Gastrointestinally Digested Protein from the Insect Alphitobius diaperinus Stimulates a Different Intestinal Secretome than Beef or Almond, Producing a Differential Response in Food Intake in Rats

 , , ,
, , , 
 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. In Vitro Digestion
2.2.1. Gastric Digestion
2.2.2. Intestinal Digestion
2.3. Characterization of Digestion Products
2.4. Tissue Collection
2.4.1. Pig Tissue
2.4.2. Human Tissue
2.5. Ex Vivo Experiments with Intestinal Segments
2.6. Food Intake Study in Rats
2.7. Statistical Analysis
3. Results
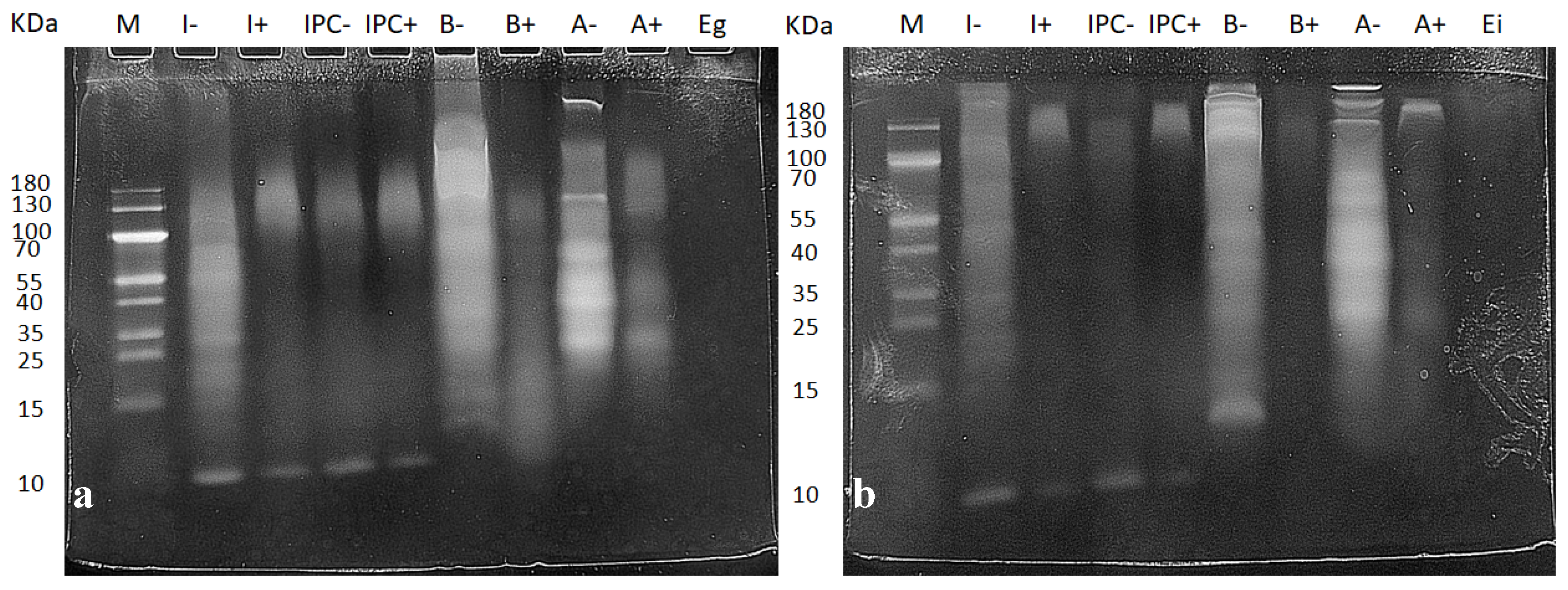
3.1. Characterization of Digested Samples
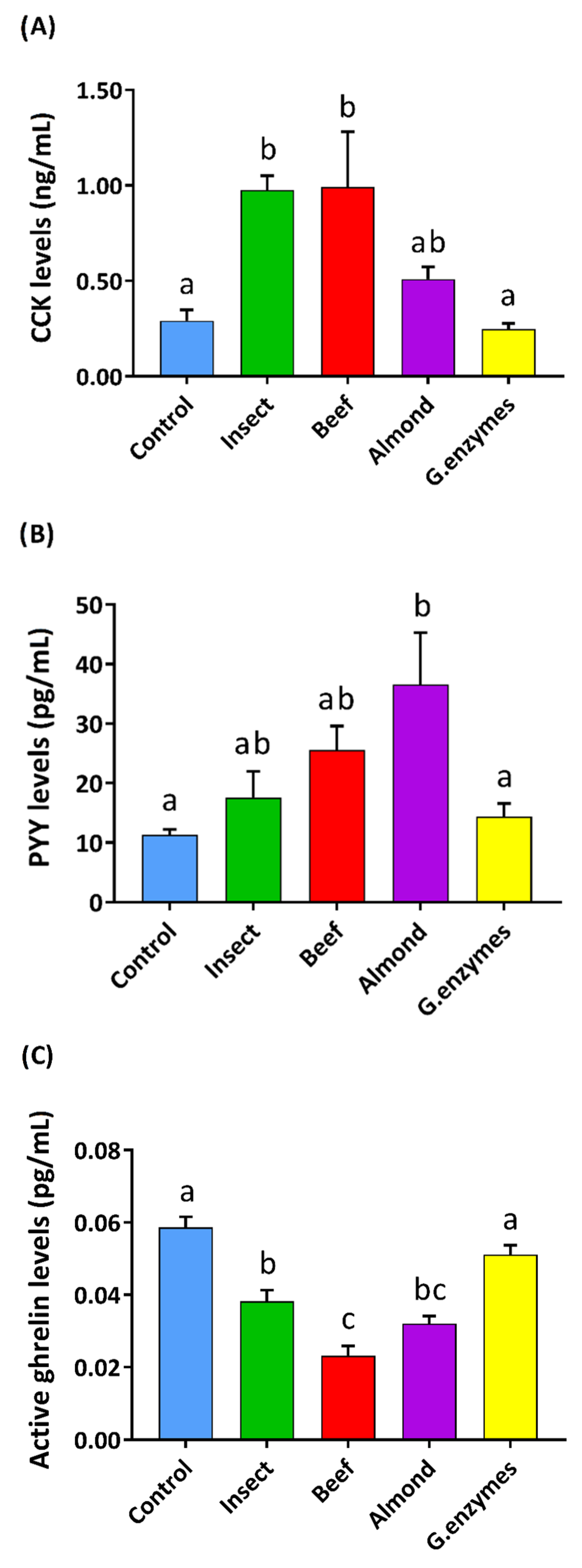
3.2. Insect Gastric Digestion Stimulated Pig Duodenal Enteroendocrine Secretions Differently than Beef or Almond
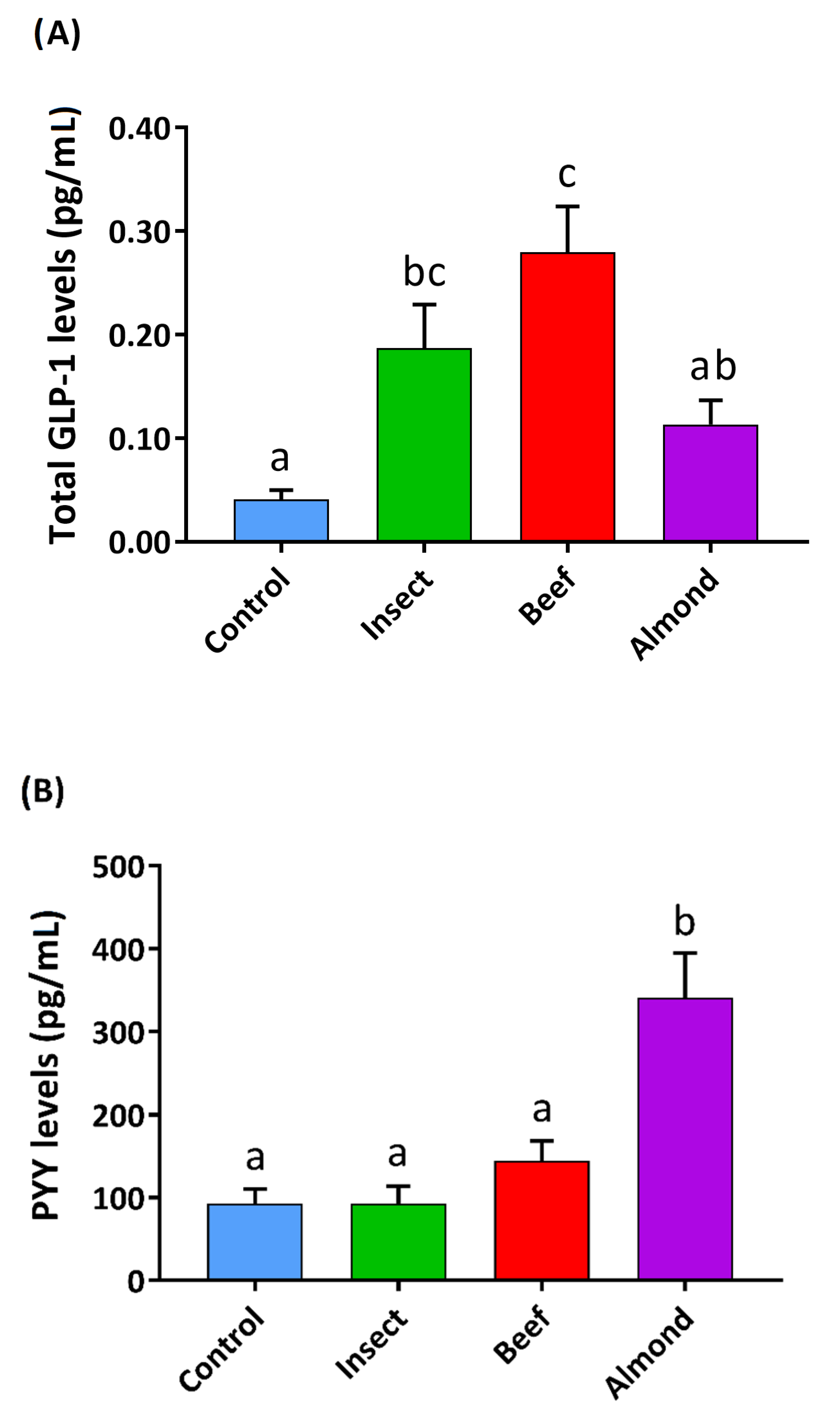
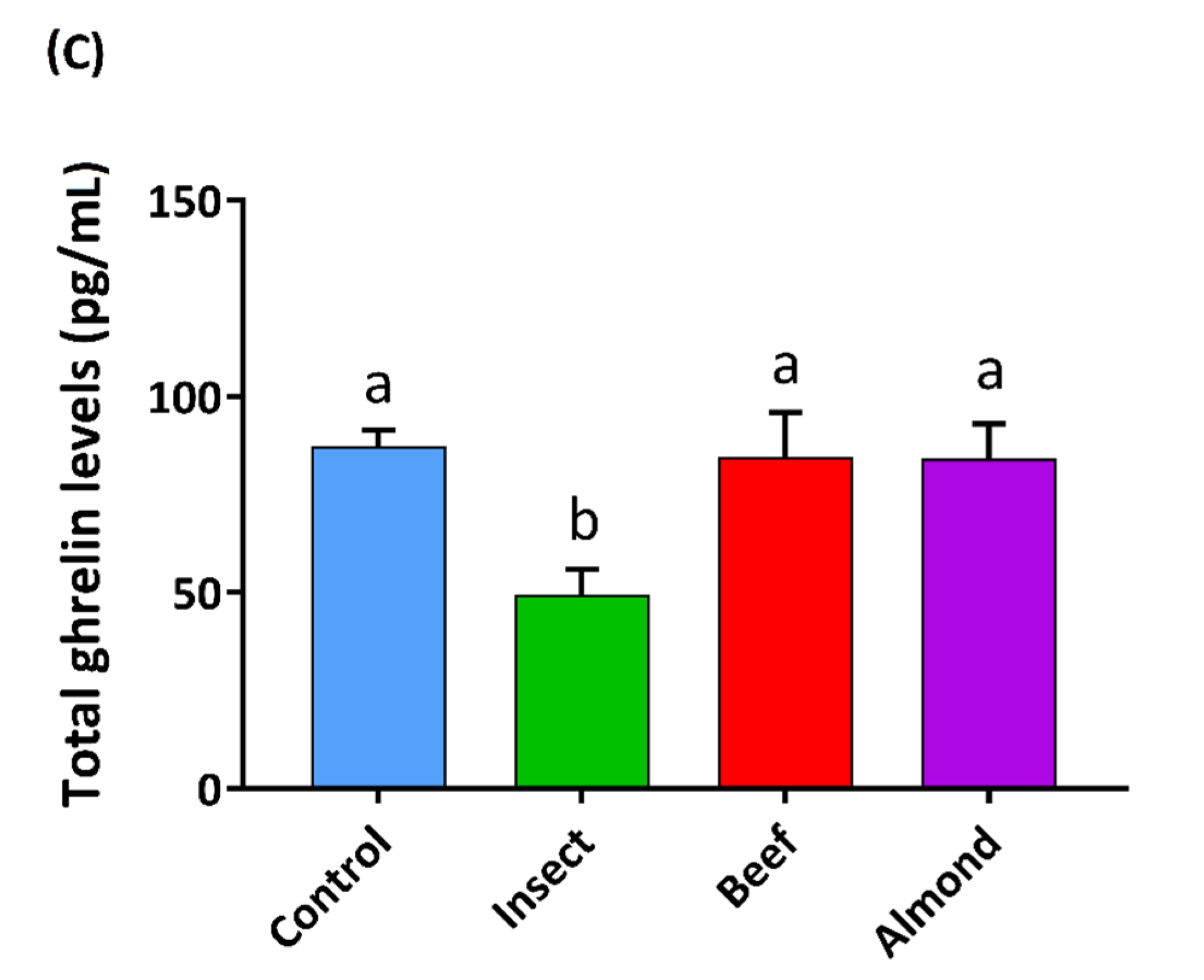
3.3. Insect Intestinal Digestion Induced GLP1 Secretion and Reduced Ghrelin Secretion in Human Colon Samples
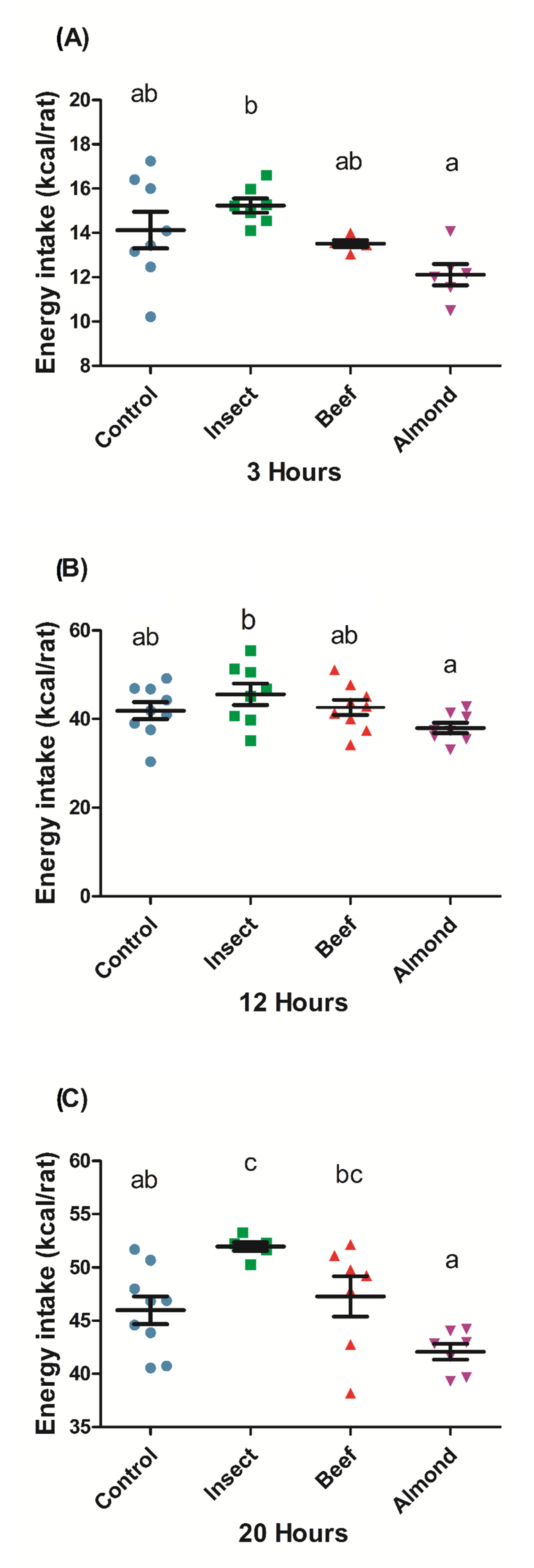
3.4. Buffalo Alphitobius diaperinus Powder Stimulated Food Intake in Rats
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- United Nations. Department of Economic and Social Affairs World Population Prospects 2019: Highlights; United Nations Publisher: New York, NY, USA, 2019. [Google Scholar]
- FAO Edible Insects. Future Prospects for Food and Feed Security; FAO: Rome, Italy, 2013; Volume 171, ISBN 9789251075951. [Google Scholar]
- Oonincx, D.G.A.B.; van Itterbeeck, J.; Heetkamp, M.J.W.; van den Brand, H.; van Loon, J.J.A.; van Huis, A. An exploration on greenhouse gas and ammonia production by insect species suitable for animal or human consumption. PLoS ONE 2010, 5, e14445. [Google Scholar] [CrossRef] [Green Version]
- Sun-Waterhouse, D.; Waterhouse, G.I.N.; You, L.; Zhang, J.; Liu, Y.; Ma, L.; Gao, J.; Dong, Y. Transforming insect biomass into consumer wellness foods: A review. Food Res. Int. 2016, 89, 129–151. [Google Scholar] [CrossRef]
- Van Huis, A. Edible insects are the future? Proc. Nutr. Soc. 2016, 75, 294–305. [Google Scholar] [CrossRef] [Green Version]
