Exploring Associations between Interindividual Differences in Taste Perception, Oral Microbiota Composition, and Reported Food Intake



Abstract
:1. Introduction
2. Material and Methods
2.1. Participants
2.2. Gustatory Function Assessments
2.3. Food Intake Evaluation
2.4. Oral Sample Collection, DNA Extraction, and Microbiota Composition Evaluation
2.5. Data Analysis
3. Results
3.1. Participant Characteristics
3.2. Association among Gustatory Functions and Their Relationship with Food Intake
3.2.1. Salty Sensitivity
3.2.2. Sweet Sensitivity
3.2.3. Bitter Sensitivity
3.2.4. Sour Sensitivity
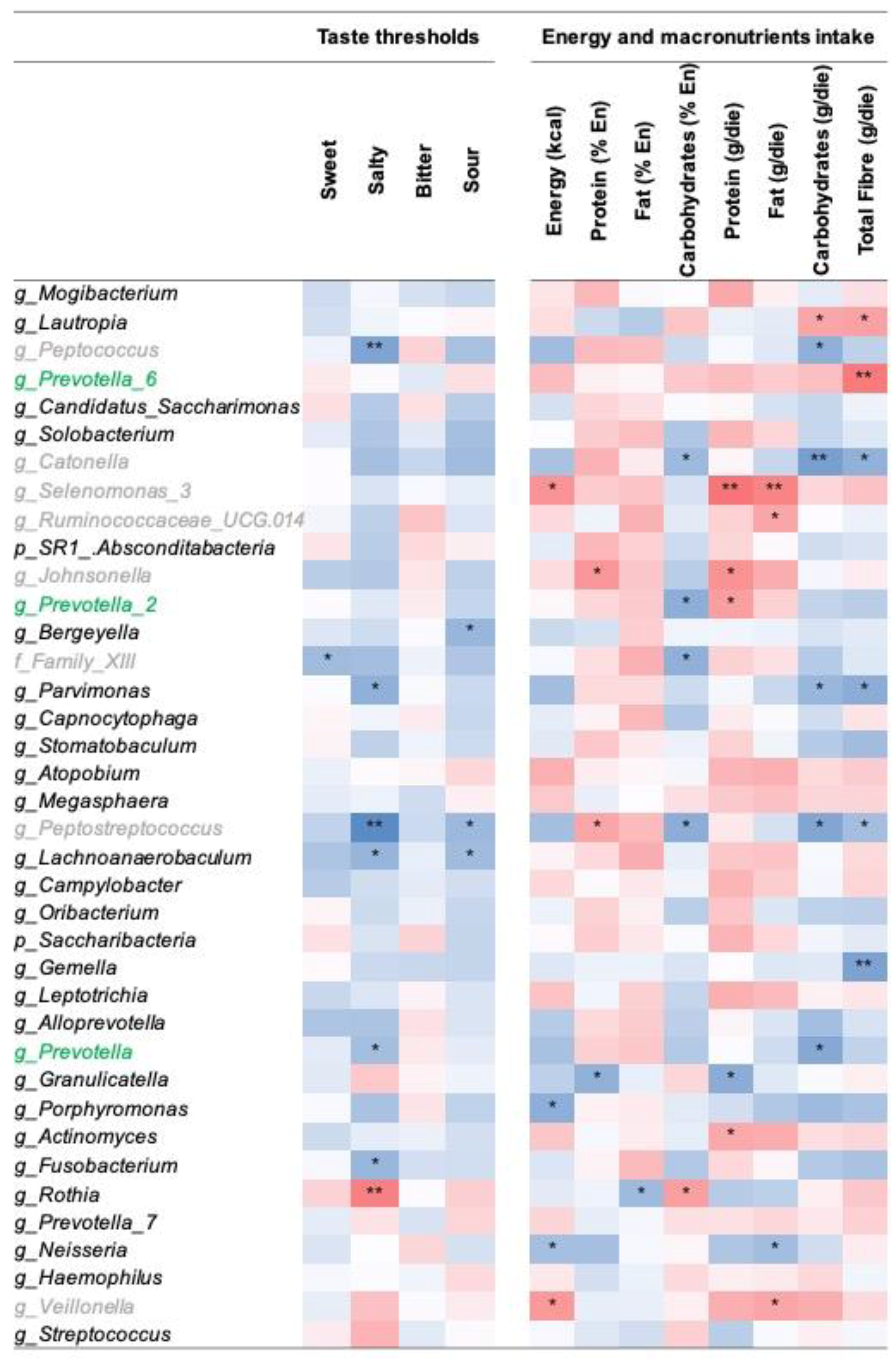
3.3. Correlation between Tongue Dorsum Microbiota, Gustatory Functions, and Dietary Intake
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mennella, J.A.; Pepino, M.Y.; Reed, D.R. Genetic and environmental determinants of bitter perception and sweet preferences. Pediatrics 2005, 115, e216–e222. [Google Scholar] [CrossRef]
- Chaudhari, N.; Roper, S.D. The cell biology of taste. J. Cell Biol. 2010, 190, 285–296. [Google Scholar] [CrossRef] [Green Version]
- Reed, D.R.; Tanaka, T.; McDaniel, A.H. Diverse tastes: Genetics of sweet and bitter perception. Physiol Behav. 2006, 88, 215–226. [Google Scholar] [CrossRef] [Green Version]
- Van Dongen, M.V.; van den Berg, M.C.; Vink, N.; Kok, F.J.; de Graaf, C. Taste–nutrient relationships in commonly consumed foods. Br. J. Nutr. 2012, 108, 140–147. [Google Scholar] [CrossRef]
- Tepper, B.J. Nutritional implications of genetic taste variation: The role of PROP sensitivity and other taste phenotypes. Annu. Rev. Nutr. 2008, 28, 367–388. [Google Scholar] [CrossRef]
- Bartoshuk, L.M. The biological basis of food perception and acceptance. Food Qual. Prefer. 1993, 4, 21–32. [Google Scholar] [CrossRef]
- Dinnella, C.; Monteleone, E.; Piochi, M.; Spinelli, S.; Prescott, J.; Pierguidi, L.; Gasperi, F.; Laureati, M.; Pagliarini, E.; Predieri, S. Individual variation in PROP status, fungiform papillae density, and responsiveness to taste stimuli in a large population sample. Chem. Senses. 2018, 43, 697–710. [Google Scholar] [CrossRef]
- Bajec, M.R.; Pickering, G.J. Thermal taste, PROP responsiveness, and perception of oral sensations. Physiol. Behav. 2008, 95, 581–590. [Google Scholar] [CrossRef]
- Bere, E.; Brug, J.; Klepp, K.I. Why do boys eat less fruit and vegetables than girls? Public Health Nutr. 2008, 11, 321–325. [Google Scholar] [CrossRef] [Green Version]
- Feeney, E.; O’brien, S.; Scannell, A.; Markey, A.; Gibney, E.R. Genetic variation in taste perception: Does it have a role in healthy eating? Proc. Nutr. Soc. 2011, 70, 135–143. [Google Scholar] [CrossRef]
