Short-Term High-Starch, Low-Protein Diet Induces Reversible Increase in β-cell Mass Independent of Body Weight Gain in Mice

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Diets
2.2. Intraperitoneal Glucose Tolerance Test (IPGTT) and Insulin Tolerance Test (ITT)
2.3. Plasma Biochemical Analyses
2.4. Isolation of Ribonucleic Acid (RNA) and Quantitative Real-Time Reverse Transcription Polymerase Chain Reaction (RT-PCR)
2.5. Immunohistochemistry and Morphological Analyses
2.6. Statistical Analysis
3. Results
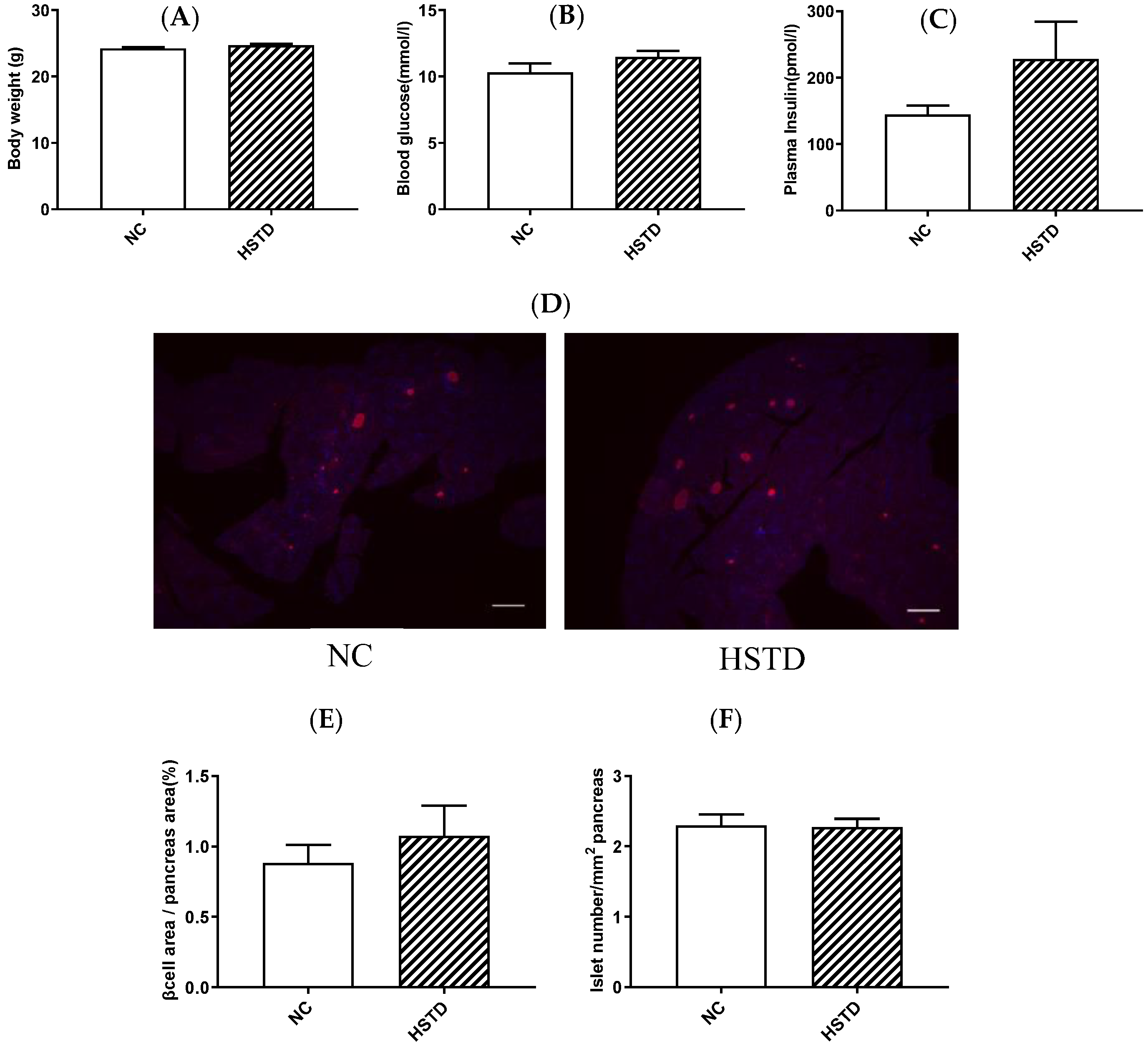
3.1. Pancreatic Islet Number and BCM Were not Increased in HSTD-Fed Mice 3 Weeks after Intervention
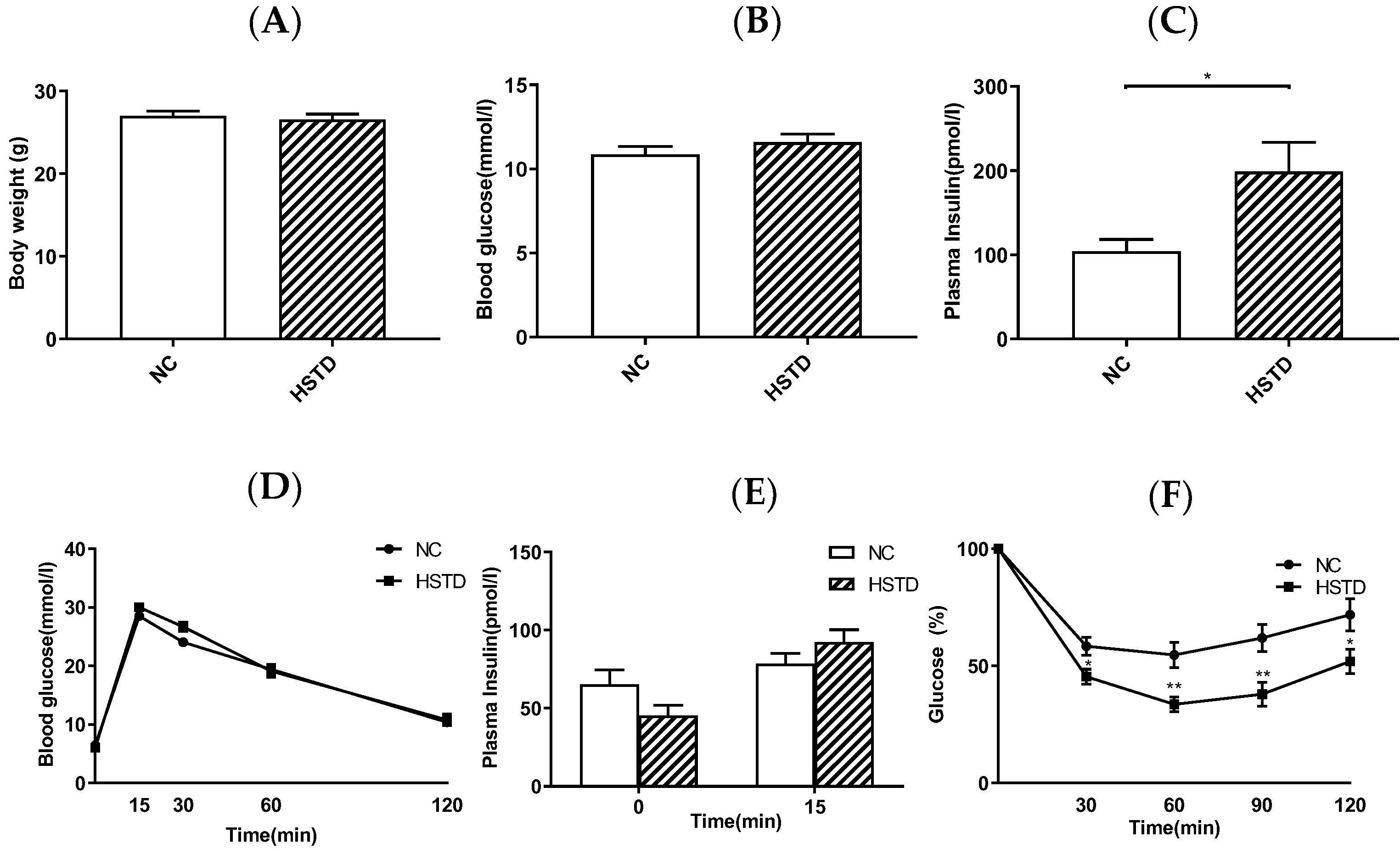
3.2. Plasma Insulin Levels Were Elevated in HSTD-Fed Mice 5 Weeks after Intervention
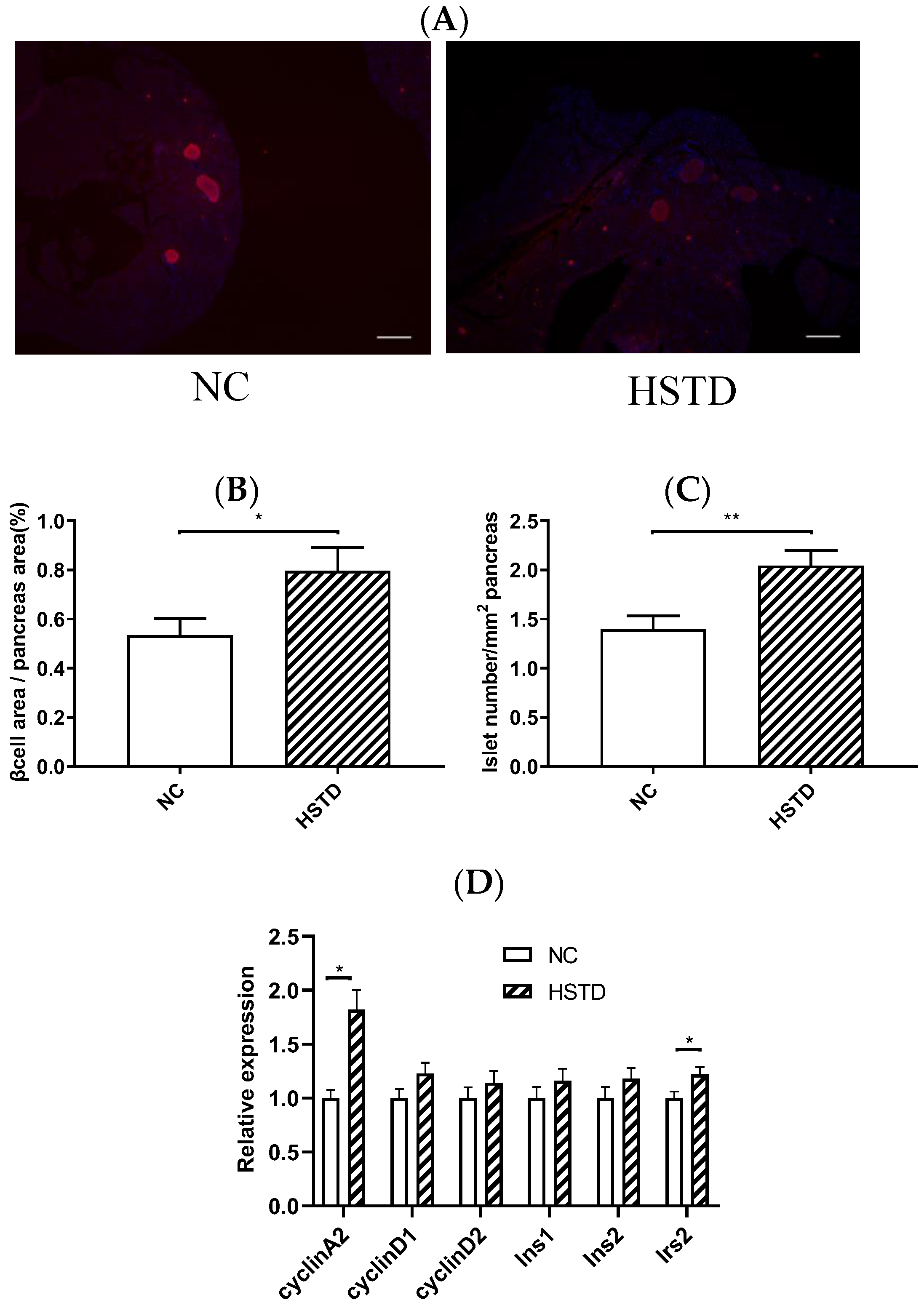
3.3. Pancreatic Islet Number and BCM Were Increased in HSTD-Fed Mice 5 Weeks after Intervention
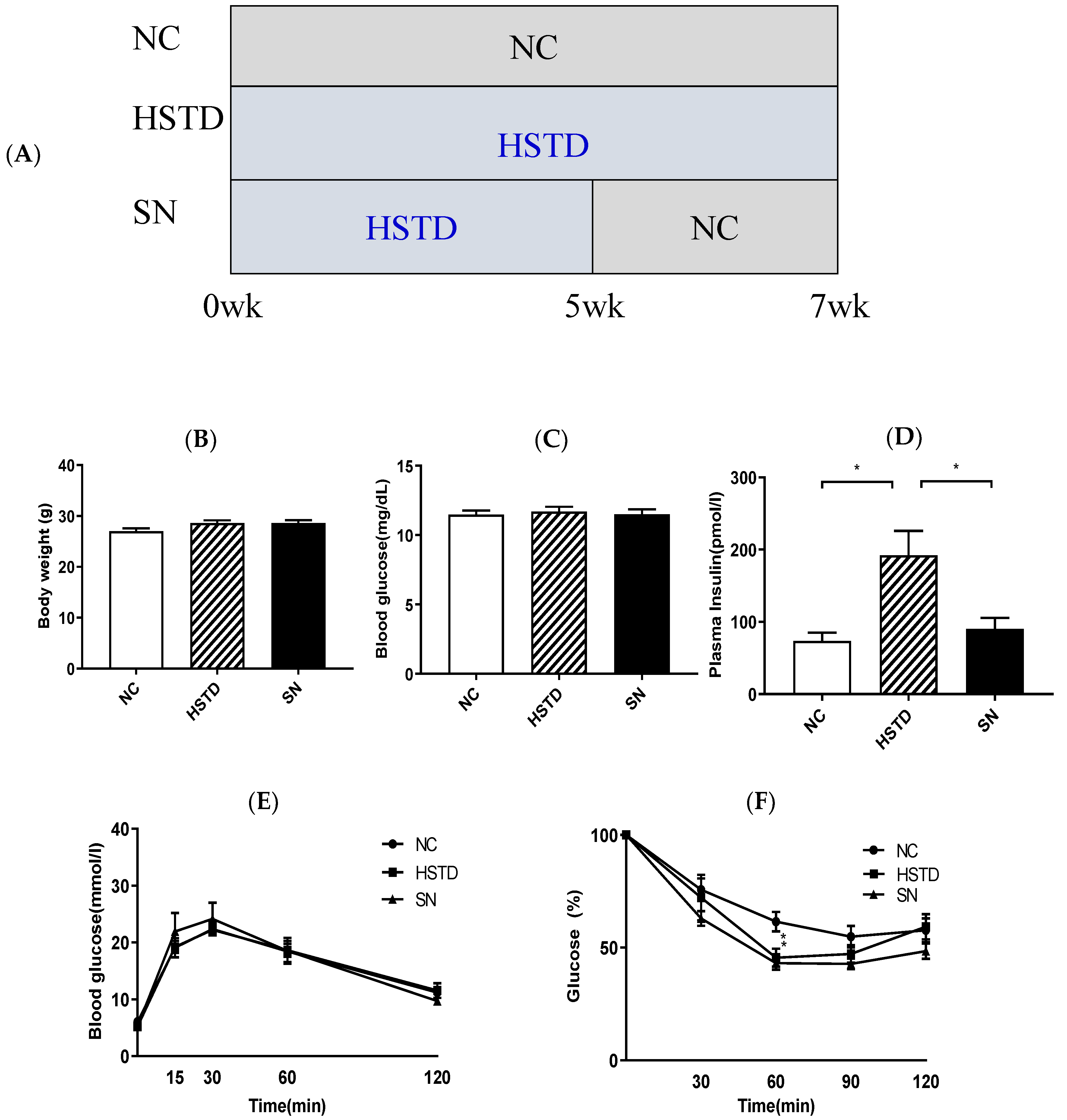
3.4. Switching from HSTD to NC Decreased Pancreatic Islet Number and BCM
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Seino, S.; Sugawara, K.; Yokoi, N.; Takahashi, H. β-Cell signalling and insulin secretagogues: A path for improved diabetes therapy. Diabetes Obes. Metab. 2017, 19, 22–29. [Google Scholar] [CrossRef] [Green Version]
- Mehran, A.E.; Templeman, N.M.; Brigidi, G.S.; Lim, G.E.; Chu, K.Y.; Hu, X.; Botezelli, J.D.; Asadi, A.; Hoffman, B.G.; Kieffer, T.J.; et al. Hyperinsulinemia Drives Diet-Induced Obesity Independently of Brain Insulin Production. Cell Metab. 