Role of Fermented Goat Milk on Liver Gene and Protein Profiles Related to Iron Metabolism during Anemia Recovery

 , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
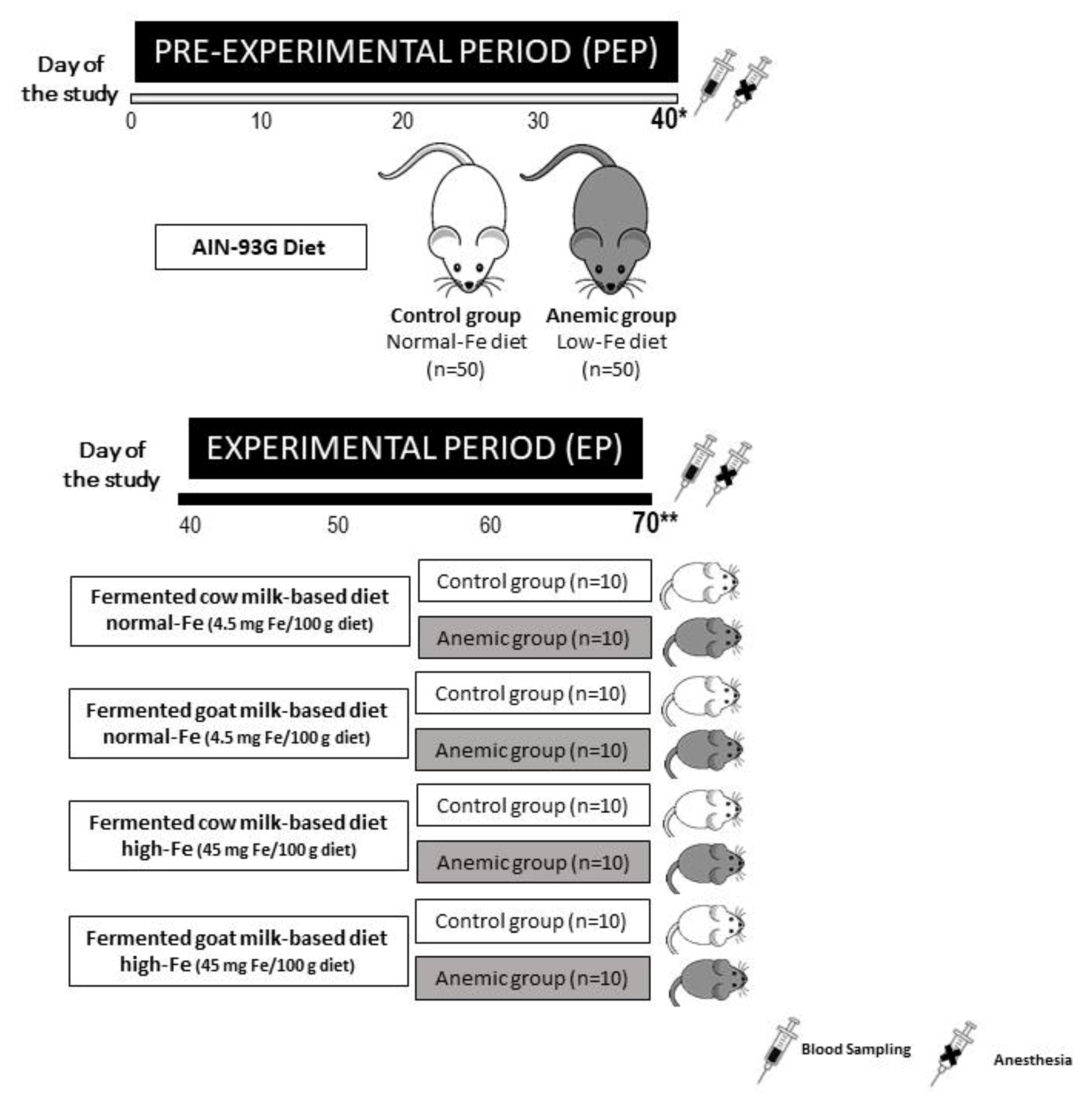
2.1. Animals and Experimental Design
2.2. Diets Preparation
2.3. Hematological Tests
2.4. Transferrin Saturation, Serum Iron, Total Iron Binding Capacity (TIBC), Serum Ferritin and Serum Hepcidin
2.5. Hepatosomatic Index, Hepatic Iron Concentration and Transaminases Analysis
2.6. RNA Extraction and Quantitative Real Time PCR
2.7. Western Blotting and Immunocytochemistry
2.8. Statistical Analysis
3. Results
3.1. Effect of Iron Deficiency on Hepatosomatic Index, Liver Iron Content and Serum Levels of Aspartate Aminotransferase and Alanine Aminotransferase
3.2. Effect of Fermented Milk-Based Diets on Hepatosomatic Index, Liver Iron Content and Serum Levels of Aspartate Aminotransferase and Alanine Aminotransferase during Anemia Recovery
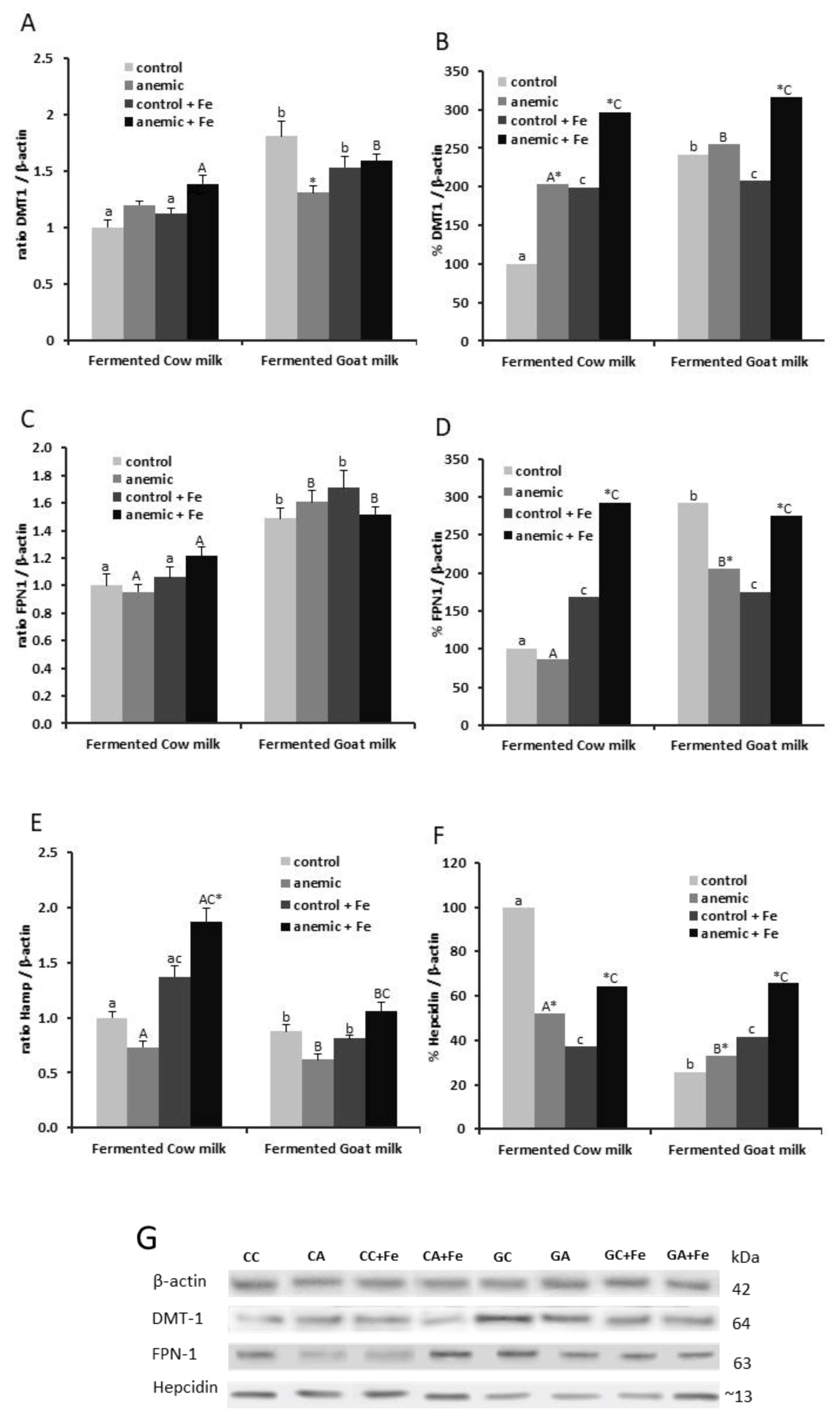
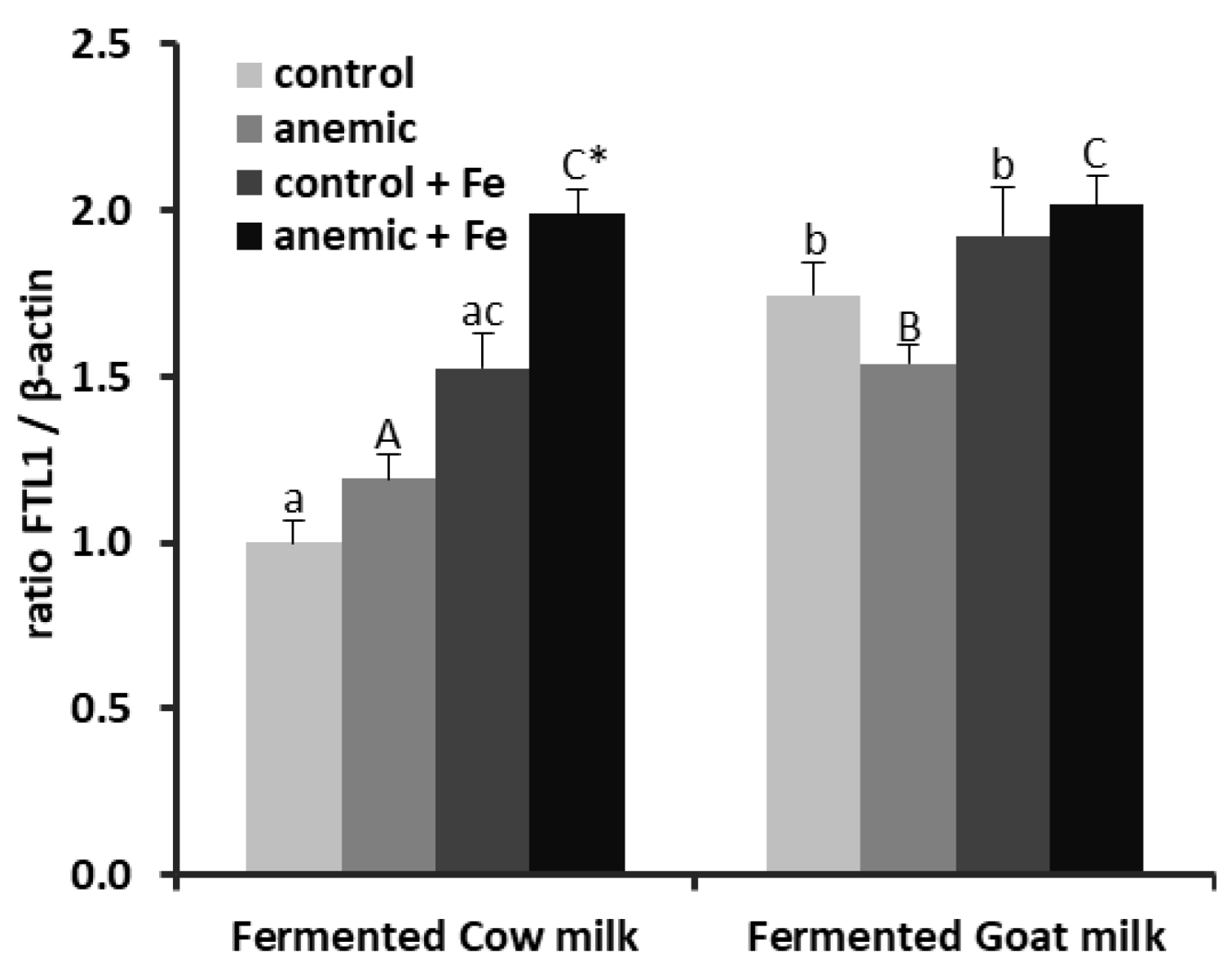
3.3. Effect of Fermented Milk-Based Diets on Liver Iron Homeostasis during Anemia Recovery
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McLean, E.; Cogswell, M.; Egli, I.; Wojdyla, D.; de Benoist, B. Worldwide prevalence of anaemia, WHO Vitamin and Mineral Nutrition Information System, 1993–2005. Public Health Nutr. 2009, 12, 444–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, D.F. The Regulation of Iron Absorption and Homeostasis. Clin. Biochem. Rev. 2016, 37, 51–62. [Google Scholar]
- Hentze, M.W.; Muckenthaler, M.U.; Galy, B.; Camaschella, C. Two to tango: Regulation of Mammalian iron metabolism. Cell 2010, 142, 24–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maio, N.; Jain, A.; Rouault, T.A. Mammalian iron-sulfur cluster biogenesis: Recent