- Wu, G. Dietary protein intake and human health. Food Funct. 2016, 7, 1251–1265. [Google Scholar] [CrossRef] [Green Version]
- Melo, V.; Garcia, M.; Sandoval, H.; Jiménez, H.D.; Calvo, C. Quality proteins from edible indigenous insect food of latin. America and Asia. Emirates J. Food Agric. 2011, 23, 283–289. [Google Scholar]
- Finke, M.D. Complete nutrient composition of commercially raised invertebrates used as food for insectivores. Zoo Biol. 2002, 21, 269–285. [Google Scholar] [CrossRef]
- Rumpold, B.A.; Schlüter, O.K. Nutritional composition and safety aspects of edible insects. Mol. Nutr. Food Res. 2013, 57, 802–823. [Google Scholar] [CrossRef] [PubMed]
- Womeni, H.M.; Linder, M.; Tiencheu, B.; Mbiapo, F.T.; Villeneuve, P.; Fanni, J.; Parmentier, M. Oils of insects and larvae consumed in Africa: Potential sources of polyunsaturated fatty acids. Oleagineux. Corps Gras. Lipides 2009, 16, 230–235. [Google Scholar] [CrossRef] [Green Version]
- Christensen, D.L.; Orech, F.O.; Mungai, M.N.; Larsen, T.; Friis, H.; Aagaard-Hansen, J. Entomophagy among the Luo of Kenya: A potential mineral source? Int. J. Food Sci. Nutr. 2006, 57, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Virginia, M.-R.; Tomás, Q.-B.; Rafael, D.-G.; César, G.-U. Consumption of Escamoles (Liometopum apiculatum M.): A Source of Vitamins A and E. J. Appl. Life Sci. Int. 2016, 9, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kulma, M.; Kouřimská, L.; Plachý, V.; Božik, M.; Adámková, A.; Vrabec, V. Effect of sex on the nutritional value of house cricket, Acheta domestica L. Food Chem. 2019, 272, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Finke, M.D. Complete nutrient content of four species of commercially available feeder insects fed enhanced diets during growth. Zoo Biol. 2015, 34, 554–564. [Google Scholar] [CrossRef] [PubMed]