- Rasmussen, M.; Krølner, R.; Klepp, K.I.; Lytle, L.; Brug, J.; Bere, E.L.; Due, P. Determinants of fruit and vegetable consumption among children and adolescents: A review of the literature. Part I: Quantitative studies. Int. J. Behav. Nutr. Phys. Act. 2006, 3, 22. [Google Scholar] [CrossRef]
- Dinehart, M.E.; Hayes, J.E.; Bartoshuk, L.M.; Lanier, S.L.; Duffy, V.B. Bitter taste markers explain variability in vegetable sweetness, bitterness, and intake. Physiol. Behav. 2006, 87, 304–313. [Google Scholar] [CrossRef]
- Drewnowski, A.; Henderson, S.A.; Shore, A.B.; Barratt-Fornell, A. Sensory responses to 6-n-propylthiouracil (PROP) or sucrose solutions and food preferences in young women. Ann. N. Y. Acad. Sci. 1998, 855, 797–801. [Google Scholar] [CrossRef]
- Jerzsa-Latta, M.; Krondl, M.; Coleman, P. Use and perceived attributes of cruciferous vegetables in terms of genetically-mediated taste sensitivity. Appetite 1990, 15, 127–134. [Google Scholar] [CrossRef]
- Yackinous, C.A.; Guinard, J.X. Relation between PROP (6-n-propylthiouracil) taster status, taste anatomy and dietary intake measures for young men and women. Appetite 2002, 38, 201–209. [Google Scholar] [CrossRef]
- Garcia-Bailo, B.; Toguri, C.; Eny, K.M.; El-Sohemy, A. Genetic variation in taste and its influence on food selection. OMICS 2009, 13, 69–80. [Google Scholar] [CrossRef]
- Tan, S.Y.; Tucker, R. Sweet Taste as a predictor of dietary intake: A systematic review. Nutrients 2019, 11, 94. [Google Scholar] [CrossRef]
- Cicerale, S.; Riddell, L.J.; Keast, R.S. The association between perceived sweetness intensity and dietary intake in young adults. J. Food Sci. 2012, 77, H31–H35. [Google Scholar] [CrossRef]
- Holt, S.H.A.; Cobiac, L.; Beaumont-Smith, N.E.; Easton, K.; Best, D.J. Dietary habits and the perception and liking of sweetness among Australian and Malaysian students: A cross-cultural study. Food Qual. Prefer. 2000, 11, 299–312. [Google Scholar] [CrossRef]
- Jayasinghe, S.N.; Kruger, R.; Walsh, D.C.; Cao, G.; Rivers, S.; Richter, M.; Bernhard, H.B. Is sweet taste perception associated with sweet food liking and intake? Nutrients 2017, 9, 750. [Google Scholar] [CrossRef]
- Low, J.Y.; Lacy, K.E.; McBride, R.; Keast, R.S. The association between sweet taste function, anthropometry, and dietary intake in adults. Nutrients 2016, 8, 241. [Google Scholar] [CrossRef]
- Mahar, A.; Duizer, L.M. The effect of frequency of consumption of artificial sweeteners on sweetness liking by women. J. Food Sci. 2007, 72, S714–S718. [Google Scholar] [CrossRef]
- Martinez-Cordero, E.; Malacara-Hernandez, J.M.; Martinez-Cordero, C. Taste perception in normal and overweight Mexican adults. Appetite 2015, 89, 192–195. [Google Scholar] [CrossRef]
