Bioactive Compounds of Cooked Tomato Sauce Modulate Oxidative Stress and Arachidonic Acid Cascade Induced by Oxidized LDL in Macrophage Cultures
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
2.2. Culture of Human Monocyte/Macrophage
2.3. Isolation of LDL and LDL Oxidation Assay
2.4. Assay for Binding and/or Uptake of oxLDL by Macrophages
2.5. Determination of Reactive Oxygen Species (ROS) and Nitric Oxide (NO) Production
2.6. Measurement of Prostaglandin E2 (PGE2) and Leukotriene B4 (LTB4)
2.7. Statistical Analysis
3. Results
3.1. Sofrito Compounds Modulate LDL Oxidation Induced by Cu2+
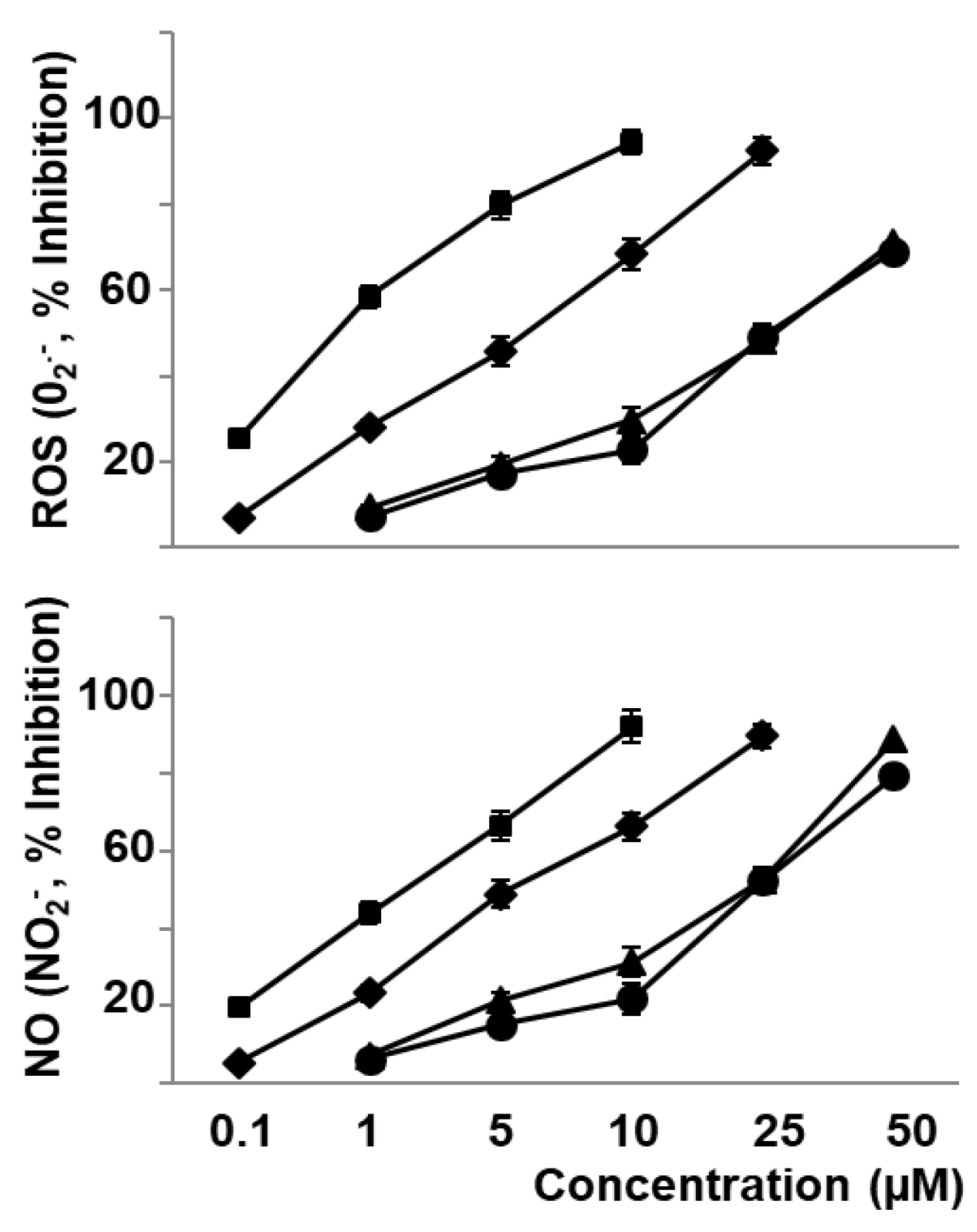
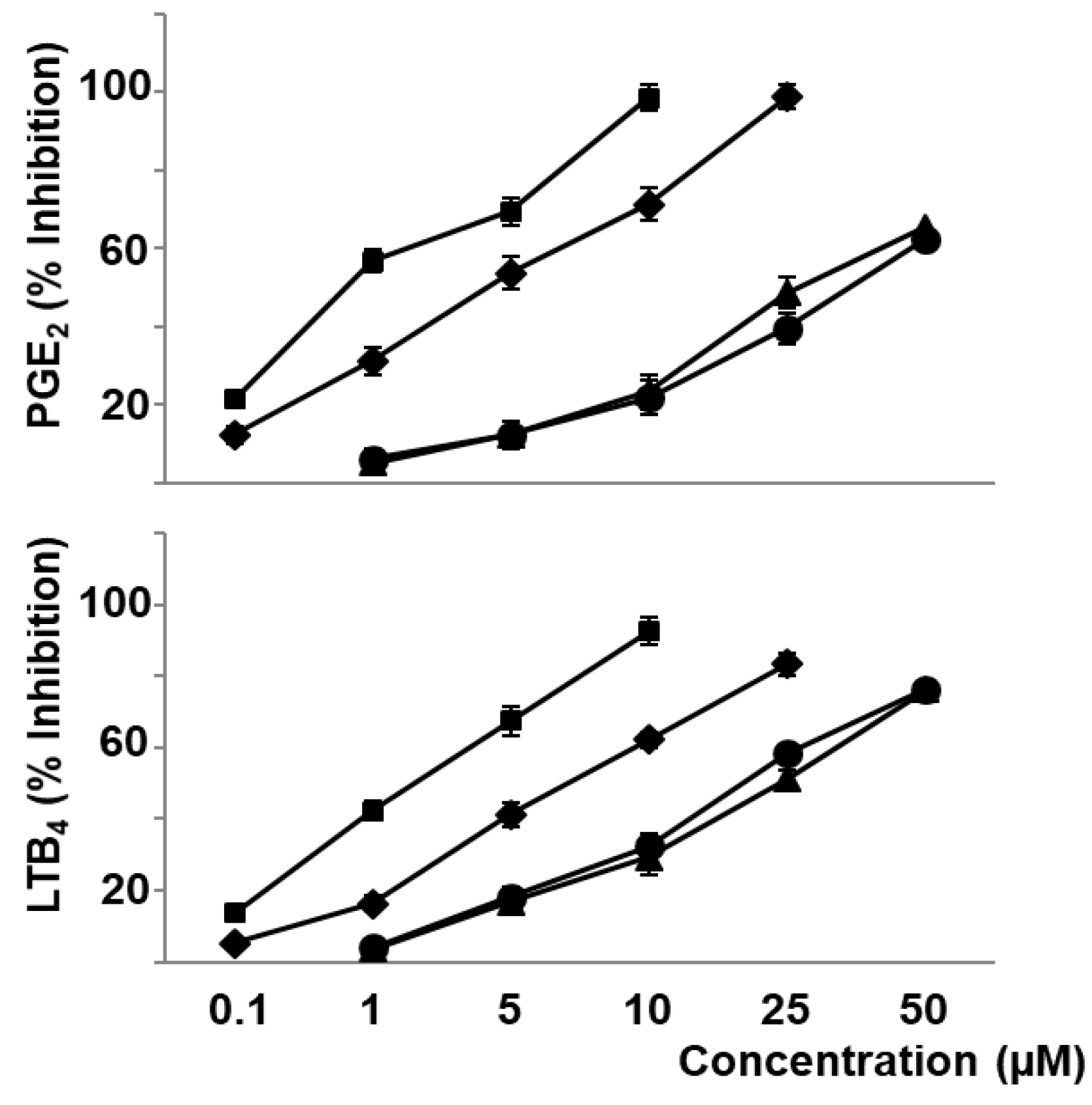
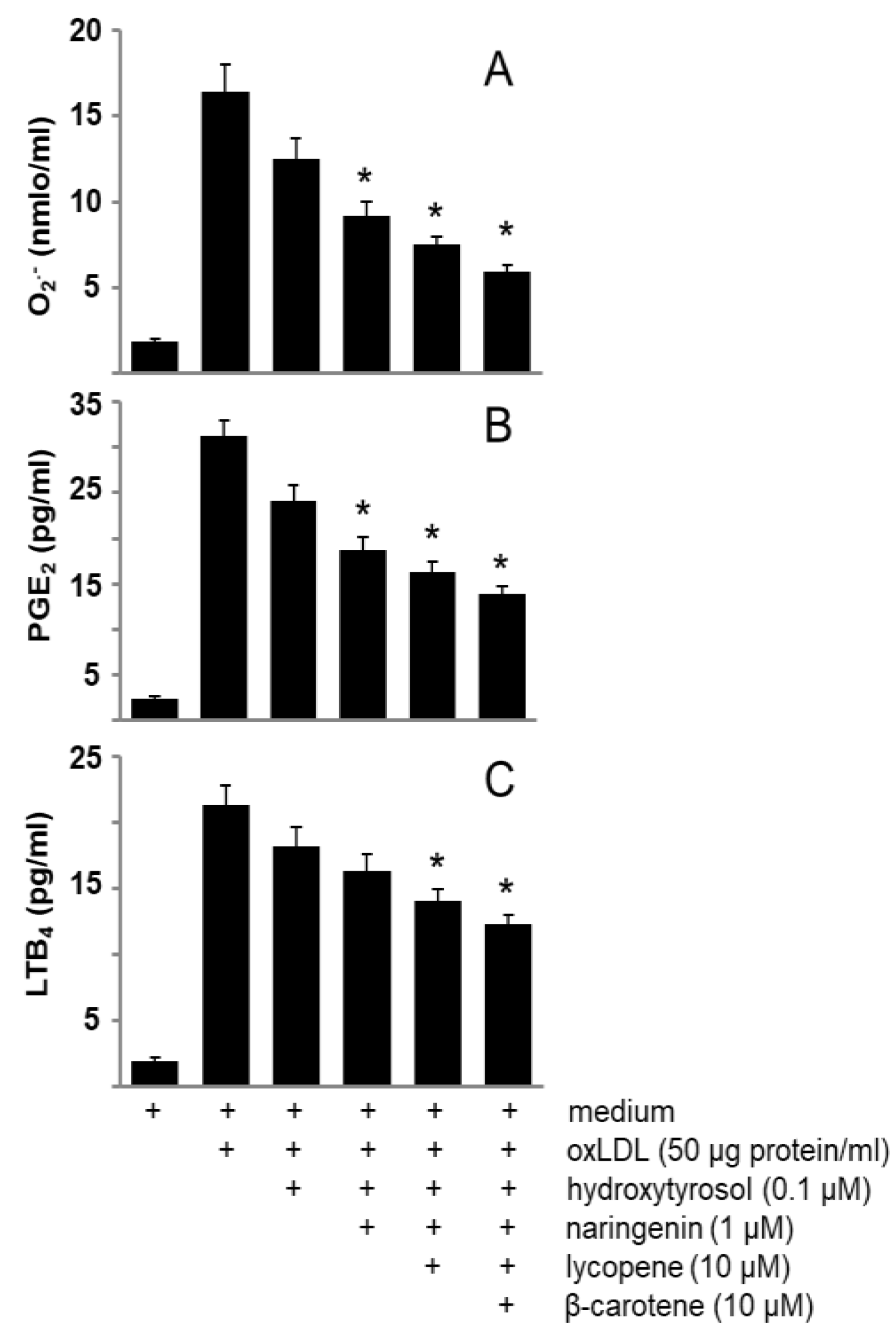
3.2. Phenolic Compounds and Carotenoids of Sofrito Reduce Oxidative Stress and Eicosanoid Synthesis Induced by oxLDL
3.3. Phenolic Compounds and Carotenoids of Sofrito Did Not Affect the Binding and Uptake of oxLDL by Macrophages
3.4. Naringenin 7-O-β-d-glucuronide Has Similar Activity on Oxidative Stress and Eicosanoid Biosynthesis to Naringenin
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- He, F.J.; Nowson, C.A.; Lucas, M.; MacGregor, G.A. Increased consumption of fruit and vegetables is related to a reduced risk of coronary heart disease: Meta-analysis of cohort studies. J. Hum. Hypertens. 2007, 21, 717–728. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, R.; Anad, S.; Ounpuu, S.; Islam, S.; Zhang, X.; Rangarajan, S.; Chifamba, J.; Al-Hinai, A.; Keltai, M. Dietary pattern and the risk of acute myocardial infarction in 52 countries: Results of the interheart study. Circulation 2008, 118, 1929–1937. [Google Scholar] [CrossRef] [PubMed]
- Estruch, R.; Ros, E.; Salas-Salvadó, J.; Covas, M.I.; Corella, D.; Arós, F.; Gómez-Gracia, E.; Ruiz-Gutiérrez, V.; Fiol, M.; Lapetra, J.; et al. Primary prevention of cardiovascular disease with a Mediterranean diet. New Engl. J. Med. 2013, 368, 1279–1290. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.M.; Koutsidis, G.; Lodge, J.K.; Ashor, A.; Siervo, M.; Lara, J. Tomato and lycopene supplementation and cardiovascular risk factors: A systematic review and meta-analysis. Atherosclerosis 2017, 257, 100–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salehi, B.; Sharifi-Rad, R.; Sharopov, F.; Namiesnik, J.; Roointan, A.; Kamle, M.; Kumar, P.; Martins, N.; Darifi-Rad, J. Beneficial effects and potential risks of tomato consumption for human health: An overview. Nutrition 2019, 62, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.M.; Kotsidis, G.; Lodge, J.K.; Ashor, A.W.; Siervo, M.; Lara, J. Lycopene and tomato and risk of cardiovascular diseases: A systematic review and meta-analysis of epidemiological evidences. Crit. Rev. Food Sci. Nutr. 2019, 59, 141–158. [Google Scholar] [CrossRef]
- Vallverdú-Queralt, A.; Rinaldi de Alvarenga, J.F.; Estruch, R.; Lamuela-Raventos, R.M. Bioactive compounds present in the Mediterranean sofrito. Food Chem. 2013, 141, 3365–3372. [Google Scholar]
- Martínez-Huelamo, M.; Tulipani, S.; Estruch, R.; Escribano, E.; Ilan, M.; Corella, D.; Lamuela-Raventos, R.M. The tomato sauce making process affect the bioaccessibility and bioavailability of tomato phenolics: A pharmacokinetic study. Food Chem. 2015, 173, 864–872. [Google Scholar] [CrossRef]
- Yoshimura, M.; Sano, A.; Kamei, J.I.; Obata, A. Identification and quantification of metabolites of orally administered naringenin chalcone in rats. J. Agric. Food Chem. 2009, 57, 6432–6437. [Google Scholar] [CrossRef]
- Mitjavila, M.T.; Moreno, J.J. The effect of polyphenols on oxidative stress and the arachidonic acid cascade. Implications for the prevention/treatment of high prevalence diseases. Biochem. Pharmacol. 2012, 84, 1113–1122. [Google Scholar] [CrossRef]
- Rao, A.V. Lycopene, tomatoes, and the prevention of coronary heart disease. Exp. Biol. Med. 2002, 227, 908–913. [Google Scholar] [CrossRef] [PubMed]
