Dietary Protein, Fiber and Coffee Are Associated with Small Intestine Microbiome Composition and Diversity in Patients with Liver Cirrhosis


 , , ,
, , ,
Abstract
:1. Introduction
2. Methods
2.1. Study Population
2.2. Duodenal Biopsy Collection and Processing
2.3. Food Frequency Questionnaire
2.4. Microbiome Characterization
2.5. Statistical Analysis
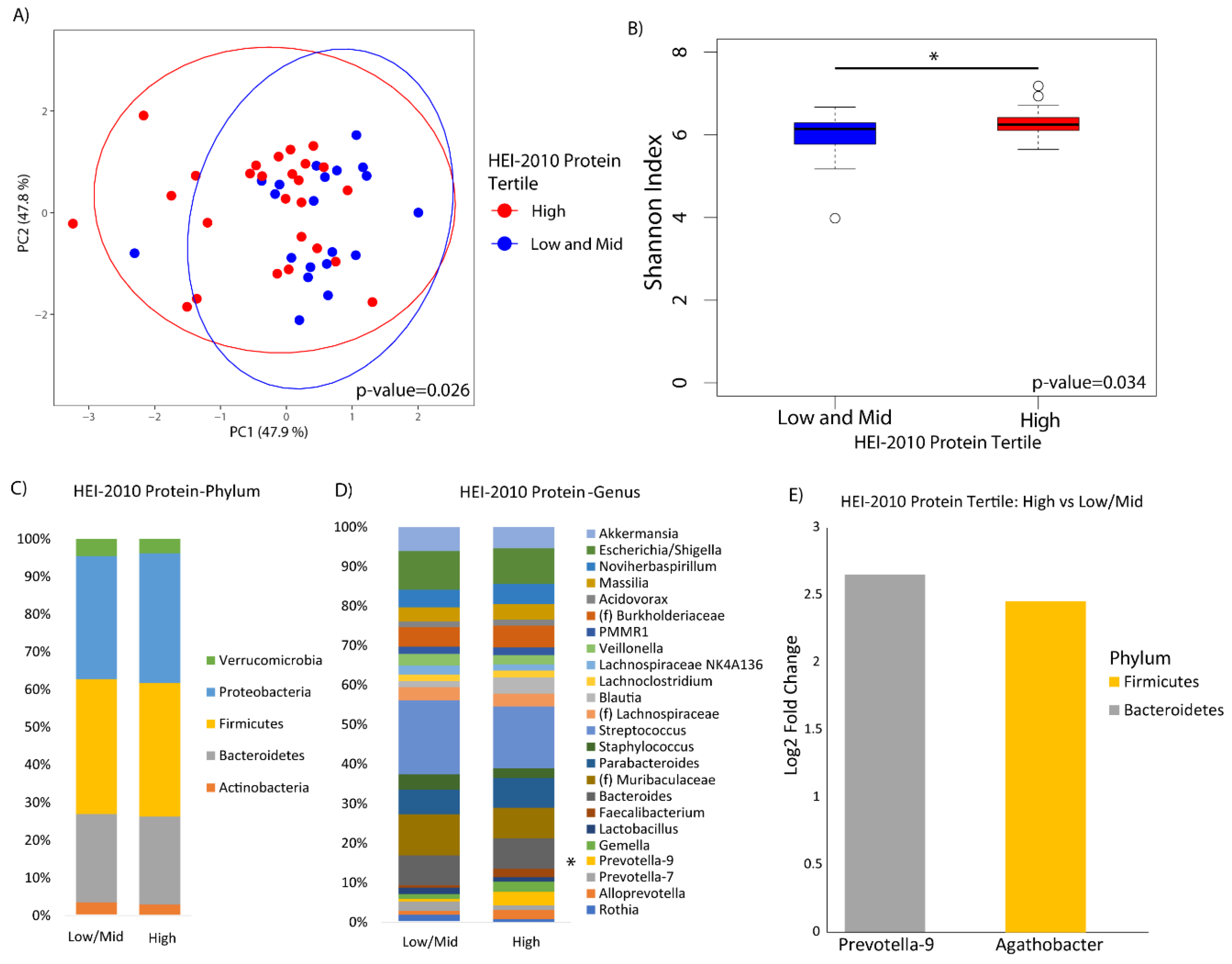
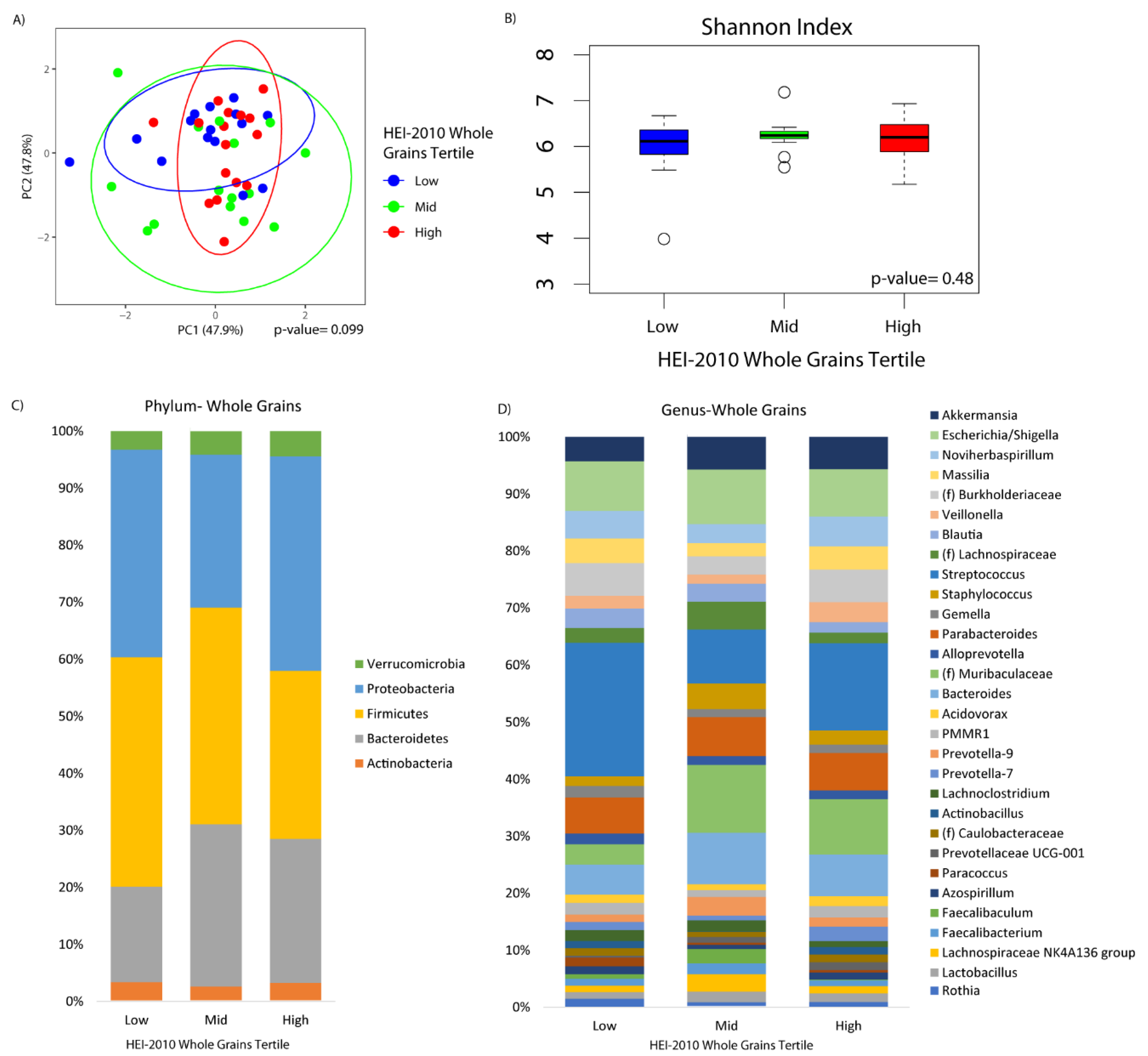
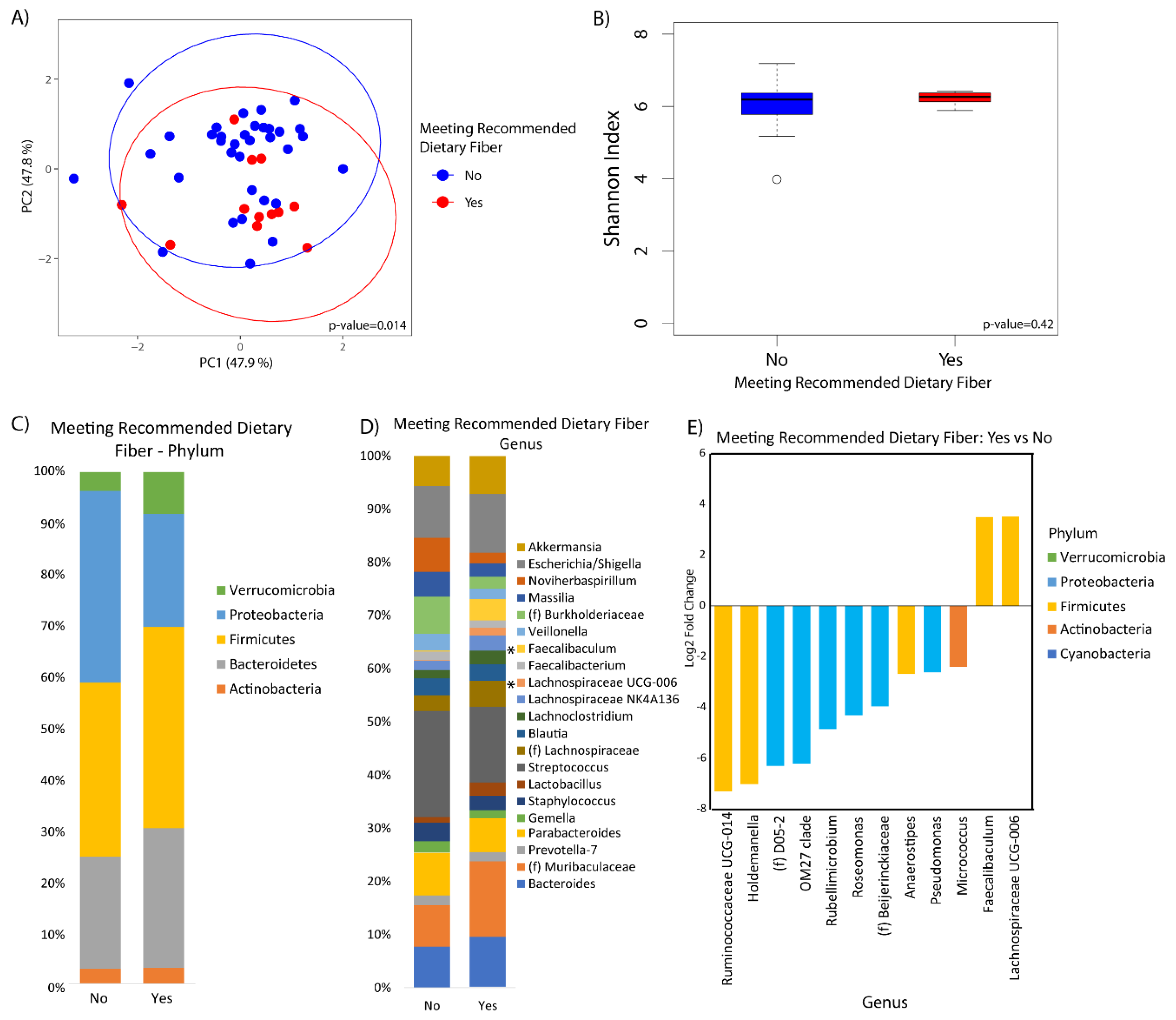
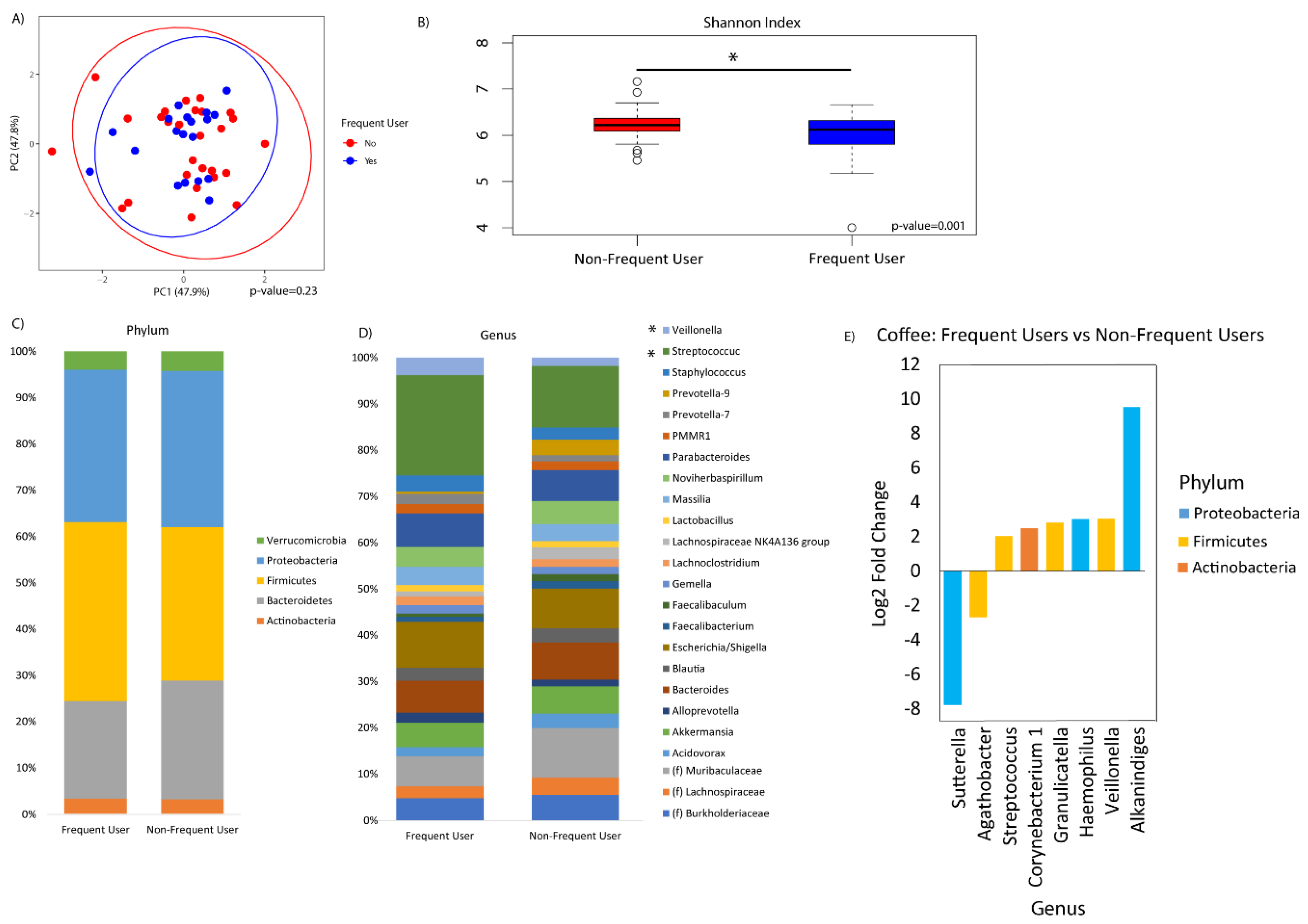
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Mokdad, A.H.; Dwyer-Lindgren, L.; Fitzmaurice, C.; Stubbs, R.W.; Bertozzi-Villa, A.; Morozoff, C.; Charara, R.; Allen, C.; Naghavi, M.; Murray, C.J.L. Trends and patterns of disparities in cancer mortality among US Counties, 1980–2014. JAMA 2017, 317, 388–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvatore, M.; Jeon, J.; Meza, R. Changing trends in liver cancer incidence by race/ethnicity and sex in the US: 1992–2016. Cancer Causes Control. 2019, 30, 1377–1388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Serag, H.B.; Kramer, J.; Duan, Z.; Kanwal, F. Racial Differences in the progression to cirrhosis and hepatocellular carcinoma in HCV-infected veterans. Am. J. Gastroenterol. 2014, 109, 1427–1435. [Google Scholar] [CrossRef] [PubMed]
- Ohtani, N.; Kawada, N. Role of the gut-liver axis in liver inflammation, fibrosis, and cancer: A special focus on the gut microbiota relationship. Hepatol. Commun. 2019, 3, 456–470. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.-W.; Chen, X.-H.; Ren, Z.-G.; Zheng, S.-S. Gut microbial dysbiosis associates hepatocellular carcinoma via the gut-liver axis. Hepatobiliary Pancreat. Dis. Int. 2019, 18, 19–27. [Google Scholar] [CrossRef]
- Pijls, K.E.; Jonkers, D.M.; Elamin, E.E.; Masclee, A.A.M.; Koek, G.H. Intestinal epithelial barrier function in liver cirrhosis: An extensive review of the literature. Liver Int. 2013, 33, 1457–1469. [Google Scholar] [CrossRef]
- Yoshimoto, S.; Loo, T.M.; Atarashi, K.; Kanda, H.; Sato, S.; Oyadomari, S.; Iwakura, Y.; Oshima, K.; Morita, H.; Hattori, M.; et al. Obesity-induced gut microbial metabolite promotes liver cancer through senescence secretome. Nature 2013, 499, 97–101. [Google Scholar] [CrossRef]
- Ma, C.; Han, M.; Heinrich, B.; Fu, Q.; Zhang, Q.; Sandhu, M.; Agdashian, D.; Terabe, M.; Berzofsky, J.A.; Fako, V.; et al. Gut microbiome–mediated bile acid metabolism regulates liver cancer via NKT cells. Science 2018, 360, eaan5931. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.-X.; Schwabe, R.F. The gut microbiome and liver cancer: Mechanisms and clinical translation. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 527–539. [Google Scholar] [CrossRef]
