The Preventive Effects of Greenshell Mussel (Perna canaliculus) on Early-Stage Metabolic Osteoarthritis in Rats with Diet-Induced Obesity

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Analysis of GSM Composition
2.2. Animal Study
2.3. Plasma Analysis
2.4. Histopathological Assessment
2.5. Data Analysis
3. Results
3.1. GSM Composition
3.2. Animal Study
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Briggs, A.M.; Cross, M.J.; Hoy, D.G.; Sanchez-Riera, L.; Blyth, F.M.; Woolf, A.D.; March, L. Musculoskeletal health conditions represent a global threat to healthy aging: A report for the 2015 world health organization world report on ageing and health. Gerontologist 2016, 243–255. [Google Scholar] [CrossRef] [PubMed]
- Cicuttini, F.M.; Baker, J.R.; Spector, T.D. The association of obesity with osteoarthritis of the hand and knee in women: A twin study. J. Rheumatol. 1996, 23, 1221–1226. [Google Scholar] [PubMed]
- Gregor, M.F.; Hotamisligil, G.S. Inflammatory mechanisms in obesity. Annu. Rev. Immunol. 2011, 29, 415–445. [Google Scholar] [CrossRef] [PubMed]
- Collins, K.H.; Reimer, R.A.; Seerattan, R.A.; Herzog, W.; Hart, D.A. Response to diet-induced obesity produces time-dependent induction and progression of metabolic osteoarthritis in rat knees. J. Orthop. Res. 2016, 34, 1010–1018. [Google Scholar] [CrossRef] [PubMed]
- Eun-Young, K.; Su-Kyung, S.; Yun-Young, C.; Un Ju, J.; Eunjung, K.; Taesun, P.; Jung Han Yoon, P.; Jong Won, Y.; McGregor, R.A.; Yong Bok, P.; et al. Time-course microarrays reveal early activation of the immune transcriptome and adipokine dysregulation leads to fibrosis in visceral adipose depots during diet-induced obesity. BMC Genom. 2012, 13, 450–465. [Google Scholar] [CrossRef]
- Barboza, E.; Hudson, J.; Chang, W.-P.; Kovats, S.; Towner, R.A.; Silasi-Mansat, R.; Lupu, F.; Kent, C.; Griffin, T.M. Profibrotic Infrapatellar Fat Pad Remodeling Without M1 Macrophage Polarization Precedes Knee Osteoarthritis in Mice with Diet-Induced Obesity. Arthritis Rheumatol. 2017, 69, 1221–1232. [Google Scholar] [CrossRef] [PubMed]
- Pearson, M.J.; Herndler-Brandstetter, D.; Tariq, M.A.; Nicholson, T.A.; Philp, A.M.; Smith, H.L.; Davis, E.T.; Jones, S.W.; Lord, J.M. IL-6 secretion in osteoarthritis patients is mediated by chondrocyte-synovial fibroblast cross-talk and is enhanced by obesity. Sci. Rep. 2017, 7, 3451–3461. [Google Scholar] [CrossRef] [PubMed]