- Stull, V.J.; Finer, E.; Bergmans, R.S.; Febvre, H.P.; Longhurst, C.; Manter, D.K.; Patz, J.A.; Weir, T.L. Impact of Edible Cricket Consumption on Gut Microbiota in Healthy Adults, a Double-blind, Randomized Crossover Trial. Sci. Rep. 2018, 8, 10762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; Jia, J.; Yan, H.; Du, J.; Gui, Z. A novel angiotensin-I converting enzyme (ACE) inhibitory peptide from gastrointestinal protease hydrolysate of silkworm pupa (Bombyx mori) protein: Biochemical characterization and molecular docking study. Peptides 2015, 68, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Yi, H.Y.; Chowdhury, M.; Huang, Y.D.; Yu, X.Q. Insect antimicrobial peptides and their applications. Appl. Microbiol. Biotechnol. 2014, 98, 5807–5822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youn, K.; Yun, E.-Y.; Lee, J.; Kim, J.-Y.; Hwang, J.-S.; Jeong, W.-S.; Jun, M. Oleic acid and linoleic acid from Tenebrio molitor larvae inhibit BACE1 activity in vitro: Molecular docking studies. J. Med. Food 2014, 17, 284–289. [Google Scholar] [CrossRef]
- Cheseto, X.; Kuate, S.P.; Tchouassi, D.P.; Ndung’u, M.; Teal, P.E.A.; Torto, B. Potential of the Desert Locust Schistocerca gregaria (Orthoptera: Acrididae) as an Unconventional Source of Dietary and Therapeutic Sterols. PLoS ONE 2015, 10, e0127171. [Google Scholar] [CrossRef]
- Rehfeld, J.F. The New Biology of Gastrointestinal Hormones. Phys. Rev. 1998, 78, 1087–1108. [Google Scholar] [CrossRef] [Green Version]
- Depoortere, I. Taste receptors of the gut: Emerging roles in health and disease. Gut 2014, 63, 179–190. [Google Scholar] [CrossRef]
- Gribble, F.M. The gut endocrine system as a coordinator of postprandial nutrient homoeostasis. Proc. Nutr. Soc. 2012, 71, 456–462. [Google Scholar] [CrossRef] [Green Version]
- Adriaenssens, A.E.; Reimann, F.; Gribble, F.M. Distribution and Stimulus Secretion Coupling of Enteroendocrine Cells along the Intestinal Tract. Compr. Physiol. 2018, 8, 1603–1638. [Google Scholar] [PubMed]
- Steinert, R.E.; Beglinger, C. Nutrient sensing in the gut: Interactions between chemosensory cells, visceral afferents and the secretion of satiation peptides. Physiol. Behav. 2011, 105, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Gribble, F.M.; Reimann, F. Enteroendocrine Cells: Chemosensors in the Intestinal Epithelium. Annu. Rev. Physiol. 2016, 78, 277–299. [Google Scholar] [CrossRef] [PubMed]
