Disparities in Head and Neck Cancer: A Case for Chemoprevention with Vitamin D

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Characteristics of the Head and Neck Cancer Patient Population
2.2. Dietary Intake Information and Risk Factor Characterization
2.3. Blood Measures
2.4. Head and Neck Cancer Cell Lines
2.5. RNA Isolation and microRNA Assay
2.6. MiR Gene Targets and Enrichment Analysis
2.7. Proteomic Profiling
2.8. Lysate Collection and Western Blot Analysis of HNC Cancer Cells Following Vitamin D
3. Results
3.1. Patient Characteristics
3.2. Blood Levels of Vitamin D, Parathyroid, and Calcium Among Head and Neck Cancer Patients
3.3. Nutrient Levels of Vitamin D and Calcium Based on Dietary Screeners
3.4. Top miRs Dysregulated in HNC Cell Lines by Vitamin D
3.5. DAVID Enrichment Analysis
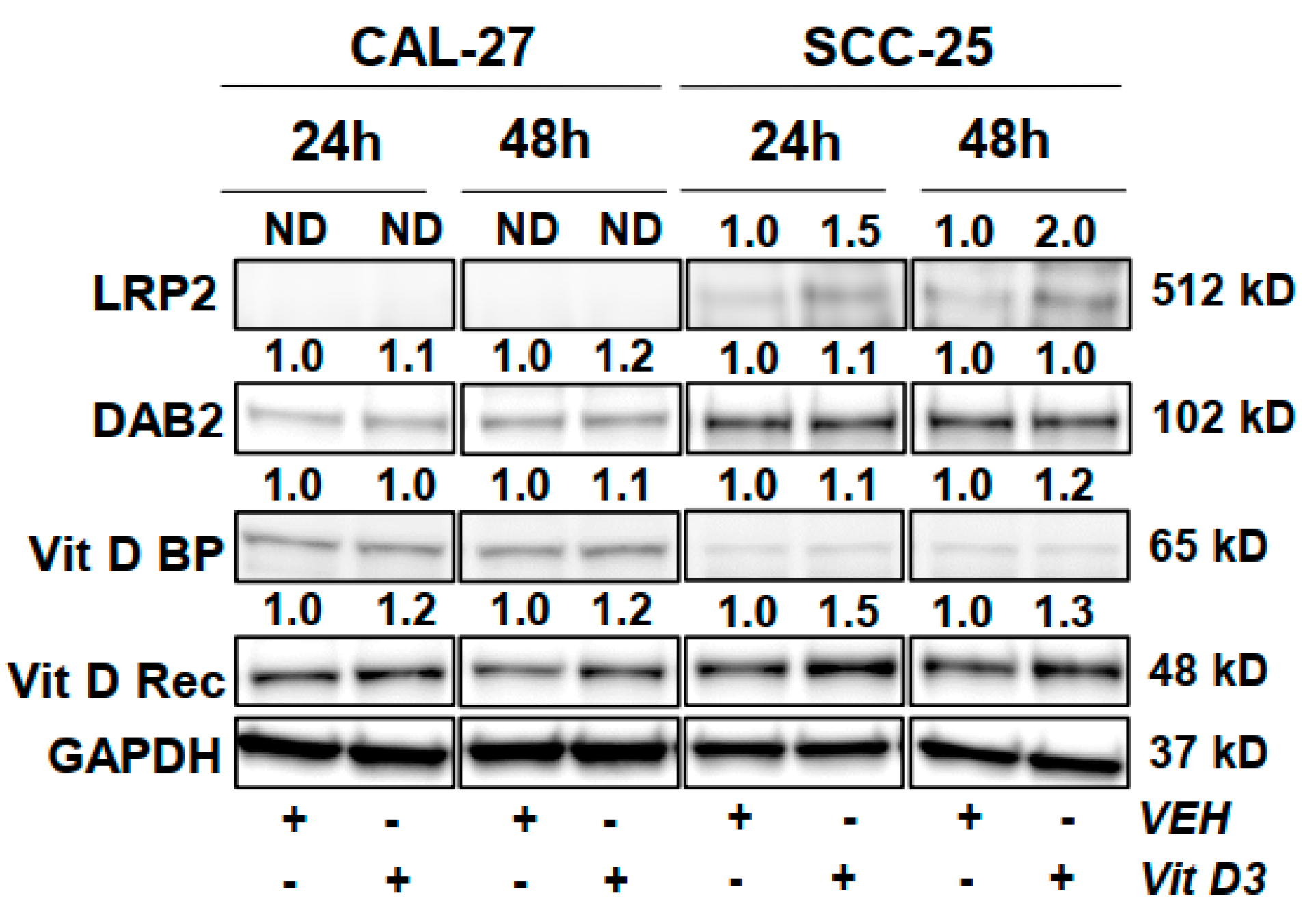
3.6. Protein Level Changes in HNC Cells Following Vitamin D Treatment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rettig, E.M.; D’Souza, G. Epidemiology of Head and Neck Cancer. Surg. Oncol. Clin. N. Am. 2015, 24, 379–396. [Google Scholar] [CrossRef] [PubMed]
- Wissinger, E.; Griebsch, I.; Lungershausen, J.; Foster, T.; Pashos, C.L. The Economic Burden of Head and Neck Cancer: A Systematic Literature Review. Pharmacoeconomics 2014, 32, 865–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daraei, P.; Moore, C.E. Racial Disparity among the Head and Neck Cancer Population. J. Cancer Educ. 2015, 30, 546–551. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, A.K.; Engels, E.A.; Pfeiffer, R.M.; Hernandez, B.Y.; Xiao, W.; Kim, E.; Jiang, B.; Goodman, M.T.; Sibug-Saber, M.; Cozen, W.; et al. Human Papillomavirus and Rising Oropharyngeal Cancer Incidence in the United States. J. Clin. Oncol. 2011, 29, 4294–4301. [Google Scholar] [CrossRef] [PubMed]
- Benson, E.; Li, R.; Eisele, D.; Fakhry, C. The Clinical Impact of HPV Tumor Status upon Head and Neck Squamous Cell Carcinomas. Oral Oncol. 2014, 50, 565–574. [Google Scholar] [CrossRef] [Green Version]
- O’Rorke, M.A.; Ellison, M.V.; Murray, L.J.; Moran, M.; James, J.; Anderson, L.A. Human Papillomavirus Related Head and Neck Cancer Survival: A Systematic Review and Meta-Analysis. Oral Oncol. 2012, 48, 1191–1201. [Google Scholar] [CrossRef]
- Al-Othman, M.O.F.; Morris, C.G.; Logan, H.L.; Hinerman, R.W.; Amdur, R.J.; Mendenhall, W.M. Impact of Race on Outcome after Definitive Radiotherapy for Squamous Cell Carcinoma of the Head and Neck. Cancer 2003, 98, 2467–2472. [Google Scholar] [CrossRef]
- Goodwin, W.J.; Thomas, G.R.; Parker, D.F.; Joseph, D.; Levis, S.; Franzmann, E.; Anello, C.; Hu, J.J. Unequal Burden of Head and Neck Cancer in the United States. Head Neck 2008, 30, 358–371. [Google Scholar] [CrossRef]
- Kompelli, A.; Cartmell, K.B.; Sterba, K.R.; Alberg, A.J.; Xiao, C.C.; Sood, A.J.; Garrett-Mayer, E.; White-Gilbertson, S.J.; Rosenzweig, S.A.; Day, T.A. An Assessment of Racial Differences in Epidemiological, Clinical and Psychosocial Factors among Head and Neck Cancer Patients at the Time of Surgery. World J. Otorhinolaryngol. Head Neck Surg. 2020, 6, 41–48. [Google Scholar] [CrossRef]
- Mondul, A.M.; Weinstein, S.J.; Layne, T.M.; Albanes, D. Vitamin D and Cancer Risk and Mortality: State of the Science, Gaps, and Challenges. Epidemiol. Rev. 2017, 39, 28–48. [Google Scholar] [CrossRef]