- Yu, L.-X.; Yan, H.-X.; Liu, Q.; Yang, W.; Wu, H.-P.; Dong, W.; Tang, L.; Lin, Y.; He, Y.-Q.; Zou, S.-S.; et al. Endotoxin accumulation prevents carcinogen-induced apoptosis and promotes liver tumorigenesis in rodents. Hepatology 2010, 52, 1322–1333. [Google Scholar] [CrossRef]
- Dapito, D.H.; Mencin, A.; Gwak, G.-Y.; Pradère, J.-P.; Jang, M.-K.; Mederacke, I.; Caviglia, J.M.; Khiabanian, H.; Adeyemi, A.; Bataller, R.; et al. Promotion of Hepatocellular Carcinoma by the Intestinal Microbiota and TLR4. Cancer Cell 2012, 21, 504–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, J.G.; Feng, Y.; Theve, E.J.; Raczynski, A.R.; Fiala, J.; Doernte, A.L.; Williams, M.; McFaline, J.L.; Essigmann, J.M.; Schauer, D.B.; et al. Gut microbes define liver cancer risk in mice exposed to chemical and viral transgenic hepatocarcinogens. Gut 2010, 59, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, J.S.; Heuman, U.M.; Hylemon, P.B.; Sanyal, A.J.; White, M.B.; Monteith, P.; Noble, N.A.; Unser, A.B.; Daita, K.; Fisher, A.R.; et al. Altered profile of human gut microbiome is associated with cirrhosis and its complications. J. Hepatol. 2013, 60, 940–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Yang, F.; Lu, H.; Wang, B.; Chen, Y.; Lei, D.; Wang, Y.; Zhu, B.; Li, L. Characterization of fecal microbial communities in patients with liver cirrhosis. Hepatology 2011, 54, 562–572. [Google Scholar] [CrossRef] [PubMed]
- Ridlon, J.M.; Alves, J.; Hylemon, P.B.; Bajaj, J.S. Cirrhosis, bile acids and gut microbiota: Unraveling a complex relationship. Gut Microbes 2013, 4, 382–387. [Google Scholar] [CrossRef] [Green Version]
- Ren, Z.; Li, A.; Jiang, J.; Zhou, L.; Yu, Z.; Lu, H.; Xie, H.; Chen, X.; Shao, L.; Zhang, R.; et al. Gut microbiome analysis as a tool towards targeted non-invasive biomarkers for early hepatocellular carcinoma. Gut 2018, 68, 1014–1023. [Google Scholar] [CrossRef]
- Zhang, L.; Wu, Y.-N.; Chen, T.; Ren, C.-H.; Li, X.; Liu, G.-X. Relationship between intestinal microbial dysbiosis and primary liver cancer. Hepatobiliary Pancreat. Dis. Int. 2019, 18, 149–157. [Google Scholar] [CrossRef]