- Durack, E.; Alonso-Gomez, M.; Wilkinson, M.G. Salt: A review of its role in food science and public health. Curr. Nutr. Food Sci. 2008, 4, 290–297. [Google Scholar] [CrossRef]
- Kim, G.H.; Lee, H.M. Frequent consumption of certain fast foods may be associated with an enhanced preference for salt taste. J. Hum. Nutr. Diet. 2009, 22, 475–480. [Google Scholar] [CrossRef]
- Kobayashi, C.; Kennedy, L.M.; Halpern, B.P. Experience-induced changes in taste identification of monosodium glutamate (MSG) are reversible. Chem. Senses. 2006, 31, 301–306. [Google Scholar] [CrossRef]
- Pittman, D.W.; Contreras, R.J. Dietary NaCl influences the organization of chorda tympani neurons projecting to the nucleus of the solitary tract in rats. Chem. Senses. 2002, 27, 333–341. [Google Scholar] [CrossRef]
- Wise, P.M.; Hansen, J.L.; Reed, D.R.; Breslin, P.A. Twin study of the heritability of recognition thresholds for sour and salty taste. Chem. Senses 2007, 32, 749–754. [Google Scholar] [CrossRef]
- Running, C.A.; Craig, B.A.; Mattes, R.D. Oleogustus: The unique taste of fat. Chem. Senses 2015, 40, 507–516. [Google Scholar] [CrossRef]
- Tucker, R.M.; Kaiser, K.A.; Parman, M.A.; George, B.J.; Allison, D.B.; Mattes, R.D. Comparisons of fatty acid taste detection thresholds in people who are lean vs. overweight or obese: A systematic review and meta-analysis. PLoS ONE 2017, 12, e0169583. [Google Scholar] [CrossRef]
- Keast, R.S.; Costanzo, A. Is fat the sixth taste primary? Evidence and implications. Flavour 2015, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Ruiz, N.R.; López-Díaz, J.A.; Wall-Medrano, A.; Jiménez-Castro, J.A.; Angulo, O. Oral fat perception is related with body mass index, preference and consumption of high-fat foods. Physiol. Behav. 2014, 129, 36–42. [Google Scholar] [CrossRef]
- Besnard, P.; Christensen, J.E.; Brignot, H.; Bernard, A.; Passilly-Degrace, P.; Nicklaus, S.; Pais de Barros, J.P.; Collet, X.; Lelouvier, B.; Servant, F. Obese subjects with specific gustatory papillae microbiota and salivary cues display an impairment to sense lipids. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef]
- Solemdal, K.; Sandvik, L.; Willumsen, T.; Mowe, M.; Hummel, T. The impact of oral health on taste ability in acutely hospitalized elderly. PLoS ONE 2012, 7, e36557. [Google Scholar] [CrossRef]
- Cattaneo, C.; Gargari, G.; Koirala, R.; Laureati, M.; Riso, P.; Guglielmetti, S.; Pagliarini, E. New insights into the relationship between taste perception and oral microbiota composition. Sci Rep. 2019, 9. [Google Scholar] [CrossRef]