- Zhao, Y.; Chen, C.; Pan, W.; Gao, M.; He, W.; Mao, R.; Lin, T.; Huang, J. Comparative Efficacy of Vitamin D Status in Reducing the Risk of Bladder Cancer: A Systematic Review and Network Meta-Analysis. Nutrition 2016, 32, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Shao, X.; Yao, Y.; Xu, L.; Chang, L.; Jiang, Z.; Lin, Z. Positive Association between Circulating 25-Hydroxyvitamin D Levels and Prostate Cancer Risk: New Findings from an Updated Meta-Analysis. J. Cancer Res. Clin. Oncol. 2014, 140, 1465–1477. [Google Scholar] [CrossRef] [PubMed]
- Yokosawa, E.B.; Arthur, A.E.; Rentschler, K.M.; Wolf, G.T.; Rozek, L.S.; Mondul, A.M. Vitamin D Intake and Survival and Recurrence in Head and Neck Cancer Patients. Laryngoscope 2018, 128, E371–E376. [Google Scholar] [CrossRef] [PubMed]
- Vaughan-Shaw, P.G.; O’Sullivan, F.; Farrington, S.M.; Theodoratou, E.; Campbell, H.; Dunlop, M.G.; Zgaga, L. The Impact of Vitamin D Pathway Genetic Variation and Circulating 25-HydroxyVitamin D on Cancer Outcome: Systematic Review and Meta-Analysis. Br. J. Cancer 2017, 116, 1092–1110. [Google Scholar] [CrossRef]
- Chan, J.; Jaceldo-Siegl, K.; Fraser, G.E. Determinants of Serum 25 Hydroxyvitamin D Levels in a Nationwide Cohort of Blacks and Non-Hispanic Whites. Cancer Causes Control 2010, 21, 501–511. [Google Scholar] [CrossRef] [Green Version]
- Freedman, D.M.; Cahoon, E.K.; Rajaraman, P.; Major, J.M.; Doody, M.M.; Alexander, B.H.; Hoffbeck, R.W.; Kimlin, M.G.; Graubard, B.I.; Linet, M.S. Sunlight and Other Determinants of Circulating 25-Hydroxyvitamin D Levels in Black and White Participants in a Nationwide US Study. Am. J. Epidemiol. 2013, 177, 180–192. [Google Scholar] [CrossRef] [Green Version]
- Aranow, C. Vitamin D and the Immune System. J. Investig. Med. 2011, 59, 881–886. [Google Scholar] [CrossRef] [Green Version]
- Dawson-Hughes, B.; Harris, S.S.; Krall, E.A.; Dallal, G.E. Effect of Calcium and Vitamin D Supplementation on Bone Density in Men and Women 65 Years of Age or Older. N. Engl. J. Med. 1997, 337, 670–676. [Google Scholar] [CrossRef]
- Bochen, F.; Balensiefer, B.; Körner, S.; Bittenbring, J.T.; Neumann, F.; Koch, A.; Bumm, K.; Marx, A.; Wemmert, S.; Papaspyrou, G.; et al. Vitamin D Deficiency in Head and Neck Cancer Patients–Prevalence, Prognostic Value and Impact on Immune Function. Oncoimmunology 2018, 7. [Google Scholar] [CrossRef]
- Coutinho-Camillo, C.M.; Lourenço, S.V.; Nishimoto, I.N.; Kowalski, L.P.; Soares, F.A. Nucleophosmin, P53, and Ki-67 Expression Patterns on an Oral Squamous Cell Carcinoma Tissue Microarray. Hum. Pathol. 2010, 41, 1079–1086. [Google Scholar] [CrossRef]
- Zheng, G.; Zhang, Z.; Liu, H.; Xiong, Y.; Luo, L.; Jia, X.; Peng, C.; Zhang, Q.; Li, N.; Gu, Y.; et al. Hsp27-Mediated Extracellular and Intracellular Signaling Pathways Synergistically Confer Chemoresistance in Squamous Cell Carcinoma of Tongue. Clin. Cancer Res. 2018, 24, 1163–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanagawa, T.; Omura, K.; Harada, H.; Ishii, T.; Uwayama, J.; Nakaso, K.; Iwasa, S.; Koyama, Y.; Onizawa, K.; Yusa, H.; et al. Peroxiredoxin I Expression in Tongue Squamous Cell Carcinomas as Involved in Tumor Recurrence. Int. J. Oral Maxillofac. Surg. 2005, 34, 915–920. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.T.; Tong, X.; Zhang, Z.X.; Sun, Y.Y.; Yan, W.; Xu, Z.M.; Fu, W.N. MYCT1 Represses Apoptosis of Laryngeal Cancerous Cells through the MAX/MiR-181a/NPM1 Pathway. FEBS J. 2019, 286, 3892–3908. [Google Scholar] [CrossRef] [PubMed]
- QuestAssureD TM 25-Hydroxy and 1,25-Dihydroxyvitamin D. Test Detail. Quest Diagnostics. Available online: https://testdirectory.questdiagnostics.com/test/test-detail/16761/questassured-25-hydroxy-and-125-dihydroxyvitamin-d?p=r&q=25-hydroxy vitamin D&cc=MASTER (accessed on 23 April 2020).