- Teichtahl, A.J.; Wluka, A.E.; Proietto, J.; Cicuttini, F.M. Obesity and the female sex, risk factors for knee osteoarthritis that may be attributable to systemic or local leptin biosynthesis and its cellular effects. Med. Hypotheses 2005, 65, 312–315. [Google Scholar] [CrossRef]
- Treschow, A.P.; Hodges, L.D.; Kalafatis, N.; Macrides, T.A.; Wright, P.F.A.; Wynne, P.M. Novel anti-inflammatory ω-3 PUFAs from the New Zealand green-lipped mussel, Perna canaliculus. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2007, 147, 645–656. [Google Scholar] [CrossRef]
- Whitehouse, M.W.; Macrides, T.A.; Kalafatis, N.; Betts, W.H.; Haynes, D.R.; Broadbent, J. Anti-inflammatory activity of a lipid fraction (Lyprinol) from the NZ green-lipped mussel. Inflammopharmacology 1997, 5, 237–246. [Google Scholar] [CrossRef]
- McPhee, S.; Hodges, L.D.; Wright, P.F.A.; Wynne, P.M.; Kalafatis, N.; Harney, D.W.; Macrides, T.A. Anti-cyclooxygenase effects of lipid extracts from the New Zealand green-lipped mussel, Perna canaliculus. Comp. Biochem. Physiol. B 2007, 146, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Bao, C.; Cho, S.H.; Lee, H.J. Green lipped mussel oil complex suppresses lipopolysaccharide stimulated inflammation via regulating nuclear factor-kappa B and mitogen activated protein kinases signaling in RAW264.7 murine macrophages. Food Sci. Biotechnol. 2017, 26, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Chi-Ho, L.; John Hon-Kei, L.; Curtise Kin-Cheung, N.; Janice, M.; Yoki Kwok-Chu, B.; Man-Sau, W.; Samuel Chun-Lap, L. Pain Controlling and Cytokine-regulating Effects of Lyprinol, a Lipid Extract of Perna Canaliculus, in a Rat Adjuvant-induced Arthritis Model. Evid.-Based Complement. Altern. Med. 2009, 6, 239–245. [Google Scholar] [CrossRef]
- Wakimoto, T.; Kondo, H.; Nii, H.; Kimura, K.; Egami, Y.; Oka, Y.; Yoshida, M.; Kida, E.; Yiping, Y.; Akahoshi, S.; et al. Furan fatty acid as an anti-inflammatory component from the green-lipped mussel Perna canaliculus. Proc. Natl. Acad. Sci. USA 2011, 108, 17533–17537. [Google Scholar] [CrossRef] [PubMed]