- Hardikar, S.; Höchenberger, R.; Villringer, A.; Ohla, K. Higher sensitivity to sweet and salty taste in obese compared to lean individuals. Appetite 2017, 111, 158–165. [Google Scholar] [CrossRef]
- Proserpio, C.; Laureati, M.; Bertoli, S.; Battezzati, A.; Pagliarini, E. Determinants of obesity in Italian adults: The role of taste sensitivity, food liking, and food neophobia. Chem. Senses. 2016, 41, 169–176. [Google Scholar] [CrossRef]
- Webb, J.; Bolhuis, D.P.; Cicerale, S.; Hayes, J.E.; Keast, R. The relationships between common measurements of taste function. Chemosens Percept. 2015, 8, 11–18. [Google Scholar] [CrossRef]
- International Organisation for Standardization. Sensory analysis—Methodology—General guidance for measuring odour, flavour and taste detection thresholds by a three-alternative forced-choice (3-AFC) procedure. In ISO International Standard; N° 13301/International Organization for Standardizatio: Geneva, Switzerland, 2018; p. 28. [Google Scholar]
- Porrini, M.; Gentile, M.G.; Fidanza, F. Biochemical validation of a self-administered semi-quantitative food-frequency questionnaire. Br. J. Nutr. 1995, 74, 323–333. [Google Scholar] [CrossRef] [Green Version]
- Daly, A.M.; Parsons, J.E.; Wood, N.A.; Gill, T.K.; Taylor, A.W. Food consumption habits in two states of Australia, as measured by a Food Frequency Questionnaire. BMC Res. Notes. 2011, 4, 507. [Google Scholar] [CrossRef]
- Drayna, D. Human taste genetics. Annu. Rev. Genomics Hum. Genet. 2005, 6, 217–235. [Google Scholar] [CrossRef]
- Hladik, C.M.; Pasquet, P.; Simmen, B. New perspectives on taste and primate evolution: The dichotomy in gustatory coding for perception of beneficent versus noxious substances as supported by correlations among human thresholds. Am. J. Phys. Anthropol. 2002, 117, 342–348. [Google Scholar] [CrossRef]
- Bertoli, S.; Laureati, M.; Battezzati, A.; Bergamaschi, V.; Cereda, E.; Spadafranca, A.; Laila, V.; Ella, P. Taste sensitivity, nutritional status and metabolic syndrome: Implication in weight loss dietary interventions. World J. Diabetes. 2014, 5, 717. [Google Scholar] [CrossRef]
- Donaldson, L.F.; Bennett, L.; Baic, S.; Melichar, J.K. Taste and weight: Is there a link? Am. J. Clin. Nutr. 2009, 90, S800–S803. [Google Scholar] [CrossRef]
- Proserpio, C.; Laureati, M.; Invitti, C.; Pagliarini, E. Reduced taste responsiveness and increased food neophobia characterize obese adults. Food Qual. Prefer. 2018, 63, 73–79. [Google Scholar] [CrossRef]
- Hayes, J.E.; Sullivan, B.S.; Duffy, V.B. Explaining variability in sodium intake through oral sensory phenotype, salt sensation and liking. Physiol. Behav. 2010, 100, 369–380. [Google Scholar] [CrossRef] [Green Version]
- Mattes, R.D. The taste for salt in humans. Am. J. Clin. Nutr. 1997, 65, 692S–697S. [Google Scholar] [CrossRef] [Green Version]