- Liu, Q.; Li, F.; Zhuang, Y.; Xu, J.; Wang, J.; Mao, X.; Zhang, Y.; Liu, X. Alteration in gut microbiota associated with hepatitis B and non-hepatitis virus related hepatocellular carcinoma. Gut Pathog. 2019, 11, 1. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2013, 505, 559–563. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.K.; Chang, H.-W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 2017, 15, 73. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.-Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking Long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koumbi, L. Dietary factors can protect against liver cancer development. World J. Hepatol. 2017, 9, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Cheung, K.; Lee, S.S.; Raman, M. Prevalence and mechanisms of malnutrition in patients with advanced liver disease, and nutrition management strategies. Clin. Gastroenterol. Hepatol. 2012, 10, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Henkel, A.S.; Buchman, A.L. Nutritional support in patients with chronic liver disease. Nat. Clin. Pract. Gastroenterol. Hepatol. 2006, 3, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Sam, J.; Nguyen, G.C. Protein-calorie malnutrition as a prognostic indicator of mortality among patients hospitalized with cirrhosis and portal hypertension. Liver Int. 2009, 29, 1396–1402. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Li, S.; Gan, R.; Zhao, C.-N.; Meng, X.; Li, H.-B. Targeting gut microbiota with dietary components on cancer: Effects and potential mechanisms of action. Crit. Rev. Food Sci. Nutr. 2019, 60, 1025–1037. [Google Scholar] [CrossRef]
- Fedirko, V.; Lukanova, A.; Bamia, C.; Trichopolou, A.; Trepo, E.; Nöthlings, U.; Schlesinger, S.; Aleksandrova, K.; Boffetta, P.; Tjønneland, A.; et al. Glycemic index, glycemic load, dietary carbohydrate, and dietary fiber intake and risk of liver and biliary tract cancers in Western Europeans. Ann. Oncol. 2013, 24, 543–553. [Google Scholar] [CrossRef]
- Yang, W.; Ma, Y.; Liu, Y.; Smith-Warner, S.A.; Simon, T.G.; Chong, D.Q.; Qi, Q.; Meyerhardt, J.A.; Giovannucci, E.L.; Chan, A.T.; et al. Association of intake of whole grains and dietary fiber with risk of hepatocellular carcinoma in US Adults. JAMA Oncol. 2019, 5, 879. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, O.J.; Roderick, P.; Buchanan, R.; Fallowfield, J.A.; Hayes, P.C.; Parkes, J. Coffee, including caffeinated and decaffeinated coffee, and the risk of hepatocellular carcinoma: A systematic review and dose-response meta-analysis. BMJ Open 2017, 7, e013739. [Google Scholar] [CrossRef]
- Cox, I.J.; Idilman, R.; Fagan, A.; Turan, D.; Ajayi, L.; Le Guennec, A.D.; Taylor-Robinson, S.D.; Karakaya, F.; Gavis, E.; Atkinson, R.A.; et al. Metabolomics and microbial composition increase insight into the impact of dietary differences in cirrhosis. Liver Int. 2019, 40, 416–427. [Google Scholar] [CrossRef]
