Metformin Ameliorates Testicular Function and Spermatogenesis in Male Mice with High-Fat and High-Cholesterol Diet-Induced Obesity



Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Design
2.3. Histological Analysis
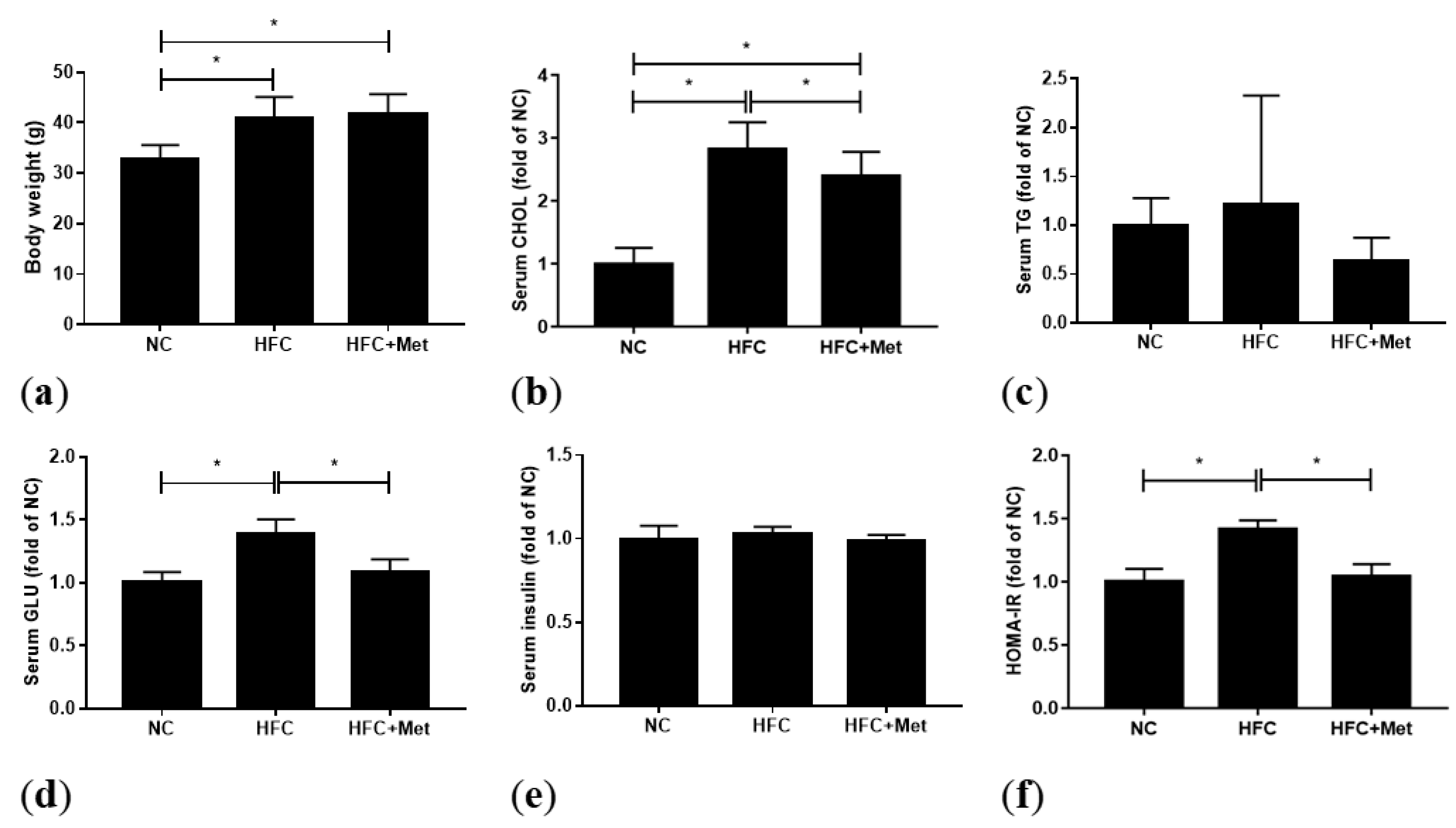
2.4. Serum Analysis
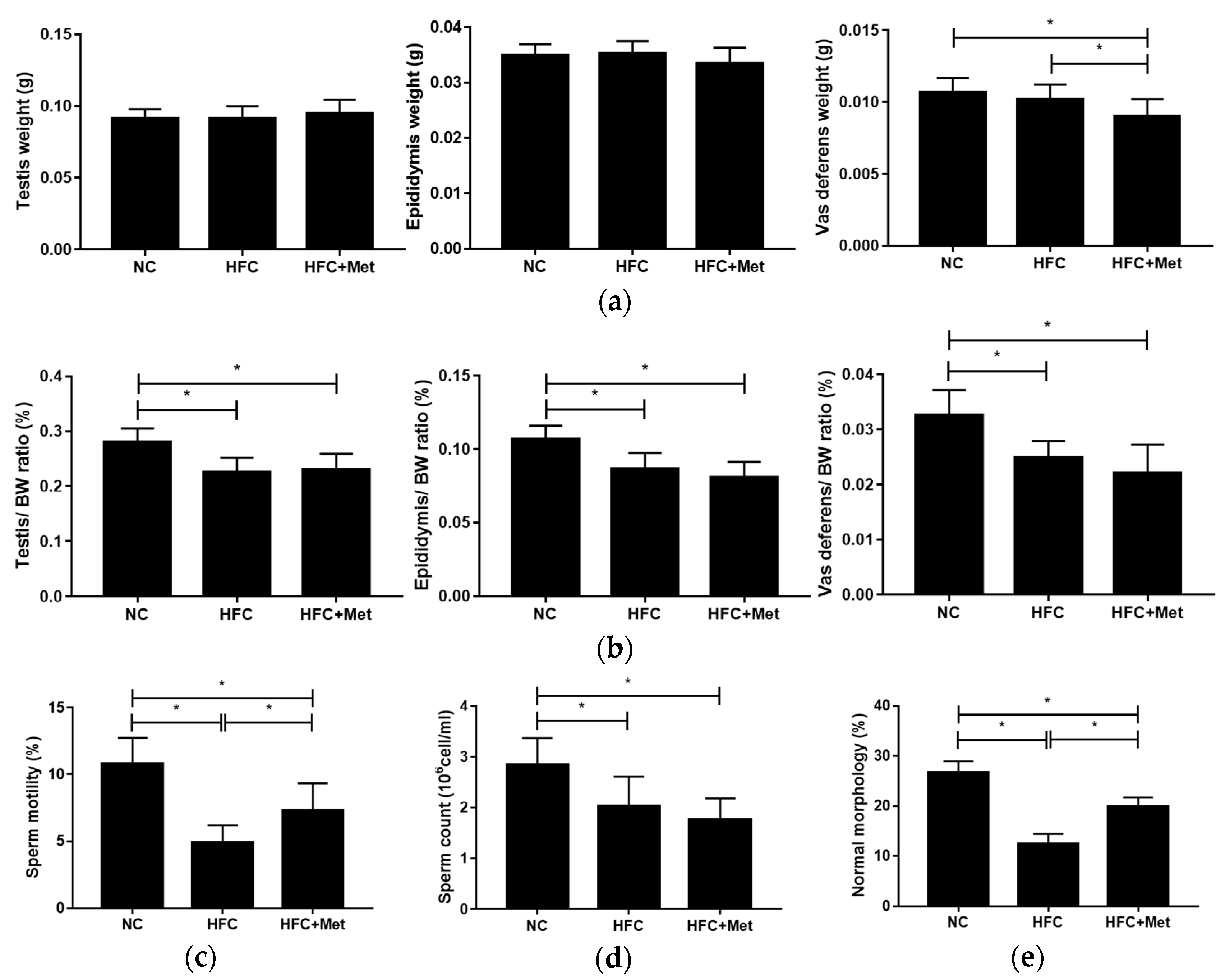
2.5. Semen Quality Analysis
2.6. Testicular Cholesterol and Sex Hormone Analysis
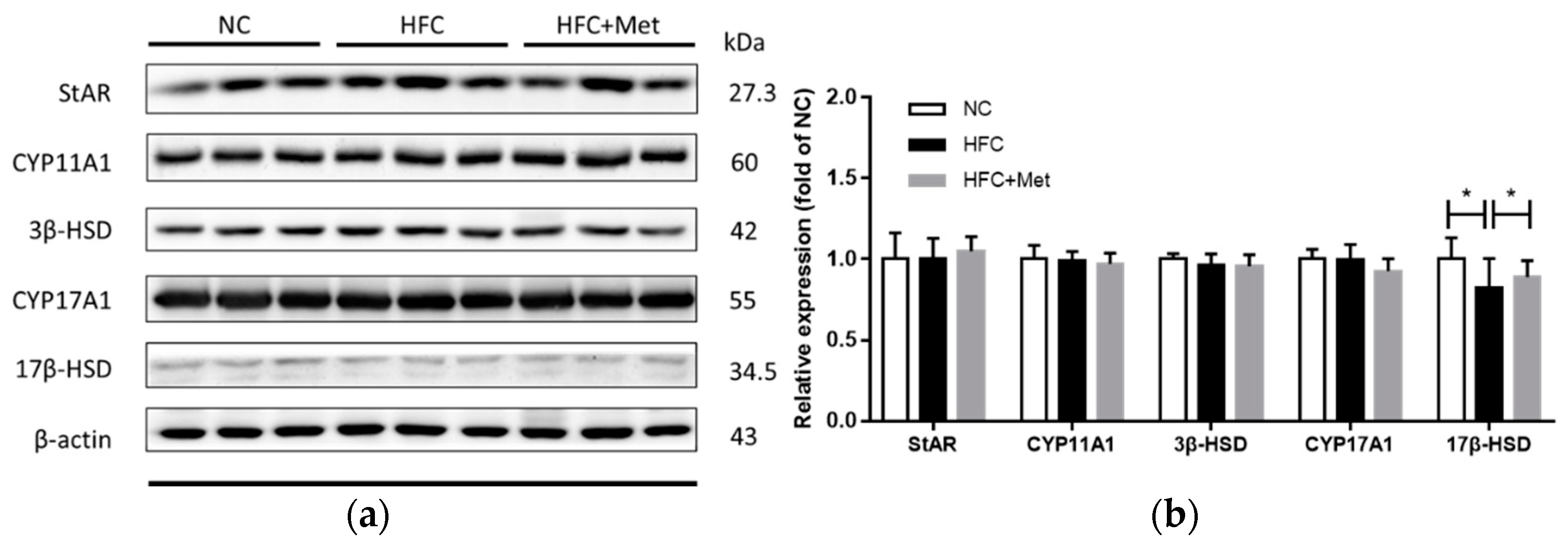
2.7. Western Blotting Analysis
2.8. Statistical Analysis
3. Results
3.1. Metformin Ameliorates the Glucose Level and HOMA-IR But Not Body Weight and Lipid Profiles
3.2. Metformin Treatment Reverses neither Liver Weight nor Liver Steatosis in Obese Mice
3.3. Metformin Treatment Improves the Testicular Spermatogenesis and Semen Quality in Obese Mice
3.4. Metformin Acts to Increase Testosterone Level in Obese Mice through Upregulating 17β-HSD Expression
3.5. Metformin Suppresses the Oxidative Stress Level in the Testis but Dose Not Reverse the Testicular Apoptosis and Inflammation
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mascarenhas, M.N.; Flaxman, S.R.; Boerma, T.; Vanderpoel, S.; Stevens, G.A. National, Regional, and Global Trends in Infertility Prevalence Since 1990: A Systematic Analysis of 277 Health Surveys. PLoS Med. 2012, 9, e1001356. [Google Scholar] [CrossRef] [PubMed]
- Poongothai, J.; Gopenath, T.S.; Manonayaki, S. Genetics of human male infertility. Singap. Med. J. 2009, 50, 336–347. [Google Scholar]
- Kumar, N.; Singh, A.K. Trends of male factor infertility, an important cause of infertility: A review of literature. J. Hum. Reprod. Sci. 2015, 8, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Mulgund, A.; Hamada, A.; Chyatte, M. A unique view on male infertility around the globe. Reprod. Boil. Endocrinol. 2015, 13, 37. [Google Scholar] [CrossRef] [Green Version]
- Levine, H.; Jørgensen, N.; Martino-Andrade, A.J.; Mendiola, J.; Weksler-Derri, D.; Mindlis, I.; Pinotti, R.; Swan, S. Temporal trends in sperm count: A systematic review and meta-regression analysis. Hum. Reprod. Update 2017, 23, 646–659. [Google Scholar] [CrossRef]