- Pangborn, R.M.; Pecore, S.D. Taste perception of sodium chloride in relation to dietary intake of salt. Am. J. Clin. Nutr. 1982, 35, 510–520. [Google Scholar] [CrossRef] [Green Version]
- Noel, C.A.; Sugrue, M.; Dando, R. Participants with pharmacologically impaired taste function seek out more intense, higher calorie stimuli. Appetite 2017, 117, 74–81. [Google Scholar] [CrossRef]
- Stevenson, R.J.; Boakes, R.A.; Oaten, M.J.; Yeomans, M.R.; Mahmut, M.; Francis, H.M. Chemosensory abilities in consumers of a western-style diet. Chem. Senses. 2016, 41, 505–513. [Google Scholar] [CrossRef]
- Törnwall, O.; Silventoinen, K.; Keskitalo-Vuokko, K.; Perola, M.; Kaprio, J.; Tuorila, H. Genetic contribution to sour taste preference. Appetite 2012, 58, 687–694. [Google Scholar] [CrossRef]
- Fagerli, R.A.; Wandel, M. Gender differences in opinions and practices with regard to a ‘healthy diet’. Appetite 1999, 32, 171–190. [Google Scholar] [CrossRef]
- Leblanc, V.; Bégin, C.; Corneau, L.; Dodin, S.; Lemieux, S. Gender differences in dietary intakes: What is the contribution of motivational variables? J. Hum. Nutr. Diet. 2015, 28, 37–46. [Google Scholar] [CrossRef]
- Li, K.K.; Concepcion, R.Y.; Lee, H.; Cardinal, B.J.; Ebbeck, V.; Woekel, E.; Readdy, R.T. An examination of sex differences in relation to the eating habits and nutrient intakes of university students. J. Nutr. Educ. Behav. 2012, 44, 246–250. [Google Scholar] [CrossRef]
- Rolls, B.J.; Fedoroff, I.C.; Guthrie, J.F. Gender differences in eating behavior and body weight regulation. Health Psychol. 1991, 10, 133–142. [Google Scholar] [CrossRef]
- Dewhirst, F.E.; Chen, T.; Izard, J.; Paster, B.J.; Tanner, A.C.; Yu, W.H.; Lakshmanan, A.; Wade, W.G. The human oral microbiome. J. Bacteriol. 2010, 192, 5002–5017. [Google Scholar] [CrossRef]
- Alcock, J.; Maley, C.C.; Aktipis, C.A. Is eating behavior manipulated by the gastrointestinal microbiota? Evolutionary pressures and potential mechanisms. Bioessays 2014, 36, 940–949. [Google Scholar] [CrossRef]
- Duca, F.A.; Swartz, T.D.; Sakar, Y.; Covasa, M. Increased oral detection, but decreased intestinal signaling for fats in mice lacking gut microbiota. PLoS ONE 2012, 7, e39748. [Google Scholar] [CrossRef]
- Swartz, T.D.; Duca, F.A.; De Wouters, T.; Sakar, Y.; Covasa, M. Up-regulation of intestinal type 1 taste receptor 3 and sodium glucose luminal transporter-1 expression and increased sucrose intake in mice lacking gut microbiota. Br. J. Nutr. 2012, 107, 621–630. [Google Scholar] [CrossRef]
- Takahashi, N. Oral microbiome metabolism: From “who are they?” to “what are they doing?”. J. Dent. Res. 2015, 94, 1628–1637. [Google Scholar] [CrossRef]