- Hutchison, A.T.; Feinle-Bisset, C.; Fitzgerald, P.C.E.; Standfield, S.; Horowitz, M.; Clifton, P.M.; Luscombe-Marsh, N.D. Comparative effects of intraduodenal whey protein hydrolysate on antropyloroduodenal motility, gut hormones, glycemia, appetite, and energy intake in lean and obese men. Am. J. Clin. Nutr. 2015, 102, 1323–1331. [Google Scholar] [CrossRef]
- Calbet, J.A.L.; Holst, J.J. Gastric emptying, gastric secretion and enterogastrone response after administration of milk proteins or their peptide hydrolysates in humans. Eur. J. Nutr. 2004, 43, 127–139. [Google Scholar] [CrossRef]
- Geraedts, M.C.P.; Troost, F.J.; Fischer, M.A.J.G.; Edens, L.; Saris, W.H.M. Direct induction of CCK and GLP-1 release from murine endocrine cells by intact dietary proteins. Mol. Nutr. Food Res. 2010, 55. [Google Scholar] [CrossRef]
- Foltz, M.; Ansems, P.; Schwarz, J.; Tasker, M.C.; Lourbakos, A.; Gerhardt, C.C. Protein hydrolysates induce CCK release from enteroendocrine cells and act as partial agonists of the CCK 1 receptor. J. Agric. Food Chem. 2008, 56, 837–843. [Google Scholar] [CrossRef]
- Wang, T.Y.; Hsieh, C.H.; Hung, C.C.; Jao, C.L.; Chen, M.C.; Hsu, K.C. Fish skin gelatin hydrolysates as dipeptidyl peptidase IV inhibitors and glucagon-like peptide-1 stimulators improve glycaemic control in diabetic rats: A comparison between warm- and cold-water fish. J. Funct. Foods 2015, 19, 330–340. [Google Scholar] [CrossRef]
- Moreiras, O.; Carbajal, A.; Cabrera, L.; Cuadrado, C. Tablas de Composición de Alimentos, 16th ed.; Ediciones Pirámide: Madrid, Spain, 2013; ISBN 978-84-368-2182-6. [Google Scholar]
- Egger, L.; Ménard, O.; Delgado-Andrade, C.; Alvito, P.; Assunção, R.; Balance, S.; Barberá, R.; Brodkorb, A.; Cattenoz, T.; Clemente, A.; et al. The harmonized INFOGEST in vitro digestion method: From knowledge to action. Food Res. Int. 2016, 88, 217–225. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [Green Version]
