Adipose Tissue Development and Expansion from the Womb to Adolescence: An Overview

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Adipose Tissue Development
2.1. Intrauterine Adipose Tissue Accrual
2.1.1. Health and Nutrition during Pregnancy as Determinants of Adipose Tissue
2.1.2. Effects of Intrauterine Exposure to Toxins on Adipose Tissue Development
2.1.3. Genetic Defects
2.2. Postnatal and Infant Adipose Tissue Accrual
2.2.1. Associations between Feeding Practices and Adipose Tissue in Infancy
2.2.2. Gut Microbiome
2.3. Adipose Tissue Development in Childhood and Adolescence
2.3.1. Hormonal Influences on Adipose Tissue Expansion among Boys and Girls
2.3.2. Dietary Intake and Interactions with Gut Microbiome
2.3.3. Physical Activity
3. Associations between Adipose Tissue in Childhood and Adulthood
4. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- NCD Risk Factor Collaboration (NCD-RisC). Worldwide trends in body-mass index, underweight, overweight, and obesity from 1975 to 2016: A pooled analysis of 2416 population-based measurement studies in 128·9 million children, adolescents, and adults. Lancet 2017, 390, 2627–2642. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Kelly, A.S. Review of childhood obesity: From epidemiology, etiology, and comorbidities to clinical assessment and treatment. Mayo Clin. Proc. 2017, 92, 251–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinonen, S.; Jokinen, R.; Rissanen, A.; Pietiläinen, K.H. White adipose tissue mitochondrial metabolism in health and in obesity. Obes. Rev. 2019, 21, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Min, S.Y.; Desai, A.; Yang, Z.; Sharma, A.; DeSouza, T.; Genga, R.M.; Kucukural, A.; Lifshitz, L.M.; Nielsen, S.; Scheele, C.; et al. Diverse repertoire of human adipocyte subtypes develops from transcriptionally distinct mesenchymal progenitor cells. Proc. Natl. Acad. Sci. USA 2019, 116, 17970–17979. [Google Scholar] [CrossRef] [Green Version]
- Virtanen, K.A.; Lidell, M.E.; Orava, J.; Heglind, M.; Westergren, R.; Niemi, T.; Taittonen, M.; Laine, J.; Savisto, N.J.; Enerbäck, S.; et al. Functional brown adipose tissue in healthy adults. N. Engl. J. Med. 2009, 360, 1518–1525. [Google Scholar] [CrossRef]
- Kindler, J.M.; Lobene, A.J.; Vogel, K.A.; Martin, B.R.; McCabe, L.D.; Peacock, M.; Warden, S.J.; McCabe, G.P.; Weaver, C.M. Adiposity, insulin resistance, and bone mass in children and adolescents. J. Clin. Endocrinol. Metab. 2018, 104, 892–899. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.J.; Arslanian, S.A. Cardiorespiratory fitness and abdominal adiposity in youth. Eur. J. Clin. Nutr. 2007, 61, 561–565. [Google Scholar] [CrossRef] [Green Version]
- Schvey, N.A.; Marwitz, S.E.; Mi, S.J.; Galescu, O.A.; Broadney, M.M.; Young-Hyman, D.; Brady, S.M.; Reynolds, J.C.; Tanofsky-Kraff, M.; Yanovski, S.Z.; et al. Weight-based teasing is associated with gain in BMI and fat mass among children and adolescents at-risk for obesity: A longitudinal study. Pediatr. Obes. 2019, 14, 1–14. [Google Scholar] [CrossRef]
- Lenz, M.; Arts, I.C.; Peeters, R.L.; de Kok, T.M.; Ertaylan, G. Adipose tissue in health and disease through the lens of its building blocks. Sci. Rep. 2020, 10, 1–4. [Google Scholar] [CrossRef]
- Cleal, L.; Aldea, T.; Chau, Y.Y. Fifty shades of white: Understanding heterogeneity in white adipose stem cells. Adipocyte 2017, 6, 205–216. [Google Scholar] [CrossRef] [Green Version]
