Mitochondrial Dysfunction as a Novel Target for Neuroprotective Nutraceuticals in Ocular Diseases


1
School of Chinese Medicine, College of Chinese Medicine, China Medical University, Taichung 404, Taiwan
2
Graduate Institute of Acupuncture Science, College of Chinese Medicine, China Medical University, Taichung 404, Taiwan
3
Department of Medical Research, China Medical University Hospital, Taichung 404, Taiwan
4
Children’s Hospital of China Medical University, Taichung 404, Taiwan
5
Department of Medical Genetics, China Medical University Hospital, Taichung 404, Taiwan
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Nutrients 2020, 12(7), 1950; https://0-doi-org.brum.beds.ac.uk/10.3390/nu12071950
Submission received: 4 June 2020
/
Revised: 23 June 2020
/
Accepted: 29 June 2020
/
Published: 30 June 2020
(This article belongs to the Special Issue Nutrition for Eye Health)
Abstract
:The eyes require a rich oxygen and nutrient supply; hence, the high-energy demand of the visual system makes it sensitive to oxidative stress. Excessive free radicals result in mitochondrial dysfunction and lead to retinal neurodegeneration, as an early stage of retinal metabolic disorders. Retinal cells are vulnerable because of their coordinated interaction and intricate neural networks. Nutraceuticals are believed to target multiple pathways and have shown neuroprotective benefits by scavenging free radicals and promoting mitochondrial gene expression. Furthermore, encouraging results demonstrate that nutraceuticals improve the organization of retinal cells and visual functions. This review discusses the mitochondrial impairments of retinal cells and the mechanisms underlying the neuroprotective effects of nutraceuticals. However, some unsolved problems still exist between laboratory study and clinical therapy. Poor bioavailability and bioaccessibility strongly limit their development. A new delivery system and improved formulation may offer promise for health care applications.
1. Introduction
Retinal neurodegeneration is one of the major causes of visual impairment and is highly associated with atrophy or cell death of the retina in ocular diseases, such as glaucoma, age-related macular degeneration (AMD), and diabetic retinopathy (DR) [1]. The metabolic rate of the visual system is higher than that of others within the brain, and further disruptions in the metabolic homeostasis can lead to vulnerability of the retina [2] In this review, we endeavored to focus on the neuropathies of retinal cell types induced by metabolic impairment, as well as the potential neuroprotective nutraceuticals based on evidence from animal experiments and clinical studies.
1.1. The High-Energy Demands of the Retina
The process of visualizing information is achieved by transmitting light into electrical impulses, triggered mainly by rods and cones, through the optic nerve to the brain. Bipolar cells and amacrine cells collect the signals from photoreceptor cells and synapse with retinal ganglion cells (RGCs). RGCs are a type of neuron and propagate action potentials (AP) to axons. The axons form the optic nerve and project to the lateral geniculate nucleus (LGN), the medial geniculate body, and superior colliculus. There are five neuronal cell types: photoreceptor, horizontal, bipolar, amacrine, and ganglion cells in the retina [3]. In the visual system, phototransduction, neurotransmitter utilization, protein synthesis transport, and repolarization-after-depolarization are energy-dependent [2]. The amount of oxygen and nutrient consumption in the retina is vast because of the intricate neural networks. Mitochondria are the major endogenous sources of adenosine triphosphate (ATP) and reactive oxygen species (ROS) in mammalian cells. Mitochondria are dynamic organelles and mitochondrial integrity is maintained by fusion, fission, mitophagy, and biogenesis. Fusion stimulates the formation of elongated mitochondria to produce more ATP. Excessive accumulation of ROS causes fission and triggers mitochondrial fragmentation. Damaged mitochondria are further self-destructed for turnover by the mitophagy pathway and then biogenesis replenishes number and/or mass of de novo mitochondria [4,5,6]. Energy metabolism overexertion causes an imbalance between the generation and elimination of ROS. Furthermore, excess ROS causes oxidative stress, which leads to retinal neurodegeneration, visual loss, and eventual blindness [7].
1.2. Disruptions of the Metabolic Homeostasis Cause Mitochondrial Dysfunction in the Retina
Under normal physiological conditions, mitochondrial ROS (mtROS) is quickly scavenged by manganese superoxide dismutase (SOD2), maintaining metabolic homeostasis. Overproduction of mtROS damages the cells by increasing levels of oxygen free radicals and further causes lipid peroxidation, protein carbonylation, and DNA breakage [8], especially mitochondrial DNA (mtDNA) without protection by histones and repair mechanisms [9]. A burst of ROS is a primary event in glutamate-induced neurotoxicity [10], and vice versa in optic atrophy gene 1 (OPA1) mutation animal models [11]. Furthermore, activation of ionotropic glutamate receptors raises Ca2+ entry into both the cytosol and mitochondria, and induces the process of neuronal death by apoptosis or necrosis [12,13]. The increasing number of free radicals also attack lipoproteins and form intracellular lipid deposits to alter autophagy or phagocytosis in the retina [14]. The term ‘mitochondrial optic neuropathy’ (MON) suggested the need to investigate the pathogenetic role of the mitochondria. There are three major acquired etiologies, including toxic, nutritional, and metabolic insults which cause MON [15]. Moreover, chronic and metabolic disorders, such as dyslipidemia or hyperglycemia, disturb the homeostasis of the mitochondrial dynamic and develop neurodegeneration as an early event in the pathogenesis of glaucoma [16], AMD [14] and DR [6,17].
1.3. The Characteristics of Mitochondrial Dysfunction in Retinal Neurodegeneration
Glucose and lipid metabolism are mostly dependent on the mitochondria, therefore, mitochondrial abnormality is seen in metabolic syndromes [18]. Mitochondrial oxidative stress alters the delicate balance between fusion and fission, mitochondrial fragmentation, and further increases the number of autophagosomes and accumulation of dysfunctional mitochondria [19,20,21]. In parallel, it accompanies the cytosolic increase in the activity of the polyol pathway, advanced glycation end products (AGEs), and protein kinase C (PKC) activation [22,23]. The susceptibility to oxidative stress is closely associated with the distribution and amount of mitochondria in RGCs [17,24]. The anatomical structure of RGCs is another important factor which causes optic neuropathy. The anterior part of the axon is unmyelinated and has slower conduction velocities. That means more energy consumption is necessary to maintain the generation and propagation of AP. These neuropathic phenotypes of RGCs include loss of dendritic arborization, axonal loss, optic nerve atrophy, and retinal nerve fiber layer thinning to cause clinical signs of reduction in visual evoked potential (VEP) and optic nerve degeneration [17].
In the retina, mitochondria are also found to be abundant at the distal ends of the retinal pigment epithelium (RPE) and photoreceptors, especially the basement membrane, cilium, and outer feet [25]. Mitochondrial dysfunction of a coordinated ecosystem between RPE and photoreceptors has recently been described. Photoreceptors utilize glucose by glycolysis and convert it to lactate, which transports back to the RPE cells as a fuel through oxidative phosphorylation [26]. Conditional knockout of Sod2 in the RPE elevates levels of oxidative stress and dysfunctional mitochondria in both RPE and photoreceptors [27]. In addition, the metabolism of photoreceptors becomes maximal to generate dark current at night [28]. Therefore, photoreceptors are the major site of ROS generation and show apoptosis in the early stages of diabetes [29] and atrophy in the later stages of AMD [17]. In brief, the high energy requirement and electrophysiological function of retinal cells make them more vulnerable to oxidative stress.
1.4. Nutraceuticals as Neuroprotectants for Retinal Neurodegeneration
Metabolic impairments cause numerous retinal manifestations in chronic progression. This results in irreversible damage of retinal cells if oxidative stress is not reduced with treatment. Some clinical medications or surgeries are approved to treat ocular diseases. However, these therapies are used for the late stages of disease progression. There is a clear need for new strategies to act at the molecular or cellular target to prevent the development of the disorder. Mitochondrial dysfunction has been shown to be one of the early events in retinal neurodegeneration [30,31]. Targeting the mitochondrial function brings the promise of new options. Nutraceuticals are believed to target multiple pathways and attenuate the progression of neuronal destruction through mitochondrial dysfunction [32,33]. Therefore, nutraceuticals could be considered as positive neuroprotectants of retinal cells. Here, we summarize the retinopathy progression related to mitochondrial dysfunction and the effects of nutraceuticals with improved retinal neurodegeneration as below.
2. Mitochondrial Dysfunction in the Pathogenesis of Ocular Diseases
2.1. Glaucoma
Glaucoma is the second leading cause of blindness worldwide and is characterized by the progressive degeneration or loss of RGCs [34]. Most glaucoma cases are classified into primary open-angle glaucoma (POAG) and angle-closure glaucoma (ACG) caused by an increase in the intraocular pressure (IOP). Mitochondrial dysfunction in the trabecular meshwork may impair their cytoarchitecture and lead to alteration in the drainage of aqueous humor, further raising the IOP [16,35]. IOP–induced stress and strain are biomechanical factors of damage in the lamina cribrosa and adjacent tissues [34]. It also induces metabolic stress causing mitochondrial dysfunction in mouse RGCs [36]. In some individuals with normal range IOP, particularly in Asians, they are classified as normal-tension glaucoma (NTG) [37]. The pathogenic mechanism of NTG is not fully understood; the low pressure of cerebrospinal fluid in the optic nerve subarachnoid space may cause trans-lamina cribrosa pressure difference and compress the optic nerve [38]. Numerous biomarkers related to oxidative stress are reported to be significantly higher in glaucoma patients [39]. Therefore, recent scientific literature demonstrates that mitochondrial dysfunction and oxidative stress are both a cause and consequence and play a central role in the process of glaucoma [40].
2.2. Age-Related Macular Degeneration
AMD is a progressive ocular disease with loss of central vision and is a major cause of visual impairment in the developed world. It is clinically classified as early-stage (formation of drusen deposits between the Bruch’s membrane and RPE) and late-stage AMD owing to atrophy of the RPE/photoreceptors (dry) or choroidal neovascularization (wet) [41]. Complement factor H (CFH) is an important component of drusen, indicating a local complement-activation at the RPE [42]. Oxidative stress suppresses the expression of CFH [43] and promotes complement system activation [44], abolishing its protective function from the lipid peroxidation product [45]. Smoking is believed to be the strongest risk for developing AMD [46] and leads to oxidative stress and complement activation, resulting in the endoplasmic reticulum (ER) stress-mediated lipid accumulation [47]. Initial RPE mitochondrial abnormalities have been revealed in AMD patients [48]. Furthermore, mtDNA damage is found in the macular and peripheral RPE of AMD human samples [49]. Therefore, AMD could be seen as a progressive neurodegenerative disease primarily causing damage to mtDNA and further affecting the mitochondrial function of RPE [50].
2.3. Diabetic Retinopathy
DR is one of the most common complications of diabetes and remains the leading cause of vision loss among working-age adults in developed countries [51]. It is traditionally characterized as a microvascular disease [52] and has recently been recognized as a disruption of the interdependence between multiple retinal cell-types, causing neurodegeneration at the endpoint [53,54]. Hyperglycemia and poor glucose control are fundamental in the development of DR [55,56]. RGCs, amacrine cells, and photoreceptors have an increased apoptotic rate at the early stages of DR development in humans [57,58,59]. This apoptotic death causes pericentral macular thinning of both the inner retinal layers and the nerve fiber layer (NFL), otherwise, hypertrophy (swelling) of Müller cells increases the thickness of the inner nuclear layer (INL) [60,61,62]. The detailed mechanisms of apoptosis in the development of DR are still not clear [63]. There are multiple factors involved in the pathogenesis, including AGEs, free radicals, excitotoxicity, and mitochondrial damage as mentioned above. Finally, hyperglycemia-induced metabolic stress may initiate a vicious cycle to amplify mitochondrial dysfunction, and further accelerate the apoptosis of retinal cells [64,65]. Severe loss of retinal cells causes failures in orchestrating intimate communication and promotes compensatory over-angiogenesis in proliferative DR [66,67] (Figure 1).
3. Neuroprotective Effects of Nutraceuticals in Animal Studies and Clinical Trails
3.1. Resveratrol
Resveratrol is a plant polyphenol found in grapes and red wine [68], and is reported to improve mitochondrial function by activating SIRT1 and peroxisome proliferator-activated receptor-γ coactivator 1α (PGC-1α) [69]. It is also effective for age-related ocular diseases through anti-oxidant and anti-inflammatory properties [70]. Evidence demonstrates that resveratrol raises the survival of RGCs [71] and protects the loss of dendrite complexity [72] in glaucoma experimental models. Reports have indicated that resveratrol causes a reduction in IOP in steroid-induced ocular hypertension rats [73] and normal normotensive rabbits [74]. Resveratrol has neuroprotective effects through suppressing apoptosis-related molecule activator protein 1 (AP-1) [75] and elevating neuroprotective factors such as leukemia inhibitory factor (LIF), brain-derived neurotrophic factor (BDNF), oncostatin M (OSM), cardiotrophin 1 (CT-1), and cardiotrophin-like cytokine (CLC) [76] in light-induced retinal degeneration mouse models. It has also demonstrated benefits in alleviating hyperglycemia, oxidative biomarkers, vascular damage, anti-inflammation, anti-apoptosis, and reduction in the thickness of retinal layers in DR animal models. The proposed mechanisms include reduction in nuclear factor κB (NF-κB), paraoxonase 1 (PON1), and vascular endothelial growth factor (VEGF) [77,78,79,80].