- Chavarro, J.E.; Toth, T.L.; Wright, D.L.; Meeker, J.D.; Hauser, R. Body mass index in relation to semen quality, sperm DNA integrity, and serum reproductive hormone levels among men attending an infertility clinic. Fertil. Steril. 2009, 93, 2222–2231. [Google Scholar] [CrossRef] [Green Version]
- Hofny, E.R.; Ali, M.E.; Abdel-Hafez, H.Z.; Kamal, E.E.-D.; Mohamed, E.E.; El-Azeem, H.G.A.; Mostafa, T. Semen parameters and hormonal profile in obese fertile and infertile males. Fertil. Steril. 2010, 94, 581–584. [Google Scholar] [CrossRef]
- Mulligan, T.; Frick, M.F.; Zuraw, Q.C.; Stemhagen, A.; McWhirter, C. Prevalence of hypogonadism in males aged at least 45 years: The HIM study. Int. J. Clin. Pract. 2008, 60, 762–769. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Liu, Y.; Xue, K.; Gu, G.; Fan, W.; Xu, Y.; Ding, Z. Diet-Induced Obesity in Male C57BL/6 Mice Decreases Fertility because of Disrupted Blood-Testis Barrier. PLoS ONE 2015, 10, e0120775. [Google Scholar] [CrossRef]
- Bakos, H.W.; Mitchell, M.; Setchell, B.P.; Lane, M. The effect of paternal diet-induced obesity on sperm function and fertilization in a mouse model. Int. J. Androl. 2010, 34, 402–410. [Google Scholar] [CrossRef]
- Mu, Y.; Yan, W.-J.; Yin, T.-L.; Zhang, Y.; Li, J.; Yang, J. Diet-induced obesity impairs spermatogenesis: A potential role for autophagy. Sci. Rep. 2017, 7, 43475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupont, C.; Faure, C.; Sermondade, N.; Boubaya, M.; Eustache, F.; Clement, P.; Briot, P.; Berthaut, I.; Levy, V.; Cédrin-Durnerin, I.; et al. Obesity leads to higher risk of sperm DNA damage in infertile patients. Asian J. Androl. 2013, 15, 622–625. [Google Scholar] [CrossRef] [Green Version]
- Taha, E.; Sayed, S.K.; Gaber, H.D.; Hafez, H.K.A.; Ghandour, N.; Zahran, A.; Mostafa, T. Does being overweight affect seminal variables in fertile men? Reprod. Biomed. Online 2016, 33, 703–708. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Said, T.M. Role of sperm chromatin abnormalities and DNA damage in male infertility. Hum. Reprod. Update 2003, 9, 331–345. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Said, T.M. Oxidative stress, DNA damage and apoptosis in male infertility: A clinical approach. BJU Int. 2005, 95, 503–507. [Google Scholar] [CrossRef]
