Dietary Micronutrient Supplementation for 12 Days in Obese Male Mice Restores Sperm Oxidative Stress

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Diet, and Micronutrient Supplementation
2.2. Glucose and Insulin Tolerance Testing
2.3. Body Composition and Serum Metabolites and Hormones
2.4. Collection of Mouse Sperm
2.5. Sperm Motility and Morphology
2.6. Sperm Capacitation
2.7. Sperm Binding to the Zona Pellucida of MII Oocyte and Fertilisation Rates
2.8. Sperm Intracellular ROS Concentrations (DCFDA)
2.9. Sperm Mitochondrial ROS Concentrations (Superoxide-MitoSOX Red)
2.10. Sperm Oxidative DNA Damage (8OHdG)
2.11. Oxidative DNA Damage (8OhdG) in the Pronuclear Embryo
2.12. Pregnancy
2.13. Statistical Analysis
3. Results
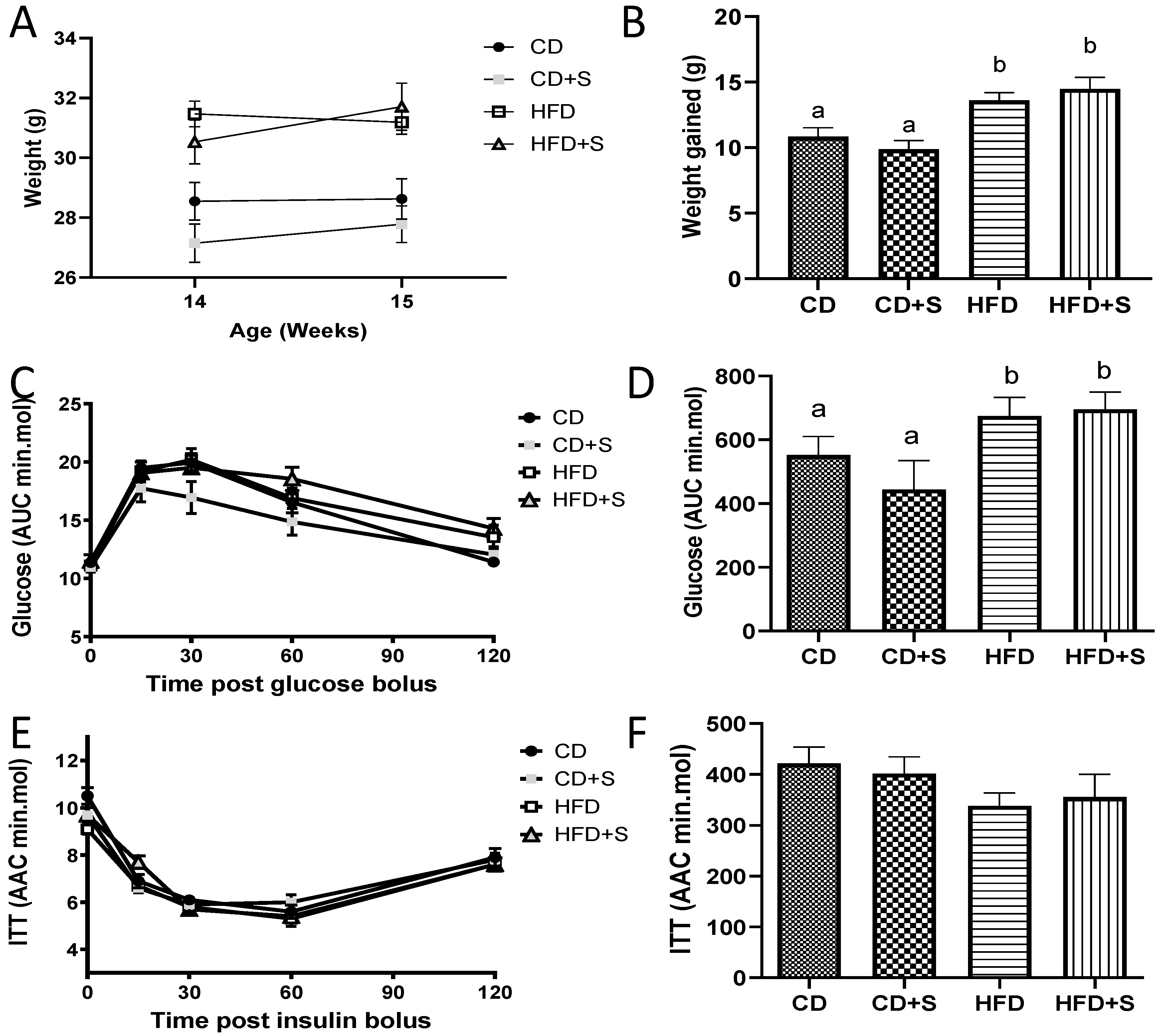
3.1. Short-Term Micronutrient Supplementation Has No Effect on Body Composition or Serum Metabolites Outside of the Original Diets
3.1.1. Pre-Intervention
3.1.2. Post-Intervention
3.2. Short-Term Micronutrient Supplementation Improves Aspects of Sperm Function in CD-Fed Mice
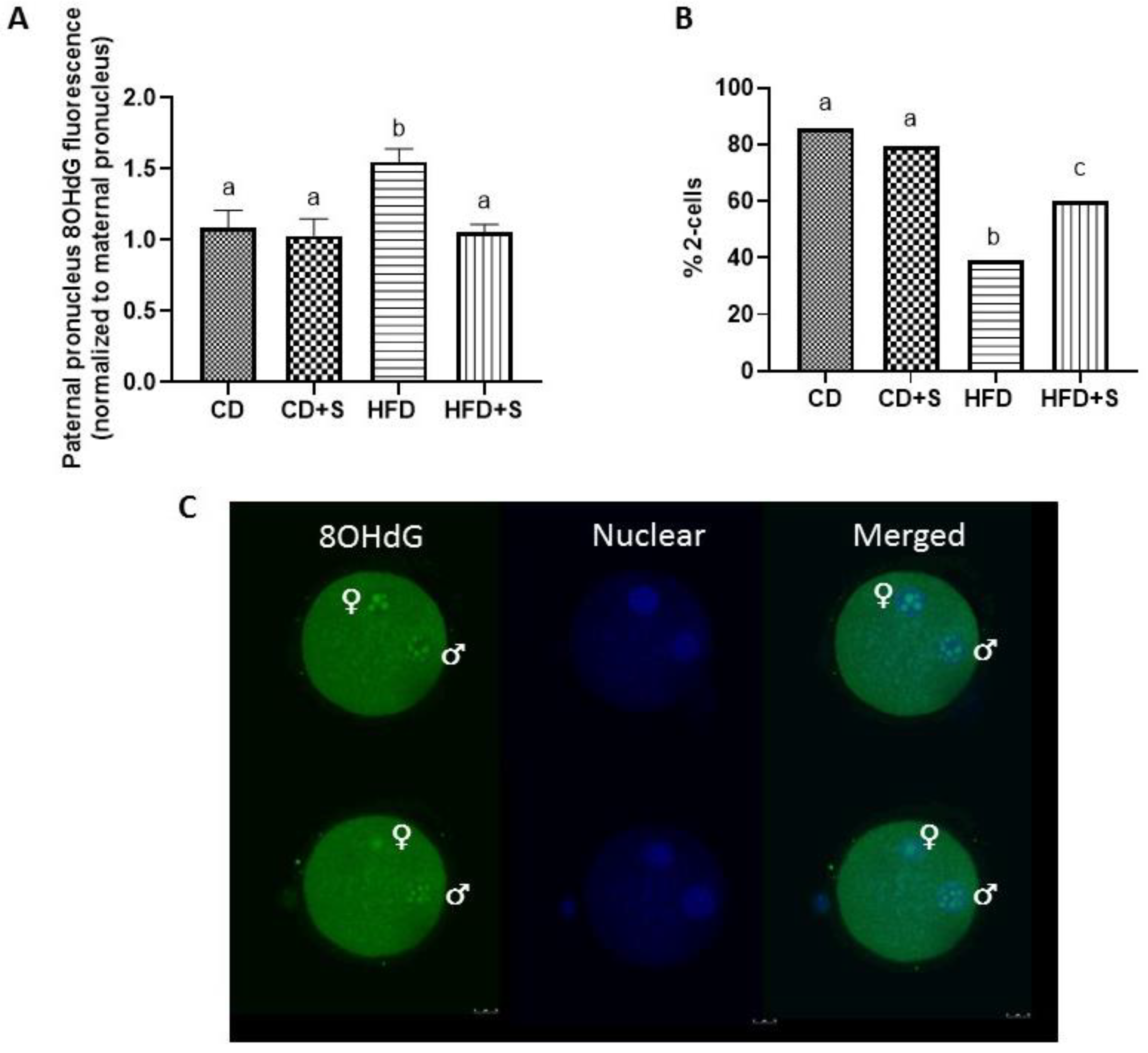
3.3. Short-Term Micronutrient Supplementation Reduces Intracellular Sperm ROS Concentrations and Oxidative DNA Damage in Males Fed a HFD
3.4. Short-Term Micronutrient Supplementation Reduces Oxidative DNA Damage in the Early Pronuclear Embryo and Increases 2-Cell Cleavage Rates from Males Fed a HFD
3.5. Short-Term Micronutrient Supplementation in Fathers Fed a HFD Partially Restored Fetal Weights Similar to Fathers Fed a CD
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Obesity and Overweight; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- Australian Institute of Health and Welfare. A Picture of Overwight and Obesity in Australia 2017; Australian Institute of Health and Welfare: Canberra, ACT, Australia, 2017. [Google Scholar]
- Ng, M.; Fleming, T.; Robinson, M.; Thomson, B.; Graetz, N.; Margono, C.; Mullany, E.C.; Biryukov, S.; Abbafati, C.; Abera, S.F.; et al. Global, regional, and national prevalence of overweight and obesity in children and adults during 1980–2013: A systematic analysis for the global burden of disease study 2013. Lancet 2014, 384, 766–781. [Google Scholar] [CrossRef]
- Egger, G.; Dixon, J. Beyond obesity and lifestyle: A review of 21st century chronic disease determinants. Biomed. Res. Int. 2014, 2014, 731685. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.M.; Lane, M.; Owens, J.A.; Bakos, H.W. Paternal obesity negatively affects male fertility and assisted reproduction outcomes: A systematic review and meta-analysis. Reprod. Biomed. Online 2015, 31, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.P.; Xiao, X.M.; Li, J.; Reichetzeder, C.; Wang, Z.N.; Hocher, B. Paternal body mass index (bmi) is associated with offspring intrauterine growth in a gender dependent manner. PLoS ONE 2012, 7, e36329. [Google Scholar] [CrossRef] [PubMed]
- McCowan, L.M.E.; North, R.A.; Kho, E.M.; Black, M.A.; Chan, E.H.; Dekker, G.A.; Poston, L.; Taylor, R.S.; Roberts, C.T. Paternal contribution to small for gestational age babies: A multicenter prospective study. Obesity 2011, 19, 1035–1039. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.F.; Lin, R.C.; Laybutt, D.R.; Barres, R.; Owens, J.A.; Morris, M.J. Chronic high-fat diet in fathers programs beta-cell dysfunction in female rat offspring. Nature 2010, 467, 963–966. [Google Scholar] [CrossRef] [PubMed]