Resveratrol has numerous beneficial effects on anti-cancer, cardiovascular diseases, obesity, diabetes, and neurological disorders in humans. It is reported to be safe at a dose of 1 g or more per day, however, the major obstacle for clinical therapy is the rapid metabolism and poor bioavailability [81,82,83]. Until now, research on resveratrol has been limited to animal models and in vitro experiments in ocular diseases [70,84]. Case report observations have shown resveratrol based nutritional supplements have benefits to improve RPE functions in AMD patients [85]. In addition, a recent double-blind randomized control trial indicates resveratrol notably reduces muscle fat and improves mitochondrial function in diabetes type 2 (T2D) patients [86]. Further investigations into the retino-protective effects of resveratrol should include more clinical studies.
3.2. Quercetin
Quercetin is a dietary flavonoid compound found in fruits, vegetables and beverages [87]. It has a substantial antioxidant ability to scavenge ROS [88] and ameliorates mitochondrial dysfunction through an AMP-activated protein kinase (AMPK)/SIRT1 signaling pathway [89,90]. An increasing number of studies show that quercetin reduces ROS [91,92], mitochondrial membrane potential (ΔΨm) and has anti-apoptotic effects on RGCs [93]. Zhou and colleagues recently report quercetin alleviates the excitability of RGCs through increased miniature GABAergic neurotransmission and decreasing miniature glutamatergic neurotransmission [94]. On the other hand, some studies have demonstrated that quercetin has an inhibitory effect of heat shock protein 72 (HSP 72) in RGCs [95,96,97]. Quercetin has neuroprotective effects of retinal layers [98] and cytoprotective effects of photoreceptor, RPE and RGCs through inhibiting activity of AP-1 pathway [99] in light-induced retinal degeneration rodent models. It attenuates hyperglycemia [100] and dyslipidemia [101], and also has anti-retinal oxidative stress, anti-neuroinflammation and anti-apoptosis protective effects in diabetic animal models [102].
Clinical trials on quercetin have shown multiple effects, such as anti-inflammatory effects through the reduction in plasma C-reactive protein [103] or oxidative stress markers [104], anti-cancer effects, and cancer chemoprevention [105,106,107]. In a recent cohort study with a 15-year follow-up, dietary intake of quercetin was shown to reduce the prevalence of any AMD (OR: 0.76; 95% CI: 0.58, 0.99) [108]. However, the lack of clinical data limits its application in ocular diseases; thus, more clinical studies are required in the future.
3.3. Xanthophylls (Lutein and Zeaxanthin)
Lutein and zeaxanthin stereoisomer are oxygenated carotenoids (xanthophylls) and are present at the macula as macular pigments [109]. Xanthophylls cannot be synthesized in humans and their supplements depend on dietary sources. They are abundant in various foods such as spinach, egg yolk, and wolfberry [110]. Xanthophylls play a key role in ROS scavenging and have anti-inflammatory and neuroprotective functions [110,111,112]. They are cleaved by β,β-carotene 9′,10′-oxygenase 2 (BCO2), however, inactivity of human BCO2 causes carotenoid accumulation [113]. This phenomenon may be an important mechanism for protecting the macula from short-wavelength light-induced damage [114]. Lutein has multiple benefits via anti-apoptosis [115], antioxidant [116] and reducing ER stress [117] in the retina. Recent studies demonstrate that xanthophylls could upregulate carotenoid metabolic genes and also improve mitochondrial biogenesis in primate animal models [118,119].
Many studies show multi-ingredient formulations for individuals could increase the concentrations of lutein or xanthophylls in the plasma and macular pigment (reviewed by Bernstein et al. [120]). Epidemiologic studies support lutein for the prevention of developing AMD in the early or intermediate stage [121,122]. It is also reported that lutein/zeaxanthin may be protective against late AMD [123]. A recent systematic review reported that there are at least 47 publications from 1946 to October 2016 and its conclusions show a strong relationship between lutein/zeaxanthin supplementation and evaluation of both macular pigment density and visual function [124]. There are controversial results of lutein and zeaxanthin in an Age-Related Eye Disease Study 2 (AREDS2) [125]. Their data showed that AREDS2 formulation in primary analyses did not prevent process of advanced AMD. However, when participants were limited to those with the lowest dietary intake of lutein + zeaxanthin, results of exploratory subgroup analyses showed a protective effect for progression to advanced AMD (HR: 0.74; 95% CI, 0.59–0.94; p = 0.01). These inconsistent results of xanthophylls in clinical trials need further design approaches to confirm their benefits.
3.4. Omega-3 Fatty Acid
Docosahexaenoic acid (DHA, C22:6) and eicosapentaenoic acid (EPA, C20:5), belonging to omega-3 (α-linolenic acid, n-3) polyunsaturated fatty acids (PUFAs), are required for membrane organization and cell integrity. PUFAs intake from dietary supplementation is essential as mammals lack the enzymes for its generation [126]. Omega-3 PUFAs improve the mitochondrial dysfunction by upregulating mitochondrial biogenesis, ATP production, and dissipating the proton gradient uncoupling proteins (UCPs) gene expression in vivo [127,128,129]. DHA is enriched in the retina, where it has both structural and neuroprotective functions, and is converted by lipoxygenase (LO) to 10,17S-docosatriene (neuroprotectin D1, NPD1) under oxidative stress conditions [130]. Furthermore, NPD1 inhibits pro-inflammatory and apoptotic gene expression, and consequently promotes the survival of photoreceptors [131]. Dietary manipulation of omega-3 PUFAs lowers IOP in aged rats and is associated with a significant increase in the outflow facility and a decrease in ocular rigidity [132]. Another study showed that a dietary combination of omega-3 and omega-6 PUFAs are more effective for preventing retinal cell structure and decreasing the glial cell activation [133]. An omega-3 fatty acid diet has been shown to have a retinal protective function in the AMD-like retinal lesions [134] and type 2 diabetic mice [135]. A recent study indicated that omega-3 PUFAs reduce lipofuscin granule formation and protect the photoreceptor layer. Its mechanism may involve an increase in the myelin basic protein (MBP), myelin proteolipid protein (MPP), myelin regulatory factor-like protein (MRFLP), and glial fibrillar acidic protein (GFAP) expression [136].
Omega-3 PUFAs intake is associated with a 30% decrease in the development of central geographic atrophy (CGA) and neovascular AMD [137]. Dietary supplementation with 4 g of omega-3 PUFAs for 6 months increases the serum omega-3 in patients with dry AMD by an average of 7.6%, however, there are no statistically significant changes in the retinal function of visual acuity or ERG [138]. The modified AREDS formulation also has a similar outcome in that the addition of omega-3 PUFAs in primary analyses did not reduce risk of progression to advanced AMD [125]. A recent AREDS2 study showed similar results, where omega-3 PUFAs did not demonstrate any significant benefit in the reduction in their risk for progression to late AMD in participants with CFH or age-related maculopathy susceptibility 2 (ARMS2) risk genotype [139]. Conversely, increasing dietary PUFA, rather than saturated FA, is associated with a reduced likelihood of the presence and severity of DR [140]. A Mediterranean diet with omega-3 PUFAs (≥500 mg/day) supplements also showed a 48% relatively reduced risk in the incidence sight-threatening DR in individuals with type 2 diabetes [141].
3.5. Curcumin
Curcumin is a polyphenol extracted from turmeric (Curcuma longa), which is used as a spice and as a traditional herbal medicine in Asia. It is a hydrophobic molecule and is almost insoluble in water (approximately 30 nM). Curcumin has strong free radical scavenging activity due to its functional groups and sequentially improves mitochondrial functions through the nuclear factor erythroid 2-related factor 2 (Nrf2) [142]. However, its poor solubility and low bioavailability have limited the clinical applications of curcumin (see a recent review in detail [143]). New strategies, including liposomes and nanoparticle carriers, or modified formulations may be an ideal approach to deliver curcumin to the lesions. For example, Davis et al. developed a curcumin nanocarrier combined with D-α-tocopherol polyethene glycol 1000 succinate (TPGS), a non-ionic surfactant, and Pluronic F127, a difunctional block copolymer surfactant, which increased curcumin solubility by 400,000 times and that enhanced curcumin transport across ocular barriers. A topically administered curcumin nanocarrier has neuroprotective effects of retinal cells in vitro and in vivo [144]. In addition, Cheng and colleagues recently reported a dual-drug delivery system which consisted of thermosensitive chitosan–gelatin-based hydrogel containing curcumin-loaded nanoparticles and latanoprost, which release medicine and was extended to 7 days. Treatment with curcumin-containing hydrogel effectively decreased the oxidative stress-mediated damage in trabecular meshwork cells [145].
Clinical application of curcumin has been broadly discussed in multiple malignant diseases [146]. However, there are few studies which report that curcumin has clinical benefits for eye disorders. Improved formulation may overcome this problem. One example is that oral administration of a curcumin-phospholipid delivery system is effective in the management of central serous chorioretinopathy (CSCR). The results show administration of curcumin significantly improves visual acuity and retinal thickness [147]. A recent review examined this issue and the authors categorize three broad formulation strategies to enhance bioavailability and metabolism of curcumin [148]. These well-designed formulations require more clinical trials to confirm their substantial benefits.
3.6. Crocetin
Crocetin is an apocarotenoid, which is found both in the saffron crocus (Crocus starus L.) and in gardenia fruit (Gardenia jasminoides Ellis) [149,150]. Saffron and its components (crocetin, crocins, and safranal) have therapeutic properties of liver, nervous and cardiovascular systems, including anti-oxidant, anti-inflammatory, and anti-apoptotic properties [151,152]. Crocetin rescues disruption of the ΔΨm induced by tunicamycin or hydrogen peroxide (H2O2) in vitro and has protective effects against retinal degeneration in vivo [153]. It is reported to inhibit oxidative stress via mitogen-activated protein kinases (MAPK), extracellular signal-regulated protein kinases (ERK), c-Jun N-terminal kinases (JNK), p38, and the redox-sensitive NF-κB and c-Jun pathway in an ischemia/reperfusion (I/R) mouse model [154]. A hydrophilic saffron extract standardized to 3% crocin reduces higher IOP values and activated microglia cells [155]. Saffron has been shown to have beneficial effects for ocular diseases in clinical studies (see a recent review [156]). However, a recent clinical study shows short-term saffron supplementation had no significant effects on the visual acuity and focal ERG in Stargardt disease/fundus flavimaculatus (STG/FF) patients with ATP binding cassette subfamily A member 4 (ABCA4) gene mutations [157].
3.7. Other Potential Nutraceuticals
Some potential nutraceuticals, like traditional Chinese medicine, have effects including neural and mitochondrial protection. Ginkgo biloba extract (GBE) contains the flavone glycosides and terpenoids, and showed stabilization and protection of mitochondrial function in Alzheimer’s disease [158]. GBE has various effects of antioxidant, microcirculation and neuroprotection activities in ocular diseases [159,160]. A topical formulation of GBE reduces IOP elevation and accumulation of extracellular materials in dexamethasone-induced ocular hypertension rabbits [161]. GBE administration has been shown to improve pre-existing visual field damage [162] and visual acuity analyzed by Humphrey Visual Field (HVF) [163] in patients with NTG. However, there is a recent study that showed no effect on mean defect or contrast sensitivity in Chinese patients with NTG [164].
Danshen (Salvia miltiorrhiza) is used for treating hyperlipidemia, acute ischemia, and stroke in traditional Chinese medicine [165]. Danshen extracts increase the levels of glutathione (GSH) and reduce the levels of malondialdehyde (MDA) in the eye tissues of hyperglycemic rats [166]. Salvianolic acids are a natural compound extracted from Danshen and more than 10 different salvianolic acids have been identified to date. Salvianolic acid A and B are the most effective and abundant compounds [167]. Salvianolic acid A has anti-oxidative stress potential, through the activation of Nrf2 and hemeoxygenase-1 (HO-1) expressions in RPE cells [168]. It also has an anti-angiogenesis function through the downregulation of cylindromatosis (CYLD) signaling pathways in choroidal neovascularization (CNV) mice [169]. A recent study showed that salvianolic acid A improved the mitochondrial function of high glucose-injured Schwann cells and diabetic peripheral neuropathy (DPN) in KK-Ay diabetic mice via upregulation of nuclear Nrf2 expression [170]. Salvianolic acid B has been reported to protect against oxidative injury through Nrf2 and glutaredoxin 1 (a thiol repair enzyme, Grx1) in primary human RPE cells [171]. A multiple-formula containing Danshen, notoginseng, and borneol (Compound Danshen Dripping Pill, CDDP) significantly improved the best corrected visual acuity and retinal pathogenesis in non-proliferative diabetic retinopathy (NPDR) patients [172]. Another study showed that CDDP significantly improved fluorescence fundus angiography and funduscopic examination parameters in NPDR patients [173].