- Bogumil, D.; Park, S.-Y.; Le Marchand, L.; Haiman, C.A.; Wilkens, L.R.; Boushey, C.J.; Setiawan, V.W. High-Quality diets are associated with reduced risk of hepatocellular carcinoma and chronic liver disease: The multiethnic cohort. Hepatol. Commun. 2019, 3, 437–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, J.P.; Dong, T.S.; Agopian, V.; Lagishetty, V.; Sundaram, V.; Noureddin, M.; Ayoub, W.S.; Durazo, F.; Benhammou, J.; Enayati, P.; et al. Microbiome and bile acid profiles in duodenal aspirates from patients with liver cirrhosis: The Microbiome, Microbial Markers and Liver Disease Study. Hepatol. Res. 2018, 48, 1108–1117. [Google Scholar] [CrossRef] [Green Version]
- Kristal, A.; Kolar, A.S.; Fisher, J.L.; Plascak, J.J.; Stumbo, P.J.; Weiss, R.; Paskett, E.D. Evaluation of web-based, self-administered, graphical food frequency questionnaire. J. Acad. Nutr. Diet. 2014, 114, 613–621. [Google Scholar] [CrossRef] [Green Version]
- Tong, M.; Jacobs, J.P.; McHardy, I.; Braun, J.M. Sampling of intestinal microbiota and targeted amplification of bacterial 16S rRNA genes for microbial ecologic analysis. Curr. Protoc. Immunol. 2014, 107, 7.41.1–7.41.11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callahan, B.J.; McMurdie, P.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Guenther, P.M.; Casavale, K.O.; Reedy, J.; Kirkpatrick, S.; Hiza, H.A.; Kuczynski, K.J.; Kahle, L.L.; Krebs-Smith, S.M. Update of the Healthy Eating Index: HEI-2010. J. Acad. Nutr. Diet. 2013, 113, 569–580. [Google Scholar] [CrossRef] [Green Version]
- U.S. Department of Health and Human Services; U.S. Department of Agriculture. 2015–2020 Dietary Guidelines for Americans, 8th ed.; December 2015. Available online: http://health.gov/dietaryguidelines/2015/guidelines/ (accessed on 1 May 2020).
- Lozupone, C.A.; Knight, R. Species divergence and the measurement of microbial diversity. FEMS Microbiol. Rev. 2008, 32, 557–578. [Google Scholar] [CrossRef]
- Goodrich, J.K.; Di Rienzi, S.C.; Poole, A.C.; Koren, O.; Walters, W.A.; Caporaso, J.G.; Knight, R.; Ley, R.E. Conducting a microbiome study. Cell 2014, 158, 250–262. [Google Scholar] [CrossRef] [Green Version]
- Martino, C.; Morton, J.T.; Marotz, C.A.; Thompson, L.R.; Tripathi, A.; Knight, R.; Zengler, K. A novel sparse compositional technique reveals microbial perturbations. mSystems 2019, 4, e00016-19. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storey, J.D.; Tibshirani, R. Statistical significance for genomewide studies. Proc. Natl. Acad. Sci. USA 2003, 100, 9440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2: An improved and extensible approach for metagenome inference. bioRxiv 2019, 672295. [Google Scholar]