- Fullston, T.; Teague, E.M.; Palmer, N.O.; de Blasio, M.J.; Mitchell, M.; Print, C.G.; Owens, J.A.; Lane, M. Paternal obesity initiates metabolic disturbances in two generations of mice and alters the transcription profile of tesis and sperm microrna content. FASEB J. 2013, 27, 4226–4243. [Google Scholar] [CrossRef] [PubMed]
- McPherson, N.O.; Fullston, T.; Owens, J.A.; Lane, M. Paternal lifestyle interventions in obese males restores early embryo development and fetal weights, improving the metabolic health and adiposity status in subsequent female offspring. PLoS ONE 2013, 8, e71459. [Google Scholar]
- McPherson, N.O.; Lane, M.; Sandeman, L.; Owens, J.A.; Fullston, T. An exercise-only intervention in obese fathers restores glucose and insulin regulation in conjunction with the rescue of pancreatic islet cell morphology and microrna expression in male offspring. Nutrients 2017, 9, 122. [Google Scholar] [CrossRef]
- McPherson, N.O.; Owens, J.A.; Fullston, T.; Lane, M. Preconception diet or exercise interventions in obese fathers normalizes sperm microrna profile and metabolic syndrome in female offspring. Am. J. Physiol. Endocrinol. Metab. 2015, 308, 805–821. [Google Scholar] [CrossRef]
- Binder, N.K.; Hannan, N.J.; Gardner, D.K. Paternal diet-induced obesity retards early mouse embryo development, mitochondrial activity and pregnancy health. PLoS ONE 2012, 7, e52304. [Google Scholar] [CrossRef] [PubMed]
- Rath, S.R.; Marsh, J.A.; Newnham, J.P.; Zhu, K.; Atkinson, H.C.; Mountain, J.; Oddy, W.H.; Hughes, I.P.; Harris, M.; Leong, G.M.; et al. Parental pre-pregnancy bmi is a dominant early-life risk factor influencing bmi of offspring in adulthood. Obes. Sci. Pract. 2016, 2, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Fullston, T.; Palmer, N.O.; Owens, J.A.; Mitchell, M.; Bakos, H.W.; Lane, M. Diet-induced paternal obesity in the absence of diabetes diminishes the reproductive health of two subsequent generations of mice. Hum. Reprod. 2012, 27, 1391–1400. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Li, H.G.; Fu, L.; Di, F.S. Influence of high-fat diet in paternal c57bl/6 mice on liver fat deposition in offspring. Chin. J. Hepatol. 2017, 25, 139–144. [Google Scholar]
- Chowdhury, S.S.; Lecomte, V.; Erlich, J.H.; Maloney, C.A.; Morris, M.J. Paternal high fat diet in rats leads to renal accumulation of lipid and tubular changes in adult offspring. Nutrients 2016, 8, 521. [Google Scholar] [CrossRef]
- Pataia, V.; Papacleovoulou, G.; Nikolova, V.; Samuelsson, A.M.; Chambers, S.; Jansen, E.; Taylor, P.D.; Poston, L.; Williamson, C. Paternal cholestasis exacerbates obesity-associated hypertension in male offspring but is prevented by paternal ursodeoxycholic acid treatment. Int. J. Obes. 2019, 43, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Lane, M.; McPherson, N.O.; Fullston, T.; Spillane, M.; Sandeman, L.; Xian, W.; Zander, D.L. Oxidative stress in mouse sperm impairs embryo development, fetal growth and alters adiposity and glucose regulation in female offspring. PLoS ONE 2014, 9, e100832. [Google Scholar] [CrossRef] [PubMed]