- Kruger, M.C.; Morel, P.C.H. Experimental Control for the Ovariectomized Rat Model: Use of Sham Versus Nonmanipulated Animal. J. Appl. Anim. Welf. Sci. 2016, 19, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Mankin, H.J.; Dorfman, H.; Lippiello, L.; Zarins, A. Biochemical and metabolic abnormalities in articular cartilage from osteo-arthritic human hips. II. Correlation of morphology with biochemical and metabolic data. J. Bone Jt. Surg. Am. 1971, 53, 523–537. [Google Scholar] [CrossRef]
- Collins, K.H.; Paul, H.A.; Reimer, R.A.; Seerattan, R.A.; Hart, D.A.; Herzog, W. Relationship between inflammation, the gut microbiota, and metabolic osteoarthritis development: Studies in a rat model. Osteoarthr. Cartil. 2015, 23, 1989–1998. [Google Scholar] [CrossRef]
- Sun, A.R.; Panchal, S.K.; Friis, T.; Sekar, S.; Crawford, R.; Brown, L.; Xiao, Y.; Prasadam, I. Obesity-associated metabolic syndrome spontaneously induces infiltration of pro-inflammatory macrophage in synovium and promotes osteoarthritis. PLoS ONE 2017, 12, e0183693. [Google Scholar] [CrossRef]
- de Visser, H.M.; Kozijn, A.E.; Pouran, B.; van Rijen, M.H.; Weinans, H.; Mastbergen, S.C.; Coeleveld, K.; Lafeber, F.P.J.G. Metabolic dysregulation accelerates injury-induced joint degeneration, driven by local inflammation; an in vivo rat study. J. Orthop. Res. 2017. [Google Scholar] [CrossRef]
- Sekar, S.; Shafie, S.R.; Prasadam, I.; Crawford, R.; Panchal, S.K.; Brown, L.; Xiao, Y. Saturated fatty acids induce development of both metabolic syndrome and osteoarthritis in rats. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Ma, H.L.; Blanchet, T.J.; Peluso, D.; Hopkins, B.; Morris, E.A.; Glasson, S.S. Osteoarthritis severity is sex dependent in a surgical mouse model. Osteoarthr. Cartil. 2007, 15, 695–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ràfols, M.E. Adipose tissue: Cell heterogeneity and functional diversity. Endocrinol. Nutr. 2014, 61, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Large, V.; Peroni, O.; Letexier, D.; Ray, H.; Beylot, M. Metabolism of lipids in human white adipocyte. Diabetes Metab. 2004, 30, 294–309. [Google Scholar] [CrossRef]
- BrahmaNaidu, P.; Nemani, H.; Meriga, B.; Mehar, S.K.; Potana, S.; Ramgopalrao, S. Mitigating efficacy of piperine in the physiological derangements of high fat diet induced obesity in Sprague Dawley rats. Chem. Biol. Interact. 2014, 221, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Kopp, H.P.; Krzyzanowska, K.; Mohlig, M.; Spranger, J.; Pfeiffer, A.F.H.; Schernthaner, G. Effects of marked weight loss on plasma levels of adiponectin, markers of chronic subclinical inflammation and insulin resistance in morbidly obese women. Int. J. Obes. 2005, 29, 766–771. [Google Scholar] [CrossRef] [PubMed]