- Visioli, F.; Riso, P.; Grande, S.; Galli, C.; Porrini, M. Protective action of tomato products in in vivo markers of lipid oxidation. Eur. J. Nutr. 2003, 42, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Mulvihill, E.E.; Assini, J.M.; Sutherland, B.G.; DiMattia, A.S.; Khami, M.; Koppes, J.B.; Sawyez, C.G.; Whitman, S.C.; Huff, M.W. Naringenin decreases progression of atherosclerosis by improving dyslipidemia in high-fat-fed low-density lipoprotein receptor-null mice. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 742–748. [Google Scholar] [CrossRef] [PubMed]
- Müller, L.; Caris-Veyrat, C.; Lowe, G.; Böhm, V. Lycopene and its antioxidant role in the prevention of cardiovascular diseases-A critical review. Crit. Rev. Food Sci. Nutr. 2016, 56, 1868–1879. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, D. Low density lipoprotein oxidation and its pathobiological significance. J. Biol. Chem. 1997, 272, 20963–20966. [Google Scholar] [CrossRef] [PubMed]
- Toshima, S.; Haserawa, A.; Kurabayashi, M.; Itabe, H.; Takano, T.; Sugano, J.; Shimamura, K.; Kimura, J.; Michishita, I.; Suzuki, T.; et al. Circulating oxidized low density lipoprotein levels. A biochemical risk marker for coronary heart disease. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 2243–2247. [Google Scholar] [CrossRef]
- Ehara, S.; Vada, M.; Naruko, T. Elevated levels of oxidized low density lipoprotein show a positive relationship with the severity of acute coronary syndromes. Circulation 2001, 103, 1955–1960. [Google Scholar] [CrossRef] [PubMed]
- Henrikse, T.; Mahoney, E.M.; Steinberg, D. Enhanced macrophage degradation of low density lipoprotein previously incubated with cultured endothelial cells: Recognition by receptors for the acetylated low density lipoproteins. Proc. Natl. Acad. Sci. USA 1981, 78, 6499–6503. [Google Scholar] [CrossRef] [PubMed]
- Osberud, B.; Bjorklid, E. Role of monocytes in atherogenesis. Physiol. Rev. 2003, 83, 1069–1112. [Google Scholar] [CrossRef]
- Tsuchiya, S.; Kobayashi, Y.; Goto, Y.; Okumura, H.; Nakae, S.; Konno, T.; Tada, K. Induction of maturation in cultured human monocytic leukemia cells by a phorbol diester. Cancer Res. 1982, 42, 1530–1536. [Google Scholar]