- Aitken, J.; Fisher, H. Reactive oxygen species generation and human spermatozoa: The balance of benefit and risk. Bioessays 1994, 16, 259–267. [Google Scholar] [CrossRef]
- Aitken, R.J.; Baker, M.A. Reactive oxygen species generation by human spermatozoa: A continuing enigma. Int. J. Androl. 2002, 25, 191–194. [Google Scholar] [CrossRef]
- Oakberg, E.F. Duration of spermatogenesis in the mouse. Nature 1957, 180, 1137–1138. [Google Scholar] [CrossRef]
- Bedford, J.M. The status and the state of the human epididymis. Hum. Reprod. 1994, 9, 2187–2199. [Google Scholar] [CrossRef]
- Aitken, R.J.; De Iuliis, G.N. On the possible origins of DNA damage in human spermatozoa. Mol. Hum. Reprod. 2010, 16, 3–13. [Google Scholar] [CrossRef]
- Hedger, M.P. Immunophysiology and pathology of inflammation in the testis and epididymis. J. Androl. 2011, 32, 625–640. [Google Scholar] [CrossRef]
- Palmer, N.O.; Bakos, H.W.; Owens, J.A.; Setchell, B.P.; Lane, M. Diet and exercise in an obese mouse fed a high-fat diet improve metabolic health and reverse perturbed sperm function. Am. J. Physiol. Endocrinol. Metab. 2012, 302, 768–780. [Google Scholar] [CrossRef]
- McPherson, N.O.; Fullston, T.; Bakos, H.W.; Setchell, B.P.; Lane, M. Obese father’s metabolic state, adiposity, and reproductive capacity indicate son’s reproductive health. Fertil. Steril. 2014, 101, 865–873. [Google Scholar] [CrossRef]
- McPherson, N.O.; Bakos, H.W.; Setchell, B.P.; Owens, J.A.; Lane, M. Improving metabolic health in obese male mice via diet and exercise restores embryo development and fetal growth. PLoS ONE 2013, 8, e71459. [Google Scholar] [CrossRef]
- Wu, T.; Gao, X.; Chen, M.; van Dam, R.M. Long-term effectiveness of diet-plus-exercise interventions vs. Diet-only interventions for weight loss: A meta-analysis. Obes. Rev. 2009, 10, 313–323. [Google Scholar] [CrossRef]
- Sies, H.; Stahl, W.; Sevanian, A. Nutritional, dietary and postprandial oxidative stress. J. Nutr. 2005, 135, 969–972. [Google Scholar] [CrossRef]
- Salas-Huetos, A.; Bullo, M.; Salas-Salvado, J. Dietary patterns, foods and nutrients in male fertility parameters and fecundability: A systematic review of observational studies. Hum. Reprod. Update 2017, 23, 371–389. [Google Scholar] [CrossRef]
- Salas-Huetos, A.; Moraleda, R.; Giardina, S.; Anton, E.; Blanco, J.; Salas-Salvado, J.; Bullo, M. Effect of nut consumption on semen quality and functionality in healthy men consuming a western-style diet: A randomized controlled trial. Am. J. Clin. Nutr. 2018, 108, 953–962. [Google Scholar] [CrossRef]
- Mir, J.; Franken, D.; Andrabi, S.W.; Ashraf, M.; Rao, K. Impact of weight loss on sperm DNA integrity in obese men. Andrologia 2018, 50, e12957. [Google Scholar] [CrossRef]
- Garcia, O.P.; Long, K.Z.; Rosado, J.L. Impact of micronutrient deficiencies on obesity. Nutr. Rev. 2009, 67, 559–572. [Google Scholar] [CrossRef]
- Kaidar-Person, O.; Rosenthal, R.J. Malnutrition in morbidly obese patients: Fact or fiction? Minerva Chir. 2009, 64, 297–302. [Google Scholar]
- Martin-Hidalgo, D.; Bragado, M.J.; Batista, A.R.; Oliveira, P.F.; Alves, M.G. Antioxidants and male fertility: From molecular studies to clinical evidence. Antioxidants 2019, 8, 89. [Google Scholar] [CrossRef]