- Ashley, D.T.; O’Sullivan, E.P.; Davenport, C.; Devlin, N.; Crowley, R.K.; McCaffrey, N.; Moyna, N.M.; Smith, D.; O’Gorman, D.J. Similar to adiponectin, serum levels of osteoprotegerin are associated with obesity in healthy subjects. Metabolism 2011, 60, 994–1000. [Google Scholar] [CrossRef]
- Reverchon, M.; Rame, C.; Bertoldo, M.; Dupont, J. Adipokines and the Female Reproductive Tract. Int. J. Endocrinol. 2014, 2014, 232454. [Google Scholar] [CrossRef]
- Azamar-Llamas, D.; Hernandez-Molina, G.; Ramos-Avalos, B.; Furuzawa-Carballeda, J. Adipokine contribution to the pathogenesis of osteoarthritis. Mediat. Inflamm. 2017, 2017, 5468023. [Google Scholar] [CrossRef]
- Viney, M.; Riley, E.M. The Immunology of Wild Rodents: Current Status and Future Prospects. Front. Immunol. 2017, 8, 1481. [Google Scholar] [CrossRef]
- Ferreira, M.R.; Alvarez, S.M.; Illesca, P.; Giménez, M.S.; Lombardo, Y.B. Dietary Salba (Salvia hispanica L.) ameliorates the adipose tissue dysfunction of dyslipemic insulin-resistant rats through mechanisms involving oxidative stress, inflammatory cytokines and peroxisome proliferator-activated receptor γ. Eur. J. Nutr. 2018, 57, 83–94. [Google Scholar] [CrossRef]
- Lu, Y.; Ho, C.S.; Liu, X.; Chua, A.N.; Wang, W.; McIntyre, R.S.; Ho, R.C. Chronic administration of fluoxetine and pro-inflammatory cytokine change in a rat model of depression. PLoS ONE 2017, 12, e0186700. [Google Scholar] [CrossRef] [PubMed]
- Chin, K.-Y.; Ima-Nirwana, S. The Effects of Testosterone Deficiency and Its Replacement on Inflammatory Markers in Rats: A Pilot Study. Int. J. Endocrinol. Metab. 2017, 15, e43053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calder, P.C.; Ahluwalia, N.; Albers, R.; Bosco, N.; Bourdet-Sicard, R.; Haller, D.; Holgate, S.T.; Jo¨nsson, L.S.; Latulippe, M.E.; Marcos, A.; et al. A consideration of biomarkers to be used for evaluation of inflammation in human nutrition. Br. J. Nutr. 2013, 109, S1–S34. [Google Scholar] [CrossRef] [PubMed]