- Ginés, I.; Gil-Cardoso, K.; Robles, P.; Arola, L.; Terra, X.; Blay, M.; Ardévol, A.; Pinent, M. Novel ex Vivo Experimental Setup to Assay the Vectorial Transepithelial Enteroendocrine Secretions of Different Intestinal Segments. J. Agric. Food Chem. 2018, 66, 11622–11629. [Google Scholar] [CrossRef] [PubMed]
- González, C.; Carme, Q.; Bové, G.; Jorba, R.; Aleidis, M.; Tarragó, C. Protective properties of grape—Seed proanthocyanidins in human ex vivo acute colonic dysfunction induced by dextran sodium sulfate. Eur. J. Nutr. 2020, 34, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Casanova-Martí, À.; Bravo, F.I.; Serrano, J.; Ardévol, A.; Pinent, M.; Muguerza, B. Antihyperglycemic effect of a chicken feet hydrolysate: Via the incretin system: DPP-IV-inhibitory activity and GLP-1 release stimulation. Food Funct. 2019, 10, 4062–4070. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.; Zhou, G.; Song, S.; Xu, X.; Voglmeir, J.; Liu, L.; Zhao, F.; Li, M.; Li, L.; Yu, X.; et al. Discrimination of in vitro and in vivo digestion products of meat proteins from pork, beef, chicken, and fish. Proteomics 2015, 15, 3688–3698. [Google Scholar] [CrossRef] [Green Version]
- De Angelis, E.; Bavaro, S.L.; Forte, G.; Pilolli, R.; Monaci, L. Heat and Pressure Treatments on Almond Protein Stability and Change in Immunoreactivity after Simulated Human Digestion. Nutrients 2018, 10, 1679. [Google Scholar] [CrossRef] [Green Version]
- Janssen, R.H.; Vincken, J.P.; Arts, N.J.G.; Fogliano, V.; Lakemond, C.M.M. Effect of endogenous phenoloxidase on protein solubility and digestibility after processing of Tenebrio molitor, Alphitobius diaperinus and Hermetia illucens. Food Res. Int. 2019, 121, 684–690. [Google Scholar] [CrossRef]
- Roura, E.; Koopmans, S.-J.; Lallès, J.-P.; Le Huerou-Luron, I.; de Jager, N.; Schuurman, T.; Val-Laillet, D. Critical review evaluating the pig as a model for human nutritional physiology. Nutr. Res. Rev. 2016, 29, 60–90. [Google Scholar] [CrossRef]
- Wewer Albrechtsen, N.J.; Kuhre, R.E.; Toräng, S.; Holst, J.J. The intestinal distribution pattern of appetite and glucose regulatory peptides in mice, rats and pigs. BMC Res. Notes 2016, 9, 60. [Google Scholar] [CrossRef] [Green Version]