- Majzoub, A.; Agarwal, A. Systematic review of antioxidant types and doses in male infertility: Benefits on semen parameters, advanced sperm function, assisted reproduction and live-birth rate. Arab. J. Urol. 2018, 16, 113–124. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi Roushandeh, A.; Salehi, I.; Mortazavi, M. Protective effects of restricted diet and antioxidants on testis tissue in rats fed with high-fat diet. Iran. Biomed. J. 2015, 19, 96–101. [Google Scholar]
- Ibrahim, H.A.; Zhu, Y.; Wu, C.; Lu, C.; Ezekwe, M.O.; Liao, S.F.; Haung, K. Selenium-enriched probiotics improves murine male fertility compromised by high fat diet. Biol. Trace Elem. Res. 2011, 147, 251–260. [Google Scholar] [CrossRef]
- Sohrabi, M.; Hosseini, M.; Inan, S.; Alizadeh, Z.; Vahabian, M.; Vahidinia, A.A.; Lahoutian, H. Effect of antioxidants on testicular inos and enos after high-fat diet in rat. Pak. J. Biol. Sci 2017, 20, 289–297. [Google Scholar] [CrossRef]
- Bakos, H.W.; Mitchell, M.; Setchell, B.P.; Lane, M. The effect of paternal diet-induced obesity on sperm function and fertilization in a mouse model. Int. J. Androl. 2011, 34, 402–410. [Google Scholar] [CrossRef]
- Kerns, K.; Zigo, M.; Sutovsky, P. Zinc: A necessary ion for mammalian sperm fertilization competency. Int. J. Mol. Sci. 2018, 19, 4097. [Google Scholar] [CrossRef]
- Jueraitetibaike, K.; Ding, Z.; Wang, D.D.; Peng, L.P.; Jing, J.; Chen, L.; Ge, X.; Qiu, X.H.; Yao, B. The effect of vitamin d on sperm motility and the underlying mechanism. Asian J. Androl. 2019, 21, 400. [Google Scholar] [PubMed]
- Perrotta, I.; Perri, M.; Santoro, M.; Panza, S.; Caroleo, M.C.; Guido, C.; Mete, A.; Cione, E.; Aquila, S. Expression and subcellular localization of retinoic acid receptor-alpha (raralpha) in healthy and varicocele human spermatozoa: Its possible regulatory role in capacitation and survival. Appl. Immunohistochem. Mol. Morphol. 2015, 23, 374–381. [Google Scholar] [CrossRef] [PubMed]
- De Amicis, F.; Santoro, M.; Guido, C.; Russo, A.; Aquila, S. Epigallocatechin gallate affects survival and metabolism of human sperm. Mol. Nutr. Food Res. 2012, 56, 1655–1664. [Google Scholar] [CrossRef] [PubMed]
- WHO. Laboratory manual of the who for the examination of human semen and sperm-cervical mucus interaction. Ann. dell’Istituto Super. di Sanita 2010, 37, 1–123. [Google Scholar]
- Baker, S.S.; Thomas, M.; Thaler, C.D. Sperm membrane dynamics assessed by changes in lectin fluorescence before and after capacitation. J. Androl. 2004, 25, 744–751. [Google Scholar] [CrossRef] [PubMed]
- Lane, M.; Gardner, D.K. Ammonium induces aberrant blastocyst differentiation, metabolism, pH regulation, gene expression and subsequently alters fetal development in the mouse. Biol. Reprod. 2003, 69, 1109–1117. [Google Scholar] [CrossRef]
- Koppers, A.J.; Garg, M.L.; Aitken, R.J. Stimulation of mitochondrial reactive oxygen species production by unesterified, unsaturated fatty acids in defective human spermatozoa. Free Radic. Biol. Med. 2010, 48, 112–119. [Google Scholar] [CrossRef] [PubMed]
- McPherson, N.O.; Fullston, T.; Kang, W.X.; Sandeman, L.Y.; Corbett, M.A.; Owens, J.A.; Lane, M. Paternal under-nutrition programs metabolic syndrome in offspring which can be reversed by antioxidant/vitamin food fortification in fathers. Sci. Rep. 2016, 6, 27010. [Google Scholar] [CrossRef] [Green Version]