- Alahmar, A.T. Role of Oxidative Stress in Male Infertility: An Updated Review. J. Hum. Reprod. Sci. 2019, 12, 4–18. [Google Scholar] [CrossRef] [PubMed]
- Azenabor, A.; Ekun, A.O.; Akinloye, O. Impact of Inflammation on Male Reproductive Tract. J. Reprod. Infertil. 2016, 16, 123–129. [Google Scholar]
- De Kretser, D.; Loveland, K.; Meinhardt, A.; Simorangkir, D.; Wreford, N. Spermatogenesis. Hum. Reprod. 1998, 13, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bain, J. The many faces of testosterone. Clin. Interv. Aging 2007, 2, 567–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyagi, V.; Scordo, M.; Yoon, R.S.; Liporace, F.A.; Greene, L.W. Revisiting the role of testosterone: Are we missing something? Rev. Urol. 2017, 19, 16–24. [Google Scholar]
- Hanukoglu, I. Steroidogenic enzymes: Structure, function, and role in regulation of steroid hormone biosynthesis. J. Steroid Biochem. Mol. Boil. 1992, 43, 779–804. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Torres, M.A.; Carrera, A.; Zambrana, M. High incidence of hyperestrogenemia and dyslipidemia in a group of infertile men. Ginecol. Obstet. 2000, 68, 224–229. [Google Scholar]
- Maqdasy, S.; Baptissart, M.; Vega, A.; Baron, S.; Lobaccaro, J.-M.A.; Volle, D.H. Cholesterol and male fertility: What about orphans and adopted? Mol. Cell. Endocrinol. 2013, 368, 30–46. [Google Scholar] [CrossRef]
- Wang, Y.-W.; He, S.-J.; Feng, X.; Cheng, J.; Luo, Y.-T.; Tian, L.; Huang, Q. Metformin: A review of its potential indications. Drug Des. Dev. Ther. 2017, 11, 2421–2429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attia, S.M.; Helal, G.; Alhaider, A. Assessment of genomic instability in normal and diabetic rats treated with metformin. Chem. Interact. 2009, 180, 296–304. [Google Scholar] [CrossRef]