- Karhunen, L.J.; Juvonen, K.R.; Huotari, A.; Purhonen, A.K.; Herzig, K.H. Effect of protein, fat, carbohydrate and fibre on gastrointestinal peptide release in humans. Regul. Pept. 2008, 149, 70–78. [Google Scholar] [CrossRef]
- Degen, L.; Oesch, S.; Casanova, M.; Graf, S.; Ketterer, S.; Drewe, J.; Beglinger, C. Effect of peptide YY3-36 on food intake in humans. Gastroenterology 2005, 129, 1430–1436. [Google Scholar] [CrossRef]
- Monteleone, P.; Bencivenga, R.; Longobardi, N.; Serritella, C.; Maj, M. Differential Responses of Circulating Ghrelin to High-Fat or High-Carbohydrate Meal in Healthy Women. J. Clin. Endocrinol. Metab. 2003, 88, 5510–5514. [Google Scholar] [CrossRef] [PubMed]
- Foster-Schubert, K.E.; Overduin, J.; Prudom, C.E.; Liu, J.; Callahan, H.S.; Gaylinn, B.D.; Thorner, M.O.; Cummings, D.E. Acyl and Total Ghrelin Are Suppressed Strongly by Ingested Proteins, Weakly by Lipids, and Biphasically by Carbohydrates. J. Clin. Endocrinol. Metab. 2008, 93, 1971–1979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egger, L.; Schlegel, P.; Baumann, C.; Stoffers, H.; Guggisberg, D.; Brügger, C.; Dürr, D.; Stoll, P.; Vergères, G.; Portmann, R. Physiological comparability of the harmonized INFOGEST in vitro digestion method to in vivo pig digestion. Food Res. Int. 2017, 102, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Caron, J.; Domenger, D.; Belguesmia, Y.; Kouach, M.; Lesage, J.; Goossens, J.F.; Dhulster, P.; Ravallec, R.; Cudennec, B. Protein digestion and energy homeostasis: How generated peptides may impact intestinal hormones? Food Res. Int. 2016, 88, 310–318. [Google Scholar] [CrossRef]
- Hollingworth, S.; Dalton, M.; Blundell, J.; Finlayson, G. Evaluation of the Influence of Raw Almonds on Appetite Control: Satiation, Satiety, Hedonics and Consumer Perceptions. Nutrients 2019, 11, 2030. [Google Scholar] [CrossRef] [Green Version]
- Du, K.; Markus, E.; Fecych, M.; Rhodes, J.S.; Beverly, J.L. Satiety and memory enhancing effects of a high-protein meal depend on the source of protein. Nutr. Neurosci. 2018, 21, 257–267. [Google Scholar] [CrossRef]