2012, 16, 723–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blüher, M.; Dodson Michael, M.; Peroni, O.D.; Ueki, K.; Carter, N.; Kahn, B.B.; Ronald Kahn, C. Adipose Tissue Selective Insulin Receptor Knockout Protects against Obesity and Obesity-Related Glucose Intolerance insulin resistance, moderate glucose intolerance, and a failure of insulin to suppress hepatic glucose production and regulate hepatic gene. Dev. Cell 2002, 3, 25–38. [Google Scholar] [CrossRef]
- Miyawaki, K.; Yamada, Y.; Ban, N.; Ihara, Y.; Tsukiyama, K.; Zhou, H.; Fujimoto, S.; Oku, A.; Tsuda, K.; Toyokuni, S.; et al. Inhibition of gastric inhibitory polypeptide signaling prevents obesity. Nat. Med. 2002, 8, 738–742. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, R.; Ogata, H.; Murase, M.; Harada, N.; Suzuki, K.; Joo, E.; Sankoda, A.; Iida, A.; Izumoto, T.; Tsunekawa, S.; et al. Glucose-dependent insulinotropic polypeptide is required for moderate high-fat diet- but not high-carbohydrate diet-induced weight gain. Am. J. Physiol. Metab. 2018, 314, E572–E583. [Google Scholar] [CrossRef] [PubMed]
- Nasteska, D.; Harada, N.; Suzuki, K.; Yamane, S.; Hamasaki, A.; Joo, E.; Iwasaki, K.; Shibue, K.; Harada, T.; Inagaki, N. Chronic Reduction of GIP Secretion Alleviates Obesity and Insulin Resistance Under High-Fat Diet Conditions. Diabetes 2014, 63, 2332–2343. [Google Scholar] [CrossRef] [Green Version]
- Murase, M.; Seino, Y.; Maekawa, R.; Iida, A.; Hosokawa, K.; Hayami, T.; Tsunekawa, S.; Hamada, Y.; Yokoi, N.; Seino, S.; et al. Functional adenosine triphosphate-sensitive potassium channel is required in high-carbohydrate diet-induced increase in β-cell mass. J. Diabetes Investig. 2019, 10, 238–250. [Google Scholar] [CrossRef]
