The Combination of Soy Isoflavones and Resveratrol Preserve Bone Mineral Density in Hindlimb-Unloaded Mice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Diets, and Experimental Design
2.2. Microcomputed Tomography (μCT) Analysis of the Distal Femur
2.3. Bone Histological Stain Analysis
2.4. Analysis of Plasma Albumin and Total Protein Concentrations
2.5. RNA Extraction and Quantitative Real-time PCR
2.6. Statistical Analysis
3. Results
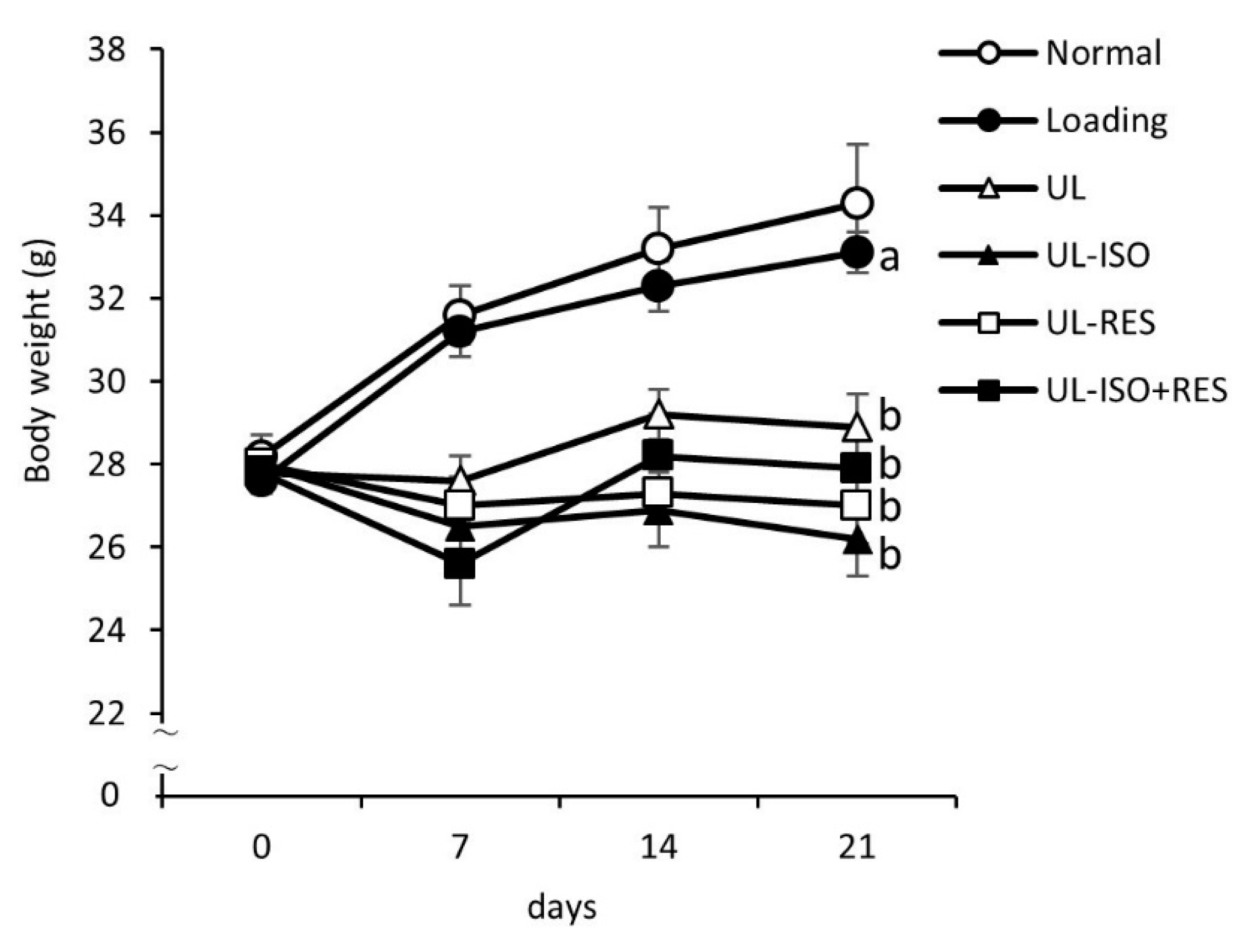
3.1. Body Weight, Food Intake, and Plasma Albumin and Total Protein Concentrations
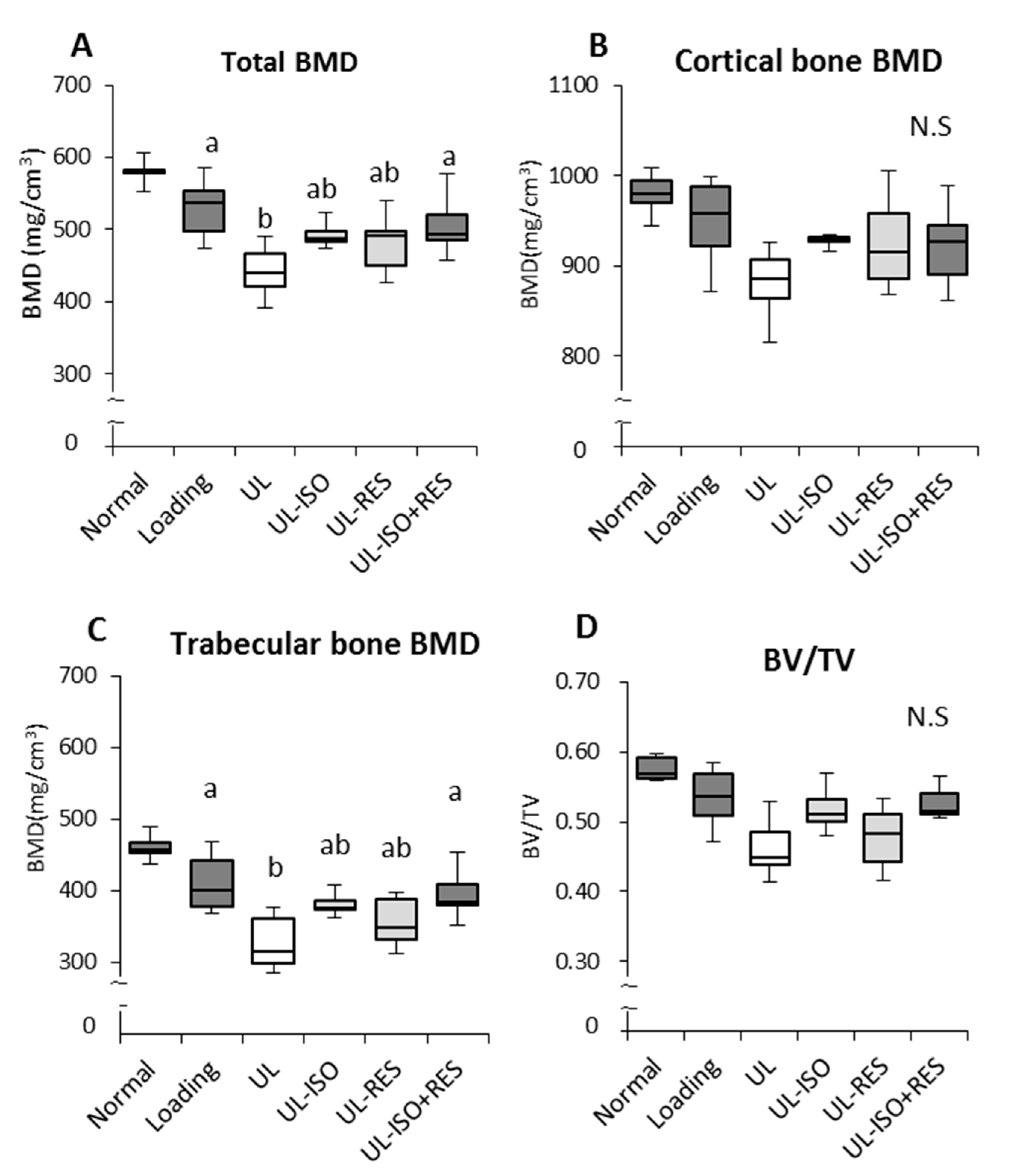
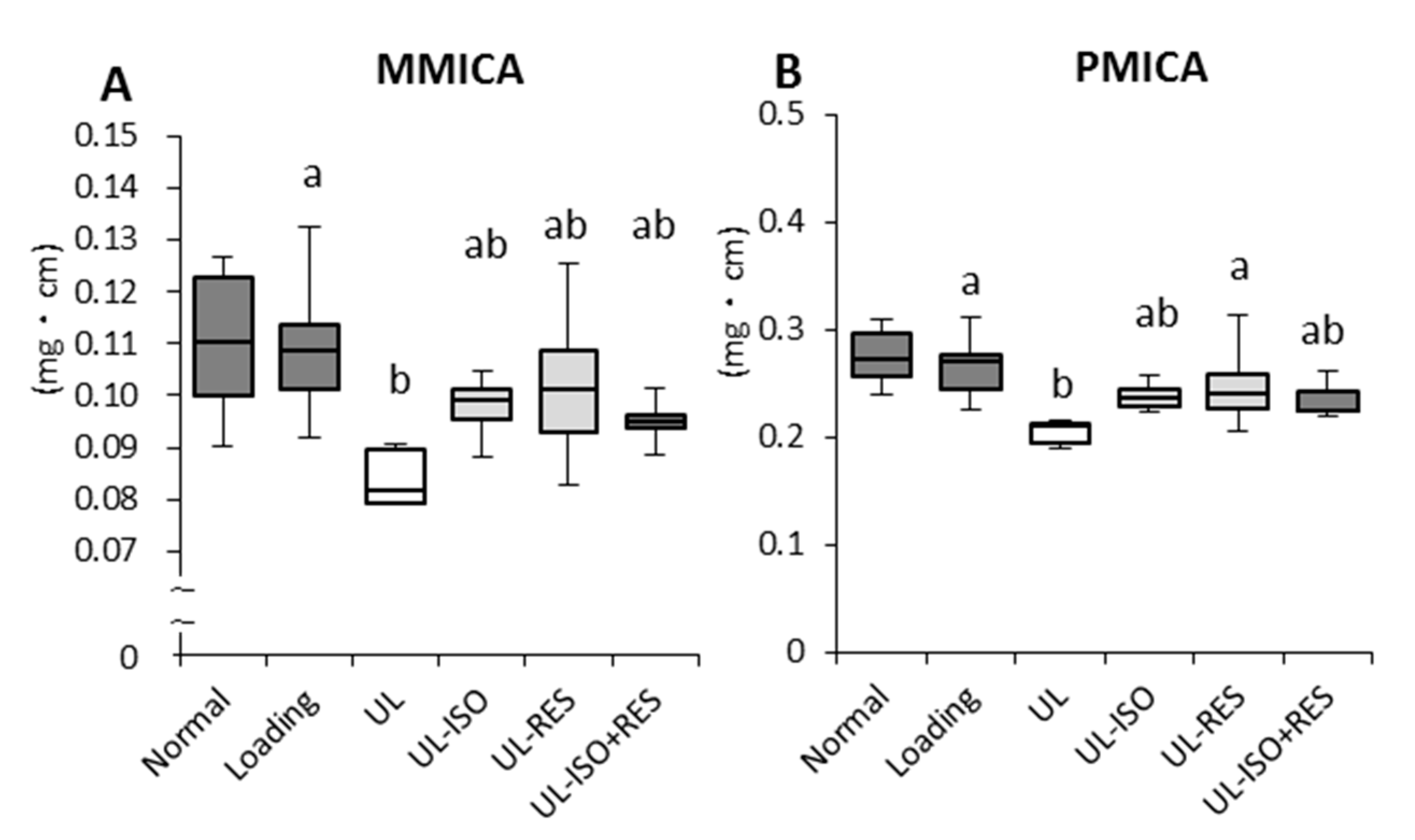
3.2. Femur BMD and Bone Strength by uCT
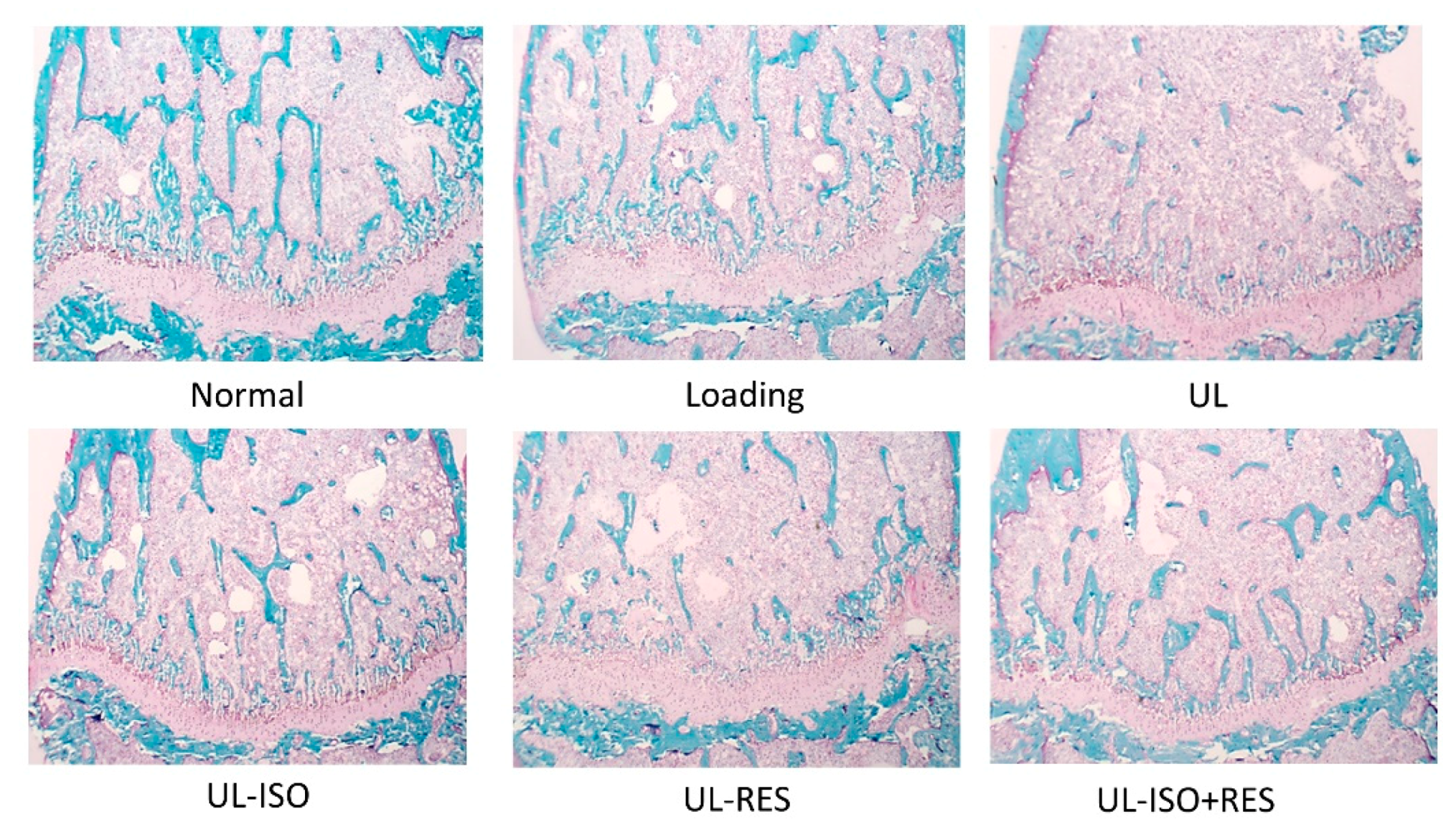
3.3. Bone Histological Stain Analysis
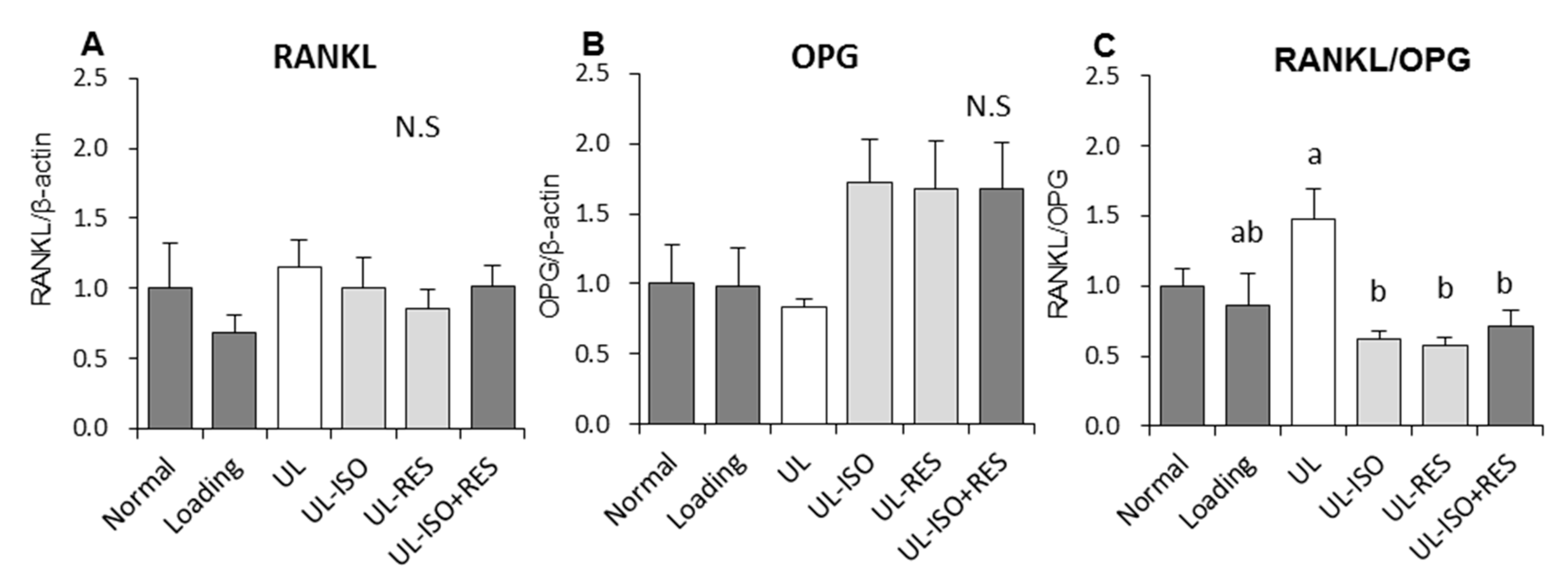
3.4. Quantitation of mRNA Expression in Bone Marrow Cells from the Tibia
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bikle, D.D.; Harris, J.; Halloran, B.P.; Morey-Holton, E. Altered skeletal pattern of gene expression in response to spaceflight and hindlimb elevation. Am. J. Physiol. 1994, 267, 822–827. [Google Scholar] [CrossRef] [PubMed]
- Globus, R.K.; Bikle, D.D.; Morey-Holton, E. The temporal response of bone to unloading. Endocrinology 1986, 118, 733–742. [Google Scholar] [CrossRef] [Green Version]
- Leblanc, A.D.; Schneider, V.S.; Evans, H.J.; Engelbretson, D.A.; Krebs, J.M. Bone mineral loss and recovery after 17 weeks of bed rest. J. Bone Miner. Res. 1990, 5, 843–850. [Google Scholar] [CrossRef]