- Lord, T.; Aitken, R.J. Fertilization stimulates 8-hydroxy-2′-deoxyguanosine repair and antioxidant activity to prevent mutagenesis in the embryo. Dev. Biol. 2015, 406, 1–13. [Google Scholar] [CrossRef]
- Edwards, R.G.; Sirlin, J.L. Labelled pronuclei in mouse eggs fertilized by labelled sperm. Nature 1956, 177, 429. [Google Scholar] [CrossRef]
- Sirlin, J.L.; Edwards, R.G. Timing of DNA synthesis in ovarian oocyte nuclei and pronuclei of the mouse. Exp. Cell Res. 1959, 18, 190–194. [Google Scholar] [CrossRef]
- Levine, H.; Jorgensen, N.; Martino-Andrade, A.; Mendiola, J.; Weksler-Derri, D.; Mindlis, I.; Pinotti, R.; Swan, S.H. Temporal trends in sperm count: A systematic review and meta-regression analysis. Hum. Reprod. Update 2017, 23, 646–659. [Google Scholar] [CrossRef]
- Hakonsen, L.B.; Thulstrup, A.M.; Aggerholm, A.S.; Olsen, J.; Bonde, J.P.; Andersen, C.Y.; Bungum, M.; Ernst, E.H.; Hansen, M.L.; Ramlau-Hansen, C.H. Does weight loss improve semen quality and reproductive hormones? Results from a cohort of severely obese men. Reprod. Health 2011, 8, 24. [Google Scholar] [CrossRef]
- Crean, A.J.; Senior, A.M. High-fat diets reduce male reproductive success in animal models: A systematic review and meta-analysis. Obes. Rev. 2019, 20, 921–933. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Xu, Q.Y.; Jia, C.; Peng, Y.F. Metformin improves epididymal sperm quality and antioxidant function of the testis in diet-induced obesity rats. Natl. J. Androl. 2012, 18, 146–149. [Google Scholar]
- Tunc, O.; Thompson, J.; Tremellen, K. Development of the nbt assay as a marker of sperm oxidative stress. Int. J. Androl. 2010, 33, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Kim, D.Y.; Choue, R.; Lim, H. Effects of vitamin c supplementation on plasma and urinary vitamin c concentration in korean women. Clin. Nutr. Res. 2017, 6, 198–205. [Google Scholar] [CrossRef]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef]
- Fraga, C.G.; Motchnik, P.A.; Shigenaga, M.K.; Helbock, H.J.; Jacob, R.A.; Ames, B.N. Ascorbic acid protects against endogenous oxidative DNA damage in human sperm. Proc. Natl. Acad. Sci. USA 1991, 88, 11003–11006. [Google Scholar] [CrossRef] [PubMed]
- Raad, G.; Mansour, J.; Ibrahim, R.; Azoury, J.; Azoury, J.; Mourad, Y.; Fakih, C.; Azoury, J. What are the effects of vitamin C on sperm functional properties during direct swim-up procedure? Zygote 2019, 27, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Adami, L.N.G.; Belardin, L.B.; Lima, B.T.; Jeremias, J.T.; Antoniassi, M.P.; Okada, F.K.; Bertolla, R.P. Effect of in vitro vitamin e (alpha-tocopherol) supplementation in human spermatozoon submitted to oxidative stress. Andrologia 2018, 50, e12959. [Google Scholar] [CrossRef]
- Vernet, P.; Aitken, R.J.; Drevet, J.R. Antioxidant strategies in the epididymis. Mol. Cell Endocrinol. 2004, 216, 31–39. [Google Scholar] [CrossRef]
- Alvarez, J.G.; Storey, B.T. Role of glutathione peroxidase in protecting mammalian spermatozoa from loss of motility caused by spontaneous lipid peroxidation. Gamete Res. 1989, 23, 77–90. [Google Scholar] [CrossRef]