- Haylett, W.L.; Ferris, W.F. Adipocyte–progenitor cell communication that influences adipogenesis. Cell Mol. Life Sci. 2020, 77, 115–128. [Google Scholar] [CrossRef]
- Laharrague, P.; Casteilla, L. The emergence of adipocytes. In Adipose Tissue Development: From Animal Models to Clinical Conditions; Levy-Marchal, C., Pénicaud, L., Eds.; Karger Medical and Scientific Publishers: Basel, Switzerland, 2010; Volume 19, pp. 21–30. [Google Scholar]
- Hao, G.; Wang, X.; Treiber, F.A.; Harshfield, G.; Kapuku, G.; Su, S. Body mass index trajectories in childhood is predictive of cardiovascular risk: Results from the 23-year longitudinal Georgia Stress and Heart study. Int. J. Obes. 2018, 42, 923–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koontz, M.B.; Gunzler, D.D.; Presley, L.; Catalano, P.M. Longitudinal changes in infant body composition: Association with childhood obesity. Pediatr. Obes. 2014, 9, e141–e144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ong, K.K.; Emmett, P.; Northstone, K.; Golding, J.; Rogers, I.; Ness, A.R.; Wells, J.C.; Dunger, D.B. Infancy weight gain predicts childhood body fat and age at menarche in girls. J. Clin. Endocrinol. Metab. 2009, 94, 1527–1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chomtho, S.; Wells, J.C.; Williams, J.E.; Davies, P.S.; Lucas, A.; Fewtrell, M.S. Infant growth and later body composition: Evidence from the 4-component model. Am. J. Clin. Nutr. 2008, 87, 1776–1784. [Google Scholar] [CrossRef] [PubMed]
- Admassu, B.; Ritz, C.; Wells, J.C.; Girma, T.; Andersen, G.S.; Belachew, T.; Owino, V.; Michaelsen, K.F.; Abera, M.; Wibaek, R.; et al. Accretion of fat-free mass rather than fat mass in infancy is positively associated with linear growth in childhood. J. Nutr. 2018, 148, 607–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, W.; St-Onge, M.; Wang, Z.; Heymsfield, S. Study of body composition: An overview. In Human Body Composition, 2nd ed.; Heymsfield, S., Lohman, T., Wang, Z., Going, S., Eds.; Human Kinetics: Champaign, IL, USA, 2005; pp. 3–14. [Google Scholar]
- Lee, M.J.; Wu, Y.; Fried, S.K. Adipose tissue heterogeneity: Implication of depot differences in adipose tissue for obesity complications. Mol. Aspects Med. 2013, 34, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Jung, S.M.; Sanchez-Gurmaches, J.; Guertin, D.A. Brown adipose tissue development and metabolism. In Brown Adipose Tissue; Springer: Cham, Switzerland, 2018; pp. 3–36. [Google Scholar]
- Lecoutre, S.; Petrus, P.; Rydén, M.; Breton, C. Transgenerational epigenetic mechanisms in adipose tissue development. Trends Endocrinol. Metab. 2018, 29, 675–685. [Google Scholar] [CrossRef]
- Sun, K.; Kusminski, C.M.; Scherer, P.E. Adipose tissue remodeling and obesity. J. Clin. Investig. 2011, 121, 2094–2101. [Google Scholar] [CrossRef] [Green Version]
- Fomon, S.J.; Haschke, F.; Ziegler, E.E.; Nelson, S.E. Body composition of reference children from birth to age 10 years. Am. J. Clin. Nutr. 1982, 35, 1169–1175. [Google Scholar] [CrossRef]
- Ellis, K.J. Human body composition: In vivo methods. Physiol. Rev. 2000, 80, 649–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sethi, J.K.; Vidal-Puig, A.J. Thematic review series: Adipocyte biology. Adipose tissue function and plasticity orchestrate nutritional adaptation. J. Lipid Res. 2007, 48, 1253–1262. [Google Scholar] [PubMed] [Green Version]
- Nouws, J.; Fitch, M.; Mata, M.; Santoro, N.; Galuppo, B.; Kursawe, R.; Narayan, D.; Vash-Margita, A.; Pierpont, B.; Shulman, G.I.; et al. Altered in vivo lipid fluxes and cell dynamics in subcutaneous adipose tissues are associated with the unfavorable pattern of fat distribution in obese adolescent girls. Diabetes 2019, 68, 1168–1177. [Google Scholar] [CrossRef] [PubMed]