- Morgante, G.; Tosti, C.; Orvieto, R.; Musacchio, M.C.; Piomboni, P.; de Leo, V. Metformin improves semen characteristics of oligo-terato-asthenozoospermic men with metabolic syndrome. Fertil. Steril. 2011, 95, 2150–2152. [Google Scholar] [CrossRef]
- Johnsen, S.G. Testicular Biopsy Score Count—A Method for Registration of Spermatogenesis in Human Testes: Normal Values and Results in 335 Hypogonadal Males. Horm. Res. Paediatr. 1970, 1, 2–25. [Google Scholar] [CrossRef]
- Pacini, G.; Mari, A. Methods for clinical assessment of insulin sensitivity and β-cell function. Best Pract. Res. Clin. Endocrinol. Metab. 2003, 17, 305–322. [Google Scholar] [CrossRef]
- Woo, S.-L.; Xu, H.; Li, H.; Zhao, Y.; Hu, X.; Zhao, J.; Guo, X.; Guo, T.; Botchlett, R.; Qi, T.; et al. Metformin Ameliorates Hepatic Steatosis and Inflammation without Altering Adipose Phenotype in Diet-Induced Obesity. PLoS ONE 2014, 9, e91111. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Sharma, A.; Yin, C.; Tan, X.; Xiao, Y. Metformin ameliorates hepatic steatosis and improves the induction of autophagy in HFD-induced obese mice. Mol. Med. Rep. 2017, 16, 680–686. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.-J.; Dai, W.; Scott, M.J.; Li, R.; Zhang, Y.-Q.; Yang, Y.; Chen, L.-Z.; Huang, X.-S. Metformin improves nonalcoholic fatty liver disease in obese mice via down-regulation of apolipoprotein A5 as part of the AMPK/LXRα signaling pathway. Oncotarget 2017, 8, 108802–108809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seifarth, C.; Schehler, B.; Schneider, H.J. Effectiveness of Metformin on Weight Loss in Non-Diabetic Individuals with Obesity. Exp. Clin. Endocrinol. Diabetes 2012, 121, 27–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levri, K.M.; Slaymaker, E.; Last, A.; Yeh, J.; Ference, J.; D’Amico, F.; Wilson, S.A. Metformin as Treatment for Overweight and Obese Adults: A Systematic Review. Ann. Fam. Med. 2005, 3, 457–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinstein, M.M.; Tu, Y.; Beigneux, A.P.; Davies, B.S.; Gin, P.; Voss, C.; Walzem, R.L.; Reue, K.; Tontonoz, P.; Bensadoun, A.; et al. Cholesterol intake modulates plasma triglyceride levels in glycosylphosphatidylinositol HDL-binding protein 1-deficient mice. Arter. Thromb. Vasc. Boil. 2010, 30, 2106–2113. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Zhou, Y.; Liu, Y.X.; Ping, J.; Shou, Q.Y.; Chen, F.M.; Ruo, R. Metformin improves hepatic IRS2/PI3K/Akt signaling in insulin-resistant rats of NASH and cirrhosis. J. Endocrinol. 2016, 229, 133–144. [Google Scholar] [CrossRef] [Green Version]
- Kita, Y.; Takamura, T.; Misu, H.; Ota, T.; Kurita, S.; Takeshita, Y.; Uno, M.; Nagata, N.; Kato, K.-I.; Ando, H.; et al. Metformin Prevents and Reverses Inflammation in a Non-Diabetic Mouse Model of Nonalcoholic Steatohepatitis. PLoS ONE 2012, 7, e43056. [Google Scholar] [CrossRef]
- Haukeland, J.W.; Konopski, Z.; Eggesbø, H.B.; von Volkmann, H.L.; Raschpichler, G.; Bjøro, K.; Haaland, T.; Løberg, E.M.; Birkeland, K. Metformin in patients with non-alcoholic fatty liver disease: A randomized, controlled trial. Scand. J. Gastroenterol. 2009, 44, 853–860. [Google Scholar] [CrossRef]
- Li, Y.; Liu, L.; Wang, B.; Wang, J.; Chen, N. Metformin in non-alcoholic fatty liver disease: A systematic review and meta-analysis. Biomed. Rep. 2012, 1, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Farah, S.; Nguyen, T.; Kelsberg, G.; Safranek, S. Metformin for Nonalcoholic Fatty Liver Disease and Nonalcoholic Steatohepatitis. Am. Fam. Phys. 2019, 99, 262–263. [Google Scholar]
- Chalasani, N.; Younossi, Z.M.; LaVine, J.E.; Charlton, M.; Cusi, K.; Rinella, M.; Harrison, S.A.; Brunt, E.M.; Sanyal, A.J. The diagnosis and management of nonalcoholic fatty liver disease: Practice guidance from the American Association for the Study of Liver Diseases. Hepatology 2017, 67, 328–357. [Google Scholar] [CrossRef]
