; Nguyen, Q.V.; Otto, J.R.; Malau-Aduli, B.S.; Nichols, P.D. Correlations between growth and wool quality traits of genetically divergent Australian lambs in response to canola or flaxseed oil supplementation. PLoS ONE 2019, 14, e0208229. [Google Scholar] [CrossRef]
- Ibeagha-Awemu, E.M.; Akwanji, K.A.; Beaudoin, F.; Zhao, X. Associations between variants of FADS genes and omega-3 and omega-6 milk fatty acids of Canadian Holstein cows. BMC Genet. 2014, 15, 25. [Google Scholar] [CrossRef]
- Ibeagha-Awemu, E.M.; Peters, S.O.; Akwanji, K.A.; Imumorin, I.G.; Zhao, X. High density genome wide genotyping-by-sequencing and association identifies common and low frequency SNPs, and novel candidate genes influencing cow milk traits. Sci. Rep. 2016, 6, 31109. [Google Scholar] [CrossRef]
- Franklin, S.T.; Martin, K.R.; Baer, R.J.; Schingoethe, D.J.; Hippen, A.R. Dietary marine algae (Schizochytrium sp.) increases concentrations of conjugated linoleic, docosahexaenoic and trans vaccenic acids in milk of dairy cows. J. Nutr. 1999, 129, 2048–2054. [Google Scholar] [CrossRef] [PubMed]
- Shingfield, K.J.; Reynolds, C.K.; Hervas, G.; Griinari, J.M.; Grandison, A.S.; Beever, D.E. Examination of the persistency of milk fatty acid composition responses to fish oil and sunflower oil in the diet of dairy cows. J. Dairy Sci. 2006, 89, 714–732. [Google Scholar] [CrossRef]
- Kazama, R.; Cortes, C.; da Silva-Kazama, D.; Gagnon, N.; Benchaar, C.; Zeoula, L.M.; Santos, G.T.D.; Petit, H.V. Abomasal or ruminal administration of flax oil and hulls on milk production, digestibility, and milk fatty acid profile of dairy cows. J. Dairy Sci. 2010, 93, 4781–4790. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Bello-Pérez, E.; Íñiguez-González, G.; Fehrmann-Cartes, K.; Toro-Mujica, P.; Garnsworthy, P.C. Influence of fish oil alone or in combination with hydrogenated palm oil on sensory characteristics and fatty acid composition of bovine cheese. Anim. Feed Sci. Technol. 2015, 205, 60–68. [Google Scholar] [CrossRef]
- Kairenius, P.; Arola, A.; Leskinen, H.; Toivonen, V.; Ahvenjarvi, S.; Vanhatalo, A.; Huhtanen, P.; Hurme, T.; Griinari, J.M.; Shingfield, K.J. Dietary fish oil supplements depress milk fat yield and alter milk fatty acid composition in lactating cows fed grass silage-based diets. J. Dairy Sci. 2015, 98, 5653–5671. [Google Scholar] [CrossRef] [PubMed]
- Toral, P.G.; Hervas, G.; Gomez-Cortes, P.; Frutos, P.; Juarez, M.; de la Fuente, M.A. Milk fatty acid profile and dairy sheep performance in response to diet supplementation with sunflower oil plus incremental levels of marine algae. J. Dairy Sci. 2010, 93, 1655–1667. [Google Scholar] [CrossRef] [PubMed]
- Bichi, E.; Hervas, G.; Toral, P.G.; Loor, J.J.; Frutos, P. Milk fat depression induced by dietary marine algae in dairy ewes: Persistency of milk fatty acid composition and animal performance responses. J. Dairy Sci. 2013, 96, 524–532. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Unit | ALA |