- Kaneko, T.; Iuchi, Y.; Kobayashi, T.; Fujii, T.; Saito, H.; Kurachi, H.; Fujii, J. The expression of glutathione reductase in the male reproductive system of rats supports the enzymatic basis of glutathione function in spermatogenesis. Eur. J. Biochem. 2002, 269, 1570–1578. [Google Scholar] [CrossRef]
- Omu, A.E.; Al-Azemi, M.K.; Kehinde, E.O.; Anim, J.T.; Oriowo, M.A.; Mathew, T.C. Indications of the mechanisms involved in improved sperm parameters by zinc therapy. Med. Princ. Pract. 2008, 17, 108–116. [Google Scholar] [CrossRef]
- Awoniyi, D.O.; Aboua, Y.G.; Marnewick, J.; Brooks, N. The effects of rooibos (aspalathus linearis), green tea (camellia sinensis) and commercial rooibos and green tea supplements on epididymal sperm in oxidative stress-induced rats. Phytother. Res. 2012, 26, 1231–1239. [Google Scholar] [CrossRef]
- Kumar, K.; Deka, D.; Singh, A.; Mitra, D.K.; Vanitha, B.R.; Dada, R. Predictive value of DNA integrity analysis in idiopathic recurrent pregnancy loss following spontaneous conception. J. Assist. Reprod. Genet. 2012, 29, 861–867. [Google Scholar] [CrossRef] [Green Version]
- Bakos, H.W.; Thompson, J.G.; Feil, D.; Lane, M. Sperm DNA damage is associated with assisted reproductive technology pregnancy. Int. J. Androl. 2008, 31, 518–526. [Google Scholar] [CrossRef]
- Gallagher, J.E.; Vine, M.F.; Schramm, M.M.; Lewtas, J.; George, M.H.; Hulka, B.S.; Everson, R.B. 32p-postlabeling analysis of DNA adducts in human sperm cells from smokers and nonsmokers. Cancer Epidemiol. Biomark. Prev. 1993, 2, 581–585. [Google Scholar]
- Brahem, S.; Mehdi, M.; Landolsi, H.; Mougou, S.; Elghezal, H.; Saad, A. Semen parameters and sperm DNA fragmentation as causes of recurrent pregnancy loss. Urology 2011, 78, 792–796. [Google Scholar] [CrossRef]
- Thomson, L.K.; Zieschang, J.A.; Clark, A.M. Oxidative deoxyribonucleic acid damage in sperm has a negative impact on clinical pregnancy rate in intrauterine insemination but not intracytoplasmic sperm injection cycles. Fertil. Steril. 2011, 96, 843–847. [Google Scholar] [CrossRef]
- Gawecka, J.E.; Marh, J.; Ortega, M.; Yamauchi, Y.; Ward, M.A.; Ward, W.S. Mouse zygotes respond to severe sperm DNA damage by delaying paternal DNA replication and embryonic development. PLoS ONE 2013, 8, e56385. [Google Scholar] [CrossRef]
- Tesarik, J.; Greco, E.; Mendoza, C. Late, but not early, paternal effect on human embryo development is related to sperm DNA fragmentation. Hum. Reprod. 2004, 19, 611–615. [Google Scholar] [CrossRef] [Green Version]
- Binder, N.K.; Mitchell, M.; Gardner, D.K. Parental diet-induced obesity leads to retarded early mouse embryo development and altered carbohydrate utilisation by the blastocyst. Reprod. Fertil. Dev. 2012, 24, 804–812. [Google Scholar] [CrossRef]
- Mitchell, M.; Bakos, H.W.; Lane, M. Paternal diet-induced obesity impairs embryo development and implantation in the mouse. Fertil. Steril. 2011, 95, 1349–1353. [Google Scholar] [CrossRef]
- Lane, M.; Zander-Fox, D.L.; Robker, R.L.; McPherson, N.O. Peri-conception parental obesity, reproductive health, and transgenerational impacts. Trends Endocrinol. Metab. 2015, 26, 84–90. [Google Scholar] [CrossRef]
- McPherson, N.O.; Fullston, T.; Aitken, R.J.; Lane, M. Paternal obesity, interventions, and mechanistic pathways to impaired health in offspring. Ann. Nutr. Metab. 2014, 64, 231–238. [Google Scholar] [CrossRef]