- Holick, M.F.; Binkley, N.C.; Bischoff-Ferrari, H.A.; Gordon, C.M.; Hanley, D.A.; Heaney, R.P.; Murad, M.H.; Weaver, C.M. Evaluation, Treatment, and Prevention of Vitamin D Deficiency: An Endocrine Society Clinical Practice Guideline. J. Clin. Endocrinol. Metab. 2011, 96, 1911–1930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rheinwald, J.G.; Beckett, M.A. Tumorigenic Keratinocyte Lines Requiring Anchorage and Fibroblast Support Cultured from Human Squamous Cell Carcinomas1. Cancer Res. 1981, 41, 1657–1663. [Google Scholar]
- Gioanni, J.; Samson, M.; Zanghellini, E.; Mazeau, C.; Ettore, F.; Demard, F.; Chauvel, P.; Duplay, H.; Schneider, M.; Laurent, J.-C.; et al. Characterization of a New Surface Epitope Specific for Human Epithelial Cells Defined by a Monoclonal Antibody and Application to Tumor Diagnosis. Cancer Res. 1987, 47, 4417–4424. [Google Scholar]
- Bhoora, S.; Pather, Y.; Marais, S.; Punchoo, R. Cholecalciferol Inhibits Cell Growth and Induces Apoptosis in the CaSki Cell Line. Med. Sci. (Basel) 2020, 8. [Google Scholar] [CrossRef] [Green Version]
- Shruthi, N.; Prashanthkumar, M.; Venugopalreddy, B.; Suma, M.; Subba Rao, V. Analysis of the Cytotoxic Effects of Vitamin D3 on Colorectal, Breast and Cervical Carcinoma Cell Lines. Biochem. Anal. Biochem. 2017, 6. [Google Scholar] [CrossRef]
- Baek, S.; Lee, Y.S.; Shim, H.E.; Yoon, S.; Baek, S.Y.; Kim, B.S.; Oh, S.O. Vitamin D3 regulates cell viability in gastric cancer and cholangiocarcinoma. Anat. Cell Biol. 2011, 44, 204–209. [Google Scholar] [CrossRef] [Green Version]
- Bennett, R.G.; Wakeley, S.E.; Hamel, F.G.; High, R.R.; Korch, C.; Goldner, W.S. Gene expression of vitamin D metabolic enzymes at baseline and in response to vitamin D treatment in thyroid cancer cell lines. Oncology 2012, 83, 264–272. [Google Scholar] [CrossRef] [Green Version]
- Santos, J.M.; Khan, Z.S.; Munir, M.T.; Tarafdar, K.; Rahman, S.M.; Hussain, F. Vitamin D3 decreases glycolysis and invasiveness, and increases cellular stiffness in breast cancer cells. J. Nutr. Biochem. 2018, 53, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Charoenngam, N.; Holick, M.F. Immunologic Effects of Vitamin D on Human Health and Disease. Nutrients 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Berlin, J.L.; Shantha, G.P.; Yeager, H.; Thomas-Hemak, L. Serum vitamin D levels may not reflect tissue-level vitamin D in sarcoidosis. BMJ Case Rep. 2014, 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kresty, L.; Clarke, J.; Ezell, K.; Exum, A.; Howell, A.; Guettouche, T. MicroRNA Alterations in Barrett′s Esophagus, Esophageal Adenocarcinoma, and Esophageal Adenocarcinoma Cell Lines Following Cranberry Extract Treatment: Insights for Chemoprevention. J. Carcinog. 2011, 10. [Google Scholar] [CrossRef]
- Chou, C.H.; Shrestha, S.; Yang, C.D.; Chang, N.W.; Lin, Y.L.; Liao, K.W.; Huang, W.C.; Sun, T.H.; Tu, S.J.; Lee, W.H.; et al. MiRTarBase Update 2018: A Resource for Experimentally Validated MicroRNA-Target Interactions. Nucleic Acids Res. 2018, 46, D296–D302. [Google Scholar] [CrossRef]
- Lee, Y.J.; Kim, V.; Muth, D.C.; Witwer, K.W. Validated MicroRNA Target Databases: An Evaluation. Drug Dev. Res. 2015, 76, 389–396. [Google Scholar] [CrossRef] [Green Version]
- Tsuprykov, O.; Chen, X.; Hocher, C.F.; Skoblo, R.; Yin, L.; Hocher, B. Why Should We Measure Free 25(OH) Vitamin D? J. Steroid Biochem. Mol. Biol. 2018, 180, 87–104. [Google Scholar] [CrossRef]
- Nagai, J.; Christensen, E.I.; Morris, S.M.; Willnow, T.E.; Cooper, J.A.; Nielsen, R. Mutually Dependent Localization of Megalin and Dab2 in the Renal Proximal Tubule. Am. J. Physiol. Ren. Physiol. 2005, 289. [Google Scholar] [CrossRef]
- Rose, B.S.; Jeong, J.H.; Nath, S.K.; Lu, S.M.; Mell, L.K. Population-Based Study of Competing Mortality in Head and Neck Cancer. J. Clin. Oncol. 2011, 29, 3503–3509. [Google Scholar] [CrossRef]
- American Cancer Society. Cancer Facts & Figures 2020; American Cancer Society: Atlanta, GA, USA, 2020. [Google Scholar]
- American Cancer Society. Cancer Facts and Figures for African Americans 2019–2021; American Cancer Society: Atlanta, GA, USA, 2019. [Google Scholar]
- Mahal, B.A.; Inverso, G.; Aizer, A.A.; Bruce Donoff, R.; Chuang, S.K. Impact of African-American Race on Presentation, Treatment, and Survival of Head and Neck Cancer. Oral Oncol. 2014, 50, 1177–1181. [Google Scholar] [CrossRef]
- Zakeri, K.; Macewan, I.; Vazirnia, A.; Cohen, E.E.W.; Spiotto, M.T.; Haraf, D.J.; Vokes, E.E.; Weichselbaum, R.R.; Mell, L.K. Race and Competing Mortality in Advanced Head and Neck Cancer. Oral Oncol. 2014, 50, 40–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramanian, S.; Chen, A. Treatment Patterns and Survival among Low-Income Medicaid Patients with Head and Neck Cancer. JAMA Otolaryngol. Head Neck Surg. 2013, 139, 489–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, X.L.; Liu, C.C. Racial/Ethnic Disparities in Socioeconomic Status, Diagnosis, Treatment and Survival among Medicare-Insured Men and Women with Head and Neck Cancer. J. Health Care Poor Underserved 2010, 21, 913–930. [Google Scholar] [CrossRef] [PubMed]