- Chung, B.H.; Segrest, J.P.; Ray, M.J.; Brunzell, J.D.; Hokanson, J.E.; Krauss, R.M.; Beaudrie, K.; Cone, J.T. Single vertical spin density gradient ultracentrifugation. Methods Enzymol. 1986, 128, 181–209. [Google Scholar] [PubMed]
- Peterson, G.L. A simplification of the protein assay method of Lowry et al. which is more generally applicable. Anal. Biochem. 1977, 83, 346–356. [Google Scholar] [CrossRef]
- Esterbauer, H.; Striegl, G.; Puhl, H. Continuous monitoring of in vitro oxidation of human low density lipoprotein. Free Radic. Res. Commun. 1989, 6, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Yagi, K. A simple fluorimetric assay for lipoperoxide in blood sample. Biochem. Med. 1976, 15, 212–216. [Google Scholar] [CrossRef]
- Innerarity, T.L.; Pitas, R.E.; Mahley, R.W. Lipoprotein receptor interactions. Methods Enzymol. 1986, 129, 542–565. [Google Scholar] [PubMed]
- Beppu, M.; Watanabe, M.; Sunohara, M.; Ohishi, K.; Mishima, E.; Kawashi, H.; Fujii, M.; Kikugawa, K. Participation of the arachidonic acid cascade pathway in macrophage binding/uptake of oxidized low density lipoprotein. Biol. Pharm. Bull. 2002, 25, 710–712. [Google Scholar] [CrossRef] [PubMed]
- Moreno, J.J. Effect of olive oil minor components on oxidative stress and arachidonic acid mobilization and metabolism by macrophages RAW 264. Free Radic. Biol. Med. 2003, 35, 1073–1081. [Google Scholar] [CrossRef]
- Krieger, M.; Herz, J. Structures and functions of multiligand lipoprotein receptors: Macrophage scavenger receptor and LDL receptor-related protein (LRP). Annu. Rev. Biochem. 1994, 63, 343–346. [Google Scholar] [CrossRef]
- Ross, J.A.; Kasum, C.M. Dietary flavonoids: Bioavailability, metabolic effects, and safety. Annu. Rev. Nutr. 2002, 22, 19–34. [Google Scholar] [CrossRef]
- Getz, G.S.; Reardon, C.A. Atherogenic lipids and macrophage subsets. Curr. Opin. Lipidol. 2015, 26, 357–361. [Google Scholar] [CrossRef]
- Di Pietro, N.; Formoso, G.; Pandolfi, A. Physiology and pathophysiology of oxLDL uptake by vascular wall cells in atherosclerosis. Vascul. Pharmacol. 2016, 84, 1–7. [Google Scholar] [CrossRef]