- Li, W.; Park, Y.; McGlynn, K.A.; Hollenbeck, A.R.; Taylor, P.R.; Goldstein, A.M.; Freedman, N.D. Index-based dietary patterns and risk of incident hepatocellular carcinoma and mortality from chronic liver disease in a prospective study. Hepatology 2014, 60, 588–597. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, A.; Debelius, J.; Brenner, D.A.; Karin, M.; Loomba, R.; Schnabl, B.; Knight, R. The gut-liver axis and the intersection with the microbiome. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 397–411. [Google Scholar] [CrossRef]
- Mailhe, M.; Ricaboni, D.; Vitton, V.; Gonzalez, J.-M.; Bachar, D.; Dubourg, G.; Cadoret, F.; Robert, C.; Delerce, J.; Levasseur, A.; et al. Repertoire of the gut microbiota from stomach to colon using culturomics and next-generation sequencing. BMC Microbiol. 2018, 18, 157. [Google Scholar] [CrossRef]
- Vasapolli, R.; Schütte, K.; Schulz, C.; Vital, M.; Schomburg, D.; Pieper, D.H.; Vilchez-Vargas, R.; Malfertheiner, P. Analysis of transcriptionally active bacteria throughout the gastrointestinal tract of healthy individuals. Gastroenterology 2019, 157, 1081–1092.e3. [Google Scholar] [CrossRef] [Green Version]
- Donaldson, G.P.; Lee, S.M.; Mazmanian, S.K. Gut biogeography of the bacterial microbiota. Nat. Rev. Genet. 2015, 14, 20–32. [Google Scholar] [CrossRef] [Green Version]
- Van Baar, A.C.; Nieuwdorp, M.; Holleman, F.; Soeters, M.R.; Groen, A.K.; Bergman, J.J. The duodenum harbors a broad untapped therapeutic potential. Gastroenterology 2018, 154, 773–777. [Google Scholar] [CrossRef] [Green Version]
- Cherrington, A.D.; Rajagopalan, H.; Maggs, D.; Devière, J. Hydrothermal duodenal mucosal resurfacing. Gastrointest. Endosc. Clin. N. Am. 2017, 27, 299–311. [Google Scholar] [CrossRef]
- Martinez, K.; Hubert, N.; Frazier, K.; Urlass, S.; Musch, M.W.; Ojeda, P.; Pierre, J.F.; Miyoshi, J.; Sontag, T.J.; Cham, C.M.; et al. Small Intestine Microbiota Regulate Host Digestive and Absorptive Adaptive Responses to Dietary Lipids. Cell Host Microbe 2018, 23, 458–469.e5. [Google Scholar] [CrossRef] [PubMed]
- Gheorghe, L.; Iacob, R.; Vadan, R.; Iacob, S.; Gheorghe, C. Improvement of hepatic encephalopathy using a modified high-calorie high-protein diet. Rom. J. Gastroenterol. 2005, 14, 231–238. [Google Scholar] [PubMed]