- Molina, M.A.; Cheung, M.C.; Perez, E.A.; Byrne, M.M.; Franceschi, D.; Moffat, F.L.; Livingstone, A.S.; Goodwin, W.J.; Gutierrez, J.C.; Koniaris, L.G. African American and Poor Patients Have a Dramatically Worse Prognosis for Head and Neck Cancer: An Examination of 20,915 Patients. Cancer 2008, 113, 2797–2806. [Google Scholar] [CrossRef]
- Arbes, S.J. Factors Contributing to the Poorer Survival of Black Americans Diagnosed with Oral Cancer (United States). Cancer Causes Control 1999, 10, 513–523. [Google Scholar] [CrossRef]
- Ellis, L.; Canchola, A.J.; Spiegel, D.; Ladabaum, U.; Haile, R.; Gomez, S.L. Racial and Ethnic Disparities in Cancer Survival: The Contribution of Tumor, Sociodemographic, Institutional, and Neighborhood Characteristics. J. Clin. Oncol. 2018, 36, 25–33. [Google Scholar] [CrossRef]
- Saini, A.T.; Genden, E.M.; Megwalu, U.C. Sociodemographic Disparities in Choice of Therapy and Survival in Advanced Laryngeal Cancer. Am. J. Otolaryngol. Head Neck Med. Surg. 2016, 37, 65–69. [Google Scholar] [CrossRef]
- Russo, D.P.; Tham, T.; Bardash, Y.; Kraus, D. The Effect of Race in Head and Neck Cancer: A Meta-Analysis Controlling for Socioeconomic Status. Am. J. Otolaryngol. 2020, 41, 102624. [Google Scholar] [CrossRef]
- Yetley, E.A. Assessing the Vitamin D Status of the US Population. Am. J. Clin. Nutr. 2008, 88, 558–564. [Google Scholar] [CrossRef]
- Harris, S.S.; Soteriades, E.; Coolidge, J.A.S.; Mudgal, S.; Dawson-Hughes, B. Vitamin D Insufficiency and Hyperparathyroidism in a Low Income, Multiracial, Elderly Population. J. Clin. Endocrinol. Metab. 2000, 85, 4125–4130. [Google Scholar] [CrossRef]
- Jacobs, E.T.; Alberts, D.S.; Foote, J.A.; Green, S.B.; Hollis, B.W.; Yu, Z.; Martínez, M.E. Vitamin D Insufficiency in Southern Arizona. Am. J. Clin. Nutr. 2008, 87, 608–613. [Google Scholar] [CrossRef] [PubMed]
- Shea, M.K.; Houston, D.K.; Tooze, J.A.; Davis, C.C.; Johnson, M.A.; Hausman, D.B.; Cauley, J.A.; Bauer, D.C.; Tylavsky, F.; Harris, T.B.; et al. Correlates and Prevalence of Insufficient 25-Hydroxyvitamin D Status in Black and White Older Adults: The Health, Aging and Body Composition Study. J. Am. Geriatr. Soc. 2011, 59, 1165–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camozzi, V.; Frigo, A.C.; Zaninotto, M.; Sanguin, F.; Plebani, M.; Boscaro, M.; Schiavon, L.; Luisetto, G. 25-Hydroxycholecalciferol Response to Single Oral Cholecalciferol Loading in the Normal Weight, Overweight, and Obese. Osteoporos. Int. 2016, 27, 2593–2602. [Google Scholar] [CrossRef]
- Gallagher, J.C.; Yalamanchili, V.; Smith, L.M. The Effect of Vitamin D Supplementation on Serum 25OHD in Thin and Obese Women. J. Steroid Biochem. Mol. Biol. 2013, 195–200. [Google Scholar] [CrossRef] [Green Version]
- Blum, M.; Dawson-Hughes, B.; Dallal, G.E. Body Size and Serum 25 Hydroxy Vitamin D Response to Oral Supplements in Healthy Older Adults. J. Am. Coll. Nutr. 2008, 27, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Savastano, S.; Barrea, L.; Savanelli, M.C.; Nappi, F.; Di Somma, C.; Orio, F.; Colao, A. Low Vitamin D Status and Obesity: Role of Nutritionist. Rev. Endocr. Metab. Disord. 2017, 18, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Rock, C.L. Multivitamin-Multimineral Supplements: Who Uses Them? Am. J. Clin. Nutr. 2007, 85, 277–279. [Google Scholar] [CrossRef] [Green Version]
- Radimer, K.; Bindewald, B.; Hughes, J.; Ervin, B.; Swanson, C.; Picciano, M.F. Dietary Supplement Use by US Adults: Data from the National Health and Nutrition Examination Survey, 1999–2000. Am. J. Epidemiol. 2004, 160, 339–349. [Google Scholar] [CrossRef] [Green Version]
- Giammanco, M.; Di Majo, D.; La Guardia, M.; Aiello, S.; Crescimannno, M.; Flandina, C.; Tumminello, F.M.; Leto, G. Vitamin D in Cancer Chemoprevention. Pharm. Biol. 2015, 53, 1399–1434. [Google Scholar] [CrossRef] [Green Version]
- Kennel, K.A.; Drake, M.T. Vitamin D in the Cancer Patient. Curr. Opin. Support. Palliat. Care 2013, 7, 272–277. [Google Scholar] [CrossRef] [Green Version]
- Lenze, N.R.; Farquhar, D.R.; Mazul, A.L.; Masood, M.M.; Zevallos, J.P. Racial Disparities and Human Papillomavirus Status in Oropharyngeal Cancer: A Systematic Review and Meta-Analysis. Head Neck 2019, 41, 256–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, C.E.; Khosla, S.; Chen, L.F.; Joslin, C.E.; Davis, F.G.; Fitzgibbon, M.L.; Freels, S.; Hoskins, K. Racial Differences in Head and Neck Squamous Cell Carcinomas among Non-Hispanic Black and White Males Identified through the National Cancer Database (1998–2012). J. Cancer Res. Clin. Oncol. 2016, 142, 1715–1726. [Google Scholar] [CrossRef] [PubMed]