insights into the roles of frataxin, acyl carrier protein and ATPase-mediated transfer to recipient proteins. Curr. Opin. Chem. Biol. 2020, 55, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Weiss, G.; Goodnough, L.T. Anemia of chronic disease. N. Engl. J. Med. 2005, 352, 1011–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rishi, G.; Subramaniam, V.N. The liver in regulation of iron homeostasis. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 313, G157–G165. [Google Scholar] [CrossRef] [Green Version]
- Ross, S.L.; Tran, L.; Winters, A.; Lee, K.J.; Plewa, C.; Foltz, I.; King, C.; Miranda, L.P.; Allen, J.; Beckman, H.; et al. Molecular mechanism of hepcidin-mediated ferroportin internalization requires ferroportin lysines, not tyrosines or JAK-STAT. Cell Metab. 2012, 15, 905–917. [Google Scholar] [CrossRef] [Green Version]
- Vyoral, D.; Jiri, P. Therapeutic potential of hepcidin—The master regulator of iron metabolism. Pharmacol. Res. 2017, 115, 242–254. [Google Scholar] [CrossRef]
- Moreno-Fernandez, J.; Diaz-Castro, J.; Pulido-Moran, M.; Alferez, M.J.; Boesch, C.; Sanchez-Alcover, A.; Lopez-Aliaga, I. Fermented Goat’s Milk Consumption Improves Duodenal Expression of Iron Homeostasis Genes during Anemia Recovery. J. Agric. Food Chem. 2016, 64, 2560–2568. [Google Scholar] [CrossRef]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C., Jr. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef]
- Pallares, I.; Lisbona, F.; Aliaga, I.L.; Barrionuevo, M.; Alferez, M.J.; Campos, M.S. Effect of iron deficiency on the digestive utilization of iron, phosphorus, calcium and magnesium in rats. Br. J. Nutr. 1993, 70, 609–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raja, K.B.; Simpson, R.J.; Peters, T.J. Intestinal iron absorption studies in mouse models of iron-overload. Br. J. Haematol. 