- Hall, W.L.; Millward, D.J.; Long, S.J.; Morgan, L.M. Casein and whey exert different effects on plasma amino acid profiles, gastrointestinal hormone secretion and appetite. Hill Blundel. 2020, 89, 239–248. [Google Scholar] [CrossRef]
- Anderson, G.H.; Tecimer, S.N.; Shah, D.; Zafar, T.A. Protein Source, Quantity, and Time of Consumption Determine the Effect of Proteins on Short-Term Food Intake in Young Men. J. Nutr. 2004, 134, 3011–3015. [Google Scholar] [CrossRef] [Green Version]
- Pal, S.; Ellis, V. The acute effects of four protein meals on insulin, glucose, appetite and energy intake in lean men. Br. J. Nutr. 2010, 104, 1241–1248. [Google Scholar] [CrossRef]
- Mahé, S.; Roos, N.; Benamouzig, R.; Davin, L.; Luengo, C.; Gagnon, L.; Gaussergès, N.; Rautureau, J.; Tomé, D. Gastrojejunal kinetics and the digestion of (15N) beta-lactoglobulin and casein in humans: The influence of the nature and quantity of the protein. Am. J. Clin. Nutr. 1996, 63, 546–552. [Google Scholar] [CrossRef] [Green Version]
- Boye, J.; Wijesinha-Bettoni, R.; Burlingame, B. Protein quality evaluation twenty years after the introduction of the protein digestibility corrected amino acid score method. Br. J. Nutr. 2020, 108, S183–S211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- House, J.D.; Hill, K.; Neufeld, J.; Franczyk, A.; Nosworthy, M.G. Determination of the protein quality of almonds (Prunus dulcis L.) as assessed by in vitro and in vivo methodologies. Food Sci. Nutr. 2019, 7, 2932–2938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Vliet, S.; Burd, N.A.; van Loon, L.J. The Skeletal Muscle Anabolic Response to Plant-Versus Animal-Based Protein Consumption. J. Nutr. 2015, 145, 1981–1991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marono, S.; Piccolo, G.; Loponte, R.; Meo, C.D.; Attia, Y.A.; Nizza, A.; Bovera, F. In vitro crude protein digestibility of tenebrio molitor and hermetia illucens insect meals and its correlation with chemical composition traits. Ital. J. Anim. Sci. 2015, 14, 338–343. [Google Scholar] [CrossRef] [Green Version]