Astragali Radix (Huangqi) is one of the most frequently used herbal medicines in traditional Chinese medicine and has a wide range of biological activities [174]. The extract of Astragalus mongholicus has been reported that protected oxidative damage through ameliorating activities of the mitochondrial complexes I, II, malate dehydrogenase and ΔΨm [175]. Astragalus polysaccharides protects mitochondria by scavenging ROS, inhibiting mitochondrial permeability transition and increasing the activities of catalase (CAT), SOD, and glutathione peroxidase (GPx) [176]. Huang and colleagues further demonstrated that Astragalus polysaccharides restored the imbalance of mitochondrial fusion–fission processes, activation of mitophagy, and decrease in PGC-1α expression in vivo [177]. The active compounds astragaloside IV and formononetin extracted from huangqi have also been indicated to inhibit aldose reductase (AR) and hypoxia-induced neovascularization, respectively [178,179]. These data suggest that extracts from huangqi may have therapeutic benefits for DR. Major findings are summarized in the Table 1 (animal studies) and Table 2 (clinical trials).
4. Conclusions
In this review, we summarize the underlying mechanisms of high-energy consumption and metabolic homeostasis that play a decisive role in the retina. The loss of balance between energy production and free radicals quenching causes oxidative stress, and further leads to mitochondrial dysfunction. The structural and functional integrity of the mitochondrion is important for maintaining the organization of retinal cells. Retinal neurodegeneration is a pathogenic result of mitochondrial dysfunction and contributes to an early stage of progression in retinal metabolic disorders.
A large body of evidence demonstrates that nutraceuticals target mitochondrial function to restore the mitochondrial flexibility. Some are essential nutrients and have benefits for both forming the cellular structure and scavenging ROS. Otherwise, multiple nutraceuticals with the potential for improving mitochondrial integrity have been reported in animal studies. Unfortunately, poor bioavailability and bioaccessibility limit their current therapeutic use. A new delivery system and improved formulation may bridge the gap between laboratory study and clinical treatment [180]. Future clinical trials require an additional focus on the next generation of nutraceuticals to confirm their health benefits.
Author Contributions
F.-J.T. and Y.-C.H. conceived and supervised all works; C.-P.H. and Y.-W.L. reviewed anal drafted the article. All authors have read and agreed to the published version of the manuscript.
Funding
The study was supported by research grants from the Ministry of Science and Technology, Taiwan (MOST 106-2320-B-015-MY2, MOST 108-2320-B-039-023-MY3 and MOST 109-2811-B-039-501).
Acknowledgments
We thank the Ministry of Science and Technology, Taiwan for support the research grants.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Schmidt, K.G.; Bergert, H.; Funk, R.H. Neurodegenerative diseases of the retina and potential for protection and recovery. Curr. Neuropharmacol. 2008, 6, 164–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong-Riley, M.T. Energy metabolism of the visual system. Eye Brain 2010, 2, 99–116. [Google Scholar] [CrossRef] [Green Version]
- Seung, H.S.; Sumbul, U. Neuronal cell types and connectivity: Lessons from the retina. Neuron 2014, 83, 1262–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, Y.A.; Di Polo, A. Mitochondrial dynamics, transport, and quality control: A bottleneck for retinal ganglion cell viability in optic neuropathies. Mitochondrion 2017, 36, 186–192. [Google Scholar] [CrossRef]
- Roy, S.; Kim, D.; Sankaramoorthy, A. Mitochondrial structural changes in the pathogenesis of diabetic retinopathy. J. Clin. Med. 2019, 8, 1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowluru, R.A. Mitochondrial stability in diabetic retinopathy: Lessons learned from epigenetics. Diabetes 2019, 68, 241–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Federico, A.; Cardaioli, E.; Da Pozzo, P.; Formichi, P.; Gallus, G.N.; Radi, E. Mitochondria, oxidative stress and neurodegeneration. J. Neurol. Sci. 2012, 322, 254–262. [Google Scholar] [CrossRef]
- Wang, Y.; Branicky, R.; Noe, A.; Hekimi, S. Superoxide dismutases: Dual roles in controlling ros damage and regulating ros signaling. J. Cell Biol. 2018, 217, 1915–1928. [Google Scholar] [CrossRef]
- Raha, S.; Robinson, B.H. Mitochondria, oxygen free radicals, disease and ageing. Trends Biochem. Sci. 2000, 25, 502–508. [Google Scholar] [CrossRef]
- Schinder, A.F.; Olson, E.C.; Spitzer, N.C.; Montal, M. Mitochondrial dysfunction is a primary event in glutamate neurotoxicity. J. Neurosci. 1996, 16, 6125–6133. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, D.; Alavi, M.V.; Kim, K.Y.; Kang, T.; Scott, R.T.; Noh, Y.H.; Lindsey, J.D.; Wissinger, B.; Ellisman, M.H.; Weinreb, R.N.; et al. A new vicious cycle involving glutamate excitotoxicity, oxidative stress and mitochondrial dynamics. Cell Death Dis. 2011, 2, e240. [Google Scholar] [CrossRef]
- Rego, A.C.; Oliveira, C.R. Mitochondrial dysfunction and reactive oxygen species in excitotoxicity and apoptosis: Implications for the pathogenesis of neurodegenerative diseases. Neurochem. Res. 2003, 28, 1563–1574. [Google Scholar] [CrossRef] [Green Version]
- Abramov, A.Y.; Scorziello, A.; Duchen, M.R. Three distinct mechanisms generate oxygen free radicals in neurons and contribute to cell death during anoxia and reoxygenation. J. Neurosci. 2007, 27, 1129–1138. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, E.M.; Emri, E.; Merle, B.M.J.; Colijn, J.M.; Kersten, E.; Cougnard-Gregoire, A.; Dammeier, S.; Meester-Smoor, M.; Pool, F.M.; De Jong, E.K.; et al. A new perspective on lipid research in age-related macular degeneration. Prog. Retin. Eye Res. 2018, 67, 56–86. [Google Scholar] [CrossRef] [PubMed]
- Pilz, Y.L.; Bass, S.J.; Sherman, J. A review of mitochondrial optic neuropathies: From inherited to acquired forms. J. Optom. 2017, 10, 205–214. [Google Scholar] [CrossRef]
- Chrysostomou, V.; Rezania, F.; Trounce, I.A.; Crowston, J.G. Oxidative stress and mitochondrial dysfunction in glaucoma. Curr. Opin. Pharmacol. 2013, 13, 12–15. [Google Scholar] [CrossRef] [PubMed]
- Barot, M.; Gokulgandhi, M.R.; Mitra, A.K. Mitochondrial dysfunction in retinal diseases. Curr. Eye Res. 2011, 36, 1069–1077. [Google Scholar] [CrossRef] [Green Version]
- Joyal, J.S.; Gantner, M.L.; Smith, L.E.H. Retinal energy demands control vascular supply of the retina in development and disease: The role of neuronal lipid and glucose metabolism. Prog. Retin. Eye Res. 2018, 64, 131–156. [Google Scholar] [CrossRef]
- Wu, S.; Zhou, F.; Zhang, Z.; Xing, D. Mitochondrial oxidative stress causes mitochondrial fragmentation via differential modulation of mitochondrial fission-fusion proteins. FEBS J. 2011, 278, 941–954. [Google Scholar] [CrossRef]
- Willems, P.H.; Rossignol, R.; Dieteren, C.E.; Murphy, M.P.; Koopman, W.J. Redox homeostasis and mitochondrial dynamics. Cell Metab. 2015, 22, 207–218. [Google Scholar] [CrossRef] [Green Version]
- Yu-Wai-Man, P.; Griffiths, P.G.; Chinnery, P.F. Mitochondrial optic neuropathies—disease mechanisms and therapeutic strategies. Prog. Retin. Eye Res. 2011, 30, 81–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaens, K.H.; Stehouwer, C.D.; Schalkwijk, C.G. Advanced glycation endproducts and its receptor for advanced glycation endproducts in obesity. Curr. Opin. Lipidol. 2013, 24, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Geraldes, P.; King, G.L. Activation of protein kinase c isoforms and its impact on diabetic complications. Circ. Res. 2010, 106, 1319–1331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bristow, E.A.; Griffiths, P.G.; Andrews, R.M.; Johnson, M.A.; Turnbull, D.M. The distribution of mitochondrial activity in relation to optic nerve structure. Arch. Ophthalmol. 2002, 120, 791–796. [Google Scholar] [CrossRef] [PubMed]
- Stone, J.; van Driel, D.; Valter, K.; Rees, S.; Provis, J. The locations of mitochondria in mammalian photoreceptors: Relation to retinal vasculature. Brain Res. 2008, 1189, 58–69. [Google Scholar] [CrossRef]
- Kanow, M.A.; Giarmarco, M.M.; Jankowski, C.S.; Tsantilas, K.; Engel, A.L.; Du, J.; Linton, J.D.; Farnsworth, C.C.; Sloat, S.R.; Rountree, A.; et al. Biochemical adaptations of the retina and retinal pigment epithelium support a metabolic ecosystem in the vertebrate eye. Elife 2017, 6, e28899. [Google Scholar] [CrossRef]
- Brown, E.E.; DeWeerd, A.J.; Ildefonso, C.J.; Lewin, A.S.; Ash, J.D. Mitochondrial oxidative stress in the retinal pigment epithelium (rpe) led to metabolic dysfunction in both the rpe and retinal photoreceptors. Redox Biol. 2019, 24, 101201. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.Y.; Cringle, S.J. Oxygen distribution and consumption within the retina in vascularised and avascular retinas and in animal models of retinal disease. Prog. Retin. Eye Res. 2001, 20, 175–208. [Google Scholar] [CrossRef]
- Kern, T.S.; Berkowitz, B.A. Photoreceptors in diabetic retinopathy. J. Diabetes Investig. 2015, 6, 371–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, K.C.; Rosales, M.A.; Biswas, S.K.; Lopes de Faria, J.B.; Lopes de Faria, J.M. Diabetic retinal neurodegeneration is associated with mitochondrial oxidative stress and is improved by an angiotensin receptor blocker in a model combining hypertension and diabetes. Diabetes 2009, 58, 1382–1390. [Google Scholar] [CrossRef] [Green Version]
- Kong, G.Y.; Van Bergen, N.J.; Trounce, I.A.; Crowston, J.G. Mitochondrial dysfunction and glaucoma. J. Glaucoma 2009, 18, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Dadhania, V.P.; Trivedi, P.P.; Vikram, A.; Tripathi, D.N. Nutraceuticals against neurodegeneration: A mechanistic insight. Curr. Neuropharmacol. 2016, 14, 627–640. [Google Scholar] [CrossRef] [PubMed]