- Butler, A.E.; Janson, J.; Soeller, W.C.; Butler, P.C. Increased beta-cell apoptosis prevents adaptive increase in beta-cell mass in mouse model of type 2 diabetes: evidence for role of islet amyloid formation rather than direct action of amyloid. Diabetes 2003, 52, 2304–2314. [Google Scholar] [CrossRef]
- Saisho, Y.; Butler, A.E.; Manesso, E.; Elashoff, D.; Rizza, R.A.; Butler, P.C. β-Cell Mass and Turnover in Humans: Effects of obesity and aging. Diabetes Care 2013, 36, 111–117. [Google Scholar] [CrossRef]
- Maekawa, R.; Seino, Y.; Ogata, H.; Murase, M.; Iida, A.; Hosokawa, K.; Joo, E.; Harada, N.; Tsunekawa, S.; Hamada, Y.; et al. Chronic high-sucrose diet increases fibroblast growth factor 21 production and energy expenditure in mice. J. Nutr. Biochem. 2017, 49, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Niwa, Y.; Ishikawa, K.; Ishigami, M.; Honda, T.; Achiwa, K.; Izumoto, T.; Maekawa, R.; Hosokawa, K.; Iida, A.; Seino, Y.; et al. Effect of hyperglycemia on hepatocellular carcinoma development in diabetes. Biochem. Biophys. Res. Commun. 2015, 463, 344–350. [Google Scholar] [CrossRef] [Green Version]
- Fukami, A.; Seino, Y.; Ozaki, N.; Yamamoto, M.; Sugiyama, C.; Sakamoto-Miura, E.; Himeno, T.; Takagishi, Y.; Tsunekawa, S.; Ali, S.; et al. Ectopic Expression of GIP in Pancreatic β-Cells Maintains Enhanced Insulin Secretion in Mice With Complete Absence of Proglucagon-Derived Peptides. Diabetes 2013, 62, 510–518. [Google Scholar] [CrossRef] [Green Version]