- Sui, Y.; Kong, X.; Fan, R.; Ye, Y.; Mai, H.; Zhuo, S.; Lu, W.; Ruan, P.; Fang, S.; Yang, T. Long-term treatment with metformin in the prevention of fatty liver in Zucker diabetic fatty rats. Diabetol. Metab. Syndr. 2019, 11, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, J.; Luo, D.; Xu, X.; Sun, M.; Su, X.; Tian, Z.; Zhang, M.; Yu, C.; Guan, Q. Metformin Improves Fertility in Obese Males by Alleviating Oxidative Stress-Induced Blood-Testis Barrier Damage. Oxid. Med. Cell. Longev. 2019, 9151067-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, W.-J.; Mu, Y.; Yu, N.; Yi, T.-L.; Zhang, Y.; Pang, X.-L.; Cheng, D.; Yang, J. Protective effects of metformin on reproductive function in obese male rats induced by high-fat diet. J. Assist. Reprod. Genet. 2015, 32, 1097–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosman, E.; Esterhuizen, A.D.; Rodrigues, F.A.; Becker, P.J.; Hoffmann, W.A. Effect of metformin therapy and dietary supplements on semen parameters in hyperinsulinaemic males. Andrologia 2014, 47, 974–979. [Google Scholar] [CrossRef] [Green Version]
- McLachlan, R.I.; O’Donnell, L.; Meachem, S.J.; Stanton, P.G.; de Kretser, D.M.; Pratis, K.; Robertson, D.M. Hormonal regulation of spermatogenesis in primates and man: Insights for development of the male hormonal contraceptive. J. Androl. 2002, 23, 149–162. [Google Scholar] [PubMed]
- Morgan, D.H.; Ghribi, O.; Hui, L.; Geiger, J.D.; Chen, X. Cholesterol-enriched diet disrupts the blood-testis barrier in rabbits. Am. J. Physiol. Metab. 2014, 307, E1125–E1130. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.; Jiang, F.; Zhang, M.; Luo, D.; Shao, S.; Zhao, J.; Gao, L.; Zuo, C.; Guan, Q. HC diet inhibited testosterone synthesis by activating endoplasmic reticulum stress in testicular Leydig cells. J. Cell. Mol. Med. 2019, 23, 3140–3150. [Google Scholar] [CrossRef]
- Fejes, I.; Koloszár, S.; Závaczki, Z.; Daru, J.; Szöllösi, J.; Pal, A. Effect of body weight on testosterone/estradiol ratio in oligozoospermic patients. Arch. Androl. 2006, 52, 97–102. [Google Scholar] [CrossRef] [Green Version]
- Zumoff, B. Hormonal Abnormalities in Obesity. Acta Med. Scand. 2009, 222, 153–160. [Google Scholar] [CrossRef]