- Poissonnet, C.M.; Burdi, A.R.; Bookstein, F.L. Growth and development of human adipose tissue during early gestation. Early Hum. Dev. 1983, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Billon, N.; Monteiro, M.C.; Dani, C. Developmental origin of adipocytes: New insights into a pending question. Biol. Cell 2008, 100, 563–575. [Google Scholar] [CrossRef]
- Ziegler, E.E.; O’donnell, A.M.; Nelson, S.E.; Fomon, S.J. Body composition of the reference fetus. Growth 1976, 40, 329–341. [Google Scholar]
- Toro-Ramos, T.; Paley, C.; Pi-Sunyer, F.X.; Gallagher, D. Body composition during fetal development and infancy through the age of 5 years. Eur. J. Clin. Nutr. 2015, 69, 1279–1289. [Google Scholar] [CrossRef] [Green Version]
- Berger-Kulemann, V.; Brugger, P.C.; Reisegger, M.; Klein, K.; Hachemian, N.; Koelblinger, C.; Weber, M.; Prayer, D. Quantification of the subcutaneous fat layer with MRI in fetuses of healthy mothers with no underlying metabolic disease vs. fetuses of diabetic and obese mothers. J. Perinat. Med. 2012, 40, 179–184. [Google Scholar]
- Kwon, E.J.; Kim, Y.J. What is fetal programming?: A lifetime health is under the control of in utero health. Obs. Gynecol. Sci. 2017, 60, 506–519. [Google Scholar] [CrossRef]
- Harding, J.; Johnston, B. Nutrition and fetal growth. Reprod. Fertil. Dev. 1995, 7, 539–547. [Google Scholar] [CrossRef]
- Dalrymple, K.V.; Thompson, J.M.; Begum, S.; Godfrey, K.M.; Poston, L.; Seed, P.T.; McCowan, L.M.; Wall, C.; Shelling, A.; North, R.; et al. Relationships of maternal body mass index and plasma biomarkers with childhood body mass index and adiposity at 6 years: The Children of SCOPE study. Pediatr. Obes. 2019, 14, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Jarvie, E.M.; Stewart, F.M.; Ramsay, J.E.; Brown, E.A.; Meyer, B.J.; Olivecrona, G.; Griffin, B.A.; Freeman, D.J. Maternal adipose tissue expansion, a missing link in the prediction of birth weight centile. J. Clin. Endocrinol. Metab. 2020, 105, e814–e825. [Google Scholar] [CrossRef] [PubMed]
- Tahir, M.J.; Haapala, J.L.; Foster, L.P.; Duncan, K.M.; Teague, A.M.; Kharbanda, E.O.; McGovern, P.M.; Whitaker, K.M.; Rasmussen, K.M.; Fields, D.A.; et al. Higher maternal diet quality during pregnancy and lactation is associated with lower infant weight-for-length, body fat percent, and fat mass in early postnatal life. Nutrients 2019, 11, 632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrestha, N.; Sleep, S.L.; Cuffe, J.S.; Holland, O.J.; Perkins, A.V.; Yau, S.Y.; McAinch, A.J.; Hryciw, D.H. Role of omega-6 and omega-3 fatty acids in fetal programming. Clin. Exp. Pharmacol. Physiol. 2020, 47, 907–915. [Google Scholar] [CrossRef] [Green Version]
- Vahdaninia, M.; Mackenzie, H.; Dean, T.; Helps, S. The effectiveness of ω-3 polyunsaturated fatty acid interventions during pregnancy on obesity measures in the offspring: An up-to-date systematic review and meta-analysis. Eur. J. Nutr. 2019, 58, 2597–2613. [Google Scholar] [CrossRef] [Green Version]
- Lowe, W.L., Jr.; Lowe, L.P.; Kuang, A.; Catalano, P.M.; Nodzenski, M.; Talbot, O.; Tam, W.H.; Sacks, D.A.; McCance, D.; Linder, B.; et al. Maternal glucose levels during pregnancy and childhood adiposity in the Hyperglycemia and Adverse Pregnancy Outcome Follow-up Study. Diabetologia 2019, 62, 598–610. [Google Scholar] [CrossRef] [Green Version]