- Sheridan, K.J. Osteoporosis in adults with cerebral palsy. Dev. Med. Child Neurol. 2009, 51 (Suppl. 4), 38–51. [Google Scholar] [CrossRef]
- Vico, L.; Collet, P.; Guignandon, A.; Lafage-Proust, M.H.; Thomas, T.; Rehaillia, M.; Alexandre, C. Effects of long-term microgravity exposure on cancellous and cortical weight-bearing bones of cosmonauts. Lancet 2000, 355, 1607–1611. [Google Scholar] [CrossRef]
- LeBlanc, A.; Schneider, V.; Shackelford, L.; West, S.; Oganov, V.; Bakulin, A.; Voronin, L. Bone mineral and lean tissue loss after long duration space flight. J. Musculoskelet. Neuronal. Interact. 2000, 1, 157–160. [Google Scholar] [PubMed]
- Morey-Holton, E.R.; Globus, R.K. Hindlimb unloading rodent model: Technical aspects. J. Appl. Physiol. 2002, 92, 1367–1377. [Google Scholar] [CrossRef] [PubMed]
- Halloran, B.P.; Bikle, D.D.; Wronski, T.J.; Globus, R.K.; Levens, M.J.; Morey-Holton, E. The role of 1,25-dihydroxyvitamin D in the inhibition of bone formation induced by skeletal unloading. Endocrinology 1986, 118, 948–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurokouchi, K.; Ito, T.; Ohmori, S.; Kanda, K.; Murata, Y.; Seo, H. Changes in the markers of bone metabolism following skeletal unloading. Environ. Med. 1995, 39, 21–24. [Google Scholar]
- Ishijima, M.; Rittling, S.R.; Yamashita, T.; Tsuji, K.; Kurosawa, H.; Nifuji, A.; Denhardt, D.T.; Noda, M. Enhancement of osteoclastic bone resorption and suppression of osteoblastic bone formation in response to reduced mechanical stress do not occur in the absence of osteopontin. J. Exp. Med. 2001, 193, 399–404. [Google Scholar] [CrossRef] [Green Version]