- Sugamura, K.; Keaney, J.F., Jr. Reactive oxygen species in cardiovascular disease. Free Radic. Biol. Med. 2011, 51, 978–992. [Google Scholar] [CrossRef] [Green Version]
- Yuhki, K.; Kojima, F.; Kashiwagi, H.; Kawabe, J.; Fujino, T.; Narumiya, S.; Ushikubi, F. Roles of prostanoids in the pathogenesis of cardiovascular diseases: Novel insights from knockout mouse studies. Pharmacol. Ther. 2011, 129, 195–205. [Google Scholar] [CrossRef]
- Sies, H.; Stahl, W.; Sundquist, A.R. Antioxidant functions of vitamins. Vitamins E and C, beta-carotene, and other carotenoids. Ann. N. Y. Acad. Sci. 1992, 669, 7–20. [Google Scholar] [CrossRef]
- Turk, K.L.; Hayball, R.J. Major phenolic compounds in olive oil: Metabolism and health effects. J. Nutr. Biochem. 2002, 13, 636–644. [Google Scholar]
- Cavia-Saiz, M.; Busto, M.D.; Pilar-Izquierdo, M.C.; Ortega, N.; Perez-Mateos, M.; Muñiz, P. Antioxidant properties, radical scavenging activity and biomolecule protection capacity of flavonoid naringenin and its glycoside naringin: A comparative study. J. Sci. Food Agric. 2010, 90, 1238–1244. [Google Scholar] [CrossRef]
- Martínez-Huelamo, M.; Vallverdú-Queralt, A.; Di Lecce, G.; Valderas-Martínez, P.; Tulipani, S.; Jáuregui, O.; Escribano-Ferrer, E.; Estruch, R.; Ilán, M.; Lamuela-Raventos, R.M. Bioavailability of tomato polyphenols is enhanced by processing and fat addition: Evidence from a randomized feeding trial. Mol. Nutr. Food Res. 2016, 60, 1578–1589. [Google Scholar] [CrossRef]
- Fuhrman, B.; Volkova, N.; Rosenblat, M.; Aviram, M. Lycopene synergistically inhibits LDL oxidation in combination with vitamin E, glabridin, rosmarinic acid, carnosic acid, or garlic. Antioxid. Redox Signal. 2000, 2, 491–506. [Google Scholar] [CrossRef]
- Vivancos, M.; Moreno, J.J. Effect of resveratrol, tyrosol and β-sitosterol on oxidised low-density lipoprotein-stimulated oxidative stress, arachidonic acid release and prostaglandin E2 synthesis by RAW 264.7 macrophages. Br. J. Nutr. 2008, 99, 1199–1207. [Google Scholar] [CrossRef]