- Bonner-Weir, S.; Deery, D.; Leahy, J.L.; Weir, G.C. Compensatory growth of pancreatic beta-cells in adult rats after short-term glucose infusion. Diabetes 1989, 38, 49–53. [Google Scholar] [CrossRef]
- Lipsett, M.; Finegood, D.T. B-Cell Neogenesis during Prolonged Hyperglycemia in Rats. Diabetes 2002, 51, 1834–1841. [Google Scholar] [CrossRef]
- Jetton, T.L.; Everill, B.; Lausier, J.; Roskens, V.; Habibovic, A.; LaRock, K.; Gokin, A.; Peshavaria, M.; Leahy, J.L. Enhanced beta-cell mass without increased proliferation following chronic mild glucose infusion. Am. J. Physiol. Endocrinol. Metab. 2008, 294, E679–E687. [Google Scholar] [CrossRef]
- Moullé, V.S.; Vivot, K.; Tremblay, C.; Zarrouki, B.; Ghislain, J.; Poitout, V. Glucose and fatty acids synergistically and reversibly promote beta cell proliferation in rats. Diabetologia 2017, 60, 879–888. [Google Scholar] [CrossRef] [Green Version]
- Hull, R.L.; Kodama, K.; Utzschneider, K.M.; Carr, D.B.; Prigeon, R.L.; Kahn, S.E. Dietary-fat-induced obesity in mice results in beta cell hyperplasia but not increased insulin release: Evidence for specificity of impaired beta cell adaptation. Diabetologia 2005, 48, 1350–1358. [Google Scholar] [CrossRef]
- Terauchi, Y.; Takamoto, I.; Kubota, N.; Matsui, J.; Suzuki, R.; Komeda, K.; Hara, A.; Toyoda, Y.; Miwa, I.; Aizawa, S.; et al. Glucokinase and IRS-2 are required for compensatory β cell hyperplasia in response to high-fat diet-induced insulin resistance. J. Clin. Investig. 2007, 117, 246–257. [Google Scholar] [CrossRef]
- Tajima, K.; Shirakawa, J.; Okuyama, T.; Kyohara, M.; Yamazaki, S.; Togashi, Y.; Terauchi, Y. Effects of metformin on compensatory pancreatic β-cell hyperplasia in mice fed a high-fat diet. Am. J. Physiol. Metab. 2017, 313, E149–E159. [Google Scholar] [CrossRef]