- Sturgis, E.M.; Cinciripini, P.M. Trends in Head and Neck Cancer Incidence in Relation to Smoking Prevalence: An Emerging Epidemic of Human Papillomavirus-Associated Cancers? Cancer 2007, 110, 1429–1435. [Google Scholar] [CrossRef] [PubMed]
- Morse, D.E.; Kerr, A.R. Disparities in Oral and Pharyngeal Cancer Incidence, Mortality and Survival among Black and White Americans. J. Am. Dent. Assoc. 2006, 137, 203–212. [Google Scholar] [CrossRef] [Green Version]
- Gillison, M.L.; D’Souza, G.; Westra, W.; Sugar, E.; Xiao, W.; Begum, S.; Viscidi, R. Distinct Risk Factor Profiles for Human Papillomavirus Type 16-Positive and Human Papillomavirus Type 16-Negative Head and Neck Cancers. J. Natl. Cancer Inst. 2008, 100, 407–420. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, S.R.; Yueh, B.; McDougall, J.K.; Daling, J.R.; Schwartz, S.M. Human Papillomavirus Infection and Survival in Oral Squamous Cell Cancer: A Population-Based Study. Otolaryngol. Head Neck Surg. 2001, 125, 1–9. [Google Scholar] [CrossRef]
- Dayyani, F.; Etzel, C.J.; Liu, M.; Ho, C.H.; Lippman, S.M.; Tsao, A.S. Meta-Analysis of the Impact of Human Papillomavirus (HPV) on Cancer Risk and Overall Survival in Head and Neck Squamous Cell Carcinomas (HNSCC). Head Neck Oncol. 2010, 2. [Google Scholar] [CrossRef] [Green Version]
- Fakhry, C.; Westra, W.H.; Li, S.; Cmelak, A.; Ridge, J.A.; Pinto, H.; Forastiere, A.; Gillison, M.L. Improved Survival of Patients with Human Papillomavirus-Positive Head and Neck Squamous Cell Carcinoma in a Prospective Clinical Trial. J. Natl. Cancer Inst. 2008, 100, 261–269. [Google Scholar] [CrossRef] [Green Version]
- Huang, Q.; Yang, J.; Zheng, J.; Hsueh, C.; Guo, Y.; Zhou, L. Characterization of Selective Exosomal MicroRNA Expression Profile Derived from Laryngeal Squamous Cell Carcinoma Detected by next Generation Sequencing. Oncol. Rep. 2018, 40, 2584–2594. [Google Scholar] [CrossRef] [Green Version]
- Zeljic, K.; Supic, G.; Magic, Z. New Insights into Vitamin D Anticancer Properties: Focus on MiRNA Modulation. Mol. Genet. Genom. 2017, 292, 511–524. [Google Scholar] [CrossRef]
- Kalinowski, F.C.; Giles, K.M.; Candy, P.A.; Ali, A.; Ganda, C.; Epis, M.R.; Webster, R.J.; Leedman, P.J. Regulation of Epidermal Growth Factor Receptor Signaling and Erlotinib Sensitivity in Head and Neck Cancer Cells by MiR-7. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Horsham, J.; Kalinowski, F.; Epis, M.; Ganda, C.; Brown, R.; Leedman, P. Clinical Potential of MicroRNA-7 in Cancer. J. Clin. Med. 2015, 4, 1668–1687. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.H.; Lin, L.K.; Zhang, S. Epigenetic Silencing of MicroRNA-335 Contributes to Nasopharyngeal Carcinoma Metastasis. Am. J. Otolaryngol. Head Neck Med. Surg. 2020, 41. [Google Scholar] [CrossRef] [PubMed]
- Ou, D.; Wu, Y.; Liu, J.; Lao, X.; Zhang, S.; Liao, G. MiRNA-335 and MiRNA-182 Affect the Occurrence of Tongue Squamous Cell Carcinoma by Targeting Survivin. Oncol. Lett. 2016, 12, 2531–2537. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.M.; Byrom, M.W.; Shelton, J.; Ford, L.P. Antisense Inhibition of Human MiRNAs and Indications for an Involvement of MiRNA in Cell Growth and Apoptosis. Nucleic Acids Res. 2005, 33, 1290–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sethi, N.; Wright, A.; Wood, H.; Rabbitts, P. MicroRNAs and Head and Neck Cancer: Reviewing the First Decade of Research. Eur. J. Cancer 2014, 2619–2635. [Google Scholar] [CrossRef] [PubMed]
- Ayaz, L.; Görür, A.; Yaroǧlu, H.Y.; Özcan, C.; Tamer, L. Differential Expression of MicroRNAs in Plasma of Patients with Laryngeal Squamous Cell Carcinoma: Potential Early-Detection Markers for Laryngeal Squamous Cell Carcinoma. J. Cancer Res. Clin. Oncol. 2013, 139, 1499–1506. [Google Scholar] [CrossRef]
- MacLellan, S.A.; Lawson, J.; Baik, J.; Guillaud, M.; Poh, C.F.Y.; Garnis, C. Differential Expression of MiRNAs in the Serum of Patients with High-Risk Oral Lesions. Cancer Med. 2012, 1, 268–274. [Google Scholar] [CrossRef]
- Yang, Y.; Li, Y.X.; Yang, X.; Jiang, L.; Zhou, Z.J.; Zhu, Y.Q. Progress Risk Assessment of Oral Premalignant Lesions with Saliva MiRNA Analysis. BMC Cancer 2013, 13. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.M.; Lin, P.M.; Wang, Y.M.; Chen, Z.J.; Lin, S.F.; Yang, M.Y. Circulating MiRNA Is a Novel Marker for Head and Neck Squamous Cell Carcinoma. Tumour Biol. 2012, 33, 1933–1942. [Google Scholar] [CrossRef]