- Kelsey, N.A.; Wilkins, H.M.; Linseman, D.A. Nutraceutical antioxidants as novel neuroprotective agents. Molecules 2010, 15, 7792–7814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinreb, R.N.; Aung, T.; Medeiros, F.A. The pathophysiology and treatment of glaucoma: A review. Jama 2014, 311, 1901–1911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sacca, S.C.; Izzotti, A. Oxidative stress and glaucoma: Injury in the anterior segment of the eye. Prog. Brain Res. 2008, 173, 385–407. [Google Scholar]
- Ju, W.K.; Kim, K.Y.; Lindsey, J.D.; Angert, M.; Duong-Polk, K.X.; Scott, R.T.; Kim, J.J.; Kukhmazov, I.; Ellisman, M.H.; Perkins, G.A.; et al. Intraocular pressure elevation induces mitochondrial fission and triggers opa1 release in glaucomatous optic nerve. Investig. Ophthalmol. Vis. Sci. 2008, 49, 4903–4911. [Google Scholar] [CrossRef]
- Chen, Y.; Hughes, G.; Chen, X.; Qian, S.; Cao, W.; Wang, L.; Wang, M.; Sun, X. Genetic variants associated with different risks for high tension glaucoma and normal tension glaucoma in a chinese population. Investig. Ophthalmol. Vis. Sci. 2015, 56, 2595–2600. [Google Scholar] [CrossRef] [Green Version]
- Marek, B.; Harris, A.; Kanakamedala, P.; Lee, E.; Amireskandari, A.; Carichino, L.; Guidoboni, G.; Tobe, L.A.; Siesky, B. Cerebrospinal fluid pressure and glaucoma: Regulation of trans-lamina cribrosa pressure. Br. J. Ophthalmol. 2014, 98, 721–725. [Google Scholar] [CrossRef]
- Pinazo-Duran, M.D.; Shoaie-Nia, K.; Zanon-Moreno, V.; Sanz-Gonzalez, S.M.; Del Castillo, J.B.; Garcia-Medina, J.J. Strategies to reduce oxidative stress in glaucoma patients. Curr. Neuropharmacol. 2018, 16, 903–918. [Google Scholar] [CrossRef]
- Tezel, G. Oxidative stress in glaucomatous neurodegeneration: Mechanisms and consequences. Prog. Retin. Eye Res. 2006, 25, 490–513. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, P.; Liew, G.; Gopinath, B.; Wong, T.Y. Age-related macular degeneration. Lancet 2018, 392, 1147–1159. [Google Scholar] [CrossRef]
- Donoso, L.A.; Vrabec, T.; Kuivaniemi, H. The role of complement factor h in age-related macular degeneration: A review. Surv. Ophthalmol. 2010, 55, 227–246. [Google Scholar] [CrossRef]
- Wu, Z.; Lauer, T.W.; Sick, A.; Hackett, S.F.; Campochiaro, P.A. Oxidative stress modulates complement factor h expression in retinal pigmented epithelial cells by acetylation of foxo3. J. Biol. Chem. 2007, 282, 22414–22425. [Google Scholar] [CrossRef] [Green Version]
- Thurman, J.M.; Renner, B.; Kunchithapautham, K.; Ferreira, V.P.; Pangburn, M.K.; Ablonczy, Z.; Tomlinson, S.; Holers, V.M.; Rohrer, B. Oxidative stress renders retinal pigment epithelial cells susceptible to complement-mediated injury. J. Biol. Chem. 2009, 284, 16939–16947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weismann, D.; Hartvigsen, K.; Lauer, N.; Bennett, K.L.; Scholl, H.P.; Charbel Issa, P.; Cano, M.; Brandstatter, H.; Tsimikas, S.; Skerka, C.; et al. Complement factor h binds malondialdehyde epitopes and protects from oxidative stress. Nature 2011, 478, 76–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, P.; Wang, J.J.; Smith, W.; Leeder, S.R. Smoking and the 5-year incidence of age-related maculopathy: The blue mountains eye study. Arch. Ophthalmol. 2002, 120, 1357–1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunchithapautham, K.; Atkinson, C.; Rohrer, B. Smoke exposure causes endoplasmic reticulum stress and lipid accumulation in retinal pigment epithelium through oxidative stress and complement activation. J. Biol. Chem. 2014, 289, 14534–14546. [Google Scholar] [CrossRef] [Green Version]
- Feher, J.; Kovacs, I.; Artico, M.; Cavallotti, C.; Papale, A.; Balacco Gabrieli, C. Mitochondrial alterations of retinal pigment epithelium in age-related macular degeneration. Neurobiol. Aging 2006, 27, 983–993. [Google Scholar] [CrossRef] [PubMed]
- Terluk, M.R.; Kapphahn, R.J.; Soukup, L.M.; Gong, H.; Gallardo, C.; Montezuma, S.R.; Ferrington, D.A. Investigating mitochondria as a target for treating age-related macular degeneration. J. Neurosci. 2015, 35, 7304–7311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaarniranta, K.; Pawlowska, E.; Szczepanska, J.; Jablkowska, A.; Blasiak, J. Role of mitochondrial DNA damage in ros-mediated pathogenesis of age-related macular degeneration (amd). Int. J. Mol. Sci. 2019, 20, 2374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collaboration, N.C.D.R.F. Worldwide trends in diabetes since 1980: A pooled analysis of 751 population-based studies with 4.4 million participants. Lancet 2016, 387, 1513–1530. [Google Scholar]
- Mohamed, Q.; Gillies, M.C.; Wong, T.Y. Management of diabetic retinopathy: A systematic review. JAMA 2007, 298, 902–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solomon, S.D.; Chew, E.; Duh, E.J.; Sobrin, L.; Sun, J.K.; VanderBeek, B.L.; Wykoff, C.C.; Gardner, T.W. Diabetic retinopathy: A position statement by the american diabetes association. Diabetes Care 2017, 40, 412–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simo, R.; Stitt, A.W.; Gardner, T.W. Neurodegeneration in diabetic retinopathy: Does it really matter? Diabetologia 2018, 61, 1902–1912. [Google Scholar] [CrossRef] [Green Version]
- Diabetes, C.; Nathan, D.M.; Genuth, S.; Lachin, J.; Cleary, P.; Crofford, O.; Davis, M.; Rand, L.; Siebert, C. Complications Trial Research Group. The effect of intensive treatment of diabetes on the development and progression of long-term complications in insulin-dependent diabetes mellitus. N. Engl. J. Med. 1993, 329, 977–986. [Google Scholar]
- Nathan, D.M.; Group, D.E.R. The diabetes control and complications trial/epidemiology of diabetes interventions and complications study at 30 years: Overview. Diabetes Care 2014, 37, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Carrasco, E.; Hernandez, C.; Miralles, A.; Huguet, P.; Farres, J.; Simo, R. Lower somatostatin expression is an early event in diabetic retinopathy and is associated with retinal neurodegeneration. Diabetes Care 2007, 30, 2902–2908. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Ramirez, M.; Hernandez, C.; Villarroel, M.; Canals, F.; Alonso, M.A.; Fortuny, R.; Masmiquel, L.; Navarro, A.; Garcia-Arumi, J.; Simo, R. Interphotoreceptor retinoid-binding protein (irbp) is downregulated at early stages of diabetic retinopathy. Diabetologia 2009, 52, 2633–2641. [Google Scholar] [CrossRef] [Green Version]
- Barber, A.J.; Lieth, E.; Khin, S.A.; Antonetti, D.A.; Buchanan, A.G.; Gardner, T.W. Neural apoptosis in the retina during experimental and human diabetes. Early onset and effect of insulin. J. Clin. Investig. 1998, 102, 783–791. [Google Scholar] [CrossRef] [Green Version]
- Vujosevic, S.; Midena, E. Retinal layers changes in human preclinical and early clinical diabetic retinopathy support early retinal neuronal and muller cells alterations. J. Diabetes Res. 2013, 2013, 905058. [Google Scholar] [CrossRef]
- Sohn, E.H.; Van Dijk, H.W.; Jiao, C.; Kok, P.H.; Jeong, W.; Demirkaya, N.; Garmager, A.; Wit, F.; Kucukevcilioglu, M.; Van Velthoven, M.E.; et al. Retinal neurodegeneration may precede microvascular changes characteristic of diabetic retinopathy in diabetes mellitus. Proc. Natl. Acad. Sci. USA 2016, 113, E2655–E2664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanek, J.; Blair, N.P.; Chau, F.Y.; Lim, J.I.; Leiderman, Y.I.; Shahidi, M. Alterations in retinal layer thickness and reflectance at different stages of diabetic retinopathy by en face optical coherence tomography. Investig. Ophthalmol. Vis. Sci. 2016, 57, OCT341–OCT347. [Google Scholar] [CrossRef]
- Chen, S.Y.; Hsu, Y.M.; Lin, Y.J.; Huang, Y.C.; Chen, C.J.; Lin, W.D.; Liao, W.L.; Chen, Y.T.; Lin, W.Y.; Liu, Y.H.; et al. Current concepts regarding developmental mechanisms in diabetic retinopathy in Taiwan. BioMedicine 2016, 6, 7. [Google Scholar] [CrossRef] [PubMed]
- Bek, T. Mitochondrial dysfunction and diabetic retinopathy. Mitochondrion 2017, 36, 4–6. [Google Scholar] [CrossRef] [PubMed]
- Kowluru, R.A.; Mishra, M. Therapeutic targets for altering mitochondrial dysfunction associated with diabetic retinopathy. Expert. Opin. Ther. Targets 2018, 22, 233–245. [Google Scholar] [CrossRef]
- Durham, J.T.; Herman, I.M. Microvascular modifications in diabetic retinopathy. Curr. Diabetes Rep. 2011, 11, 253–264. [Google Scholar] [CrossRef]
- Simo, R.; Hernandez, C. European Consortium for the Early Treatment of Diabetic, R. Neurodegeneration in the diabetic eye: New insights and therapeutic perspectives. Trends Endocrinol. Metab. 2014, 25, 23–33. [Google Scholar] [CrossRef]
- Lekli, I.; Ray, D.; Das, D.K. Longevity nutrients resveratrol, wines and grapes. Genes Nutr. 2010, 5, 55–60. [Google Scholar] [CrossRef] [Green Version]
- Lagouge, M.; Argmann, C.; Gerhart-Hines, Z.; Meziane, H.; Lerin, C.; Daussin, F.; Messadeq, N.; Milne, J.; Lambert, P.; Elliott, P.; et al. Resveratrol improves mitochondrial function and protects against metabolic disease by activating sirt1 and pgc-1alpha. Cell 2006, 127, 1109–1122. [Google Scholar] [CrossRef]
- Abu-Amero, K.K.; Kondkar, A.A.; Chalam, K.V. Resveratrol and ophthalmic diseases. Nutrients 2016, 8, 200. [Google Scholar] [CrossRef] [Green Version]
- Pirhan, D.; Yuksel, N.; Emre, E.; Cengiz, A.; Kursat Yildiz, D. Riluzole-and resveratrol-induced delay of retinal ganglion cell death in an experimental model of glaucoma. Curr. Eye Res. 2016, 41, 59–69. [Google Scholar] [CrossRef]
- Lindsey, J.D.; Duong-Polk, K.X.; Hammond, D.; Leung, C.K.; Weinreb, R.N. Protection of injured retinal ganglion cell dendrites and unfolded protein response resolution after long-term dietary resveratrol. Neurobiol. Aging 2015, 36, 1969–1981. [Google Scholar] [CrossRef] [PubMed]