1994, 86, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Fernandez, J.; Diaz-Castro, J.; Alferez, M.J.M.; Lopez-Aliaga, I. Iron Deficiency and Neuroendocrine Regulators of Basal Metabolism, Body Composition and Energy Expenditure in Rats. Nutrients 2019, 11, 631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolman, K.G.; Re, J.R. Liver function. In Tietz Textbook of Clinical Chemistry; Burtis, C.A., Ashwood, E.R., Eds.; W.B. Saunders: Philadelphia, PA, USA, 1999; pp. 1125–1177. [Google Scholar]
- Diaz-Castro, J.; Moreno-Fernandez, J.; Pulido-Moran, M.; Alferez, M.J.M.; Robles-Rebollo, M.; Ochoa, J.J.; Lopez-Aliaga, I. Changes in Adiposity and Body Composition during Anemia Recovery with Goat or Cow Fermented Milks. J. Agric. Food Chem. 2017, 65, 4057–4065. [Google Scholar] [CrossRef]
- Blankenhaus, B.; Braza, F.; Martins, R.; Bastos-Amador, P.; Gonzalez-Garcia, I.; Carlos, A.R.; Mahu, I.; Faisca, P.; Nunes, J.M.; Ventura, P.; et al. Ferritin regulates organismal energy balance and thermogenesis. Mol. Metab. 2019, 24, 64–79. [Google Scholar] [CrossRef]
- Trinder, D.; Oates, P.S.; Thomas, C.; Sadleir, J.; Morgan, E.H. Localisation of divalent metal transporter 1 (DMT1) to the microvillus membrane of rat duodenal enterocytes in iron deficiency, but to hepatocytes in iron overload. Gut 2000, 46, 270–276. [Google Scholar] [CrossRef]
- Kelleher, S.L.; Lonnerdal, B. Low vitamin a intake affects milk iron level and iron transporters in rat mammary gland and liver. J. Nutr. 2005, 135, 27–32. [Google Scholar] [CrossRef] [Green Version]
- Alférez, M.J.M.; López-Aliaga, I.; Nestares, T.; Díaz-Castro, J.; Barrionuevo, M.; Ros, P.B.; Campos, M.S. Dietary goat milk improves iron bioavailability in rats with induced ferropenic anaemia in comparison with cow milk. Int. Dairy J. 2006, 16, 813–821. [Google Scholar] [CrossRef]