|---|---|---|
| Flaxseed oil | g/tbsp | 7.26 |
| Chia seed | g/ounce | 5.06 |
| English walnuts | g/ounce | 2.57 |
| Whole flaxseed | g/tbsp | 2.35 |
| Canola oil | g/tbsp | 1.28 |
| Soybean oil | g/tbsp | 0.92 |
| Black walnut | g/ounce | 0.76 |
| Item | Unit | EPA | DHA | DPA | Total n-3 LC-PUFA | Reference |
|---|---|---|---|---|---|---|
| Wild seafood | Nichols et al. [1] | |||||
| Fish | mg/150 g | - | - | - | 350 | |
| Shellfish | mg/150 g | - | - | - | 225 | |
| Prawns | mg/150 g | - | - | - | 180 | |
| Lobster | mg/150 g | - | - | - | 160 | |
| Farmed fish | ||||||
| Atlantic salmon | mg/100 g | - | - | - | 980 | Nichols et al. [38] |
| Barramundi | mg/100 g | - | - | - | 790 | |
| Other animal sources | ||||||
| Beef | mg/100 g | 15 | 12 | 20 | 47 | Garcia et al. [39] |
| Chicken breast | mg/100 g | - | - | - | 62.04 | Konieczka et al. [40] |
| Pork | mg/100 g | 23.3 | 3.9 | 21.1 | 48.3 | Dugan et al. [41] |
| Feedlot lamb meat | mg/100 g | 17.9 | 4.9 | 15.6 | 38.4 | Nguyen et al. [42] |
| 28.9 | 13.3 | 19.6 | 61.8 | Le et al. [43] | ||
| Grazing lamb meat | mg/100 g | 25 | 7.1 | 23.7 | 55.8 | Le et al. [44] |
| Sheep milk | mg/250 mL | 17.8 | 19.8 | 24.1 | 61.7 | Nguyen et al. [45] |
| Sheep cheese | mg/40 g | 14.3 | 12.8 | 17.1 | 44.2 | Nguyen et al. [46] |
| Cow milk | mg/100 g | 3.3 | - | 4.4 | - | Benbrook et al. [47] |
| Forage Source/Feeding System | Species | ALA | EPA | DHA | DPA | References |
|---|---|---|---|---|---|---|
| Ryegrass-dominated pastures | Bovine | 0.703 | 0.083 | 0.009 | 0.109 | Leiber et al. [105] |
| Freshly harvested ryegrass | 0.619 | 0.073 | 0.009 | 0.113 | ||
| Alpine pastures | 1.146 | 0.083 | 0.009 | 0.120 | ||
| Freshly harvested Alpine | 0.950 | 0.083 | 0.010 | 0.118 | ||
| Silage-concentrate diet (control) 1 | 0.516 | 0.063 | ND | 0.082 | ||
| Ryegrass pasture | Bovine | 0.68 | 0.05 | 0.02 | 0.07 | Mohammed et al. [107] |
| Freshly harvested ryegrass | 0.82 | 0.07 | 0.02 | 0.08 | ||
| Ryegrass silage | 0.34 | 0.05 | 0.02 | 0.09 | ||
| Indoor hay based diet | Bovine | 0.72 | 0.08 | - | 0.147 | Coppa et al. [119] |
| Rotational grazing system | 0.727 | 0.070 | - | 0.137 | ||
| Continuous grazing system | 0.940 | 0.087 | - | 0.150 | ||
| Indoor conventional system | Bovine | 0.579 | 0.072 | - | 0.118 | Stergiadis et al. [120] |
| Indoor organic system | 1.199 | 0.098 | - | 0.098 | ||
| Mixed forage 2 | Bovine | 0.47 | - | - | - | Liu et al. [121] |
| Corn stalk1 diet (35%) | 0.58 | - | - | - | ||
| Corn stalk2 diet (53.8%) | 0.63 | - | - | - | ||
| Daisy forb − winter | Ovine | 1.62 | - | - | - | Addis et al. [109] |
| Ryegrass − winter | 1.47 | - | - | - | ||
| Burr medic − winter | 2.19 | - | - | - | ||
| Sulla − winter | 2.98 | - | - | - | ||
| Daisy forb − spring | 1.26 | - | - | - | ||
| Ryegrass − spring | 1.44 | - | - | - | ||
| Burr medic − spring | 1.84 | - | - | - | ||
| Sulla − spring | 3.15 | - | - | - | ||
| Pasture | Ovine | 1.07 | 0.06 | - | 0.13 | Gomez-Cortes et al. [104] |