- Smith, B.J.; King, J.B.; Lucas, E.A.; Akhter, M.P.; Arjmandi, B.H.; Stoecker, B.J. Skeletal unloading and dietary copper depletion are detrimental to bone quality of mature rats. J. Nutr. 2002, 132, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Bikle, D.D.; Morey-Holton, E.R.; Doty, S.B.; Currier, P.A.; Tanner, S.J.; Halloran, B.P. Alendronate increases skeletal mass of growing rats during unloading by inhibiting resorption of calcified cartilage. J. Bone Miner. Res. 1994, 9, 1777–1787. [Google Scholar] [CrossRef] [PubMed]
- Kodama, Y.; Nakayama, K.; Fuse, H.; Fukumoto, S.; Kawahara, H.; Takahashi, H.; Kurokawa, T.; Sekiguchi, C.; Nakamura, T.; Matsumoto, T. Inhibition of bone resorption by pamidronate cannot restore normal gain in cortical bone mass and strength in tail-suspended rapidly growing rats. J. Bone Miner. Res. 1997, 12, 1058–1067. [Google Scholar] [CrossRef] [PubMed]
- Momken, I.; Stevens, L.; Bergouignan, A.; Desplanches, D.; Rudwill, F.; Chery, I.; Zahariev, A.; Zahn, S.; Stein, T.P.; Sebedio, J.L.; et al. Resveratrol prevents the wasting disorders of mechanical unloading by acting as a physical exercise mimetic in the rat. FASEB J. 2011, 25, 3646–3660. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, Y.; Tousen, Y.; Ishimi, Y. Beta-Carotene prevents bone loss in hind limb unloading mice. J. Clin. Biochem. Nutr. 2018, 63, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Sugiyama, F.; Wu, J.; Fujioka, M.; Ezaki, J.; Takeda, K.; Miyaura, C.; Ishida, T.; Yamada, K.; Ishimi, Y. Soybean isoflavones preserve bone mass in hindlimb-unloaded mice. J. Bone Miner. Metab. 