- Maharshi, S.; Sharma, P.; Sachdeva, S.; Srivastava, S.; Sharma, P. Efficacy of nutritional therapy for patients with cirrhosis and minimal hepatic encephalopathy in a randomized trial. Clin. Gastroenterol. Hepatol. 2016, 14, 454–460.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Córdoba, J.; López-Hellín, J.; Planas, M.; Sabín, P.; Sanpedro, F.; Castro, F.; Esteban, R.; Guardia, J. Normal protein diet for episodic hepatic encephalopathy: Results of a randomized study. J. Hepatol. 2004, 41, 38–43. [Google Scholar] [CrossRef]
- Chen, Y.; Ji, F.; Guo, J.; Shi, D.; Fang, D.; Li, L. Dysbiosis of small intestinal microbiota in liver cirrhosis and its association with etiology. Sci. Rep. 2016, 6, 34055. [Google Scholar] [CrossRef]
- Dong, T.S.; Katzka, W.; Lagishetty, V.; Luu, K.; Hauer, M.; Pisegna, J.; Jacobs, J.P. A microbial signature identifies advanced fibrosis in patients with chronic liver disease mainly due to NAFLD. Sci. Rep. 2020, 10, 2771. [Google Scholar] [CrossRef]
- Larsen, J.M. The immune response toPrevotellabacteria in chronic inflammatory disease. Immunology 2017, 151, 363–374. [Google Scholar] [CrossRef] [Green Version]
- Velikonja, A.; Lipoglavšek, L.; Zorec, M.; Orel, R.; Avguštin, G. Alterations in gut microbiota composition and metabolic parameters after dietary intervention with barley beta glucans in patients with high risk for metabolic syndrome development. Anaerobe 2019, 55, 67–77. [Google Scholar] [CrossRef]
- El Khoury, D.; Cuda, C.; Luhovyy, B.L.; Anderson, G.H. Beta Glucan: Health benefits in obesity and metabolic syndrome. J. Nutr. Metab. 2011, 2012, 851362. [Google Scholar] [CrossRef] [Green Version]
- Martínez, I.; Lattimer, J.M.; Hubach, K.L.; Case, J.A.; Yang, J.; Weber, C.G.; Louk, J.A.; Rose, D.J.; Kyureghian, G.; Peterson, D.A.; et al. Gut microbiome composition is linked to whole grain-induced immunological improvements. ISME J. 2012, 7, 269–280. [Google Scholar] [CrossRef] [Green Version]
- So, D.; Whelan, K.; Rossi, M.; Morrison, M.; Holtmann, G.J.; Kelly, J.T.; Shanahan, E.R.; Staudacher, H.; Campbell, K.L. Dietary fiber intervention on gut microbiota composition in healthy adults: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2018, 107, 965–983. [Google Scholar] [CrossRef] [Green Version]
- Schnorr, S.L.; Candela, M.; Rampelli, S.; Centanni, M.; Consolandi, C.; Basaglia, G.; Turroni, S.; Biagi, E.; Peano, C.; Severgnini, M.; et al. Gut microbiome of the Hadza hunter-gatherers. Nat. Commun. 2014, 5, 3654. [Google Scholar] [CrossRef] [PubMed]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loftfield, E.; Rothwell, J.A.; Sinha, R.; Keski-Rahkonen, P.; Robinot, N.; Albanes, D.; Weinstein, S.J.; Derkach, A.; Sampson, J.N.; Scalbert, A.; et al. Prospective investigation of serum metabolites, coffee drinking, liver cancer incidence, and liver disease mortality. J. Natl. Cancer Inst. 2020, 112, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Bazzaz, B.S.F.; Fakori, M.; Khameneh, B.; Hosseinzadeh, H. Effects of omeprazole and caffeine alone and in combination with gentamicin and ciprofloxacin against antibiotic resistant staphylococcus aureus and escherichia coli strains. J. Pharmacopunct. 2019, 22, 49–54. [Google Scholar] [CrossRef]