| Pasture + oat grain | 0.59 | 0.05 | - | 0.12 | ||
| Total mixed ration 3 | 0.33 | 0.03 | - | 0.06 | ||
| Grass hay (in door) | Ovine | 1.31 | 0.19 | 0.30 | - | Mierlita [106] |
| Part-time grazing | 2.06 | 0.28 | 0.39 | - | ||
| Pasture | Ovine | 2.09 | 0.30 | 0.37 | - | Mierlita et al. [122] |
| Pasture + standard concentrate 3 | 1.04 | 0.11 | 0.18 | - | ||
| Pasture | Ovine | 0.44 | 0.01 | 0.07 | 0.13 | Mohamed et al. [123] |
| Pasture + concentrate | 0.24 | 0.00 | 0.12 | 0.07 | ||
| Concentrate | 0.21 | 0.00 | 0.00 | 0.08 | ||
| Red clover silage | Ovine | 0.92 | 0.05 | - | 0.09 | Guzatti et al. [111] |
| Lucerne silage | 0.70 | 0.05 | - | 0.09 |
| Diet | Species | Product | ALA | EPA | DHA | DPA | References |
|---|---|---|---|---|---|---|---|
| Control | Bovine | Milk | 0.61 | 0.09 | - | 0.07 | Khas et al. [124] |
| 40 g/day infused LNA-rich fatty acid | 6.49 | 0.18 | - | 0.12 | |||
| 80 g/day infused LNA-rich fatty acid | 12.42 | 0.22 | - | 0.16 | |||
| 120 g/day infused LNA-rich fatty acid | 18.75 | 0.21 | - | 0.29 | |||
| 160 g/day infused LNA-rich fatty acid | 25.38 | 0.22 | - | 0.23 | |||
| Control | Bovine | Milk | 0.75 | 0.003 | 0.001 | - | Caroprese et al. [134] |
| Whole flaxseed | 0.81 | 0.022 | 0.001 | - | |||
| Control | Bovine | Milk | 0.41 | 0.05 | - | 0.05 | Dai et al. [135] |
| Rapeseed oil | 0.38 | 0.06 | - | 0.04 | |||
| Peanut oil | 0.33 | 0.06 | - | 0.06 | |||
| Sunflower seed oil | 0.32 | 0.06 | - | 0.05 | |||
| Control | Bovine | Milk | 0.83 | 0.09 | 0.01 | 0.13 | Otto et al. [136] |
| 25mL/kg DM 1 Canola oil | 0.85 | 0.09 | 0.01 | 0.14 | |||
| 35 mL/kg DM canola oil | 0.95 | 0.08 | 0.01 | 0.12 | |||
| 50 mL/kg DM canola oil | 0.97 | 0.08 | 0.00 | 0.11 | |||
| Control | Bovine | Milk | 0.28 | 0.02 | - | - | Cattani et al. [137] |
| 500 g/day extruded flaxseed | 0.50 | 0.02 | - | - | |||
| 1000 g/day extruded flaxseed | 0.59 | 0.02 | - | - | |||
| Linseed oil | Bovine | Milk | 0.249 | 0.019 | - | 0.014 | Li et al. [138] |
| Safflower oil | 0.180 | 0.013 | - | 0.007 | |||
| Control | Bovine | Milk | 0.19 | 0.012 | 0.004 | 0.037 | Welter et al. [139] |
| 3% Canola oil | 0.36 | 0.011 | 0.003 | 0.034 | |||
| 6% Canola oil | 0.35 | 0.011 | 0.003 | 0.033 | |||
| Control | Bovine | Milk | 0.19 | - | 0.019 | - | Vanbergue et al. [140] |
| Extruded linseed | 0.51 | - | 0.008 | - | |||
| Palm oil | Ovine | Milk | 0.52 | 0.04 | 0.02 | 0.08 | Bodas et al. [141] |
| Olive oil | 0.36 | 0.03 | 0.02 | 0.06 | |||
| Soybean oil | 0.53 | 0.03 | 0.02 | 0.07 | |||
| Linseed oil | 1.07 | 0.05 | 0.04 | 0.11 | |||
| Control | Ovine | Milk | 1.21 | 0.05 | 0.05 | - | Mughetti et al. [142] |
| 100 g extruded linseed | 1.65 | 0.06 | 0.09 | - | |||
| 200 g extruded linseed | 2.26 | 0.06 | 0.10 | - | |||
| Control | Ovine | Milk | 0.57 | 0.07 | 0.05 | 0.08 | Caroprese et al. [143] |
| Seaweed | 0.59 | 0.06 | 0.04 | 0.08 | |||
| Whole flaxseed | 1.53 | 0.08 | 0.05 | 0.09 | |||
| Seaweed + Whole flaxseed | 1.32 | 0.08 | 0.06 | 0.10 | |||