2006, 24, 439–446. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Tousen, Y.; Nishide, Y.; Tadaishi, M.; Kato, K.; Ishimi, Y. Combined effects of soy isoflavones and milk basic protein on bone mineral density in hind-limb unloaded mice. J. Clin. Biochem. Nutr. 2016, 58, 141–145. [Google Scholar] [CrossRef] [Green Version]
- Kostelac, D.; Rechkemmer, G.; Briviba, K. Phytoestrogens modulate binding response of estrogen receptors alpha and beta to the estrogen response element. J. Agric. Food Chem. 2003, 51, 7632–7635. [Google Scholar] [CrossRef]
- Ishimi, Y.; Miyaura, C.; Ohmura, M.; Onoe, Y.; Sato, T.; Uchiyama, Y.; Ito, M.; Wang, X.; Suda, T.; Ikegami, S. Selective effects of genistein, a soybean isoflavone, on B-lymphopoiesis and bone loss caused by estrogen deficiency. Endocrinology 1999, 140, 1893–1900. [Google Scholar] [CrossRef]
- de la Lastra, C.A.; Villegas, I. Resveratrol as an anti-inflammatory and anti-aging agent: Mechanisms and clinical implications. Mol. Nutr. Food Res. 2005, 49, 405–430. [Google Scholar] [CrossRef]
- Tseng, P.C.; Hou, S.M.; Chen, R.J.; Peng, H.W.; Hsieh, C.F.; Kuo, M.L.; Yen, M.L. Resveratrol promotes osteogenesis of human mesenchymal stem cells by upregulating RUNX2 gene expression via the SIRT1/FOXO3A axis. J. Bone Miner. Res. 2011, 26, 2552–2563. [Google Scholar] [CrossRef]
- Zhao, H.; Li, X.; Li, N.; Liu, T.; Liu, J.; Li, Z.; Xiao, H.; Li, J. Long-term resveratrol treatment prevents ovariectomy-induced osteopenia in rats without hyperplastic effects on the uterus. Br. J. Nutr. 2014, 111, 836–846. [Google Scholar] [CrossRef] [Green Version]