- Rahaman, S.O.; Lennon, D.J.; Febbraio, M.; Rodrez, E.A.; Hazen, S.L.; Silverstein, R.L. A CD36-dependent signalling cascade is necessary for macrophage foam cell formation. Cell Metab. 2006, 4, 211–221. [Google Scholar] [CrossRef]
- Martinez, J.; Sanchez, T.; Moreno, J.J. Regulation of prostaglandin E2 production by the superoxide radical and nitric oxide in mouse peritoneal macrophages. Free Radic. Res. 2000, 32, 303–311. [Google Scholar] [CrossRef]
- Martinez, J.; Moreno, J.J. Role of Ca2+-independent phospholipase A2 on arachidonic acid release induced by reactive oxygen species. Arch. Biochem. Biophys. 2001, 392, 257–262. [Google Scholar] [CrossRef]
- Vivancos, M.; Moreno, J.J. Role of Ca2+-independent phospholipase A2 and cyclooxygenase/Lipoxygenase pathways in the nitric oxide production by murine macrophages stimulated by lipopolysaccharides. Nitric Oxide 2002, 6, 255–262. [Google Scholar] [CrossRef]
- Lupo, G.; Nicotra, A.; Giurdanella, G.; Anfuso, C.D.; Romeo, L.; Biondi, G.; Tirolo, C.; Marchetti, B.; Ragusa, N.; Alberghina, M. Activation of phospholipase A2 and MAP kinases by oxidized low-density lipoproteins in immortalized GP8.39 endothelial cells. Biochim. Biophys. Acta 2005, 1735, 135–150. [Google Scholar] [CrossRef]
- Muroya, T.; Ihara, Y.; Ikeda, S.; Yasuoka, C.; Miyahara, Y.; Urata, Y.; Kondo, T.; Kohno, S. Oxidative modulation of NFkB signaling by oxidized low-density lipoproteins. Biochem. Biophys. Res. Commun. 2003, 309, 900–905. [Google Scholar] [CrossRef]
- Kamiloglu, S.; Demirci, M.; Selen, S.; Toydemir, G.; Boyacioglu, D.; Capanoglu, E. Home processing of tomatoes (solanum lycopersicum): Effects on in vitro bioaccessibility of total lycopene, phenolics, flavonoids, and antioxidant capacity. J. Sci. Food Agric. 2014, 94, 2225–2233. [Google Scholar] [CrossRef]
- Valderas-Martínez, P.; Chica-Blanch, G.; Casa, R.; Arranz, S.; Martínez-Huelamo, M.; Urpi-Sarda, M.; Torrado, X.; Corella, D.; Lamuela-Raventos, R.M.; Estruch, R. Tomato sauce enriched with olive oil exerts greater effects on cardiovascular disease risk factors than raw tomato and tomato sauce: A randomized trial. Nutrients 2016, 8, 170. [Google Scholar] [CrossRef]