- Kang, T.M.; Yuan, J.; Nguyen, A.; Becket, E.; Yang, H.; Miller, J.H. The aminoglycoside antibiotic kanamycin damages DNA bases in Escherichia coli: Caffeine potentiates the DNA-Damaging effects of kanamycin while suppressing cell killing by ciprofloxacin in Escherichia coli and Bacillus anthracis. Antimicrob. Agents Chemother. 2012, 56, 3216–3223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishitsuji, K.; Watanabe, S.; Xiao, J.; Nagatomo, R.; Ogawa, H.; Tsunematsu, T.; Umemoto, H.; Morimoto, Y.; Akatsu, H.; Inoue, K.; et al. Effect of coffee or coffee components on gut microbiome and short-chain fatty acids in a mouse model of metabolic syndrome. Sci. Rep. 2018, 8, 16173. [Google Scholar] [CrossRef] [Green Version]
- Ren, Y.; Wang, C.; Xu, J.-K.; Wang, S. Cafestol and Kahweol: A Review on Their Bioactivities and Pharmacological Properties. Int. J. Mol. Sci. 2019, 20, 4238. [Google Scholar] [CrossRef] [Green Version]
- Scheiman, J.; Luber, J.; Chavkin, T.; Macdonald, T.; Tung, A.; Pham, L.-D.; Wibowo, M.C.; Wurth, R.C.; Punthambaker, S.; Tierney, B.T.; et al. Meta-omics analysis of elite athletes identifies a performance-enhancing microbe that functions via lactate metabolism. Nat. Med. 2019, 25, 1104–1109. [Google Scholar] [CrossRef]
- Plauth, M.; Bernal, W.; Dasarathy, S.; Merli, M.; Plank, L.D.; Schütz, T.; Bischoff, S. ESPEN guideline on clinical nutrition in liver disease. Clin. Nutr. 2019, 38, 485–521. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
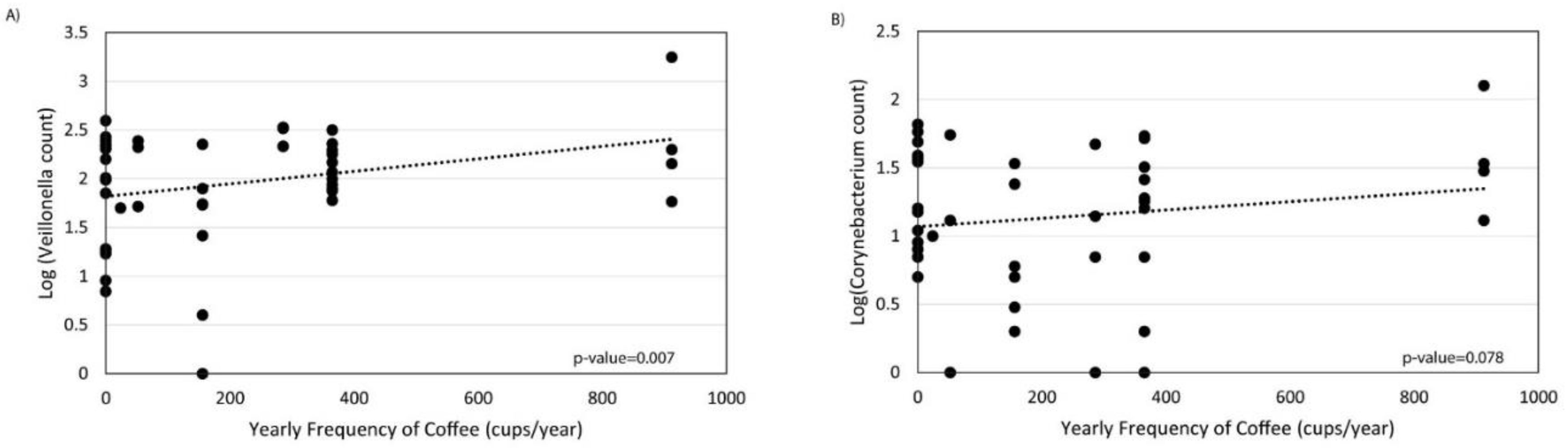{kind=link}
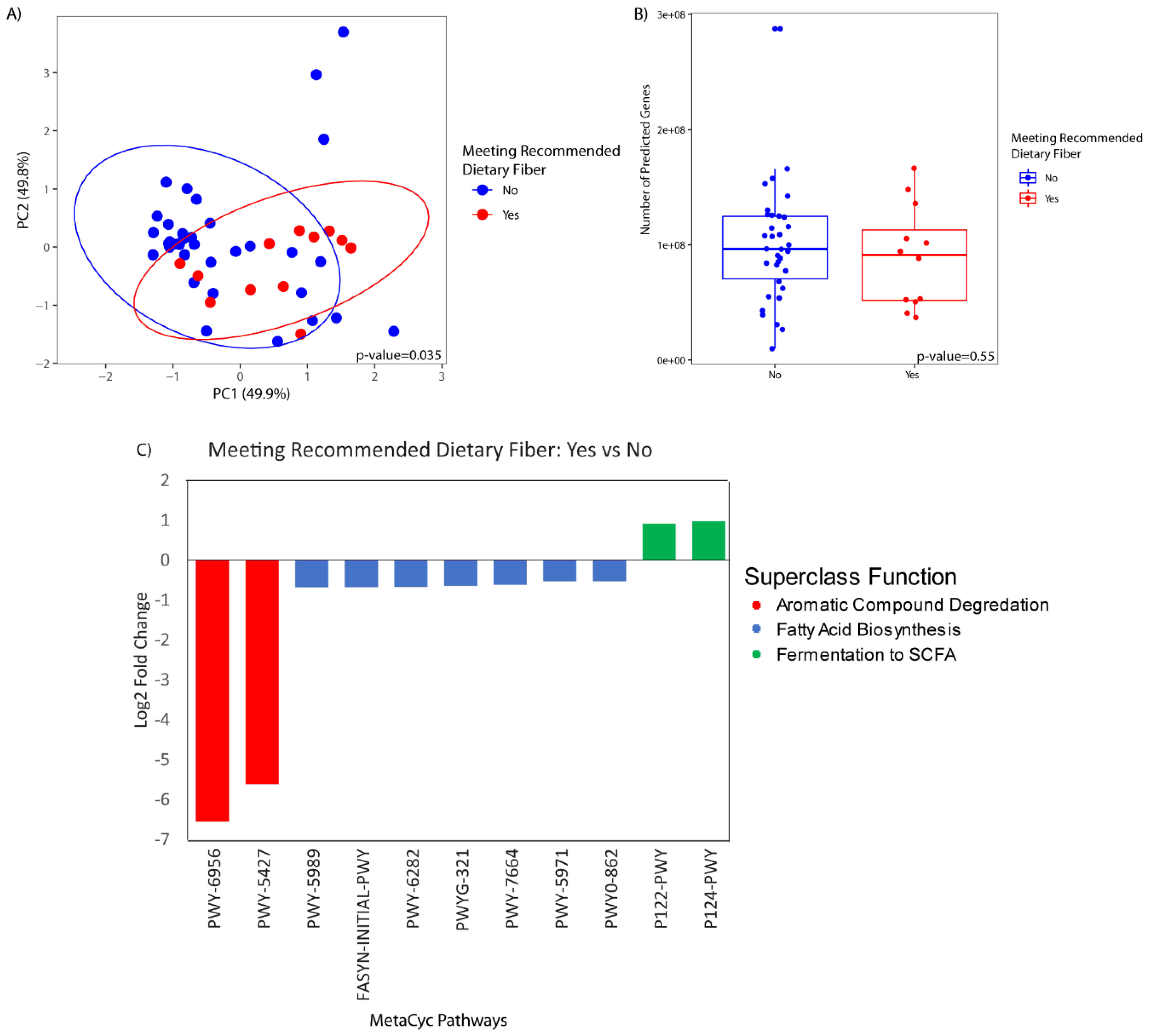{kind=link}
| Demographics | Complete | HEI-2010 | ||
|---|---|---|---|---|
| Lowest Tertile | Middle Tertile | Highest Tertile | ||
| Number | 51 | 17 | 17 | 17 |
| Site, n (%) | ||||
| CSMC | 24 (47%) | 6 (35%) | 9 (53%) | 9 (53%) |
| UCLA | 26 (51%) | 11 (65%) | 8 (47%) | 7 (41%) |
| VAGLA | 1 (2%) | 0 | 0 | 1 (6%) |
| Sex, n (%) | ||||
| Male | 29 (57%) | 11 (65%) | 10 (59%) | 8 (47%) |
| Female | 22 (43%) | 6 (35%) | 7 (41%) | 9 (53%) |
| Baseline age, mean (st dev) | 57 (11) | 52.6 (9.6) | 57.2 (11.2) | 60.5 (11.6) |
| Race, n (%) | ||||
| White | 45 (88%) | 14 (82%) | 16 (94%) | 15 (88%) |
| Non-White | 6 (12%) | 3 (18%) | 1 (6%) | 2 (12%) |