- Shakibaei, M.; Buhrmann, C.; Mobasheri, A. Resveratrol-mediated SIRT-1 interactions with p300 modulate receptor activator of NF-kappaB ligand (RANKL) activation of NF-kappaB signaling and inhibit osteoclastogenesis in bone-derived cells. J. Biol. Chem. 2011, 286, 11492–11505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.; Andersson, G.; Lindgren, U.; Li, Y. Resveratrol prevents RANKL-induced osteoclast differentiation of murine osteoclast progenitor RAW 264.7 cells through inhibition of ROS production. Biochem. Biophys. Res. Commun. 2010, 401, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; Huang, Y.M.; Xiao, B.X.; Ren, G.F. Effects of resveratrol on bone mineral density in ovarectomized rats. Int. J. Biomed. Sci. 2005, 1, 76–81. [Google Scholar] [PubMed]
- Durbin, S.M.; Jackson, J.R.; Ryan, M.J.; Gigliotti, J.C.; Alway, S.E.; Tou, J.C. Resveratrol supplementation influences bone properties in the tibia of hindlimb-suspended mature Fisher 344 x Brown Norway male rats. Appl. Physiol. Nutr. Metab. 2012, 37, 1179–1188. [Google Scholar] [CrossRef]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C., Jr. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef]
- Tousen, Y.; Uehara, M.; Kruger, M.C.; Ishimi, Y. Effects of dietary fibre and tea catechin, ingredients of the Japanese diet, on equol production and bone mineral density in isoflavone-treated ovariectomised mice. J. Nutr. Sci. 2012, 1, e13. [Google Scholar] [CrossRef]