- Feng, Y.; Licandro, H.; Martin, C.; Septier, C.; Zhao, M.; Neyraud, E.; Martine, M. The associations between biochemical and microbiological variables and taste differ in whole saliva and in the film lining the tongue. BioMed. Res. Int. 2018. [Google Scholar] [CrossRef]
- Jeffery, I.B.; O’Toole, P.W. Diet-microbiota interactions and their implications for healthy living. Nutrients 2013, 5, 234–252. [Google Scholar] [CrossRef]
- Albenberg, L.G.; Wu, G.D. Diet and the intestinal microbiome: Associations, functions, and implications for health and disease. Gastroenterol 2014, 146, 1564–1572. [Google Scholar] [CrossRef]
- Olsen, I.; Yamazaki, K. Can oral bacteria affect the microbiome of the gut? J. Oral Microbiol. 2019, 11, 1586422. [Google Scholar] [CrossRef] [Green Version]
- Segata, N.; Haake, S.K.; Mannon, P.; Lemon, K.P.; Waldron, L.; Gevers, D.; Huttenhower, C.; Izard, J. Composition of the adult digestive tract bacterial microbiome based on seven mouth surfaces, tonsils, throat and stool samples. Genome. Biol. 2012, 13, R42. [Google Scholar] [CrossRef]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ridaura, V.K.; Faith, J.J.; Rey, F.E.; Knight, R.; Gordon, J.I. The effect of diet on the human gut microbiome: A metagenomic analysis in humanized gnotobiotic mice. Sci. Transl. Med. 2009, 1, 6ra14. [Google Scholar] [CrossRef]
- Kretsch, M.J.; Fong, A.K.; Green, M.W. Behavioral and body size correlates of energy intake underreporting by obese and normal-weight women. J. Am. Diet. Assoc. 1999, 99, 300–306. [Google Scholar] [CrossRef]


{kind=link}
| Taste Quality | Reference Stimuli | Sample Concentration (g/L) a | ||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | ||
| Sweet | Sucrose | 1.6 × 10−1 | 4.0 × 10−1 | 1.02 | 2.56 | 6.4 | 16.0 | 40.0 |
| Salty | Sodium chloride | 6.25 × 10−2 | 1.25 × 10−2 | 2.5 × 10−1 | 5.0 × 10−1 | 1.0 | 2.0 | 4.0 |
| Bitter | Caffeine | 3.0 × 10−3 | 9.0 × 10−3 | 3.0 × 10−2 | 8.0 × 10−2 | 2.4 × 10−1 | 8.0 × 10−1 | 2.0 |
| Sour | Citric acid | 2.0 × 10−2 | 5.0 × 10−2 | 8.0 × 10−2 | 1.5 × 10−1 | 3.5 × 10−1 | 7.5 × 10−1 | 1.5 |
| Original Frequency used in FFQ | Daily Equivalent Frequency |
|---|---|
| Less than once per month | 0.02 |
| 1–3 times per month | 0.07 |
| 1–4 times per week | 0.43 |
| 5–7 times per week | 0.86 |
| 2–4 times per day | 3.00 |
| 5 or more times per day | 5.00 |
| Mean | SEM | |
|---|---|---|
| Age (years) | 23.3 | 0.3 |
| BMI (kg/m2) | 21.6 | 0.3 |
| Gustatory Functions | ||
| Sweet threshold (g/L) | 3.61 | 0.62 |
| Salty threshold (g/L) | 0.20 | 0.02 |
| Bitter threshold (g/L) | 0.16 | 0.04 |
| Sour threshold (g/L) | 0.09 | 0.01 |
| Food Intake | ||