- Razali, N.; Agarwal, R.; Agarwal, P.; Kumar, S.; Tripathy, M.; Vasudevan, S.; Crowston, J.G.; Ismail, N.M. Role of adenosine receptors in resveratrol-induced intraocular pressure lowering in rats with steroid-induced ocular hypertension. Clin. Exp. Ophthalmol. 2015, 43, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Natesan, S.; Pandian, S.; Ponnusamy, C.; Palanichamy, R.; Muthusamy, S.; Kandasamy, R. Co-encapsulated resveratrol and quercetin in chitosan and peg modified chitosan nanoparticles: For efficient intra ocular pressure reduction. Int. J. Biol. Macromol. 2017, 104, 1837–1845. [Google Scholar] [CrossRef]
- Kubota, S.; Kurihara, T.; Ebinuma, M.; Kubota, M.; Yuki, K.; Sasaki, M.; Noda, K.; Ozawa, Y.; Oike, Y.; Ishida, S.; et al. Resveratrol prevents light-induced retinal degeneration via suppressing activator protein-1 activation. Am. J. Pathol. 2010, 177, 1725–1731. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wu, Z.; Li, J.; Marmalidou, A.; Zhang, R.; Yu, M. Protective effect of resveratrol against light-induced retinal degeneration in aged samp8 mice. Oncotarget 2017, 8, 65778–65788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soufi, F.G.; Mohammad-Nejad, D.; Ahmadieh, H. Resveratrol improves diabetic retinopathy possibly through oxidative stress - nuclear factor kappab - apoptosis pathway. Pharmacol. Rep. 2012, 64, 1505–1514. [Google Scholar] [CrossRef]
- Sedlak, L.; Wojnar, W.; Zych, M.; Wygledowska-Promienska, D.; Mrukwa-Kominek, E.; Kaczmarczyk-Sedlak, I. Effect of resveratrol, a dietary-derived polyphenol, on the oxidative stress and polyol pathway in the lens of rats with streptozotocin-induced diabetes. Nutrients 2018, 10, 1423. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Meng, J.; Li, H.; Wei, H.; Bi, F.; Liu, S.; Tang, K.; Guo, H.; Liu, W. Resveratrol exhibits an effect on attenuating retina inflammatory condition and damage of diabetic retinopathy via pon1. Exp. Eye Res. 2019, 181, 356–366. [Google Scholar] [CrossRef]
- Kim, Y.H.; Kim, Y.S.; Roh, G.S.; Choi, W.S.; Cho, G.J. Resveratrol blocks diabetes-induced early vascular lesions and vascular endothelial growth factor induction in mouse retinas. Acta Ophthalmol. 2012, 90, e31–e37. [Google Scholar] [CrossRef]
- Berman, A.Y.; Motechin, R.A.; Wiesenfeld, M.Y.; Holz, M.K. The therapeutic potential of resveratrol: A review of clinical trials. NPJ Precis. Oncol. 2017, 1, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.P.; Singh, R.; Verma, S.S.; Rai, V.; Kaschula, C.H.; Maiti, P.; Gupta, S.C. Health benefits of resveratrol: Evidence from clinical studies. Med. Res. Rev. 2019, 39, 1851–1891. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Garza, S.L.; Laveriano-Santos, E.P.; Marhuenda-Munoz, M.; Storniolo, C.E.; Tresserra-Rimbau, A.; Vallverdu-Queralt, A.; Lamuela-Raventos, R.M. Health effects of resveratrol: Results from human intervention trials. Nutrients 2018, 10, 1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bola, C.; Bartlett, H.; Eperjesi, F. Resveratrol and the eye: Activity and molecular mechanisms. Graefe’s Arch. Clin. Exp. Ophthalmol. = Albrecht Graefes Arch. Klin. Exp. Ophthalmol. 2014, 252, 699–713. [Google Scholar] [CrossRef] [Green Version]
- Richer, S.; Stiles, W.; Ulanski, L.; Carroll, D.; Podella, C. Observation of human retinal remodeling in octogenarians with a resveratrol based nutritional supplement. Nutrients 2013, 5, 1989–2005. [Google Scholar] [CrossRef]
- Timmers, S.; De Ligt, M.; Phielix, E.; Van de Weijer, T.; Hansen, J.; Moonen-Kornips, E.; Schaart, G.; Kunz, I.; Hesselink, M.K.; Schrauwen-Hinderling, V.B.; et al. Resveratrol as add-on therapy in subjects with well-controlled type 2 diabetes: A randomized controlled trial. Diabetes Care 2016, 39, 2211–2217. [Google Scholar] [CrossRef] [Green Version]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [Green Version]
- Boots, A.W.; Haenen, G.R.; Bast, A. Health effects of quercetin: From antioxidant to nutraceutical. Eur. J. Pharmacol. 2008, 585, 325–337. [Google Scholar] [CrossRef]
- Qiu, L.; Luo, Y.; Chen, X. Quercetin attenuates mitochondrial dysfunction and biogenesis via upregulated ampk/sirt1 signaling pathway in oa rats. Biomed. Pharmacother. 2018, 103, 1585–1591. [Google Scholar] [CrossRef]
- Wang, D.M.; Li, S.Q.; Wu, W.L.; Zhu, X.Y.; Wang, Y.; Yuan, H.Y. Effects of long-term treatment with quercetin on cognition and mitochondrial function in a mouse model of alzheimer’s disease. Neurochem. Res. 2014, 39, 1533–1543. [Google Scholar] [CrossRef]
- Nakayama, M.; Aihara, M.; Chen, Y.N.; Araie, M.; Tomita-Yokotani, K.; Iwashina, T. Neuroprotective effects of flavonoids on hypoxia-, glutamate-, and oxidative stress-induced retinal ganglion cell death. Mol. Vis. 2011, 17, 1784–1793. [Google Scholar]
- Maher, P.; Hanneken, A. Flavonoids protect retinal ganglion cells from oxidative stress-induced death. Investig. Ophthalmol. Vis. Sci. 2005, 46, 4796–4803. [Google Scholar] [CrossRef]
- Gao, F.J.; Zhang, S.H.; Xu, P.; Yang, B.Q.; Zhang, R.; Cheng, Y.; Zhou, X.J.; Huang, W.J.; Wang, M.; Chen, J.Y.; et al. Quercetin declines apoptosis, ameliorates mitochondrial function and improves retinal ganglion cell survival and function in in vivo model of glaucoma in rat and retinal ganglion cell culture in vitro. Front. Mol. Neurosci. 2017, 10, 285. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Li, G.; Yang, B.; Wu, J. Quercetin enhances inhibitory synaptic inputs and reduces excitatory synaptic inputs to off- and on-type retinal ganglion cells in a chronic glaucoma rat model. Front. Neurosci. 2019, 13, 672. [Google Scholar] [CrossRef] [PubMed]
- Park, K.H.; Cozier, F.; Ong, O.C.; Caprioli, J. Induction of heat shock protein 72 protects retinal ganglion cells in a rat glaucoma model. Investig. Ophthalmol. Vis. Sci. 2001, 42, 1522–1530. [Google Scholar]
- Caprioli, J.; Ishii, Y.; Kwong, J.M. Retinal ganglion cell protection with geranylgeranylacetone, a heat shock protein inducer, in a rat glaucoma model. Trans. Am. Ophthalmol. Soc. 2003, 101, 39–50. [Google Scholar]
- Li, N.; Li, Y.; Duan, X. Heat shock protein 72 confers protection in retinal ganglion cells and lateral geniculate nucleus neurons via blockade of the sapk/jnk pathway in a chronic ocular-hypertensive rat model. Neural Regen. Res. 2014, 9, 1395–1401. [Google Scholar] [PubMed]
- Kim, J.; Jin, H.L.; Jang, D.S.; Jeong, K.W.; Choung, S.Y. Quercetin-3-o-alpha-l-arabinopyranoside protects against retinal cell death via blue light-induced damage in human rpe cells and balb-c mice. Food Funct. 2018, 9, 2171–2183. [Google Scholar] [CrossRef] [PubMed]
- Koyama, Y.; Kaidzu, S.; Kim, Y.C.; Matsuoka, Y.; Ishihara, T.; Ohira, A.; Tanito, M. Suppression of light-induced retinal degeneration by quercetin via the ap-1 pathway in rats. Antioxidants 2019, 8, 79. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Kang, M.J.; Choi, H.N.; Jeong, S.M.; Lee, Y.M.; Kim, J.I. Quercetin attenuates fasting and postprandial hyperglycemia in animal models of diabetes mellitus. Nutr. Res. Pract. 2011, 5, 107–111. [Google Scholar] [CrossRef] [Green Version]
- Jeong, S.M.; Kang, M.J.; Choi, H.N.; Kim, J.H.; Kim, J.I. Quercetin ameliorates hyperglycemia and dyslipidemia and improves antioxidant status in type 2 diabetic db/db mice. Nutr. Res. Pract. 2012, 6, 201–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, B.; Gupta, S.K.; Nag, T.C.; Srivastava, S.; Saxena, R.; Jha, K.A.; Srinivasan, B.P. Retinal neuroprotective effects of quercetin in streptozotocin-induced diabetic rats. Exp. Eye Res. 2014, 125, 193–202. [Google Scholar] [CrossRef]
- Mohammadi-Sartang, M.; Mazloom, Z.; Sherafatmanesh, S.; Ghorbani, M.; Firoozi, D. Effects of supplementation with quercetin on plasma c-reactive protein concentrations: A systematic review and meta-analysis of randomized controlled trials. Eur. J. Clin. Nutr. 2017, 71, 1033–1039. [Google Scholar] [CrossRef]
- Boots, A.W.; Drent, M.; De Boer, V.C.; Bast, A.; Haenen, G.R. Quercetin reduces markers of oxidative stress and inflammation in sarcoidosis. Clin. Nutr. 2011, 30, 506–512. [Google Scholar] [CrossRef] [Green Version]
- Ferry, D.R.; Smith, A.; Malkhandi, J.; Fyfe, D.W.; De Takats, P.G.; Anderson, D.; Baker, J.; Kerr, D.J. Phase i clinical trial of the flavonoid quercetin: Pharmacokinetics and evidence for in vivo tyrosine kinase inhibition. Clin. Cancer Res. 1996, 2, 659–668. [Google Scholar]
- Ramos, S. Cancer chemoprevention and chemotherapy: Dietary polyphenols and signalling pathways. Mol. Nutr. Food Res. 2008, 52, 507–526. [Google Scholar] [CrossRef]
- Kooshyar, M.M.; Mozafari, P.M.; Amirchaghmaghi, M.; Pakfetrat, A.; Karoos, P.; Mohasel, M.R.; Orafai, H.; Azarian, A.A. A randomized placebo- controlled double blind clinical trial of quercetin in the prevention and treatment of chemotherapy-induced oral mucositis. J. Clin. Diagn. Res. 2017, 11, ZC4–ZC50. [Google Scholar] [CrossRef]
- Gopinath, B.; Liew, G.; Kifley, A.; Flood, V.M.; Joachim, N.; Lewis, J.R.; Hodgson, J.M.; Mitchell, P. Dietary flavonoids and the prevalence and 15-y incidence of age-related macular degeneration. Am. J. Clin. Nutr. 2018, 108, 381–387. [Google Scholar] [CrossRef]
- Sommerburg, O.G.; Siems, W.G.; Hurst, J.S.; Lewis, J.W.; Kliger, D.S.; Van Kuijk, F.J. Lutein and zeaxanthin are associated with photoreceptors in the human retina. Curr. Eye Res. 1999, 19, 491–495. [Google Scholar] [CrossRef]
- Tan, B.L.; Norhaizan, M.E. Carotenoids: How effective are they to prevent age-related diseases? Molecules 2019, 24, 1801. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, M.; Ozawa, Y.; Kurihara, T.; Noda, K.; Imamura, Y.; Kobayashi, S.; Ishida, S.; Tsubota, K. Neuroprotective effect of an antioxidant, lutein, during retinal inflammation. Investig. Ophthalmol. Vis. Sci. 2009, 50, 1433–1439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnal, E.; Miranda, M.; Johnsen-Soriano, S.; Alvarez-Nolting, R.; Diaz-Llopis, M.; Araiz, J.; Cervera, E.; Bosch-Morell, F.; Romero, F.J. Beneficial effect of docosahexanoic acid and lutein on retinal structural, metabolic, and functional abnormalities in diabetic rats. Curr. Eye Res. 2009, 34, 928–938. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Vachali, P.P.; Gorusupudi, A.; Shen, Z.; Sharifzadeh, H.; Besch, B.M.; Nelson, K.; Horvath, M.M.; Frederick, J.M.; Baehr, W.; et al. Inactivity of human beta,beta-carotene-9′,10′-dioxygenase (bco2) underlies retinal accumulation of the human macular carotenoid pigment. Proc. Natl. Acad. Sci. USA 2014, 111, 10173–10178. [Google Scholar] [CrossRef] [Green Version]