- Rice, S.; Pellatt, L.; Ramanathan, K.; Whitehead, S.A.; Mason, H.D. Metformin inhibits aromatase via an extracellular signal-regulated kinase-mediated pathway. Endocrinology 2009, 150, 4794–4801. [Google Scholar] [CrossRef] [Green Version]
- Fuhrmeister, I.P.; Branchini, G.; Pimentel, A.M.; Ferreira, G.D.; Capp, E.; Brum, I.; Corleta, H.V.E. Human Granulosa Cells: Insulin and Insulin-Like Growth Factor-1 Receptors and Aromatase Expression Modulation by Metformin. Gynecol. Obstet. Investig. 2014, 77, 156–162. [Google Scholar] [CrossRef]
- Faure, M.; Bertoldo, M.J.; Khoueiry, R.; Bongrani, A.; Brion, F.; Giulivi, C.; Dupont, J.; Froment, P. Metformin in Reproductive Biology. Front. Endocrinol. 2018, 9, 675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertoldo, M.; Guibert, E.; Faure, M.; Guillou, F.; Rame, C.; Nadal-Desbarats, L.; Foretz, M.; Viollet, B.; Dupont, J.; Froment, P. Specific deletion of AMP-activated protein kinase (α1AMPK) in mouse Sertoli cells modifies germ cell quality. Mol. Cell. Endocrinol. 2016, 423, 96–112. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.G.; Martins, A.D.; Vaz, C.V.; Correia, S.; Moreira, P.; Oliveira, P.F.; Socorro, S. Metformin, and male reproduction: Effects on Sertoli cell metabolism. Br. J. Pharmacol. 2014, 171, 1033–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, X.; Xu, Q.-Y.; Jia, C.; Peng, Y.-F. Metformin improves epididymal sperm quality and antioxidant function of the testis in diet-induced obesity rats. Zhonghua Nan Ke Xue Natl. J. Androl. 2012, 18, 146–149. [Google Scholar]
- Min, Q.-Q.; Qin, L.; Sun, Z.-Z.; Zuo, W.-T.; Zhao, L.; Xu, J.-Y. Effects of Metformin Combined with Lactoferrin on Lipid Accumulation and Metabolism in Mice Fed with High-Fat Diet. Nutrients 2018, 10, 1628. [Google Scholar] [CrossRef] [Green Version]
- Asghari, A.; Akbari, G.; Meghdadi, A.; Mortazavi, P. Protective effect of metformin on testicular ischemia/reperfusion injury in rats. Acta Cir. Bras. 2016, 31, 411–416. [Google Scholar] [CrossRef] [Green Version]
- Nna, V.U.; Abu Bakar, A.B.; Azlina, A.; Mohamed, M. Diabetes-induced testicular oxidative stress, inflammation, and caspase-dependent apoptosis: The protective role of metformin. Arch. Physiol. Biochem. 2018, 1–12. [Google Scholar] [CrossRef]