| Total Energy (kcal) | 1829 | 60 |
| Protein (%) a | 15.6 | 0.4 |
| Fat (%) a | 35.2 | 0.9 |
| Carbohydrates (%) a | 45.2 | 0.7 |
| Protein (g) b | 68.0 | 2.3 |
| Fat (g) b | 70.1 | 2.7 |
| Carbohydrates (g) b | 216.0 | 8.6 |
| Total Fiber (g) b | 15.1 | 6.7 |
| Items | Daily Equivalent Frequency | Daily Equivalent Frequency | ||||||
|---|---|---|---|---|---|---|---|---|
| p value | Salty Taste Sensitivity Level | p value | Sweet Taste Sensitivity Level | |||||
| Hyper | Normal | Hypo | Hyper | Normal | Hypo | |||
| Cereal and cereal-derived products (e.g., pasta, rice, barley, spelt) | 0.07 | 1.67 | 1.33 | 2.23 | 0.88 | 1.92 | 1.58 | 1.49 |
| Salty baked products (e.g., bread, pizza, focaccia) | 0.007 | 0.99 b | 1.45 ab | 1.99 a | 0.69 | 1.37 | 1.12 | 1.40 |
| Bakery products (e.g., bakery and breakfast cereals, biscuits, croissants) | 0.04 | 0.76 b | 1.39 a | 1.42 a | 0.48 | 1.12 | 1.34 | 0.94 |
| Meats | 0.98 | 0.50 | 0.51 | 0.49 | 0.85 | 0.48 | 0.54 | 0.50 |
| Cured meats | 0.64 | 0.36 | 0.40 | 0.31 | 0.09 | 0.27 | 0.48 | 0.37 |
| Fish | 0.16 | 0.41 | 0.28 | 0.43 | 0.35 | 0.39 | 0.28 | 0.39 |
| Milk and yoghurts | 0.47 | 0.84 | 0.75 | 1.11 | 0.27 | 0.75 | 0.61 | 1.04 |
| Dairy products | 0.32 | 0.51 | 0.79 | 0.79 | 0.19 | 0.52 | 0.62 | 0.86 |
| Eggs | 0.22 | 0.34 | 0.23 | 0.29 | 0.72 | 0.33 | 0.27 | 0.30 |
| Vegetables | 0.41 | 1.80 | 1.85 | 2.40 | 0.08 | 2.62 | 1.72 | 1.71 |
| Legumes | 0.05 | 0.36 b | 0.67a | 0.41 ab | 0.03 | 0.69 a | 0.41 ab | 0.35 b |
| Potatoes | 0.20 | 0.35 | 0.58 | 0.47 | 0.65 | 0.49 | 0.35 | 0.41 |
| Fruit | 0.22 | 1.65 | 1.84 | 2.42 | 0.79 | 2.07 | 1.78 | 1.78 |
| Fruit juices | 0.31 | 0.22 | 0.43 | 0.48 | 0.80 | 0.40 | 0.26 | 0.36 |
| Nuts | 0.27 | 0.23 | 0.45 | 0.36 | 0.76 | 0.33 | 0.22 | 0.33 |
| Sweets and desserts (e.g., cakes, ice creams, chocolate) | 0.53 | 0.60 | 0.86 | 0.78 | 0.02 | 0.32 b | 0.98 a | 0.87 a |
| Fats | 0.05 | 0.26 b | 0.22 b | 0.45 a | 0.18 | 0.19 | 0.35 | 0.34 |
| Oils | 0.47 | 1.99 | 1.54 | 1.72 | 0.87 | 1.83 | 1.94 | 1.74 |
| Salty snacks (e.g., chips, salty peanuts) | 0.23 | 0.22 | 0.22 | 0.35 | 0.23 | 0.17 | 0.27 | 0.30 |
| Alcoholic beverages | 0.21 | 0.49 | 0.29 | 0.31 | 0.18 | 0.30 | 0.56 | 0.35 |
| Soft drinks | 0.007 | 0.26 b | 1.13 a | 0.16 b | 0.82 | 0.59 | 0.39 | 0.38 |
| Candies and gums | 0.82 | 0.63 | 0.54 | 0.78 | 0.32 | 0.82 | 0.81 | 0.40 |
| Items | Daily Equivalent Frequency | Daily Equivalent Frequency | ||||||
|---|---|---|---|---|---|---|---|---|
| p value | Bitter Taste Sensitivity Level | p value | Sour Taste Sensitivity Level | |||||
| Hyper | Normal | Hypo | Hyper | Normal | Hypo | |||
| Cereal and cereal-derived products (e.g., pasta, rice, barley, spelt) | 0.58 | 1.75 | 1.29 | 1.66 | 0.08 | 2.19 | 1.31 | 1.78 |