- Diaz-Castro, J.; Pulido, M.; Alferez, M.J.; Ochoa, J.J.; Rivas, E.; Hijano, S.; Lopez-Aliaga, I. Goat milk consumption modulates liver divalent metal transporter 1 (DMT1) expression and serum hepcidin during Fe repletion in Fe-deficiency anemia. J. Dairy Sci. 2014, 97, 147–154. [Google Scholar] [CrossRef] [Green Version]
- Nemeth, E.; Valore, E.V.; Territo, M.; Schiller, G.; Lichtenstein, A.; Ganz, T. Hepcidin, a putative mediator of anemia of inflammation, is a type II acute-phase protein. Blood 2003, 101, 2461–2463. [Google Scholar] [CrossRef]
- Ganz, T. Hepcidin and iron regulation, 10 years later. Blood 2011, 117, 4425–4433. [Google Scholar] [CrossRef] [Green Version]
- Ueda, N.; Takasawa, K. Impact of Inflammation on Ferritin, Hepcidin and the Management of Iron Deficiency Anemia in Chronic Kidney Disease. Nutrients 2018, 10, 1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.Y.; Uprichard, J. Ferritin and iron studies in anaemia and chronic disease. Ann. Clin. Biochem. 2017, 54, 43–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Aliaga, I.; Garcia-Pedro, J.D.; Moreno-Fernandez, J.; Alferez, M.J.M.; Lopez-Frias, M.; Diaz-Castro, J. Fermented goat milk consumption improves iron status and evokes inflammatory signalling during anemia recovery. Food Funct. 2018, 9, 3195–3201. [Google Scholar] [CrossRef] [PubMed]
- Nasopoulou, C.; Smith, T.; Detopoulou, M.; Tsikrika, C.; Papaharisis, L.; Barkas, D.; Zabetakis, I. Structural elucidation of olive pomace fed sea bass (Dicentrarchus labrax) polar lipids with cardioprotective activities. Food Chem. 2014, 145, 1097–1105. [Google Scholar] [CrossRef] [PubMed]