- Chen, Q.; Yan, M.; Cao, Z.; Li, X.; Zhang, Y.; Shi, J.; Feng, G.H.; Peng, H.; Zhang, X.; Zhang, Y.; et al. Sperm tsrnas contribute to intergenerational inheritance of an acquired metabolic disorder. Science 2016, 351, 397–400. [Google Scholar] [CrossRef]
- Sharma, U.; Conine, C.C.; Shea, J.M.; Boskovic, A.; Derr, A.G.; Bing, X.Y.; Belleannee, C.; Kucukural, A.; Serra, R.W.; Sun, F.; et al. Biogenesis and function of trna fragments during sperm maturation and fertilization in mammals. Science 2016, 351, 391–396. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.; Shi, J.; Tuorto, F.; Li, X.; Liu, Y.; Liebers, R.; Zhang, L.; Qu, Y.; Qian, J.; et al. Dnmt2 mediates intergenerational transmission of paternally acquired metabolic disorders through sperm small non-coding rnas. Nat. Cell Biol. 2018, 20, 535–540. [Google Scholar] [CrossRef]
- Sarker, G.; Sun, W.F.; Rosenkranz, D.; Pelczar, P.; Opitz, L.; Efthymiou, V.; Wolfrum, C.; Peleg-Raibstein, D. Maternal overnutrition programs hedonic and metabolic phenotypes across generations through sperm tsrnas. Proc. Natl. Acad. Sci. USA 2019, 116, 10547–10556. [Google Scholar] [CrossRef]
- Thompson, D.M.; Lu, C.; Green, P.J.; Parker, R. Trna cleavage is a conserved response to oxidative stress in eukaryotes. RNA 2008, 14, 2095–2103. [Google Scholar] [CrossRef]
- Abdali, D.; Samson, S.E.; Grover, A.K. How effective are antioxidant supplements in obesity and diabetes? Med. Princ. Pract. 2015, 24, 201–215. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet | CD | CD + S | HFD | HFD + S | |
|---|---|---|---|---|---|
| Main Ingredients | |||||
| Total fat (%) | 6.0 | 6.0 | 21.0 | 21.0 | |
| Sucrose (%) | 34.1 | 34.1 | 34.1 | 34.1 | |
| Wheat starch (%) | 30.5 | 30.5 | 15.5 | 15.5 | |
| Vitamins and Minerals (micronutrients) | |||||
| Zinc (zinc sulphate monohydrate) | 52 mg/kg | 61 mg/kg | 52 mg/kg | 61 mg/kg | |
| Selenium | 0.3 mg/kg | 0.44 mg/kg | 0.3 mg/kg | 0.44 mg/kg | |
| Lycopene | - | 0.3 mg/kg | - | 0.3 mg/kg | |
| Vitamin E (alpha-tocopherol acetate) | 64 mg/kg | 78 mg/kg | 64 mg/kg | 78 mg/kg | |
| Vitamin C (ascorbic acid) | - | 700 mg/kg | - | 700 mg/kg | |
| Folic acid | 1 mg/kg | 1.5 mg/kg | 1 mg/kg | 1.5 mg/kg | |
| Green tea extract | - | 0.95 mg/kg | - | 0.95 mg/kg | |
| CD (n = 12) | CD + S (n = 12) | HFD (n = 12) | HFD + S (n = 12) | |
|---|---|---|---|---|
| Total body weight (g) | 26.2 ± 0.6 a | 25.2 ± 0.5 a | 29.2 ± 0.5 b | 30.4 ± 0.8 b |
| Adipose tissue (% of total body weight | ||||
| Peri-renal fat | 0.21 ± 0.02 a | 0.20 ± 0.0 2 a | 0.32 ± 0.02 b | 0.37 ± 0.06 b |
| Retroperitoneal fat | 0.47 ± 0.03 a | 0.45 ± 0.03 a | 0.87 ± 0.06 b | 0.84 ± 0.08 b |
| Omental fat | 0.97 ± 0.07 a | 0.95 ± 0.06 a | 1.36 ± 0.08 b | 1.46 ± 0.05 b |
| Dorsal fat | 0.62 ± 0.03 a | 0.67 ± 0.04 a | 0.82 ± 0.04 b | 0.90 ± 0.04 b |
| Gonadal fat | 2.40 ± 0.12 a | 2.21 ± 0.13 a | 4.04 ± 0.19 b | 4.28 ± 0.19 b |
| Sum of adipose tissues | 4.66 ± 0.18 a | 4.49 ± 0.23 a | 7.41 ± 0.23 b | 7.86 ± 0.30 b |
| Organs (g) | ||||
| Left testis | 0.082 ± 0.005 | 0.085 ± 0.003 | 0.085 ± 0.006 | 0.085 ± 0.001 |
| Right testis | 0.082 ± 0.002 | 0.088 ± 0.001 | 0.150 ± 0.062 | 0.148 ± 0.063 |
| Seminal vesicles | 0.286 ± 0.013 a | 0.272 ± 0.010 a | 0.291 ± 0.006 a | 0.339 ± 0.009 b |