- Mosser, R.E.; Maulis, M.F.; Moullé, V.S.; Dunn, J.C.; Carboneau, B.A.; Arasi, K.; Pappan, K.; Poitout, V.; Gannon, M. High-fat diet-induced β-cell proliferation occurs prior to insulin resistance in C57Bl/6J male mice. Am. J. Physiol. Metab. 2015, 308, E573–E582. [Google Scholar] [CrossRef] [Green Version]
- Stamateris, R.E.; Sharma, R.B.; Hollern, D.A.; Alonso, L.C. Adaptive β-cell proliferation increases early in high-fat feeding in mice, concurrent with metabolic changes, with induction of islet cyclin D2 expression. Am. J. Physiol. Metab. 2013, 305, E149–E159. [Google Scholar] [CrossRef] [Green Version]
- Kitao, N.; Nakamura, A.; Miyoshi, H.; Nomoto, H.; Takahashi, K.; Omori, K.; Yamamoto, K.; Cho, K.Y.; Terauchi, Y.; Atsumi, T. The role of glucokinase and insulin receptor substrate-2 in the proliferation of pancreatic beta cells induced by short-term high-fat diet feeding in mice. Metabolism 2018, 85, 48–58. [Google Scholar] [CrossRef]
- Georgia, S.; Hinault, C.; Kawamori, D.; Hu, J.; Meyer, J.; Kanji, M.; Bhushan, A.; Kulkarni, R.N. Cyclin D2 is essential for the compensatory β-cell hyperplastic response to insulin resistance in Rodents. Diabetes 2010, 59, 987–996. [Google Scholar] [CrossRef]
- Imai, J.; Katagiri, H.; Yamada, T.; Ishigaki, Y.; Suzuki, T.; Kudo, H.; Uno, K.; Hasegawa, Y.; Gao, J.; Kaneko, K.; et al. Regulation of Pancreatic b Cell Mass by Neuronal Signals from the Liver. Science 2008, 322, 1250–1254. [Google Scholar] [CrossRef]