- Ramdas, L.; Giri, U.; Ashorn, C.L.; Coombes, K.R.; El-Naggar, A.; Ang, K.K.; Story, M.D. MiRNA Expression Profiles in Head and Neck Squamous Cell Carcinoma and Adjacent Normal Tissue. Head Neck 2009, 31, 642–654. [Google Scholar] [CrossRef] [PubMed]
- Rentoft, M.; Fahlén, J.; Coates, P.J.; Laurell, G.; Sjöström, B.; Rydén, P.; Nylander, K. MiRNA Analysis of Formalin-Fixed Squamous Cell Carcinomas of the Tongue Is Affected by Age of the Samples. Int. J. Oncol. 2011, 38, 61–69. [Google Scholar] [CrossRef] [Green Version]
- Chou, Y.T.; Lin, H.H.; Lien, Y.C.; Wang, Y.H.; Hong, C.F.; Kao, Y.R.; Lin, S.C.; Chang, Y.C.; Lin, S.Y.; Chen, S.J.; et al. EGFR Promotes Lung Tumorigenesis by Activating MiR-7 through a Ras/ERK/Myc Pathway That Targets the Ets2 Transcriptional Repressor ERF. Cancer Res. 2010, 70, 8822–8831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alsahafi, E.; Begg, K.; Amelio, I.; Raulf, N.; Lucarelli, P.; Sauter, T.; Tavassoli, M. Clinical Update on Head and Neck Cancer: Molecular Biology and Ongoing Challenges. Cell Death Dis. 2019. [Google Scholar] [CrossRef] [Green Version]
- Lechner, M.; Frampton, G.M.; Fenton, T.; Feber, A.; Palmer, G.; Jay, A.; Pillay, N.; Forster, M.; Cronin, M.T.; Lipson, D.; et al. Targeted Next-Generation Sequencing of Head and Neck Squamous Cell Carcinoma Identifies Novel Genetic Alterations in HPV+ and HPV- Tumors. Genome Med. 2013, 5, 49. [Google Scholar] [CrossRef] [Green Version]
- Jackson-Weaver, O.; Ungvijanpunya, N.; Yuan, Y.; Qian, J.; Gou, Y.; Wu, J.; Shen, H.; Chen, Y.; Li, M.; Richard, S.; et al. PRMT1-P53 Pathway Controls Epicardial EMT and Invasion. Cell Rep. 2020, 31. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Sturgis, E.M.; Zhu, L.; Zhang, H.; Tao, Y.; Wei, P.; Wei, Q.; Li, G. Mouse Double Minute 4 Variants Modify Susceptibility to Risk of Recurrence in Patients with Squamous Cell Carcinoma of the Oropharynx. Mol. Carcinog. 2018, 57, 361–369. [Google Scholar] [CrossRef]
- Ach, T.; Schwarz-Furlan, S.; Ach, S.; Agaimy, A.; Gerken, M.; Rohrmeier, C.; Zenk, J.; Iro, H.; Brockhoff, G.; Ettl, T. Genomic Aberrations of MDM2, MDM4, FGFR1 and FGFR3 Are Associated with Poor Outcome in Patients with Salivary Gland Cancer. J. Oral Pathol. Med. 2016, 45, 500–509. [Google Scholar] [CrossRef]
- Mishra, R.; Ranjan Das, B. Early Overexpression of Cdk4 and Possible Role of KRF and C-Myc in Chewing Tobacco Mediated Oral Cancer Development. Mol. Biol. Rep. 2003, 30, 207–213. [Google Scholar] [CrossRef]
- Yu, C.; Xing, F.; Zhang, J.; Xu, J.; Li, Y. A Combination of MRNA Expression Profile and MiRNA Expression Profile Identifies Detection Biomarkers in Different Tumor Stages of Laryngeal Squamous Cell Carcinoma. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 7296–7304. [Google Scholar]
- Xuefang, Z.; Ruinian, Z.; Liji, J.; Chun, Z.; Qiaolan, Z.; Jun, J.; Yuming, C.; Junrong, H. MiR-331-3p Inhibits Proliferation and Promotes Apoptosis of Nasopharyngeal Carcinoma Cells by Targeting Elf4B-PI3K-AKT Pathway. Technol. Cancer Res. Treat. 2020, 19, 153303381989225. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhu, J.; Liu, Y.; Duan, C.; Chang, R.; Zhang, C. MicroRNA-331-3p Inhibits Epithelial-mesenchymal Transition by Targeting ErbB2 and VAV2 through the Rac1/PAK1/Β-catenin Axis in Non-small-cell Lung Cancer. Cancer Sci. 2019, 110, cas.14014. [Google Scholar] [CrossRef] [Green Version]
- Zhan, J.; Jiao, D.; Wang, Y.; Song, J.; Wu, J.; Wu, L.; Chen, Q.; Ma, S. Integrated MicroRNA and Gene Expression Profiling Reveals the Crucial MiRNAs in Curcumin Anti-Lung Cancer Cell Invasion. Thorac. Cancer 2017, 8, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Momen-Heravi, F.; Trachtenberg, A.J.; Kuo, W.P.; Cheng, Y.S. Genomewide Study of Salivary MicroRNAs for Detection of Oral Cancer. J. Dent. Res. 2014, 93, 86S–93S. [Google Scholar] [CrossRef]
- Lu, E.; Su, J.; Zhou, Y.; Zhang, C.; Wang, Y. CCL20/CCR6 Promotes Cell Proliferation and Metastasis in Laryngeal Cancer by Activating P38 Pathway. Biomed. Pharmacother. 2017, 85, 486–492. [Google Scholar] [CrossRef]
- Zhou, Z.X.; Zhang, Z.P.; Tao, Z.Z.; Tan, T.Z. MiR-632 Promotes Laryngeal Carcinoma Cell Proliferation, Migration, and Invasion through Negative Regulation of GSK3β. Oncol. Res. 2020, 28, 21–31. [Google Scholar] [CrossRef]
- Grisendi, S.; Mecucci, C.; Falini, B.; Pandolfi, P.P. Nucleophosmin and Cancer. Nat. Rev. Cancer 2006, 493–505. [Google Scholar] [CrossRef] [PubMed]
- Saini, J.; Sharma, P.K. Clinical, Prognostic and Therapeutic Significance of Heat Shock Proteins in Cancer. Curr. Drug Targets 2018, 19, 1478–1490. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, N.; Xue, M.; Weickert, M.O.; Thornalley, P.J. Multiple Roles of Glyoxalase 1-Mediated Suppression of Methylglyoxal Glycation in Cancer Biology—Involvement in Tumour Suppression, Tumour Growth, Multidrug Resistance and Target for Chemotherapy. Semin. Cancer Biol. 2018, 83–93. [Google Scholar] [CrossRef]