- Hiltunen, T.P.; Yla-Herttuala, S. Expression of lipoprotein receptors in atherosclerotic lesions. Atherosclerosis 1998, 137, S81–S88. [Google Scholar] [CrossRef]
- Miles, E.A.; Wallace, F.A.; Calder, P.C. An olive oil-rich diet reduces scavenger receptor mRNA in murine macrophages. Br. J. Nutr. 2001, 85, 185–191. [Google Scholar] [CrossRef] [Green Version]
- Granados-Principal, S.; Quiles, J.L.; Ramírez-Tortosa, C.L.; Ochoa-Herrera, J.; Perez-Lopez, P.; Pulido-Moran, M.; Ramirez-Tortosa, M.C. Squalene ameliorates atherosclerotic lesions through the reduction of CD36 scavenger receptor expression in macrophages. Mol. Nutr. Food Res. 2012, 56, 733–740. [Google Scholar] [CrossRef]
- Storniolo, C.E.; Moreno, J.J. Resveratrol metabolites have an antiproliferative effect on intestinal epithelial cancer cells. Food Chem. 2012, 134, 1385–1391. [Google Scholar] [CrossRef]
- Tresserra-Rimbau, A.; Lamuela-Raventos, R.M.; Moreno, J.J. Polyphenols, food and pharma. Current knowledge and directions for future research. Biochem. Pharmacol. 2018, 156, 186–195. [Google Scholar] [CrossRef]
- Vilahur, G.; Cubedo, J.; Padró, T.; Casaní, L.; Mendieta, G.; González, A.; Badimon, L. Intake of cooked tomato sauce preserves coronary endothelial function and improves apolipoprotein A-I and apolipoprotein J protein profile in high-density lipoproteins. Transl. Res. 2015, 166, 44–56. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| CD | TBARs | ||
|---|---|---|---|
| Tlag (min) | ΔTlag (%) | MDA | |
| LDL | 3.2 ± 0.2 | ||
| LDL + Cu2+ | 58 ± 6 | 27.3 ± 1.3 | |
| LDL + Cu2+ + Naringenin (1 µM) | 97 ± 7 * | 67.2 | 19.5 ± 0.7 * |
| LDL + Cu2+ + Naringenin (10 µM) | 198 ± 13 * | 241.4 | 12.1 ± 0.4 * |
| LDL + Cu2+ + Naringenin Gluc. (10 μM) | 204 ± 12 * | 251.7 | 13.6 ± 0.3 * |
| LDL + Cu2+ + Hydroxytyrosol (1 μM) | 225 ± 11 * | 287.9 | 14.8 ± 0.3 * |
| LDL + Cu2+ + Hydroxytyrosol (10 μM) | >500 * | 8.3 ± 0.2 * | |
| LDL + Cu2+ + Lycopene (10 μM) | 73 ± 6 | 25.8 | 22.5 ± 0.4 |