- Habold, C.; Momken, I.; Ouadi, A.; Bekaert, V.; Brasse, D. Effect of prior treatment with resveratrol on density and structure of rat long bones under tail-suspension. J. Bone Miner. Metab. 2011, 29, 15–22. [Google Scholar] [CrossRef]
- Durbin, S.M.; Jackson, J.R.; Ryan, M.J.; Gigliotti, J.C.; Alway, S.E.; Tou, J.C. Resveratrol supplementation preserves long bone mass, microstructure, and strength in hindlimb-suspended old male rats. J. Bone Miner. Metab. 2014, 32, 38–47. [Google Scholar] [CrossRef]
- Bitto, A.; Burnett, B.P.; Polito, F.; Marini, H.; Levy, R.M.; Armbruster, M.A.; Minutoli, L.; Di Stefano, V.; Irrera, N.; Antoci, S.; et al. Effects of genistein aglycone in osteoporotic, ovariectomized rats: A comparison with alendronate, raloxifene and oestradiol. Br. J. Pharmacol. 2008, 155, 896–905. [Google Scholar] [CrossRef] [PubMed]
- Dai, R.; Ma, Y.; Sheng, Z.; Jin, Y.; Zhang, Y.; Fang, L.; Fan, H.; Liao, E. Effects of genistein on vertebral trabecular bone microstructure, bone mineral density, microcracks, osteocyte density, and bone strength in ovariectomized rats. J. Bone Miner. Metab. 2008, 26, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Teitelbaum, S.L. Bone resorption by osteoclasts. Science 2000, 289, 1504–1508. [Google Scholar] [CrossRef]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Wang, X.X.; Takasaki, M.; Ohta, A.; Higuchi, M.; Ishimi, Y. Cooperative effects of exercise training and genistein administration on bone mass in ovariectomized mice. J. Bone Miner. Res. 2001, 16, 1829–1836. [Google Scholar] [CrossRef]