- Yi, L.; Van Boekel, M.A.J.S.; Boeren, S.; Lakemond, C.M.M. Protein identification and in vitro digestion of fractions from Tenebrio molitor. Eur. Food Res. Technol. 2016, 242, 1285–1297. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Protein mg/mL | Triglycerides mg/mL | Glucose mg/mL | |
|---|---|---|---|---|
| Gastric digestion | Insect | 26.60 | 5.84 | 0.20 |
| IPC | 31.49 | 15.30 | 0.22 | |
| Beef | 21.60 | 0.92 | 0.09 | |
| Almond | 26.55 | 2.24 | 0.12 | |
| Enzymes | 0.65 | 0.05 | n.d. | |
| Intestinal digestion | Insect | 15.02 | 8.97 | 0.09 |
| IPC | 14.38 | 12.91 | 0.08 | |
| Beef | 9.28 | 0.70 | 0.06 | |
| Almond | 10.29 | 9.44 | 0.1 | |
| Enzymes | 0.62 | 0.05 | n.d. | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miguéns-Gómez, A.; Grau-Bové, C.; Sierra-Cruz, M.; Jorba-Martín, R.; Caro, A.; Rodríguez-Gallego, E.; Beltrán-Debón, R.; Blay, M.T.; Terra, X.; Ardévol, A.; et al. Gastrointestinally Digested Protein from the Insect Alphitobius diaperinus Stimulates a Different Intestinal Secretome than Beef or Almond, Producing a Differential Response in Food Intake in Rats. Nutrients 2020, 12, 2366. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12082366
Miguéns-Gómez A, Grau-Bové C, Sierra-Cruz M, Jorba-Martín R, Caro A, Rodríguez-Gallego E, Beltrán-Debón R, Blay MT, Terra X, Ardévol A, et al. Gastrointestinally Digested Protein from the Insect Alphitobius diaperinus Stimulates a Different Intestinal Secretome than Beef or Almond, Producing a Differential Response in Food Intake in Rats. Nutrients. 2020; 12(8):2366. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12082366
Chicago/Turabian StyleMiguéns-Gómez, Alba, Carme Grau-Bové, Marta Sierra-Cruz, Rosa Jorba-Martín, Aleidis Caro, Esther Rodríguez-Gallego, Raúl Beltrán-Debón, M Teresa Blay, Ximena Terra, Anna Ardévol, and et al. 2020. "Gastrointestinally Digested Protein from the Insect Alphitobius diaperinus Stimulates a Different Intestinal Secretome than Beef or Almond, Producing a Differential Response in Food Intake in Rats" Nutrients 12, no. 8: 2366. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12082366