- Tsorotioti, S.E.; Nasopoulou, C.; Detopoulou, M.; Sioriki, E.; Demopoulos, C.A.; Zabetakis, I. In vitro anti-atherogenic properties of traditional Greek cheese lipid fractions. Dairy Sci. Technol. 2014, 94, 269–281. [Google Scholar] [CrossRef] [Green Version]
- Maras, J.S.; Maiwall, R.; Harsha, H.C.; Das, S.; Hussain, M.S.; Kumar, C.; Bihari, C.; Rastogi, A.; Kumar, M.; Trehanpati, N.; et al. Dysregulated iron homeostasis is strongly associated with multiorgan failure and early mortality in acute-on-chronic liver failure. Hepatology 2015, 61, 1306–1320. [Google Scholar] [CrossRef]
- Diaz-Castro, J.; Perez-Sanchez, L.J.; Ramirez Lopez-Frias, M.; Lopez-Aliaga, I.; Nestares, T.; Alferez, M.J.; Ojeda, M.L.; Campos, M.S. Influence of cow or goat milk consumption on antioxidant defence and lipid peroxidation during chronic iron repletion. Br. J. Nutr. 2012, 108, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Alferez, M.J.; Rivas, E.; Diaz-Castro, J.; Hijano, S.; Nestares, T.; Moreno, M.; Campos, M.S.; Serrano-Reina, J.A.; Lopez-Aliaga, I. Folic acid supplemented goat milk has beneficial effects on hepatic physiology, haematological status and antioxidant defence during chronic Fe repletion. J. Dairy Res. 2015, 82, 86–94. [Google Scholar] [CrossRef]
- Knutson, M.D. Iron transport proteins: Gateways of cellular and systemic iron homeostasis. J. Biol. Chem. 2017, 292, 12735–12743. [Google Scholar] [CrossRef] [Green Version]
- Muckenthaler, M.U.; Rivella, S.; Hentze, M.W.; Galy, B. A Red Carpet for Iron Metabolism. Cell 2017, 168, 344–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gozzelino, R.; Soares, M.P. Coupling heme and iron metabolism via ferritin H chain. Antioxid. Redox Signal. 2014, 20, 1754–1769. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Constituents | Pre-Experimental Period | Experimental Period | |
|---|---|---|---|
| g/100 g Diet | Fermented Milk Diets 2 | ||
| AIN-93G 1 | Cow Milk | Goat Milk | |