- Morabito, N.; Crisafulli, A.; Vergara, C.; Gaudio, A.; Lasco, A.; Frisina, N.; D’Anna, R.; Corrado, F.; Pizzoleo, M.A.; Cincotta, M.; et al. Effects of Genistein and Hormone-Replacement Therapy on Bone Loss in Early Postmenopausal Women: A Randomized Double-Blind Placebo-Controlled Study. J. Bone Miner. Res. 2002, 17, 1904–1912. [Google Scholar] [CrossRef]
- Ishimi, Y.; Arai, N.; Wang, X.; Wu, J.; Umegaki, K.; Miyaura, C.; Takeda, A.; Ikegami, S. Difference in effective dosage of genistein on bone and uterus in ovariectomized mice. Biochem. Biophys. Res. Commun. 2000, 11, 697–701. [Google Scholar] [CrossRef]
- Picherit, C.; Chanteranne, B.; Bennetau-Pelissero, C.; Davicco, M.J.; Lebecque, P.; Barlet, J.P.; Coxam, V. Dose-dependent Bone-Sparing Effects of Dietary Isoflavones in the Ovariectomised Rat. Br. J. Nutr. 2001, 85, 307–316. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient (g) | Normal b, Loading b | ISO c | RES d | ISO + RES e |
|---|---|---|---|---|
| Cornstarch | 529.5 | 525.5 | 528.0 | 524.0 |
| Casein | 200 | 200 | 200 | 200 |
| Sucrose | 100 | 100 | 100 | 100 |
| Corn oil | 70 | 70 | 70.0 | 70 |
| Cellulose | 50 | 50 | 50.0 | 50 |
| Mineral mixture a | 35.0 | 35.0 | 35.0 | 35.0 |
| Vitamin mixture a | 10.0 | 10.0 | 10.0 | 10.0 |
| L-Cystine | 3.00 | 3.00 | 3.00 | 3.00 |
| Choline bitartrate | 2.50 | 2.50 | 2.50 | 2.50 |
| Tert-Butylhydroquinone | 0.014 | 0.014 | 0.014 | 0.014 |
| Isoflavones f | - | 4.00 | - | 4.00 |
| Resveratrol g | - | 1.50 | 1.50 | |
| Total | 1000 | 1000 | 1000 | 1000 |
| Normal | Loading | UL | UL- ISO | UL- RES | UL- ISO+RES | p | |