- Kumar, N.; Prasad, P.; Jash, E.; Saini, M.; Husain, A.; Goldman, A.; Sehrawat, S. Insights into Exchange Factor Directly Activated by CAMP (EPAC) as Potential Target for Cancer Treatment. Mol. Cell. Biochem. 2018, 447, 77–92. [Google Scholar] [CrossRef]
- Monteiro, F.L.; Baptista, T.; Amado, F.; Vitorino, R.; Jerónimo, C.; Helguero, L.A. Expression and Functionality of Histone H2A Variants in Cancer. Oncotarget 2014, 5, 3428–3443. [Google Scholar] [CrossRef] [Green Version]
- Dhar, S.K.; St. Clair, D.K. Nucleophosmin Blocks Mitochondrial Localization of P53 and Apoptosis. J. Biol. Chem. 2009, 284, 16409–16418. [Google Scholar] [CrossRef] [Green Version]
- Jeon, Y.J.; Cho, J.H.; Lee, S.Y.; Choi, Y.H.; Park, H.; Jung, S.; Shim, J.H.; Chae, J.I. Esculetin Induces Apoptosis Through EGFR/PI3K/Akt Signaling Pathway and Nucleophosmin Relocalization. J. Cell. Biochem. 2016, 117, 1210–1221. [Google Scholar] [CrossRef] [PubMed]
- Shandilya, J.; Swaminathan, V.; Gadad, S.S.; Choudhari, R.; Kodaganur, G.S.; Kundu, T.K. Acetylated NPM1 Localizes in the Nucleoplasm and Regulates Transcriptional Activation of Genes Implicated in Oral Cancer Manifestation. Mol. Cell. Biol. 2009, 29, 5115–5127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onikubo, T.; Nicklay, J.J.; Xing, L.; Warren, C.; Anson, B.; Wang, W.L.; Burgos, E.S.; Ruff, S.E.; Shabanowitz, J.; Cheng, R.H.; et al. Developmentally Regulated Post-Translational Modification of Nucleoplasmin Controls Histone Sequestration and Deposition. Cell Rep. 2015, 10, 1735–1748. [Google Scholar] [CrossRef] [Green Version]
- Lindström, M.S. NPM1/B23: A Multifunctional Chaperone in Ribosome Biogenesis and Chromatin Remodeling. Biochem. Res. Int. 2011, 2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreycy, N.; Gotzian, C.; Fleming, T.; Flechtenmacher, C.; Grabe, N.; Plinkert, P.; Hess, J.; Zaoui, K. Glyoxalase 1 Expression Is Associated with an Unfavorable Prognosis of Oropharyngeal Squamous Cell Carcinoma. BMC Cancer 2017, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geng, X.; Ma, J.; Zhang, F.; Xu, C. Glyoxalase i in Tumor Cell Proliferation and Survival and as a Potential Target for Anticancer Therapy. Oncol. Res. Treat. 2014, 37, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Ernst, B.P.; Wiesmann, N.; Gieringer, R.; Eckrich, J.; Brieger, J. HSP27 Regulates Viability and Migration of Cancer Cell Lines Following Irradiation. J. Proteom. 2020, 226, 103886. [Google Scholar] [CrossRef]
- Guttmann, D.M.; Hart, L.; Du, K.; Seletsky, A.; Koumenis, C. Inhibition of Hsp27 Radiosensitizes Head-and-Neck Cancer by Modulating Deoxyribonucleic Acid Repair. Int. J. Radiat. Oncol. Biol. Phys. 2013, 87, 168–175. [Google Scholar] [CrossRef]
- Zhu, Z.K.; Wang, Y.S.; Xu, X. Biology Behavior of Head and Neck Squamous Cell Cancer Cells Changes after Knocking down Heat Shock Protein 27. Hua Xi Kou Qiang Yi Xue Za Zhi 2020, 38, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Berggren, K.L.; Restrepo Cruz, S.; Hixon, M.D.; Cowan, A.T.; Keysar, S.B.; Craig, S.; James, J.; Barry, M.; Ozbun, M.A.; Jimeno, A.; et al. MAPKAPK2 (MK2) Inhibition Mediates Radiation-Induced Inflammatory Cytokine Production and Tumor Growth in Head and Neck Squamous Cell Carcinoma. Oncogene 2019, 38, 7329–7341. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Ren, X.; Wu, J.; Gao, X.; Cen, X.; Wang, S.; Sheng, S.; Chen, Q.; Tang, Y.J.; Liang, X.H.; et al. HSP27 Associates with Epithelial–Mesenchymal Transition, Stemness and Radioresistance of Salivary Adenoid Cystic Carcinoma. J. Cell. Mol. Med. 2018, 22, 2283–2298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Lim, H.; Kim, S.; Cho, H.; Kim, Y.; Li, X.; Choi, H.; Kim, O. Effects of HSP27 Downregulation on PDT Resistance through PDT-Induced Autophagy in Head and Neck Cancer Cells. Oncol. Rep. 2016, 35, 2237–2245. [Google Scholar] [CrossRef] [Green Version]
- da Silva, S.D.; Marchi, F.A.; Xu, B.; Bijian, K.; Alobaid, F.; Mlynarek, A.; Rogatto, S.R.; Hier, M.; Kowalski, L.P.; Alaoui-Jamali, M.A. Predominant Rab-GTPase Amplicons Contributing to Oral Squamous Cell Carcinoma Progression to Metastasis. Oncotarget 2015, 6, 21950–21963. [Google Scholar] [CrossRef]
- Lo Re, O.; Fusilli, C.; Rappa, F.; Van Haele, M.; Douet, J.; Pindjakova, J.; Rocha, S.W.; Pata, I.; Valčíková, B.; Uldrijan, S.; et al. Induction of Cancer Cell Stemness by Depletion of Macrohistone H2A1 in Hepatocellular Carcinoma. Hepatology 2018, 67, 636–650. [Google Scholar] [CrossRef]
- Niu, W.; Zhang, M.; Chen, H.; Wang, C.; Shi, N.; Jing, X.; Ge, L.; Chen, T.; Tang, X. Peroxiredoxin 1 Promotes Invasion and Migration by Regulating Epithelial-to-Mesenchymal Transition during Oral Carcinogenesis. Oncotarget 2016, 7, 47042–47051. [Google Scholar] [CrossRef]