- Fagiolo, U.; Cossarizza, A.; Scala, E.; Fanales-Belasio, E.; Ortolani, C.; Cozzi, E.; Monti, D.; Franceschi, C.; Paganelli, R. Increased cytokine production in mononuclear cells of healthy elderly people. Eur. J. Immunol. 1993, 23, 2375–2378. [Google Scholar] [CrossRef] [PubMed]
- Pietschmann, P.; Gollob, E.; Brosch, S.; Hahn, P.; Kudlacek, S.; Willheim, M.; Woloszczuk, W.; Peterlik, M.; Tragl, K.H. The effect of age and gender on cytokine production by human peripheral blood mononuclear cells and markers of bone metabolism. Exp. Gerontol. 2003, 38, 1119–1127. [Google Scholar] [CrossRef]
- Kemp, G.J.; Birrell, F.; Clegg, P.D.; Cuthbertson, D.J.; De Vito, G.; van Dieën, J.H.; Del Din, S.; Eastell, R.; Garnero, P.; Goljanek-Whysall, K.; et al. Developing a toolkit for the assessment and monitoring of musculoskeletal ageing. Age Ageing 2018, 47, iv1–iv19. [Google Scholar] [CrossRef] [PubMed]
- Garnero, P.; Conrozier, T.; Christgau, S.; Mathieu, P.; Delmas, P.D.; Vignon, E. Urinary type II collagen C-telopeptide levels are increased in patients with rapidly destructive hip osteoarthritis. Ann. Rheum. Dis. 2003, 62, 939–943. [Google Scholar] [CrossRef] [Green Version]
- Khan, H.M.; Ashraf, M.; Hashmi, A.S.; Ahmad, M.-u.-D.; Anjum, A.A. Relationship between Disease Activity and Circulating Level of Collagen II C-Telopeptide Fragments in Papain Induced Osteoarthritis Rat Model. Pak. Vet. J. 2014, 34, 92–95. [Google Scholar]
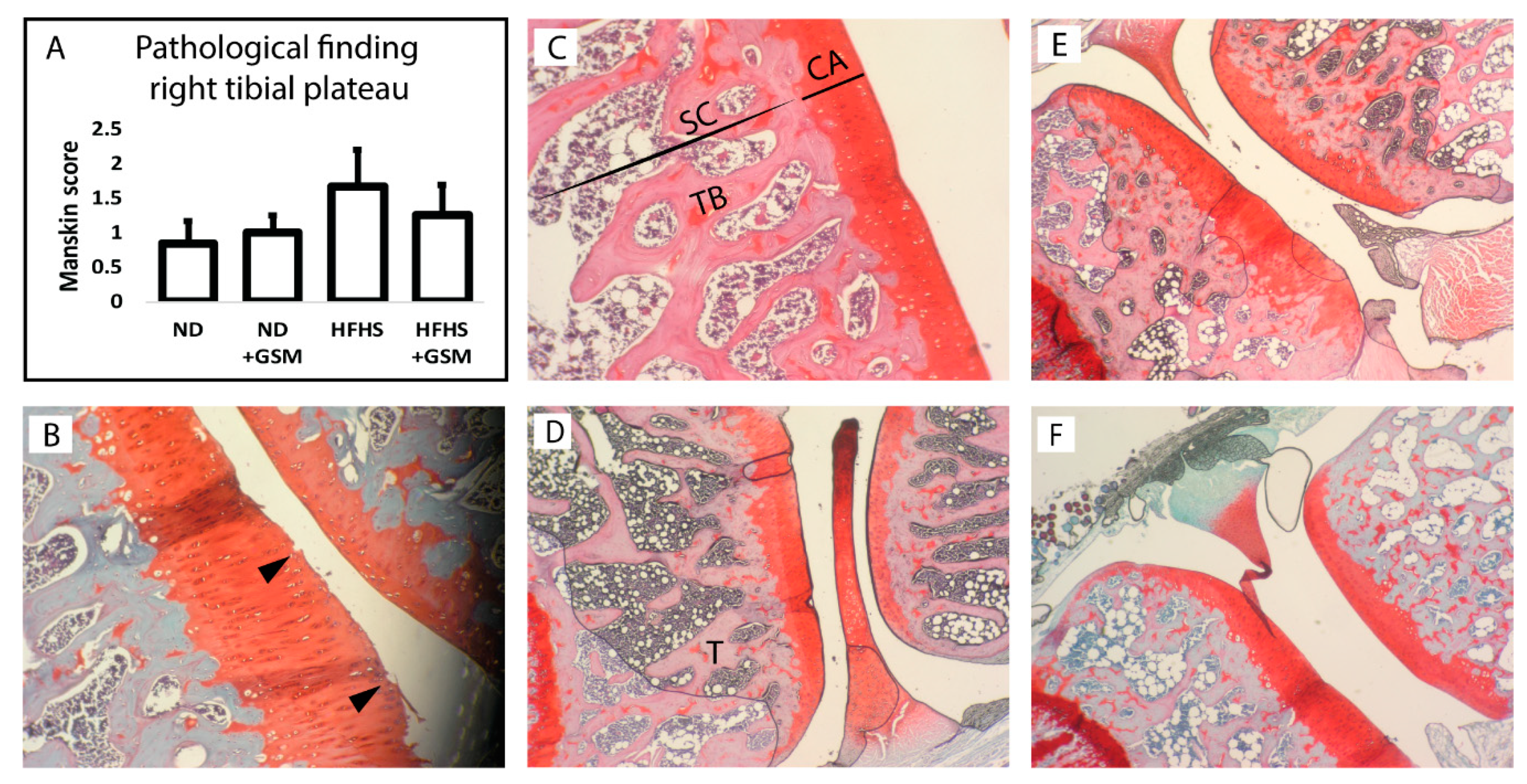
- Murat, N.; Karadam, B.; Ozkal, S.; Karatosun, V.; Gidener, S. Quantification of papain-induced rat osteoarthritis in relation to time with the Mankin score. Acta Orthop. Traumatol. Turc. 2007, 41, 233–237. [Google Scholar]
- Tortosa-Caparrós, E.; Marín, F.; Orenes-Piñero, E.; Navas-Carrillo, D. Anti-inflammatory effects of omega 3 and omega 6 polyunsaturated fatty acids in cardiovascular disease and metabolic syndrome. Crit. Rev. Food Sci. Nutr. 2017, 57, 3421–3429. [Google Scholar] [CrossRef]
- Layé, S.; Nadjar, A.; Joffre, C.; Bazinet, R.P. Anti-inflammatory effects of omega-3 fatty acids in the brain: Physiological mechanisms and relevance to pharmacology. Pharmacol. Rev. 2018, 70, 12–38. [Google Scholar] [CrossRef] [PubMed]
- Teitelbaum, J.E.; Allan Walker, W. Review: The role of omega 3 fatty acids in intestinal inflammation. J. Nutr. Biochem. 2001, 12, 21–32. [Google Scholar] [CrossRef]