| Salty baked products (e.g., bread, pizza, focaccia) | 0.70 | 1.45 | 1.25 | 1.20 | 0.11 | 0.90 | 1.43 | 1.66 |
| Bakery products (e.g., bakery and breakfast cereals, biscuits, croissants) | 0.81 | 0.92 | 1.10 | 1.10 | 0.21 | 0.69 | 1.20 | 1.29 |
| Meats | 0.65 | 0.47 | 0.48 | 0.54 | 0.75 | 0.53 | 0.51 | 0.46 |
| Cured meats | 0.41 | 0.34 | 0.46 | 0.33 | 0.37 | 0.35 | 0.40 | 0.29 |
| Fish | 0.22 | 0.42 | 0.26 | 0.36 | 0.004 | 0.49 a | 0.27 b | 0.46 a |
| Milk and yoghurt | 0.66 | 0.72 | 0.97 | 0.92 | 0.96 | 0.91 | 0.84 | 0.82 |
| Dairy products | 0.07 | 0.52 | 0.52 | 0.95 | 0.55 | 0.49 | 0.75 | 0.69 |
| Eggs | 0.52 | 0.33 | 0.24 | 0.30 | 0.54 | 0.36 | 0.31 | 0.27 |
| Vegetables | 0.08 | 1.92 | 1.04 | 2.23 | 0.33 | 1.84 | 1.77 | 2.39 |
| Legumes | 0.51 | 0.53 | 0.42 | 0.38 | 0.09 | 0.32 | 0.61 | 0.40 |
| Potatoes | 0.53 | 0.49 | 0.39 | 0.35 | 0.23 | 0.61 | 0.42 | 0.36 |
| Fruit | 0.76 | 2.06 | 1.70 | 1.81 | 0.94 | 1.77 | 1.94 | 1.94 |
| Fruit juices | 0.24 | 0.48 | 0.39 | 0.19 | 0.70 | 0.22 | 0.31 | 0.39 |
| Nuts | 0.42 | 0.43 | 0.26 | 0.26 | 0.41 | 0.43 | 0.23 | 0.36 |
| Sweets and desserts (e.g., cakes, ice creams, chocolate) | 0.85 | 0.73 | 0.61 | 0.78 | 0.44 | 0.46 | 0.75 | 0.79 |
| Fats | 0.84 | 0.29 | 0.29 | 0.25 | 0.26 | 0.26 | 0.36 | 0.22 |
| Oils | 0.007 | 1.46 b | 1.11 b | 2.22 a | 0.35 | 1.64 | 1.72 | 2.16 |
| Salty snacks (e.g., chips, salty peanuts) | 0.40 | 0.24 | 0.20 | 0.32 | 0.44 | 0.18 | 0.28 | 0.22 |
| Alcoholic beverages | 0.34 | 0.46 | 0.33 | 0.29 | 0.70 | 0.31 | 0.43 | 0.37 |
| Soft drinks | 0.22 | 0.68 | 0.20 | 0.16 | 0.81 | 0.40 | 0.56 | 0.36 |
| Candies and gums | 0.91 | 0.68 | 0.73 | 0.57 | 0.76 | 0.48 | 0.63 | 0.78 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cattaneo, C.; Riso, P.; Laureati, M.; Gargari, G.; Pagliarini, E. Exploring Associations between Interindividual Differences in Taste Perception, Oral Microbiota Composition, and Reported Food Intake. Nutrients 2019, 11, 1167. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11051167
Cattaneo C, Riso P, Laureati M, Gargari G, Pagliarini E. Exploring Associations between Interindividual Differences in Taste Perception, Oral Microbiota Composition, and Reported Food Intake. Nutrients. 2019; 11(5):1167. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11051167
Chicago/Turabian StyleCattaneo, Camilla, Patrizia Riso, Monica Laureati, Giorgio Gargari, and Ella Pagliarini. 2019. "Exploring Associations between Interindividual Differences in Taste Perception, Oral Microbiota Composition, and Reported Food Intake" Nutrients 11, no. 5: 1167. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11051167