- Widjaja-Adhi, M.A.K.; Ramkumar, S.; Von Lintig, J. Protective role of carotenoids in the visual cycle. FASEB J. 2018, 32, 6305–6315. [Google Scholar] [CrossRef]
- Woo, T.T.; Li, S.Y.; Lai, W.W.; Wong, D.; Lo, A.C. Neuroprotective effects of lutein in a rat model of retinal detachment. Graefe’s Arch. Clin. Exp. Ophthalmol. = Albrecht Graefes Arch. Klin. Exp. Ophthalmol. 2013, 251, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Kamoshita, M.; Toda, E.; Osada, H.; Narimatsu, T.; Kobayashi, S.; Tsubota, K.; Ozawa, Y. Lutein acts via multiple antioxidant pathways in the photo-stressed retina. Sci. Rep. 2016, 6, 30226. [Google Scholar] [CrossRef]
- Yu, M.; Yan, W.; Beight, C. Lutein and zeaxanthin isomers protect against light-induced retinopathy via decreasing oxidative and endoplasmic reticulum stress in balb/cj mice. Nutrients 2018, 10, 842. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Wark, L.; Ji, H.; Willard, L.; Jaing, Y.; Han, J.; He, H.; Ortiz, E.; Zhang, Y.; Medeiros, D.M.; et al. Dietary wolfberry upregulates carotenoid metabolic genes and enhances mitochondrial biogenesis in the retina of db/db diabetic mice. Mol. Nutr. Food Res. 2013, 57, 1158–1169. [Google Scholar] [CrossRef] [Green Version]
- Mohn, E.S.; Erdman, J.W., Jr.; Neuringer, M.; Kuchan, M.J.; Johnson, E.J. Brain xanthophyll content and exploratory gene expression analysis: Subspecies differences in rhesus macaque. Genes Nutr. 2017, 12, 9. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, P.S.; Li, B.; Vachali, P.P.; Gorusupudi, A.; Shyam, R.; Henriksen, B.S.; Nolan, J.M. Lutein, zeaxanthin, and meso-zeaxanthin: The basic and clinical science underlying carotenoid-based nutritional interventions against ocular disease. Prog. Retin. Eye Res. 2016, 50, 34–66. [Google Scholar] [CrossRef] [Green Version]
- Moeller, S.M.; Parekh, N.; Tinker, L.; Ritenbaugh, C.; Blodi, B.; Wallace, R.B.; Mares, J.A.; Group, C.R.S. Associations between intermediate age-related macular degeneration and lutein and zeaxanthin in the carotenoids in age-related eye disease study (careds): Ancillary study of the women’s health initiative. Arch. Ophthalmol. 2006, 124, 1151–1162. [Google Scholar] [CrossRef]
- Gale, C.R.; Hall, N.F.; Phillips, D.I.; Martyn, C.N. Lutein and zeaxanthin status and risk of age-related macular degeneration. Investig. Ophthalmol. Vis. Sci. 2003, 44, 2461–2465. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Dou, H.L.; Wu, Y.Q.; Huang, Y.M.; Huang, Y.B.; Xu, X.R.; Zou, Z.Y.; Lin, X.M. Lutein and zeaxanthin intake and the risk of age-related macular degeneration: A systematic review and meta-analysis. Br. J. Nutr. 2012, 107, 350–359. [Google Scholar] [CrossRef]
- Raman, G.; Haslam, D.; Avendano, E.; Johnson, E.J. Lutein/zeaxanthin intake and visual outcomes in adults with healthy eyes: Qualitative gap analysis. Cogent Med. 2019, 6, 1683939. [Google Scholar] [CrossRef]
- Chew, E.Y.; Clemons, T.E.; SanGiovanni, J.P.; Danis, R.; Ferris, F.L.; Elman, M.; Antoszyk, A.; Ruby, A.; Orth, D.; Bressler, S.; et al. Age-Related Eye Disease Study 2 Research, G. Lutein + zeaxanthin and omega-3 fatty acids for age-related macular degeneration: The age-related eye disease study 2 (areds2) randomized clinical trial. JAMA 2013, 309, 2005–2015. [Google Scholar]
- Bazan, N.G.; Molina, M.F.; Gordon, W.C. Docosahexaenoic acid signalolipidomics in nutrition: Significance in aging, neuroinflammation, macular degeneration, alzheimer’s, and other neurodegenerative diseases. Annu. Rev. Nutr. 2011, 31, 321–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cha, S.H.; Fukushima, A.; Sakuma, K.; Kagawa, Y. Chronic docosahexaenoic acid intake enhances expression of the gene for uncoupling protein 3 and affects pleiotropic mrna levels in skeletal muscle of aged c57bl/6njcl mice. J. Nutr. 2001, 131, 2636–2642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flachs, P.; Horakova, O.; Brauner, P.; Rossmeisl, M.; Pecina, P.; Franssen-van Hal, N.; Ruzickova, J.; Sponarova, J.; Drahota, Z.; Vlcek, C.; et al. Polyunsaturated fatty acids of marine origin upregulate mitochondrial biogenesis and induce beta-oxidation in white fat. Diabetologia 2005, 48, 2365–2375. [Google Scholar] [CrossRef] [Green Version]
- Afshordel, S.; Hagl, S.; Werner, D.; Rohner, N.; Kogel, D.; Bazan, N.G.; Eckert, G.P. Omega-3 polyunsaturated fatty acids improve mitochondrial dysfunction in brain aging--impact of bcl-2 and npd-1 like metabolites. Prostaglandins Leukot. Essent. Fat. Acids 2015, 92, 23–31. [Google Scholar] [CrossRef]
- Bazan, N.G.; Birkle, D.L.; Reddy, T.S. Docosahexaenoic acid (22:6, n-3) is metabolized to lipoxygenase reaction products in the retina. Biochem. Biophys. Res. Commun. 1984, 125, 741–747. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Marcheselli, V.L.; De Rivero Vaccari, J.C.; Gordon, W.C.; Jackson, F.E.; Bazan, N.G. Photoreceptor outer segment phagocytosis attenuates oxidative stress-induced apoptosis with concomitant neuroprotectin d1 synthesis. Proc. Natl. Acad. Sci. USA 2007, 104, 13158–13163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, C.T.; Bui, B.V.; Sinclair, A.J.; Vingrys, A.J. Dietary omega 3 fatty acids decrease intraocular pressure with age by increasing aqueous outflow. Investig. Ophthalmol. Vis. Sci. 2007, 48, 756–762. [Google Scholar] [CrossRef] [PubMed]
- Schnebelen, C.; Pasquis, B.; Salinas-Navarro, M.; Joffre, C.; Creuzot-Garcher, C.P.; Vidal-Sanz, M.; Bron, A.M.; Bretillon, L.; Acar, N. A dietary combination of omega-3 and omega-6 polyunsaturated fatty acids is more efficient than single supplementations in the prevention of retinal damage induced by elevation of intraocular pressure in rats. Graefe’s Arch. Clin. Exp. Ophthalmol. = Albrecht Graefes Arch. Klin. Exp. Ophthalmol. 2009, 247, 1191–1203. [Google Scholar] [CrossRef] [PubMed]
- Tuo, J.; Ross, R.J.; Herzlich, A.A.; Shen, D.; Ding, X.; Zhou, M.; Coon, S.L.; Hussein, N.; Salem, N., Jr.; Chan, C.C. A high omega-3 fatty acid diet reduces retinal lesions in a murine model of macular degeneration. Am. J. Pathol. 2009, 175, 799–807. [Google Scholar] [CrossRef] [Green Version]
- Sapieha, P.; Chen, J.; Stahl, A.; Seaward, M.R.; Favazza, T.L.; Juan, A.M.; Hatton, C.J.; Joyal, J.S.; Krah, N.M.; Dennison, R.J.; et al. Omega-3 polyunsaturated fatty acids preserve retinal function in type 2 diabetic mice. Nutr. Diabetes 2012, 2, e36. [Google Scholar] [CrossRef]
- Prokopiou, E.; Kolovos, P.; Georgiou, C.; Kalogerou, M.; Potamiti, L.; Sokratous, K.; Kyriacou, K.; Georgiou, T. Omega-3 fatty acids supplementation protects the retina from age-associated degeneration in aged c57bl/6j mice. BMJ Open Ophthalmol. 2019, 4, e000326. [Google Scholar] [CrossRef] [Green Version]
- Sangiovanni, J.P.; Agron, E.; Meleth, A.D.; Reed, G.F.; Sperduto, R.D.; Clemons, T.E.; Chew, E.Y.; Age-Related Eye Disease Study Research Group. {omega}-3 long-chain polyunsaturated fatty acid intake and 12-y incidence of neovascular age-related macular degeneration and central geographic atrophy: Areds report 30, a prospective cohort study from the age-related eye disease study. Am. J. Clin. Nutr. 2009, 90, 1601–1607. [Google Scholar]
- Gerstenblith, A.T.; Baskin, D.E.; Shah, C.P.; Wolfe, J.D.; Fineman, M.S.; Kaiser, R.S.; Ho, A.C. Electroretinographic effects of omega-3 fatty acid supplementation on dry age-related macular degeneration. JAMA Ophthalmol. 2013, 131, 365–369. [Google Scholar] [CrossRef] [Green Version]
- Van Asten, F.; Chiu, C.Y.; Agron, E.; Clemons, T.E.; Ratnapriya, R.; Swaroop, A.; Klein, M.L.; Fan, R.; Chew, E.Y. Age-Related Eye Disease Study 2 Research, G. No cfh or arms2 interaction with omega-3 fatty acids, low versus high zinc, or beta-carotene versus lutein and zeaxanthin on progression of age-related macular degeneration in the age-related eye disease study 2: Age-related eye disease study 2 report no. 18. Ophthalmology 2019, 126, 1541–1548. [Google Scholar]
- Sasaki, M.; Kawasaki, R.; Rogers, S.; Man, R.E.; Itakura, K.; Xie, J.; Flood, V.; Tsubota, K.; Lamoureux, E.; Wang, J.J. The associations of dietary intake of polyunsaturated fatty acids with diabetic retinopathy in well-controlled diabetes. Investig. Ophthalmol. Vis. Sci. 2015, 56, 7473–7479. [Google Scholar] [CrossRef]
- Sala-Vila, A.; Diaz-Lopez, A.; Valls-Pedret, C.; Cofan, M.; Garcia-Layana, A.; Lamuela-Raventos, R.M.; Castaner, O.; Zanon-Moreno, V.; Martinez-Gonzalez, M.A.; Toledo, E.; et al. Dietary marine omega-3 fatty acids and incident sight-threatening retinopathy in middle-aged and older individuals with type 2 diabetes: Prospective investigation from the predimed trial. JAMA Ophthalmol. 2016, 134, 1142–1149. [Google Scholar] [CrossRef] [PubMed]
- Trujillo, J.; Granados-Castro, L.F.; Zazueta, C.; Anderica-Romero, A.C.; Chirino, Y.I.; Pedraza-Chaverri, J. Mitochondria as a target in the therapeutic properties of curcumin. Arch. Pharm. 2014, 347, 873–884. [Google Scholar] [CrossRef]
- Lopez-Malo, D.; Villaron-Casares, C.A.; Alarcon-Jimenez, J.; Miranda, M.; Diaz-Llopis, M.; Romero, F.J.; Villar, V.M. Curcumin as a therapeutic option in retinal diseases. Antioxidants 2020, 9, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, B.M.; Pahlitzsch, M.; Guo, L.; Balendra, S.; Shah, P.; Ravindran, N.; Malaguarnera, G.; Sisa, C.; Shamsher, E.; Hamze, H.; et al. Topical curcumin nanocarriers are neuroprotective in eye disease. Sci. Rep. 2018, 8, 11066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.H.; Ko, Y.C.; Chang, Y.F.; Huang, S.H.; Liu, C.J. Thermosensitive chitosan-gelatin-based hydrogel containing curcumin-loaded nanoparticles and latanoprost as a dual-drug delivery system for glaucoma treatment. Exp. Eye Res. 2019, 179, 179–187. [Google Scholar] [CrossRef]