| Protein | 20.00 | 20.50 | 20.60 |
| Lactose | - | 29.50 | 29.10 |
| Fat | 10.00 | 10.00 | 29.10 |
| Wheat starch | 50.00 | 20.00 | 20.30 |
| Constant ingredients 3 | 20.00 | 20.00 | 20.00 |
| Gene | Direction | Primer Sequence (5′→3′) | Annealing Temperature | Size (bp) |
|---|---|---|---|---|
| β-Actin | Forward | GGGGTGTTGAAGGTCTCAAA | 57 °C | 165 |
| Reverse | TGTCACCAACTGGGACGATA | |||
| DMT1 | Forward | GGCATGTGGCACTGTATGTG | 59 °C | 163 |
| Reverse | CCGCTGGTATCTTCGCTCAG | |||
| FPN1 | Forward | GAACAAGAACCCACCTGTGC | 57 °C | 191 |
| Reverse | AGGATGGAACCACTCAGTCC | |||
| HAMP | Forward | CCTATCTCCGGCAACAGACG | 59 °C | 121 |
| Reverse | GGGAAGTTGGTGTCTCGCTT | |||
| FTL1 | Forward | GCCCTGGAGAAGAACCTGAA | 59 °C | 247 |
| Reverse | AGTCGTGCTTCAGAGTGAGG |
| Control Group (n = 20) | Anemic Group (n = 20) | |
|---|---|---|
| Body weight (g) | 239.7 ± 3.9 | 201.15 ± 2.9 ** |
| Liver weight (g) | 6.324 ± 0.31 | 6.129 ± 0.31 |
| HSI (%) | 2.55 ± 0.07 | 2.89 ± 0.09 * |
| Liver iron content (µg/g dry weight) | 615.25 ± 31.10 | 432.31 ± 24.07 ** |
| AST (UI/L) | 103.58 ± 8.93 | 228.04 ± 18.45 ** |
| ALT (UI/L) | 24.57 ± 1.16 | 52.28 ± 2.73 ** |
| Fermented Cow Milk | Fermented Goat Milk | 2-WAY ANOVA | ||||||
|---|---|---|---|---|---|---|---|---|
| Fe Content | Control Group | Anemic Group | Control Group | Anemic Group | Diet | Anemia | Fe Content | |
| Body weight (g) | Normal | 365.23 ± 8.61 a | 347.21 ± 8.39 A,* | 278.98 ± 3.70 b | 255.41 ± 2.85 B,* | <0.001 | <0.01 | <0.05 |
| High | 339.42 ± 5.18 a,c | 329.22 ± 5.81 A,C | 287.27 ± 4.92 b | 267.57 ± 4.03 B | <0.001 | NS 1 | ||
| Liver weight (g) | Normal | 6.528 ± 0.24 a | 6.269 ± 0.10 A | 8.391 ± 0.23 b | 8.521 ± 0.21 B | <0.001 | NS | <0.05 |
| High | 6.764 ± 0.2 a | 6.555 ± 0.12 A | 7.692 ± 0.22 b,c | 7.934 ± 0.22 B,C | <0.01 | NS | ||