| LDL + Cu2+ + Lycopene (50 μM) | 121 ± 8 * | 108.6 | 15.7 ± 0.3 * |
| LDL + Cu2+ + β Carotene (10 μM) | 72 ± 5 | 24.1 | 23.8 ± 0.4 |
| LDL + Cu2+ + β Carotene (50 μM) | 119 ± 7 * | 105.2 | 17.9 ± 0.3 * |
| LDL + Cu2+ + all compounds (1 μM) | 386 ± 8 * | 565.5 | 10.6 ± 0.2 * |
| LDL + Cu2+ + SOD + Catalase | >500 * | 5.7 ± 0.2 * | |
| ROS | NO | PGE2 | LTB4 | |
|---|---|---|---|---|
| Control | 2.0 ± 0.1 | 1.3 ± 0.1 | 2.5 ± 0.5 | 1.8 ± 0.4 |
| oxLDL | 15.4 ± 1.6 * | 32.1 ± 2.7 * | 26.8 ± 2.6 * | 21.2 ± 1.8 * |
| oxLDL + Naringenin | 6.3 ± 0.8 ≠ | 12.4 ± 1.3 ≠ | 11.6 ± 2.1 ≠ | 13.5 ± 1.4 ≠ |
| oxLDL + Naringenin 7-O-β-gluc. | 7.6 ± 1.2 ≠ | 13.6 ± 1.5 ≠ | 13.2 ± 2.5 ≠ | 15.1 ± 1.3 ≠ |
| Fluorescence Intensity (Arbitrary Units) | |
|---|---|
| oxLDL | 21.2 ± 1.5 |
| Dil-oxLDL | 532.2 ± 21.3 |
| Dil-oxLDL + Naringenin (10 µM) | 476.7 ± 16.1 |
| Dil-oxLDL + Hydroxytyrosol (10 µM) | 453.4 ± 17.3 |
| Dil-oxLDL + Lycopene (50 µM) | 503.5 ± 13.8 |
| Dil-oxLDL + β-Carotene (50 µM) | 517.1 ± 12.8 |
| Dil-oxLDL + Fucoidan (10 µg/mL) | 102.6 ± 3.7 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Storniolo, C.E.; Sacanella, I.; Mitjavila, M.T.; Lamuela-Raventos, R.M.; Moreno, J.J. Bioactive Compounds of Cooked Tomato Sauce Modulate Oxidative Stress and Arachidonic Acid Cascade Induced by Oxidized LDL in Macrophage Cultures. Nutrients 2019, 11, 1880. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11081880
Storniolo CE, Sacanella I, Mitjavila MT, Lamuela-Raventos RM, Moreno JJ. Bioactive Compounds of Cooked Tomato Sauce Modulate Oxidative Stress and Arachidonic Acid Cascade Induced by Oxidized LDL in Macrophage Cultures. Nutrients. 2019; 11(8):1880. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11081880
Chicago/Turabian StyleStorniolo, Carolina E., Ignasi Sacanella, María T. Mitjavila, Rosa M. Lamuela-Raventos, and Juan J. Moreno. 2019. "Bioactive Compounds of Cooked Tomato Sauce Modulate Oxidative Stress and Arachidonic Acid Cascade Induced by Oxidized LDL in Macrophage Cultures" Nutrients 11, no. 8: 1880. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11081880