- Agardh, C.-D.; Ahrén, B. Switching from high-fat to low-fat diet normalizes glucose metabolism and improves glucose-stimulated insulin secretion and insulin sensitivity but not body weight in C57BL/6J mice. Pancreas 2012, 41, 253–257. [Google Scholar] [CrossRef]
- Parekh, P.I.; Tiller, J.M.; Feinglos, M.N.; Design, E.; Sciences, B.; Sciences, H. Reversal of Diet-Induced Obesity and Diabetes in C57BL/6J Mice. Metabolism 1998, 9, 1089–1096. [Google Scholar] [CrossRef]
- Kowalski, G.M.; Hamley, S.; Selathurai, A.; Kloehn, J.; De Souza, D.P.; O’Callaghan, S.; Nijagal, B.; Tull, D.L.; McConville, M.J.; Bruce, C.R. Reversing diet-induced metabolic dysregulation by diet switching leads to altered hepatic de novo lipogenesis and glycerolipid synthesis. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef]
- Kim, J.; Bae, K.; Choi, Y.; Go, Y.; Choe, M.; Jeon, Y.; Lee, H.; Koo, S.; Perfield, J.W.; Harris, R.A.; et al. Fibroblast growth factor 21 analogue LY2405319 lowers blood glucose in streptozotocin-induced insulin-deficient diabetic mice by restoring brown adipose tissue function. Diabetes Obes. Metab. 2015, 161–169. [Google Scholar] [CrossRef]
- Laeger, T.; Henagan, T.M.; Albarado, D.C.; Redman, L.M.; Bray, G.A.; Noland, R.C.; Münzberg, H.; Hutson, S.M.; Gettys, T.W.; Morrison, C.D.; et al. FGF21 is an endocrine signal of protein restriction. J. Clin. Investig. 2014, 124, 3913–3922. [Google Scholar] [CrossRef] [Green Version]
- Pellizzon, M.A.; Ricci, M.R. The common use of improper control diets in diet-induced metabolic disease research confounds data interpretation: The fiber factor. Nutr. Metab. 2018, 15, 1–6. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′ to 3′) | Reverse Primer (5′ to 3′) |