- Wall, R.; Ross, R.P.; Stanton, C.; Fitzgerald, G.F. Fatty acids from fish: The anti-inflammatory potential of long-chain omega-3 fatty acids. Nutr. Rev. 2010, 68, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Rajaei, E.; Mowla, K.; Ghorbani, A.; Bahadoram, S.; Bahadoram, M.; Dargahi-Malamir, M. The Effect of Omega-3 Fatty Acids in Patients with Active Rheumatoid Arthritis Receiving DMARDs Therapy: Double-Blind Randomized Controlled Trial. Glob. J. Health Sci. 2015, 8, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Song, G.G.; Bae, S.C. Omega-3 Polyunsaturated Fatty Acids and the Treatment of Rheumatoid Arthritis: A Meta-analysis. Arch. Med. Res. 2012, 43, 356–362. [Google Scholar] [CrossRef]
- Norling, L.V.; Perretti, M. The role of omega-3 derived resolvins in arthritis. Curr. Opin. Pharmacol. 2013. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Proximate composition (g/100 g) | |
| Fat | 8.1 |
| Crude protein | 43 |
| Carbohydrate | 21.9 |
| Moisture | 5.8 |
| Ash | 21.2 |
| Fatty acid profile (% fatty acids) | |
| C14:0 myristic acid | 5.9 |
| C16:0 palmitic acid | 20.3 |
| C17:0 heptadecanoic acid | 1.2 |
| C18:0 stearic acid | 4.9 |
| C18:1n7 vaccenic acid | 3.2 |
| C18:1n9c oleic acid | 2.1 |
| C18:2n6c linoleic acid | 2.1 |
| C18:3n3 α-linolenic acid (ALA) | 1.4 |
| C18:3n4 octadecatrienoic acid | 1.3 |
| C18:4n3 stearidonic acid (SDA) | 2.5 |
| C20:1 gadoleic acid | 2.6 |
| C20:4n6 arachidonic acid (AA) | 1.0 |
| C20:5n3 eicosapentaenoic acid (EPA) | 13.5 |
| C22:5n3 docosapentaenoic acid (DPA) | 1.0 |
| C22:6n3 docosahexaenoic acid (DHA) | 10.7 |
| ∑SFA | 34.29 |
| ∑MUFA | 8.23 |
| ∑PUFA | 34.20 |
| ∑n-3 PUFA | 29.4 |
| ∑n-6 PUFA | 3.5 |
| Weight (g) | ND | ND + GSM | HFHS | HFHS + GSM | p-Value |
|---|---|---|---|---|---|
| BW (week 0) | 277.14 ± 8.24 | 280.47 ± 10.48 | 280.78 ± 6.53 | 276.28 ± 9.43 | NS |
| BW (week 13) | 343.00 ± 45.70a | 351.80 ± 51.00a | 400.80 ± 50.43b | 417.2 ± 88.08b | 0.010 |
| % BW gain | 24.05 ± 7.95a | 25.44 ± 8.62a | 42.42 ± 9.56b | 48.22 ± 15.72b | <0.001 |
| %body fat (week 0) | 11.16 ± 3.48 | 11.36 ± 4.62 | 11.24 ± 3.64 | 12.83 ± 4.03 | NS |
| %body fat (week 13) | 21.2 ± 5.43a | 22.88 ± 7.90a | 34.39 ± 4.80b | 36.46 ± 7.52b | <0.001 |
| %body fat gain | 101.13 ± 65.10a | 112.48 ± 60.96a | 232.89 ± 98.82b | 193.91 ± 51.78b | <0.001 |