|---|---|---|---|---|---|---|---|
| Body weight | |||||||
| Initial body weight (g) | 28.2 ± 0.5 | 27.6 ± 0.3 | 27.8 ± 0.4 | 28.0 ± 0.4 | 28.0 ± 0.4 | 27.8 ± 0.4 | 0.925 |
| Final body weight (g) | 31.9 ± 1.5 | 31.2 ± 0.5 a | 27.0 ± 0.7 b | 24.7 ± 0.8 b | 25.2 ± 0.8 b | 26.1 ± 0.7 b | < 0.01 |
| Total food intake (g) | 88.4 ± 0.7 | 88.8 ± 1.4 | 87.8 ± 1.5 | 85.2 ± 1.2 | 86.3 ± 1.7 | 84.0 ± 1.9 | 0.200 |
| Plasma | |||||||
| Total protein (g/dL) | 4.19 ± 0.15 | 4.45 ± 0.10 | 4.62 ± 0.17 | 4.47 ± 0.14 | 4.61 ± 0.09 | 4.59 ± 0.05 | 0.733 |
| Albumin (g/dL) | 2.48 ± 0.10 | 2.74 ± 0.12 | 2.59 ± 0.11 | 2.57 ± 0.12 | 2.87 ± 0.07 | 2.84 ± 0.05 | 0.091 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tousen, Y.; Ichimaru, R.; Kondo, T.; Inada, M.; Miyaura, C.; Ishimi, Y. The Combination of Soy Isoflavones and Resveratrol Preserve Bone Mineral Density in Hindlimb-Unloaded Mice. Nutrients 2020, 12, 2043. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12072043
Tousen Y, Ichimaru R, Kondo T, Inada M, Miyaura C, Ishimi Y. The Combination of Soy Isoflavones and Resveratrol Preserve Bone Mineral Density in Hindlimb-Unloaded Mice. Nutrients. 2020; 12(7):2043. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12072043
Chicago/Turabian StyleTousen, Yuko, Ryota Ichimaru, Takashi Kondo, Masaki Inada, Chisato Miyaura, and Yoshiko Ishimi. 2020. "The Combination of Soy Isoflavones and Resveratrol Preserve Bone Mineral Density in Hindlimb-Unloaded Mice" Nutrients 12, no. 7: 2043. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12072043