|---|---|---|
| βactin | CATCCGTAAAGACCTCTATGCCAAC | ATGGAGCCACCGATCCACA |
| cyclinA2 | TCCTTGCTTTTGACTTGGCT | ATGACTCAGGCCAGCTCTGT |
| cyclinD1 | GCCGAGAAGTTGTGCATCTA | TCACCAGAAGCAGTTCCATTT |
| cyclinD2 | CACCGACAACTCTGTGAAGC | TCCACTTCAGCTTACCCAACA |
| Ins1 | CCTTAGTGACCAGCTATAATCAGAG | AGATGCTGTTTGACAAAAGCC |
| Ins2 | AGCCCTAAGTGATCCGCTACA | AAGGTGCTGCTTGACAAAAGCC |
| IRS2 | TCCAGGCACTGGAGCTTT | GGCTGGTAGCGCTTCACT |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masuda, A.; Seino, Y.; Murase, M.; Hidaka, S.; Shibata, M.; Takayanagi, T.; Sugimura, Y.; Hayashi, Y.; Suzuki, A. Short-Term High-Starch, Low-Protein Diet Induces Reversible Increase in β-cell Mass Independent of Body Weight Gain in Mice. Nutrients 2019, 11, 1045. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11051045
Masuda A, Seino Y, Murase M, Hidaka S, Shibata M, Takayanagi T, Sugimura Y, Hayashi Y, Suzuki A. Short-Term High-Starch, Low-Protein Diet Induces Reversible Increase in β-cell Mass Independent of Body Weight Gain in Mice. Nutrients. 2019; 11(5):1045. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11051045
Chicago/Turabian StyleMasuda, Atsushi, Yusuke Seino, Masatoshi Murase, Shihomi Hidaka, Megumi Shibata, Takeshi Takayanagi, Yoshihisa Sugimura, Yoshitaka Hayashi, and Atsushi Suzuki. 2019. "Short-Term High-Starch, Low-Protein Diet Induces Reversible Increase in β-cell Mass Independent of Body Weight Gain in Mice" Nutrients 11, no. 5: 1045. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11051045