| Control | Ovine | Milk | 0.62 | 0.08 | 0.04 | 0.08 | Nguyen et al. [45] |
| Canola oil | 0.73 | 0.09 | 0.06 | 0.13 | |||
| Rice bran oil | 0.51 | 0.07 | 0.04 | 0.10 | |||
| Flaxseed oil | 1.74 | 0.11 | 0.06 | 0.15 | |||
| Safflower oil | 0.67 | 0.07 | 0.06 | 0.10 | |||
| Control | Ovine | Milk | 0.31 | 0.04 | 0.02 | 0.08 | Parentet et al. [144] |
| Canola oil | 0.26 | 0.03 | 0.02 | 0.07 | |||
| Sunflower oil | 0.24 | 0.03 | 0.02 | 0.07 | |||
| Castor oil | 0.28 | 0.05 | 0.01 | 0.08 | |||
| Control | Bovine | Cheese | 0.29 | 0.02 | - | - | Cattani et al. [137] |
| 500 g/day extruded Flaxseed at | 0.50 | 0.02 | - | - | |||
| 1000 g/day extruded Flaxseed at | 0.61 | 0.02 | - | - | |||
| Palm oil | Ovine | Cheese | 0.54 | 0.04 | 0.02 | 0.07 | Bodas et al. [141] |
| Olive oil | 0.36 | 0.03 | 0.03 | 0.06 | |||
| Soybean oil | 0.51 | 0.03 | 0.02 | 0.06 | |||
| Linseed oil | 1.04 | 0.03 | 0.03 | 0.09 | |||
| Control | Ovine | Cheese | 1.18 | 0.02 | 0.03 | - | Mughettiet al. [142] |
| 100 g extruded linseed | 1.84 | 0.04 | 0.05 | - | |||
| 200 g extruded linseed | 2.02 | 0.04 | 0.06 | - | |||
| Control | Ovine | Cheese | 0.71 | 0.11 | 0.06 | 0.12 | Nguyen et al. [46] |
| Canola oil | 0.79 | 0.11 | 0.06 | 0.13 | |||
| Rice bran oil | 0.63 | 0.10 | 0.06 | 0.12 | |||
| Flaxseed oil | 1.30 | 0.11 | 0.06 | 0.13 | |||
| Safflower oil | 0.71 | 0.11 | 0.08 | 0.13 | |||
| Control | Ovine | Yogurt | 0.0 | - | - | - | Bianchi et al. [145] |
| 2% Palm oil | 0.0 | - | - | - | |||
| 4% Palm oil | 0.28 | - | - | - | |||
| 6% Palm oil | 0.31 | - | - | - |
| Diet | Species | Product | ALA | EPA | DHA | DPA | References |
|---|---|---|---|---|---|---|---|
| Control | Bovine | Milk | 0.54 | - | 0.00 | - | Franklin et al. [161] |
| Protected algae | 0.49 | - | 0.76 | - | |||
| Unprotected algae | 0.47 | - | 0.46 | - | |||
| Control | Bovine | Milk | 0.86 | 0.0 | 0.0 | - | Kitessa et al. [126] |
| Rumen-protected tuna oil | 1.28 | 0.61 | 1.09 | - | |||
| Control | Bovine | Milk | 0.21 | 0.03 | 0.00 | 0.07 | Shingfield et al. [162] |
| Fish oil and sunflower oil | 0.23 | 0.11 | 0.07 | 0.16 | |||
| Control | Bovine | Milk | 0.50 | - | 0.09 | - | Boeckaert et al. [129] |
| Marine algae | 0.42 | - | 1.01 | - | |||
| ABO/ABO 1 | Bovine | Milk | 14.4 | 0.22 | - | 0.22 | Kazama et al. [163] |
| RUM/ABO 2 | 4.78 | 0.14 | - | 0.22 | |||
| RUM/RUM 3 | 2.33 | 0.09 | - | 0.12 | |||
| ABO/RUM 4 | 11.6 | 0.16 | - | 0.18 | |||
| Control | Bovine | Milk | 0.75 | 0.003 | 0.001 | - | Caroprese et al. [134] |
| Fish oil | 0.84 | 0.060 | 0.117 | - | |||
| Control | Bovine | Milk | 0.45 | 0.06 | 0.10 | - | Vargas-Bello-Pérez et al. [164] |
| Fish oil | 0.62 | 0.10 | 0.21 | - | |||
| Fish oil + palm oil | 0.69 | 0.09 | 0.14 | - | |||
| Control | Bovine | Milk | 0.41 | 0.06 | 0.03 | 0.09 | Kairenius et al. [165] |
| Ultrarefined fish oil at 75 g/day | 0.38 | 0.06 | 0.03 | 0.08 | |||
| Ultrarefined fish oil at 150 g/day | 0.39 | 0.07 | 0.05 | 0.10 | |||
| Ultrarefined fish oil at 300 g/day | 0.48 | 0.17 | 0.10 | 0.18 | |||