- Salehi, B.; Stojanovic-Radic, Z.; Matejic, J.; Sharifi-Rad, M.; Anil Kumar, N.V.; Martins, N.; Sharifi-Rad, J. The therapeutic potential of curcumin: A review of clinical trials. Eur. J. Med. Chem. 2019, 163, 527–545. [Google Scholar] [CrossRef]
- Mazzolani, F.; Togni, S. Oral administration of a curcumin-phospholipid delivery system for the treatment of central serous chorioretinopathy: A 12-month follow-up study. Clin. Ophthalmol. 2013, 7, 939–945. [Google Scholar]
- Stohs, S.J.; Chen, O.; Ray, S.D.; Ji, J.; Bucci, L.R.; Preuss, H.G. Highly bioavailable forms of curcumin and promising avenues for curcumin-based research and application: A review. Molecules 2020, 25, 1397. [Google Scholar] [CrossRef] [Green Version]
- Ichi, T.; Higashimura, Y.; Katayama, T.; Koda, T.; Shimizu, T.; Tada, M. Analysis of crocetin derivatives from gardenia fruits. Nippon Shokuhin Kagaku Kogaku Kaishi 1995, 42, 776–783. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Lin, G.; Kwan, Y.W.; Min, Z.D. Simultaneous quantification of five major biologically active ingredients of saffron by high-performance liquid chromatography. J. Chromatogr. A 1999, 849, 349–355. [Google Scholar] [CrossRef]
- Khazdair, M.R.; Boskabady, M.H.; Hosseini, M.; Rezaee, R.A.; Tsatsakis, A.M. The effects of crocus sativus (saffron) and its constituents on nervous system: A review. Avicenna J. Phytomed. 2015, 5, 376–391. [Google Scholar]
- Jose Bagur, M.; Alonso Salinas, G.L.; Jimenez-Monreal, A.M.; Chaouqi, S.; Llorens, S.; Martinez-Tome, M.; Alonso, G.L. Saffron: An old medicinal plant and a potential novel functional food. Molecules 2017, 23, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamauchi, M.; Tsuruma, K.; Imai, S.; Nakanishi, T.; Umigai, N.; Shimazawa, M.; Hara, H. Crocetin prevents retinal degeneration induced by oxidative and endoplasmic reticulum stresses via inhibition of caspase activity. Eur. J. Pharmacol. 2011, 650, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Ishizuka, F.; Shimazawa, M.; Umigai, N.; Ogishima, H.; Nakamura, S.; Tsuruma, K.; Hara, H. Crocetin, a carotenoid derivative, inhibits retinal ischemic damage in mice. Eur. J. Pharmacol. 2013, 703, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Albarral, J.A.; Ramirez, A.I.; De Hoz, R.; Lopez-Villarin, N.; Salobrar-Garcia, E.; Lopez-Cuenca, I.; Licastro, E.; Inarejos-Garcia, A.M.; Almodovar, P.; Pinazo-Duran, M.D.; et al. Neuroprotective and anti-inflammatory effects of a hydrophilic saffron extract in a model of glaucoma. Int. J. Mol. Sci. 2019, 20, 4110. [Google Scholar] [CrossRef] [Green Version]
- Heitmar, R.; Brown, J.; Kyrou, I. Saffron (Crocus sativus L.) in ocular diseases: A narrative review of the existing evidence from clinical studies. Nutrients 2019, 11, 649. [Google Scholar] [CrossRef] [Green Version]
- Piccardi, M.; Fadda, A.; Martelli, F.; Marangoni, D.; Magli, A.; Minnella, A.M.; Bertelli, M.; Di Marco, S.; Bisti, S.; Falsini, B. Antioxidant saffron and central retinal function in abca4-related stargardt macular dystrophy. Nutrients 2019, 11, 2461. [Google Scholar] [CrossRef] [Green Version]
- Eckert, A.; Keil, U.; Scherping, I.; Hauptmann, S.; Muller, W.E. Stabilization of mitochondrial membrane potential and improvement of neuronal energy metabolism by ginkgo biloba extract egb 761. Ann. N.Y. Acad. Sci. 2005, 1056, 474–485. [Google Scholar] [CrossRef]
- Hirooka, K.; Tokuda, M.; Miyamoto, O.; Itano, T.; Baba, T.; Shiraga, F. The ginkgo biloba extract (egb 761) provides a neuroprotective effect on retinal ganglion cells in a rat model of chronic glaucoma. Curr. Eye Res. 2004, 28, 153–157. [Google Scholar] [CrossRef]
- Chung, H.S.; Harris, A.; Kristinsson, J.K.; Ciulla, T.A.; Kagemann, C.; Ritch, R. Ginkgo biloba extract increases ocular blood flow velocity. J. Ocul. Pharmacol. Ther. J. Assoc. Ocul. Pharmacol. Ther. 1999, 15, 233–240. [Google Scholar] [CrossRef]
- Jia, L.Y.; Sun, L.; Fan, D.S.; Lam, D.S.; Pang, C.P.; Yam, G.H. Effect of topical ginkgo biloba extract on steroid-induced changes in the trabecular meshwork and intraocular pressure. Arch. Ophthalmol. 2008, 126, 1700–1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quaranta, L.; Bettelli, S.; Uva, M.G.; Semeraro, F.; Turano, R.; Gandolfo, E. Effect of ginkgo biloba extract on preexisting visual field damage in normal tension glaucoma. Ophthalmology 2003, 110, 359–362. [Google Scholar] [CrossRef]
- Shim, S.H.; Kim, J.M.; Choi, C.Y.; Kim, C.Y.; Park, K.H. Ginkgo biloba extract and bilberry anthocyanins improve visual function in patients with normal tension glaucoma. J. Med. Food 2012, 15, 818–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Kong, X.; Huang, R.; Jin, L.; Ding, X.; He, M.; Liu, X.; Patel, M.C.; Congdon, N.G. Effect of ginkgo biloba on visual field and contrast sensitivity in chinese patients with normal tension glaucoma: A randomized, crossover clinical trial. Investig. Ophthalmol. Vis. Sci. 2014, 55, 110–116. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Ng, E.S.; Kwan, Y.W.; Lau, C.B.; Cheung, D.W.; Koon, J.C.; Zhang, Z.; Zuo, Z.; Leung, P.C.; Fung, K.P.; et al. Cerebral vasodilator properties of danshen and gegen: A study of their combined efficacy and mechanisms of actions. Phytomed. Int. J. Phytother. Phytopharm. 2014, 21, 391–399. [Google Scholar] [CrossRef]
- Yue, K.K.; Lee, K.W.; Chan, K.K.; Leung, K.S.; Leung, A.W.; Cheng, C.H. Danshen prevents the occurrence of oxidative stress in the eye and aorta of diabetic rats without affecting the hyperglycemic state. J. Ethnopharmacol. 2006, 106, 136–141. [Google Scholar] [CrossRef]
- Ma, L.; Tang, L.; Yi, Q. Salvianolic acids: Potential source of natural drugs for the treatment of fibrosis disease and cancer. Front. Pharmacol. 2019, 10, 97. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Liu, Y.Y.; Jiang, Q.; Li, K.R.; Zhao, Y.X.; Cao, C.; Yao, J. Salvianolic acid a protects rpe cells against oxidative stress through activation of nrf2/ho-1 signaling. Free Radic. Biol. Med. 2014, 69, 219–228. [Google Scholar] [CrossRef]
- Mao, K.; Shu, W.; Liu, L.; Gu, Q.; Qiu, Q.; Wu, X. Salvianolic acid a inhibits ox-ldl effects on exacerbating choroidal neovascularization via downregulating cyld. Oxid. Med. Cell Longev. 2017, 2017, 6210694. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Hou, B.; He, P.; Ma, P.; Yang, X.; Yang, X.; Zhang, L.; Qiang, G.; Li, W.; Du, G. Neuroprotective effect of salvianolic acid a against diabetic peripheral neuropathy through modulation of nrf2. Oxid. Med. Cell Longev. 2020, 2020, 6431459. [Google Scholar] [CrossRef]
- Liu, X.; Xavier, C.; Jann, J.; Wu, H. Salvianolic acid b (sal b) protects retinal pigment epithelial cells from oxidative stress-induced cell death by activating glutaredoxin 1 (grx1). Int. J. Mol. Sci. 2016, 17, 1835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, D.; Qin, Y.; Yuan, W.; Deng, H.; Zhang, Y.; Jin, M. Compound danshen dripping pill for treating early diabetic retinopathy: A randomized, double-dummy, double-blind study. Evid. Based Complement. Altern. Med. Ecam. 2015, 2015, 539185. [Google Scholar] [CrossRef] [PubMed]
- Lian, F.; Wu, L.; Tian, J.; Jin, M.; Zhou, S.; Zhao, M.; Wei, L.; Zheng, Y.; Wang, Y.; Zhang, M.; et al. The effectiveness and safety of a danshen-containing chinese herbal medicine for diabetic retinopathy: A randomized, double-blind, placebo-controlled multicenter clinical trial. J. Ethnopharmacol. 2015, 164, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Wang, Z.; Huang, L.; Zheng, S.; Wang, D.; Chen, S.; Zhang, H.; Yang, S. Review of the botanical characteristics, phytochemistry, and pharmacology of astragalus membranaceus (huangqi). Phytother. Res. Ptr. 2014, 28, 1275–1283. [Google Scholar] [CrossRef]
- Aldarmaa, J.; Liu, Z.; Long, J.; Mo, X.; Ma, J.; Liu, J. Anti-convulsant effect and mechanism of astragalus mongholicus extract in vitro and in vivo: Protection against oxidative damage and mitochondrial dysfunction. Neurochem. Res. 2010, 35, 33–41. [Google Scholar] [CrossRef]
- Li, X.T.; Zhang, Y.K.; Kuang, H.X.; Jin, F.X.; Liu, D.W.; Gao, M.B.; Liu, Z.; Xin, X.J. Mitochondrial protection and anti-aging activity of astragalus polysaccharides and their potential mechanism. Int. J. Mol. Sci. 2012, 13, 1747–1761. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.F.; Lu, L.; Zhu, D.J.; Wang, M.; Yin, Y.; Chen, D.X.; Wei, L.B. Effects of astragalus polysaccharides on dysfunction of mitochondrial dynamics induced by oxidative stress. Oxidative Med. Cell Longev. 2016, 2016, 9573291. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Yuan, S.; Liu, X.; Mao, P.; Zhao, C.; Huang, Q.; Zhang, R.; Fang, Y.; Song, Q.; Yuan, D.; et al. Protective effects of astragaloside iv on db/db mice with diabetic retinopathy. PLoS ONE 2014, 9, e112207. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Ke, X.; Ma, N.; Wang, W.; Fu, W.; Zhang, H.; Zhao, M.; Gao, X.; Hao, X.; Zhang, Z. Formononetin, an active compound of astragalus membranaceus (fisch) bunge, inhibits hypoxia-induced retinal neovascularization via the hif-1alpha/vegf signaling pathway. Drug Des. Dev. Ther. 2016, 10, 3071–3081. [Google Scholar] [CrossRef] [Green Version]
- Daliu, P.; Santini, A.; Novellino, E. From pharmaceuticals to nutraceuticals: Bridging disease prevention and management. Expert Rev. Clin. Pharmacol. 2019, 12, 1–7. [Google Scholar] [CrossRef]
Figure 1.
Different pathogenetic mechanisms lead to ocular diseases. Arrow indicates pleiotropic factors, as described above, and how they interact with the retinal cells that contribute to eye diseases.
Figure 1.
Different pathogenetic mechanisms lead to ocular diseases. Arrow indicates pleiotropic factors, as described above, and how they interact with the retinal cells that contribute to eye diseases.


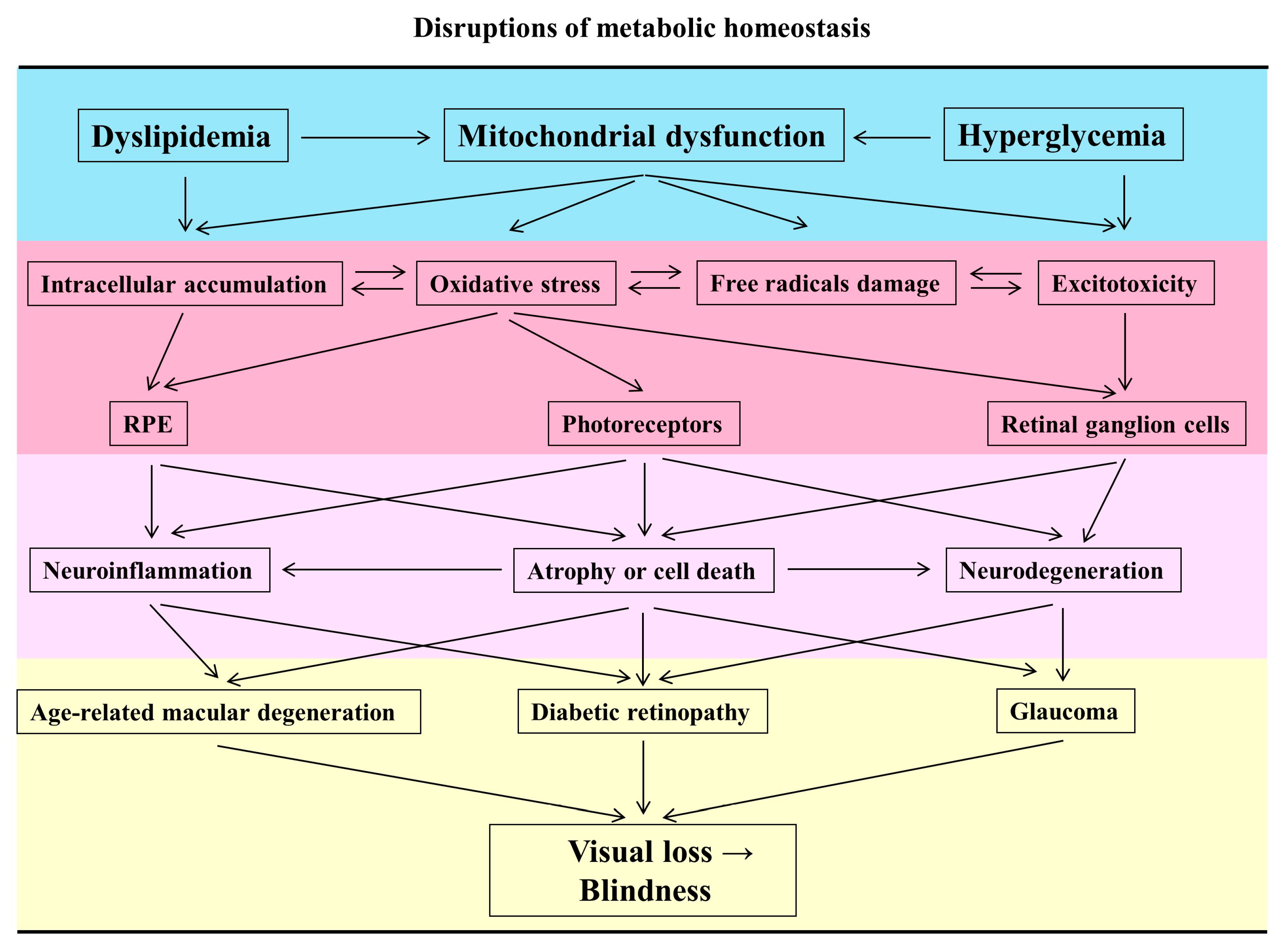{kind=link}
Table 1.