| Liver | 1.20 ± 0.05 ab | 1.11 ± 0.04 a | 1.19 ± 0.03 ab | 1.30 ± 0.07 b |
| Pancreas | 0.136 ± 0.008 | 0.135 ± 0.009 | 0.140 ± 0.008 | 0.149 ± 0.004 |
| Left kidney | 0.198 ± 0.008 | 0.183 ± 0.008 | 0.194 ± 0.005 | 0.189 ± 0.006 |
| Right kidney | 0.203 ± 0.011 | 0.190 ± 0.007 | 0.199 ± 0.006 | 0.207 ± 0.008 |
| Metabolites and hormones | ||||
| Glucose (mmol/L−1) | 10.6 ± 0.6 | 9.5 ± 0.3 | 10.7 ± 0.8 | 10.8 ± 0.7 |
| Cholesterol (mmol/L−1) | 3.1 ± 0.2 a | 3.0 ± 0.2 a | 4.2 ± 0.2 b | 4.3 ± 0.2 b |
| HDL (mmol/L−1) | 2.7 ± 0.2 a | 2.8 ± 0.1 a | 3.7 ± 0.02 b | 3.8 ± 0.3 b |
| Triglycerides (mmol/L−1) | 0.46 ± 0.04 a | 0.45 ± 0.03 a | 0.60 ± 0.05 b | 0.62 ± 0.06 b |
| NEFA (mmol/L−1) | 0.73 ± 0.03 | 0.72 ± 0.06 | 0.72 ± 0.03 | 0.77 ± 0.04 |
| Testosterone (nmol/L−1) | 0.05 ± 0.01 | 0.07 ± 0.01 | 0.05 ± 0.01 | 0.06 ± 0.01 |
| CD (n = 12) | CD + S (n = 12) | HFD (n = 12) | HFD + S (n = 12) | |
|---|---|---|---|---|
| Sperm motility | ||||
| Progressive (%) | 22.1 ± 2.9 | 16.0 ± 1.8 | 24.1 ± 3.9 | 20.4 ± 2.9 |
| Non progressive (%) | 42.3 ± 3.9 | 43.8 ± 3.2 | 36.8 ± 2.9 | 35.9 ± 3.5 |
| Immotile (%) | 35.5 ± 2.6 | 40.2 ± 4.3 | 39.1 ± 3.2 | 43.5 ± 3.2 |
| Total motile (%) | 64.5 ± 2.6 | 59.8 ± 4.3 | 60.9 ± 3.2 | 56.3 ± 3.2 |
| Sperm morphology | ||||
| Normal (%) | 53.5 ± 1.1 a | 58.5 ± 1.2 b | 45.1 ± 3.0 c | 47.6 ± 2.6 ac |
| Head defect (%) | 22.1 ± 1.1 | 21.6 ± 1.6 | 22.5 ± 2.2 | 20.7 ± 2.9 |
| Tail and mid piece defect (%) | 24.3 ± 0.6 a | 18.7 ± 1.3 b | 33.6 ± 1.4 c | 31.7 ± 1.0 c |
| Sperm Capacitation | ||||
| Capacitated sperm (%) | 90.6 ± 1.0 a | 89.5 ± 1.1 ab | 87.3 ± 0.9 b | 86.9 ± 0.6 b |
| Non-capacitated sperm (%) | 4.2 ± 0.6 a | 5.6 ± 0.6 ab | 5.3 ± 0.5 ab | 6.1 ± 0.4 b |
| Acrosome reacted (%) | 4.7 ± 0.4 a | 4.9 ± 0.6 a | 7.5 ± 0.6 b | 7.1 ± 0.6 b |
| Sperm binding to the zona pellucida of MII oocyte | ||||
| Mean of sperm | 20.4 ± 2.7 | 23.6 ± 3.5 | 22.1 ± 3.1 | 22.7 ± 3.1 |
| CD | CD + S | HFD | HFD + S | |
|---|---|---|---|---|
| Litter size | 8.8 ± 0.9 | 8.7 ± 0.7 | 8.9 ± 0.4 | 8.6 ± 0.5 |
| Fetal weight (mg) | 738.7 ± 23.4 a | 792.3 ± 22.9 ab | 805.4 ± 18.3 b | 783.5 ± 19.5 ab |
| Crown rump length (mm) | 17.4 ± 0.5 | 18.3 ± 0.6 | 18.6 ± 0.4 | 18.8 ± 0.5 |
| Placenta weight (mg) | 106.8 ± 4.1 a | 88.2 ± 4.1 b | 91.5 ± 3.2 b | 85.4 ± 3.4 b |
| Fetal:placenta weight ratio | 7.3 ± 0.4 a | 9.0 ± 0.4 b | 8.9 ± 0.3 b | 9.1 ± 0.3 b |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McPherson, N.O.; Shehadeh, H.; Fullston, T.; Zander-Fox, D.L.; Lane, M. Dietary Micronutrient Supplementation for 12 Days in Obese Male Mice Restores Sperm Oxidative Stress. Nutrients 2019, 11, 2196. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11092196
McPherson NO, Shehadeh H, Fullston T, Zander-Fox DL, Lane M. Dietary Micronutrient Supplementation for 12 Days in Obese Male Mice Restores Sperm Oxidative Stress. Nutrients. 2019; 11(9):2196. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11092196
Chicago/Turabian StyleMcPherson, Nicole O., Helana Shehadeh, Tod Fullston, Deirdre L. Zander-Fox, and Michelle Lane. 2019. "Dietary Micronutrient Supplementation for 12 Days in Obese Male Mice Restores Sperm Oxidative Stress" Nutrients 11, no. 9: 2196. https://0-doi-org.brum.beds.ac.uk/10.3390/nu11092196