- Jiang, Y.; Cao, W.; Wu, K.; Qin, X.; Wang, X.; Li, Y.; Yu, B.; Zhang, Z.; Wang, X.; Yan, M.; et al. LncRNA LINC00460 Promotes EMT in Head and Neck Squamous Cell Carcinoma by Facilitating Peroxiredoxin-1 into the Nucleus. J. Exp. Clin. Cancer Res. 2019, 38, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Monisha, J.; Kishor Roy, N.; Bordoloi, D.; Kumar, A.; Golla, R.; Kotoky, J.; Padmavathi, G.; B. Kunnumakkara, A. Nuclear Factor Kappa B: A Potential Target to Persecute Head and Neck Cancer. Curr. Drug Targets 2016, 18, 232–253. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Characteristics | Race | |
|---|---|---|
| Black | White | |
| Average Age (years, ±SD) | 53.6 (±9.4) | 64.3 (±14.4) # |
| Ratio of Men:Women (n) | 5:4 | 8:2 |
| Alcohol Use | 100% | 100% |
| Current Smoker | 33.3% | 20% |
| Ever Smoker | 66.7% | 30% |
| Body Mass Index (mean kg/m2) | 27.1 | 24.2 |
| Fruit & Vegetable Servings/Daily | 2.0 | 2.1 |
| Sun Protection, Never or Seldom | 89.9% | 20.0% |
| Sun Protection, Always or Mostly | 11.1% | 80.0% # |
| Middle School or Some High School (n) | 22.2% (2) | 0% |
| High School Graduate (n) | 55.6% (5) | 10% (1) |
| Some College (n) | 11.1% (1) | 30.0% (3) |
| College Graduate or Professional Degree (n) | 11.1% (1) | 60.0% (6) # |
| Measurement | Race | p-Value | |
|---|---|---|---|
| Black | White | ||
| 25-Hydroxyvitamin D (ng/mL) | 20.00 (±5.98) | 27.30 (±9.86) | 0.04 |
| 1,25-Dihydroxyvitamin D (pg/mL) | 43.86 (±20.80) | 36.80 (±14.33) | 0.21 |
| Patients with Low Vitamin D (<19.9 ng/mL) | 50% | 10% | |
| Patients with Intermediate Vitamin D (20.0–29.9 ng/mL) | 50% | 60% | |
| Patients with Sufficient Vitamin D (≥30 ng/mL) | 0% | 30% | 0.06 |
| Parathyroid (pg/mL) | 35.50 (±23.95) | 35.89 (±14.93) | 0.48 |
| Calcium (pg/mL) | 9.34 (0.58) | 9.09 (±0.68) | 0.21 |
| Measurement | Race | p-Value | |
|---|---|---|---|
| Black | White | ||
| Total Vitamin D (IU) | 174.70 (±154.37) | 276.93 (±206.03) | 0.23 |
| Dietary Vitamin D (IU) | 121.24 (±113.77) | 62.92 (±40.10) | 0.07 |
| Supplemental Vitamin D (IU) | 88.89 (±176.38) | 200.00 (±210.82) | 0.12 |
| Patients Supplementing Vitamin D | 22.2% | 50.0% | 0.21 |
| Total Calcium (mg) | 678.40 (±450.42) | 542.93 (±280.74) | 0.22 |
| Dietary Calcium (mg) | 535.16 (±317.30) | 447.87 (±286.89) | 0.27 |
| Supplemental Calcium (mean, mg) | 25.28 (±56.10) | 83.69 (±82.94) | 0.31 |
| Patients Supplementing Calcium | 44.4% | 60.0% | 0.34 |
| Functional Parameter | MiR Alteration | Term (n) | p-Value | Benjamini-Hochberg FDR |
|---|---|---|---|---|
| KEGG Pathway | Up-regulated | Pathways in Cancer (9) | 0.01 | 0.90 |
| Down-regulated | Steroid Biosynthesis (13) | 7.72E-06 | 2.2E-03 | |
| Biological Process | Up-regulated | Positive regulation of transcription from RNA polymerase II promoter (18) | 3.31E-03 | 0.97 |
| Down-regulated | Negative regulation of cell proliferation (95) | 5.27E-06 | 0.03 | |
| Molecular Function | Up-regulated | Protein binding (96) | 1.50E-04 | 0.04 |
| Down-regulated | Transcription factor activity, sequence-specific DNA binding (194) | 1.47E-05 | 0.03 | |
| Diseases | Up-regulated | Cancer (95) | 4.30 E-04 | 7.71E-03 |
| Down-regulated | Chemodependency (736) | 2.91E-12 | 5.53E-11 |
| Cells | Spot | Protein | Symbol | Fold-Change | Uniprot No. |
|---|---|---|---|---|---|
| SCC-25 | 15/16 * | Nucleophosmin | NPM_HUMAN | −2.36 | P06748 |
| 28 | Lactoylglutathione lyase | LGUL_HUMAN | −2.73 | Q04760 | |
| 31 | Heat shock protein beta-1 | HSPB1_HUMAN | −2.57 | P04792 | |
| 51 | Ras-related protein Rap-2b | RAP2B_HUMAN | −3.39 | P61225 | |
| CAL-27 | 30 | Peroxiredoxin-1 | PRDX1_HUMAN | −2.60 | Q06830 |
| 31 | Histone H2A type 1-J | H2A1J_HUMAN | +2.48 | Q99878 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibrahimovic, M.; Franzmann, E.; Mondul, A.M.; Weh, K.M.; Howard, C.; Hu, J.J.; Goodwin, W.J.; Kresty, L.A. Disparities in Head and Neck Cancer: A Case for Chemoprevention with Vitamin D. Nutrients 2020, 12, 2638. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12092638
Ibrahimovic M, Franzmann E, Mondul AM, Weh KM, Howard C, Hu JJ, Goodwin WJ, Kresty LA. Disparities in Head and Neck Cancer: A Case for Chemoprevention with Vitamin D. Nutrients. 2020; 12(9):2638. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12092638
Chicago/Turabian StyleIbrahimovic, Mirela, Elizabeth Franzmann, Alison M. Mondul, Katherine M. Weh, Connor Howard, Jennifer J. Hu, W. Jarrard Goodwin, and Laura A. Kresty. 2020. "Disparities in Head and Neck Cancer: A Case for Chemoprevention with Vitamin D" Nutrients 12, no. 9: 2638. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12092638