| Ethnicity, n (%) | ||||
| Hispanic or Latino | 14 (27%) | 7 (41%) | 4 (24%) | 3 (18%) |
| Not Hispanic or Latino | 37 (73%) | 10 (59%) | 13 (76%) | 14 (82%) |
| Cirrhosis etiology, n (%) | ||||
| HCV/HBV | 16 (31%) | 7 (41%) | 6 (35%) | 3 (18%) |
| ALD | 12 (24%) | 4 (24%) | 4 (24%) | 4 (24%) |
| NASH | 8 (16%) | 1 (6%) | 3 (18%) | 4 (24%) |
| PSC | 7 (14%) | 3 (18%) | 2 (12%) | 2 (12%) |
| Other | 8 (16%) | 2 (12%) | 2 (12%) | 4 (24%) |
| Cirrhosis Complications, n (%) | ||||
| Hepatic Encephalopathy | 10 (20%) | 1 (6%) | 6 (35%) | 3 (18%) |
| Esophageal Varices | 37 (73%) | 10 (59%) | 13 (76%) | 14 (82%) |
| Ascites | 27 (53%) | 9 (53%) | 10 (59%) | 8 (47%) |
| Baseline clinical labs, mean (st dev) | ||||
| AFP | 4.7 (4.3) | 4.8 (3.7) | 5.4 (6.1) | 3.8 (1.7) |
| Creatinine | 1 (0.9) | 1.3 (1.4) | 1.1 (0.45) | 0.74 (.20) |
| Bilirubin | 1.8 (2.0) | 1.8 (1.8) | 1.7 (1.5) | 1.9 (2.6) |
| AST | 42 (23) | 42 (19) | 45 (28) | 39 (21) |
| ALT | 33 (18) | 36 (18) | 32 (19) | 30 (17) |
| Platelets | 125 (78) | 125(62) | 138 (88) | 112 (86) |
| INR | 1.2 (0.2) | 1.2 (0.22) | 1.2 (0.11) | 1.3 (0.31) |
| MELD | 11.5 (5.7) | 12 (6.5) | 11 (4.7) | 11 (5.9) |
| Baseline medications, n (%) | ||||
| PPI | 23 (45%) | 6 (35%) | 10 (59%) | 7 (41%) |
| Lactulose | 9 (18%) | 1 (6%) | 3 (18%) | 5 (29%) |
| Rifaximin | 7 (14%) | 0 | 3 (18%) | 4 (24%) |
| Antibiotics | 6 (12%) | 3 (18%) | 0 | 3 (18%) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussain, S.K.; Dong, T.S.; Agopian, V.; Pisegna, J.R.; Durazo, F.A.; Enayati, P.; Sundaram, V.; Benhammou, J.N.; Noureddin, M.; Choi, G.; et al. Dietary Protein, Fiber and Coffee Are Associated with Small Intestine Microbiome Composition and Diversity in Patients with Liver Cirrhosis. Nutrients 2020, 12, 1395. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12051395
Hussain SK, Dong TS, Agopian V, Pisegna JR, Durazo FA, Enayati P, Sundaram V, Benhammou JN, Noureddin M, Choi G, et al. Dietary Protein, Fiber and Coffee Are Associated with Small Intestine Microbiome Composition and Diversity in Patients with Liver Cirrhosis. Nutrients. 2020; 12(5):1395. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12051395
Chicago/Turabian StyleHussain, Shehnaz K., Tien S. Dong, Vatche Agopian, Joseph R. Pisegna, Francisco A. Durazo, Pedram Enayati, Vinay Sundaram, Jihane N. Benhammou, Mazen Noureddin, Gina Choi, and et al. 2020. "Dietary Protein, Fiber and Coffee Are Associated with Small Intestine Microbiome Composition and Diversity in Patients with Liver Cirrhosis" Nutrients 12, no. 5: 1395. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12051395