- Pourmatroud, E. Metformin in Health Issues and Reproductive System. In Metformin; IntechOpen: Rijeka, Croatia, 2019. [Google Scholar] [CrossRef] [Green Version]
- Theas, S.; Rival, C.; Jarazo-Dietrich, S.; Jacobo, P.; Guazzone, V.; Lustig, L. Tumour necrosis factor-released by testicular macrophages induces apoptosis of germ cells in autoimmune orchitis. Hum. Reprod. 2008, 23, 1865–1872. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | NC | HFC | HFC + Met |
| Corn starch | 41.0 | 29.5 | 29.5 |
| Dextrin | 15.5 | 10.0 | 10.0 |
| Sucrose | 10.0 | 10.0 | 10.0 |
| Cellulose | 5.0 | 5.0 | 5.0 |
| Casein | 19.0 | 19.0 | 19.0 |
| Soybean oil | 4.0 | 20.0 | 20.0 |
| Mineral mix | 3.5 | 3.5 | 3.5 |
| Vitamin mix | 1.0 | 1.0 | 1.0 |
| Choline | 0.25 | 0.25 | 0.25 |
| Cholesterol | - | 1.5 | 1.5 |
| Cysteine | 0.18 | 0.18 | 0.18 |
| TBHQ | 0.25 | 0.0008 | 0.0008 |
| Metformin | - | - | 0.05 |
| Energy (%) | NC | HFC | HFC + Met |
| Carbohydrate | 70 | 43 | 43 |
| Fat | 10 | 40 | 40 |
| Protein | 20 | 17 | 17 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.-Y.; Chang, T.-C.; Lin, S.-H.; Wu, S.-T.; Cha, T.-L.; Tsao, C.-W. Metformin Ameliorates Testicular Function and Spermatogenesis in Male Mice with High-Fat and High-Cholesterol Diet-Induced Obesity. Nutrients 2020, 12, 1932. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12071932
Liu C-Y, Chang T-C, Lin S-H, Wu S-T, Cha T-L, Tsao C-W. Metformin Ameliorates Testicular Function and Spermatogenesis in Male Mice with High-Fat and High-Cholesterol Diet-Induced Obesity. Nutrients. 2020; 12(7):1932. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12071932
Chicago/Turabian StyleLiu, Chin-Yu, Ting-Chia Chang, Shyh-Hsiang Lin, Sheng-Tang Wu, Tai-Lung Cha, and Chih-Wei Tsao. 2020. "Metformin Ameliorates Testicular Function and Spermatogenesis in Male Mice with High-Fat and High-Cholesterol Diet-Induced Obesity" Nutrients 12, no. 7: 1932. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12071932