Mechanisms of ocular neuroprotection by nutraceuticals in animal models.
| Diseases | Nutraceutical | Effects | Mechanisms | Animal Models | Refs. |
|---|---|---|---|---|---|
| Glaucoma | Resveratrol | Neuroprotection | Apoptosis ↓ | Intracameral injection of hyaluronic acid-induced rats | [71] |
| Resveratrol | Neuroprotection | BiP↑, CHOP↑, XBP-1↓ | Optic nerve crush experimental mice | [72] | |
| Resveratrol | IOP reduction | Binding through A1R | Steroid-induced ocular hypertension rats | [73] | |
| Resveratrol and quercetin | IOP reduction | Synergic effects | Normal normotensive rabbits | [74] | |
| AMD | Resveratrol | Prevent retinal degeneration | ONL↑, ERG↑, Apoptosis↓, AP-1↓, SIRT1↑ | Light-induced retinal degeneration mice | [75] |
| Resveratrol | Neuroprotection | ONL↑, ERG↑, LIF, BDNF, OSM, CT-1 and CLC↑ | Light-induced retinal degeneration mice | [76] | |
| DR | Resveratrol | Alleviate oxidative stress | BG↓, BW↑, SOD↑, 8-Isoprostane↓, GSSG/GSH↓, NF-κB↓, Apoptosis↓, ONL↑ | STZ-nicotinamide-induced DR rats | [77] |
| Resveratrol | Anti-oxidative stress | AOPP↓, MDA↓, TOS↓ | STZ-induced type 1 diabetes rats | [78] | |
| Resveratrol | Anti-inflammation and retinal protection | BG↓, BW↑, AGEs↓, Insulin↑, Apoptosis↓, PON1↑, Ox-LDL↓, IL-1β, IL-6, TNF-α, VEGF, IFN-γ and MCP-1↓ | STZ-induced diabetes rats | [79] | |
| Resveratrol | Retinal vascular protection | BG↓, Pericytes↑ | STZ-induced diabetes mice | [80] | |
| Glaucoma | Quercetin | Neuroprotection | ERG↑, RGC survival↑, Apoptosis↓, ΔΨm↑ | Chronic ocular hypertension rats | [93] |
| Quercetin | Neuroprotection | GABAergic inhibitory neurotransmission↑, glutamatergic excitatory neurotransmission↓, excitability of the RGCs↓ | Electrocoagulation of the superior scleral vein rats | [94] | |
| AMD | Quercetin | Neuroprotection | Thickness of whole retina↑, Apoptosis↓, Inflammation↓ | Blue light-induced damage mice | [98] |
| Quercetin | Neuroprotection | ERG↑, ONL↑, Phagosomes in RPE↑, AP-1↓ | Light-induced retinal degeneration rats | [99] | |
| DR | Quercetin | Neuroprotection | GSH↑, SOD↑, Thickness of whole retina↑, ONL↑, INL↑, TNF-α↓, IL-1β↓, Apoptosis↓, AQP4, GFAP and caspase-3↓ | STZ-induced diabetes rats | [102] |
| Lutein or DHA | Neuroprotection | Thickness of whole retina↑, ONL↑, INL↑, Apoptosis↓ | STZ-induced diabetes rats | [112] | |
| Retinal detachment (RD) | Lutein | Neuroprotection | ONL↑, GFAP↓, RHO↑, Apoptosis↓ | Subretinal injections-induced RD rats | [115] |
| AMD | Lutein | Anti-oxidative stress | RPE tight junctions↑, ROS↓, SOD↑, Macrophage-related markers↓ | Light-induced AMD-related mice | [116] |
| Lutein and zeaxanthin | Neuroprotection | ERG↑, Apoptosis↓, p-JNK↓, Nrf2↑, GRP78, p-PERK, ATF4 and ATF6↓ | Light-induced damage mice | [117] | |
| DR | Wolfberry | Retinoprotection | Levels of zeaxanthin and lutein↑, SRB1↑, GSTP1, BCO2, and AMPK-α2↑, HIF-1α, VEGF, and HSP↓, Mitochondrial copy number↑, Citrate synthase activity↑, PGC-1α, Nrf1, and TFAM↑ | Leptin receptor-deficient (db/db) type 2 diabetic mice | [118] |
| Glaucoma | Omega-3 PUFAs | IOP reduction | IOP↓, Aqueous outflow↑ | Age-induced IOP increase rats | [132] |
| Omega-3 and omega-6 PUFAs | Anti-inflammation | GFAP↓, Thickness of whole retina↑ | Photocoagulation-induced IOP increase rats | [133] | |
| AMD | Omega-3 PUFAs | Anti-inflammation | PGE2, LTB4, TNF-α and IL-6↓, PGD2↑ | AMD-like retinal lesions mice | [134] |
| DR | Omega-3 PUFAs | Retinoprotection | ERG↑, BG↓ | Leptin receptor-deficient (db/db) type 2 diabetic mice | [135] |
| AMD | Omega-3 PUFAs | Retinoprotection | Lipofuscin↓, ONL↑, MBP, MPP, MRFLP and GFAP↑ | Aged (24-month-old) wild-type mice | [136] |
| Glaucoma | Curcumin | Neuroprotection | IOP↓, RGC density↑ | Ocular hypertension and partial optic nerve transection rats | [144] |
| Retinal degeneration | Crocetin | Neuroprotection | ERG↑, ONL↑, Apoptosis↓ | Light-induced damage mice | [153] |
| Retinal ischemia | Crocetin | Neuroprotection | GCL, INL, ONL↑, ERG↑, p-p38, p-JNK, p- ERK 1/2, p-c-Jun, p-NF-κB↓ | I/R-induced retinal damage mice | [154] |
| Glaucoma | Saffron | Neuroprotection | IOP↓, RGC↑, Iba-1 (+) microglia↓ | Laser-induced ocular hypertension mice | [155] |
| GBE | Neuroprotection | IOP↓, RGC↑, | Ocular hypertension by cautery of three episcleral vessels rats | [159] | |
| GBE | Retinoprotection | IOP↓, Apoptotic TM cells↓ | Dexamethasone–induced ocular hypertension rabbits | [161] | |
| Diabetes | Danshen | Anti-oxidative stress | GSH↑, MDA↓ | STZ-induced diabetes rats | [166] |
| CNV | Salvianolic acid A | Anti-angiogenesis | OX-LDL↓, Fluorescein angiography↓, VEGF↓, PDGF↓, Angiostatin↑, CYLD↓ | Laser photocoagulations plus OX-LDL injection-induced CNV mice | [169] |
| DPN | Salvianolic acid A | Anti-oxidative stress | BG↓, Fructosamine↓, Myelin sheath thickness↑, Nrf2↑ | KK-Ay diabetic mice | [170] |
| DR | Astragaloside IV | Neuroprotection | ERG↑, Apoptosis of RGCs↓, AR↓, p-ERK1/2, NF-kB↓ | Leptin receptor-deficient (db/db) type 2 diabetic mice | [178] |
Increase (↑), Decrease (↓), Binding immunoglobulin protein (BiP), C/EBP homologous protein (CHOP), X-box binding protein-1 (XBP-1), Adenosine receptor 1 (A1R), Optic nerve layer (ONL), Streptozotocin (STZ), Blood glucose (BG), Body weight (BW), Glycosylated hemoglobin (HbA1c), Oxidized glutathione (GSSG), Advanced oxidation protein products (AOPP), Malondialdehyde (MDA), Total Oxidant Status (TOS), Low-density lipoprotein (LDL), Interleukin (IL), Tumor necrosis factor (TNF), Interferon (IFN), Monocyte chemotactic protein (MCP), Inner nuclear layer (INL), Aquaporin-4 (AQP4), rhodopsin (RHO), phosphorylated c-Jun N-terminal kinase (p-JNK), Glucose-regulated protein (GRP78), phosphorylated protein kinase RNA-like endoplasmic reticulum kinase (p-PERK), activating transcription factor 4 (ATF4), activating transcription factor (ATF6), Scavenger receptor class B type 1 (SRB1), Glutathione S-transferase pi gene (GSTP1), Hypoxia-inducible factor-1-α (HIF-1α), Nuclear respiratory factor 1 (NRF1), Transcription factor A, mitochondrial (TFAM), Prostaglandin (PG), Leukotriene B4 (LTB4), Oxidized low-density lipoprotein (OX-LDL), Platelet-derived growth factor (PDGF).
Table 2.
Main significant outcomes of nutraceuticals in clinical trials.
| Diseases | Nutraceutical | Study Population | Study Design/Follow-Up | Results/Findings | Refs. |
|---|---|---|---|---|---|
| AMD | Resveratrol | 3 cases | Case report | Restoration of structure and visual function | [85] |
| T2D | Resveratrol | 17 subjects | Double-blind randomized cross-over study | Intrahepatic lipid↓, Intramyocellular lipid↑, Mitochondrial function (ex vivo) ↑, Metformin dose↓ | [86] |
| AMD | Quercetin | 2856 adults and 2037 followed | Population-based cohort study/15-y | Quercetin was associated with reduced odds of any AMD (OR: 0.76; 95% CI: 0.58–0.99) | [108] |
| AMD | Lutein/zeaxanthin | 93,676 women | Cohort study/7-y | Lutein/zeaxanthin may protect against intermediate AMD (OR: 0.57; 95% CI: 0.34–0.95) | [121] |
| AMD | Analysis of Lutein/zeaxanthin | 380 adults | Cohort study | Risk of AMD was associated with plasma concentrations of lutein/zeaxanthin (OR: 1.9; 95% CI: 0.9–3.5) | [122] |
| AMD | Lutein/zeaxanthin | 6 publications | Meta-analysis | Dietary intake of lutein/zeaxanthin was significantly related with a reduction in risk of late AMD (RR: 0.74; 95% CI: 0.57–0.97) | [123] |
| AMD | Lutein/zeaxanthin, Omega-3, PUFAs, Zinc | 4203 participants | multicenter, randomized, double-masked, placebo-controlled phase 3 study with a 2 × 2 factorial design/12-y | Participants were limited to those with the lowest dietary intake of lutein + zeaxanthin, results of exploratory subgroup analyses showed a protective effect for progression to advanced AMD (HR: 0.74; 95% CI, 0.59–0.94; p = 0.01) | [125] |
| AMD | Omega-3 PUFAs | 1837 participants | Nested cohort study/12-y | Omega-3 PUFAs intake reduces 30% incidence to develop CGA (OR: 0.65; 95% CI: 0.45–0.92; p < 0.02) and neovascular AMD (OR: 0.68; 95% CI:0.49–0.94; p < 0.02) | [137] |
| AMD | Omega-3 PUFAs | 17 patients | Prospective, noncomparative, descriptive pilot study | No statistically significant improvement in visual acuity and ERG; Serum omega-3 index increased by a mean of 7.6% (p < 0.001) | [138] |
| AMD | Lutein/zeaxanthin, Omega-3 PUFAs, Zinc, β-carotene | 1684 participants | AREDS2/5-y | No significant interaction between supplements and genotype with improvement of progression to late AMD | [139] |
| DR | PUFAs | 379 patients | Cohort study | PUFAs was associated with a reduction in DR severity (OR: 0.18; 95% CI: 0.06–0.59) | [140] |
| DR | Omega-3 PUFAs | 3482 participants | Prospective, randomized clinical trial | Participants meeting the LCω3PUFA recommendation at baseline (≥500 mg/d) compared with those not fulfilling this recommendation (<500 mg/d) showed a 48% relatively reduced risk of incident sight-threatening DR, with a HR of 0.52 (95% CI, 0.31–0.88; p = 0.001). This association was slightly stronger for yearly updated LCω3PUFA intake (relative risk, 0.48; 95% CI, 0.28–0.82; p = 0.007) | [141] |
| CSCR | Curcuminoids and lecithin formulation | 12 patients | Follow-up study/1-y | Visual acuity improvement (p = 0.0005 by Wilcoxon signed rank test), Reduction in neuroretinal or RPE detachment (p = 0.0004 by Wilcoxon signed rank test) | [147] |
| STG/FF | Saffron | 31 patients | Randomized, double-blind, placebo-controlled study/3-y | Saffron had no detrimental effects on the visual acuity and focal ERG | [157] |
| NTG | GBE | 27 patients | Prospective, randomized, placebo-controlled, double-masked cross-over trial | Visual fields improvement in mean deviation (t = 8.86, p = 0.0001, chi-square test) and corrected pattern standard deviation (t = 9.89, p = 0.0001, chi-square test) | [162] |
| NTG | GBE | 332 patients | Retrospective study/1-y or more | HVF mean deviation improvement from −5.25 ± 6.13 to −4.31 ± 5.60 (p = 0.002) | [163] |
| NTG | GBE | 35 patients | Prospective, randomized, placebo-controlled crossover study | No effect on mean defect or contrast sensitivity | [164] |
| NPDR | CDDP | 57 patients | Randomized, double-dummy, double-blind study | Improvement of the best corrected visual acuity (p < 0.05). | [172] |
| NPDR | CDDP | 223 patients | Randomized, double-blind, placebo-controlled clinical trial | Improvement of fluorescence fundus angiography and funduscopic examination (p < 0.001) | [173] |
Odds ratios (OR), Confidence interval (CI), Relative risk (RR), Hazard ratio (HR).
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Huang, C.-P.; Lin, Y.-W.; Huang, Y.-C.; Tsai, F.-J. Mitochondrial Dysfunction as a Novel Target for Neuroprotective Nutraceuticals in Ocular Diseases. Nutrients 2020, 12, 1950. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12071950
AMA Style
Huang C-P, Lin Y-W, Huang Y-C, Tsai F-J. Mitochondrial Dysfunction as a Novel Target for Neuroprotective Nutraceuticals in Ocular Diseases. Nutrients. 2020; 12(7):1950. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12071950
Chicago/Turabian StyleHuang, Chun-Ping, Yi-Wen Lin, Yu-Chuen Huang, and Fuu-Jen Tsai. 2020. "Mitochondrial Dysfunction as a Novel Target for Neuroprotective Nutraceuticals in Ocular Diseases" Nutrients 12, no. 7: 1950. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12071950
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.