| Lean mass (week 0) | 237.68 ± 22.77 | 236.28 ± 27.05 | 234.44 ± 20.73 | 227.39 ± 21.92 | NS |
| Lean mass (week 13) | 269.62 ± 32.55 | 275.29 ± 29.86 | 266.68 ± 26.49 | 266.04 ± 29.97 | NS |
| % lean mass gain | 13.34 ± 6.52 | 16.89 ± 8.10 | 13.77 ± 6.30 | 17.01 ± 6.54 | NS |
| Retroperitoneal | 9.05 ± 3.90a | 11.14 ± 5.89a | 19.28 ± 5.01b | 18.81 ± 5.10b | <0.001 |
| Epididymal | 8.80 ± 3.04a | 9.04 ± 3.17a | 15.31 ± 5.25b | 17.90 ± 5.73b | <0.001 |
| Inguinal | 5.23 ± 2.87a | 4.98 ± 4.42a | 7.95 ± 3.95a | 10.71 ± 8.09b | 0.023 |
| Interscapular | 0.83 ± 0.18a | 0.90 ± 0.23a | 0.97 ± 0.19a | 1.10 ± 0.31b | 0.033 |
| Analysts | ND | ND + GSM | HFHS | HFHS + GSM | p-Value |
|---|---|---|---|---|---|
| IL-1β (pg/mL) | 23.90 ± 39.34 | 4.93 ± 7.45 | 6.95 ± 15.23 | 13.46 ± 36.93 | NS |
| IL-6 (pg/mL) | 21.90 ± 19.23 | 29.40 ± 19.67 | 16.23 ± 22.27 | 19.56 ± 20.27 | NS |
| IL-10 (pg/mL) | 3.36 ± 7.16 | 7.33 ± 13.11 | 14.31 ± 38.74 | 2.39 ± 5.03 | NS |
| TNF-α (pg/mL) | 0.16 ± 0.58 | 0.35 ± 0.81 | 1.25 ± 3.30 | 0.00 ± 0.00 | NS |
| HbA1C(ng/mL) | 252.25 ± 50.46a | 227.10 ± 41.59a | 275.44 ± 67.73a | 326.76 ± 120.24b | 0.044 |
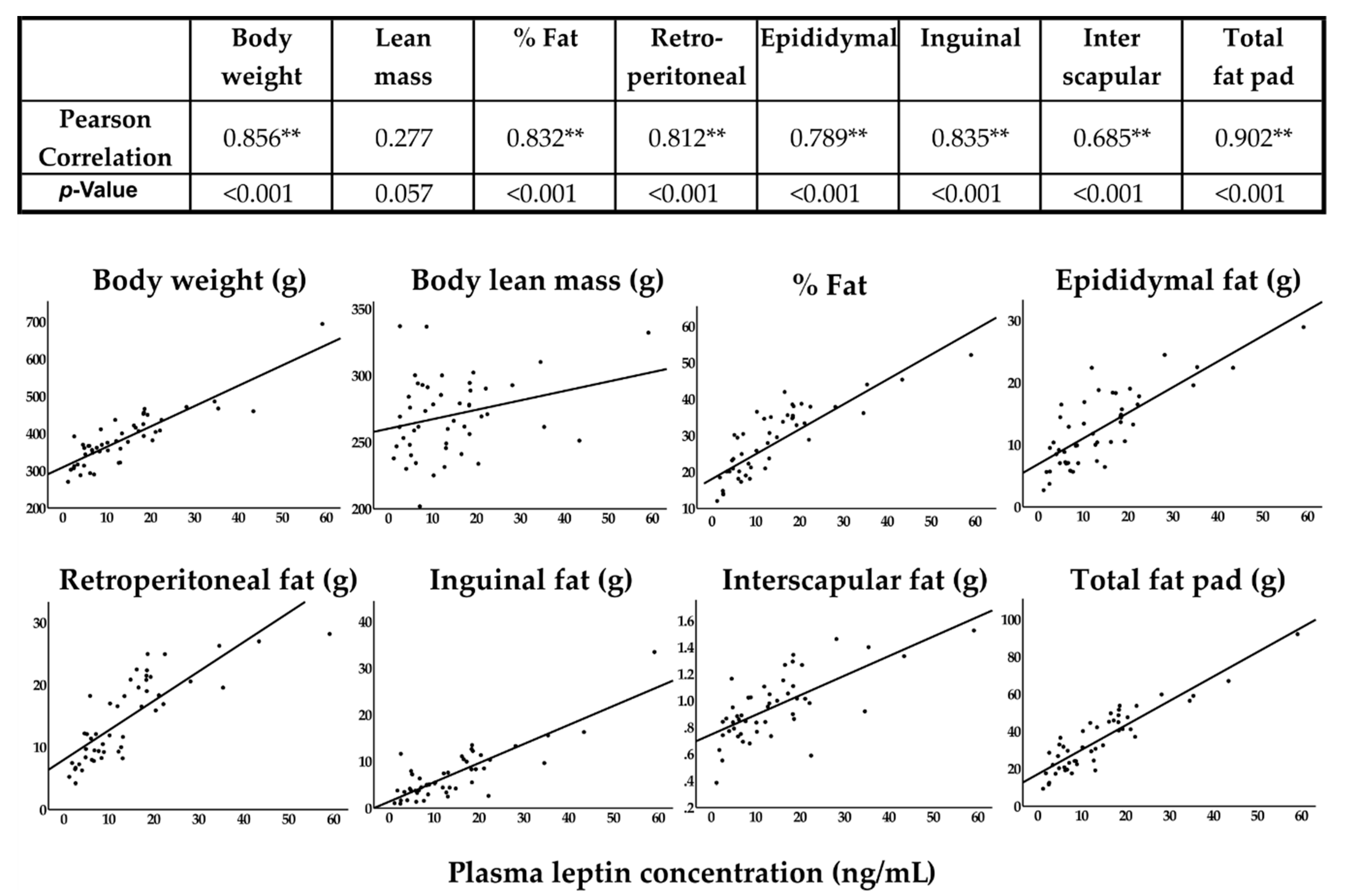
| Leptin (ng/mL) | 7.71 ± 5.62a | 8.47 ± 5.52a | 18.26 ± 10.94b | 21.72 ± 14.59b | 0.002 |
| Adiponectin (µg/mL) | 8.17 ± 1.97 | 7.85 ± 2.94 | 7.92 ± 1.94 | 9.51 ± 0.96 | NS |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siriarchavatana, P.; Kruger, M.C.; Miller, M.R.; Tian, H.S.; Wolber, F.M. The Preventive Effects of Greenshell Mussel (Perna canaliculus) on Early-Stage Metabolic Osteoarthritis in Rats with Diet-Induced Obesity. Nutrients 2019, 11, 1601. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11071601
Siriarchavatana P, Kruger MC, Miller MR, Tian HS, Wolber FM. The Preventive Effects of Greenshell Mussel (Perna canaliculus) on Early-Stage Metabolic Osteoarthritis in Rats with Diet-Induced Obesity. Nutrients. 2019; 11(7):1601. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11071601
Chicago/Turabian StyleSiriarchavatana, Parkpoom, Marlena C. Kruger, Matthew R. Miller, Hong Sabrina Tian, and Frances M. Wolber. 2019. "The Preventive Effects of Greenshell Mussel (Perna canaliculus) on Early-Stage Metabolic Osteoarthritis in Rats with Diet-Induced Obesity" Nutrients 11, no. 7: 1601. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11071601