| HSI (%) | Normal | 1.84 ± 0.04 a | 1.77 ± 0.02 A | 2.95 ± 0.03 b | 3.27 ± 0.05 B | <0.001 | NS | NS |
| High | 1.79 ± 0.03 a | 1.82 ± 0.03 A | 2.65 ± 0.04 b | 3.12 ± 0.04 B | <0.001 | NS | ||
| Liver iron content (µg/g dry weight) | Normal | 559.56 ± 28.72 a | 401.56 ± 24.50 A,* | 666.45 ± 33.21 b | 489.32 ± 29.64 B,* | <0.01 | <0.01 | <0.01 |
| High | 832.25 ± 32.56 a,c | 782.32 ± 33.55 A,C,* | 735.67 ± 29.33 b,c | 657.15 ± 29.22 B,C,* | <0.01 | <0.01 | ||
| AST (UI/L) | Normal | 107.62 ± 4.29 a | 80.86 ± 4.25 A,* | 67.99 ± 2.75 b | 61.11 ± 2.12 B | <0.01 | <0.05 | NS |
| High | 82.92 ± 4.15 a,c | 78.19 ± 3.82 A | 60.43 ± 1.10 b | 68.47 ± 2.03 B | <0.01 | NS | ||
| ALT (UI/L) | Normal | 28.91 ± 1.34 a | 27.77 ± 3.91 A | 23.14 ± 1.9 b | 16.49 ± 0.76 B,* | <0.01 | <0.01 | NS |
| High | 24.0.4 ± 1.73 a | 19.00 ± 1.21 A | 19.47 ± 0.53 b | 14.48 ± 0.35 B,* | <0.01 | <0.01 | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno-Fernandez, J.; Alférez, M.J.M.; López-Aliaga, I.; Díaz-Castro, J. Role of Fermented Goat Milk on Liver Gene and Protein Profiles Related to Iron Metabolism during Anemia Recovery. Nutrients 2020, 12, 1336. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12051336
Moreno-Fernandez J, Alférez MJM, López-Aliaga I, Díaz-Castro J. Role of Fermented Goat Milk on Liver Gene and Protein Profiles Related to Iron Metabolism during Anemia Recovery. Nutrients. 2020; 12(5):1336. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12051336
Chicago/Turabian StyleMoreno-Fernandez, Jorge, María J. M. Alférez, Inmaculada López-Aliaga, and Javier Díaz-Castro. 2020. "Role of Fermented Goat Milk on Liver Gene and Protein Profiles Related to Iron Metabolism during Anemia Recovery" Nutrients 12, no. 5: 1336. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12051336