| Control | Bovine | Milk | 0.19 | - | 0.019 | - | Vanbergue et al. [140] |
| Microalgae DHA Gold® | 0.25 | - | 0.444 | - | |||
| Extruded linseed + DHA Gold® | 0.46 | - | 0.170 | - | |||
| Control | Ovine | Milk | 0.33 | ND | ND | ND | Papadopoulos et al. [128] |
| Low algae (23.5 g) | 0.31 | 0.04 | 0.43 | 0.21 | |||
| Medium algae (47 g) | 0.33 | 0.12 | 0.69 | 0.28 | |||
| High algae (94 g) | 0.25 | 0.21 | 1.24 | 0.31 | |||
| Control | Ovine | Milk | 0.53 | 0.05 | 0.03 | 0.10 | Toral et al. [166] |
| Sunflower oil (SO) | 0.41 | 0.04 | 0.02 | 0.07 | |||
| SO + 8 g/kg DM of Marine Algae | 0.37 | 0.05 | 0.17 | 0.10 | |||
| SO + 16 g/kg DM of Marine Algae | 0.36 | 0.09 | 0.46 | 0.13 | |||
| SO + 24 g/kg DM of Marine Algae | 0.34 | 0.10 | 0.57 | 0.15 | |||
| Sunflower oil | Ovine | Milk | 0.49 | 0.04 | 0.05 | 0.10 | Bichi et al. [167] |
| Sunflower oil + Marine algae | 0.48 | 0.06 | 0.38 | 0.12 | |||
| Control | Ovine | Milk | 0.62 | 0.08 | 0.04 | 0.08 | Nguyen et al. [45] |
| Rumen-protected EPA + DHA oil | 0.74 | 0.17 | 0.19 | 0.23 | |||
| Control | Bovine | Cheese | 0.01 | 0.05 | 0.09 | - | Vargas-Bello-Pérez et al. [164] |
| Fish oil | 0.02 | 0.12 | 0.34 | - | |||
| Fish oil + palm oil | 0.01 | 0.09 | 0.18 | - | |||
| Control | Ovine | Cheese | 0.71 | 0.11 | 0.06 | 0.12 | Nguyen et al. [46] |
| Rumen-protected EPA + DHA | 1.02 | 0.16 | 0.15 | 0.19 |
| Breed | Species | Unit | ALA | EPA | DPA | n-3 | Reference |
|---|---|---|---|---|---|---|---|
| Holstein-Friesians | Bovine | %FA | 0.09 | - | - | - | Stoop et al. [147] |
| Holsteins | Bovine | % FA | 0.06 | 0.04 | 0.01 | - | Bilal et al. [148] |
| Brown Swiss | Bovine | % FA | 0.093 | 0.045 | 0.039 | 0.085 | Pegolo et al. [149] |
| Sarda | Ovine | % FA | 0.02 | - | - | - | Correddu et al. [150] |
| Holstein | Bovine | % fat | - | - | - | 0.26 | Boichard et al. [151] |
| Saanen | Caprine | - | - | - | 0.23 | ||
| Lacaune | Ovine | - | - | - | 0.18 | ||
| Alpine | Caprine | % fat | - | - | - | 0.28 | Maroteau et al. [152] |
| Saanen | Caprine | - | - | - | 0.25 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, Q.V.; Malau-Aduli, B.S.; Cavalieri, J.; Nichols, P.D.; Malau-Aduli, A.E.O. Enhancing Omega-3 Long-Chain Polyunsaturated Fatty Acid Content of Dairy-Derived Foods for Human Consumption. Nutrients 2019, 11, 743. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11040743
Nguyen QV, Malau-Aduli BS, Cavalieri J, Nichols PD, Malau-Aduli AEO. Enhancing Omega-3 Long-Chain Polyunsaturated Fatty Acid Content of Dairy-Derived Foods for Human Consumption. Nutrients. 2019; 11(4):743. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11040743
Chicago/Turabian StyleNguyen, Quang V., Bunmi S. Malau-Aduli, John Cavalieri, Peter D. Nichols, and Aduli E. O. Malau-Aduli. 2019. "Enhancing Omega-3 Long-Chain Polyunsaturated Fatty Acid Content of Dairy-Derived Foods for Human Consumption" Nutrients 11, no. 4: 743. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11040743