- Hockett, C.W.; Harrall, K.K.; Moore, B.F.; Starling, A.P.; Bellatorre, A.; Sauder, K.A.; Perng, W.; Scherzinger, A.; Garg, K.; Ringham, B.M.; et al. Persistent effects of in utero overnutrition on offspring adiposity: The Exploring Perinatal Outcomes among Children (EPOCH) study. Diabetologia 2019, 62, 2017–2024. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Pan, L.; Liu, E.; Liu, H.; Liu, J.; Wang, S.; Guo, J.; Li, N.; Zhang, C.; Hu, G. Gestational diabetes and offspring’s growth from birth to 6 years old. Int. J. Obes. 2019, 43, 663–672. [Google Scholar] [CrossRef]
- Hoepner, L.A. Bisphenol A: A narrative review of prenatal exposure effects on adipogenesis and childhood obesity via peroxisome proliferator-activated receptor gamma. Environ. Res. 2019, 173, 54–68. [Google Scholar] [CrossRef]
- Shafei, A.E.; Nabih, E.S.; Shehata, K.A.; Abd Elfatah, E.S.; Sanad, A.B.; Marey, M.Y.; Hammouda, A.A.; Mohammed, M.M.; Mostafa, R.; Ali, M.A. Prenatal exposure to endocrine disruptors and reprogramming of adipogenesis: An early-life risk factor for childhood obesity. Child. Obes. 2018, 14, 18–25. [Google Scholar] [CrossRef]
- Moore, B.F.; Starling, A.P.; Magzamen, S.; Harrod, C.S.; Allshouse, W.B.; Adgate, J.L.; Ringham, B.M.; Glueck, D.H.; Dabelea, D. Fetal exposure to maternal active and secondhand smoking with offspring early-life growth in the Healthy Start study. Int. J. Obes. 2019, 43, 652–662. [Google Scholar] [CrossRef] [PubMed]
- Starling, A.P.; Moore, B.F.; Thomas, D.S.K.; Peel, J.L.; Zhang, W.; Adgate, J.L.; Magzamen, S.; Martenies, S.E.; Allshouse, W.B.; Dabelea, D. Prenatal exposure to traffic and ambient air pollution and infant weight and adiposity: The Healthy Start study. Environ. Res. 2020, 8, 109130. [Google Scholar] [CrossRef] [PubMed]
- Pigeyre, M.; Yazdi, F.T.; Kaur, Y.; Meyre, D. Recent progress in genetics, epigenetics and metagenomics unveils the pathophysiology of human obesity. Clin. Sci. 2016, 130, 943–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pigeyere, M.; Meyre, D. Monogenic obesity. In Pediatric Obesity: Etiology, Pathogenesis and Treatment, 2nd ed.; Freemark, M.S., Ed.; Springer International Publishing: New York, NY, USA, 2018; pp. 135–152. [Google Scholar]
- Irizarry, K.A.; Haqq, A.M. Syndromic obesity. In Pediatric Obesity: Etiology, Pathogenesis and Treatment, 2nd ed.; Freemark, M.S., Ed.; Springer International Publishing: New York, NY, USA, 2018; pp. 153–182. [Google Scholar]
- Marenne, G.; Hendricks, A.E.; Perdikari, A.; Bounds, R.; Payne, F.; Keogh, J.M.; Lelliott, C.J.; Henning, E.; Pathan, S.; Ashford, S.; et al. Exome sequencing identifies genes and gene sets contributing to severe childhood obesity, linking PHIP variants to repressed POMC transcription. Cell Metab. 2020, 31, 1107–1119. [Google Scholar] [CrossRef]
- Hinney, A.; Giuranna, J. Polygenic obesity. In Pediatric Obesity: Etiology, Pathogenesis and Treatment, 2nd ed.; Freemark, M.S., Ed.; Springer International Publishing: New York, NY, USA, 2018; pp. 183–204. [Google Scholar]
- Moulton, C.R. Age and chemical development in mammals. J. Biol. Chem. 1923, 57, 79–97. [Google Scholar]
- Griffin, I.J.; Cooke, R.J. Development of whole body adiposity in preterm infants. Early Hum. Dev. 2012, 88, S19–S24. [Google Scholar] [CrossRef]
- Roggero, P.; Giannì, M.L.; Amato, O.; Orsi, A.; Piemontese, P.; Morlacchi, L.; Mosca, F. Is term newborn body composition being achieved postnatally in preterm infants? Early Hum. Dev. 2009, 85, 349–352. [Google Scholar] [CrossRef]
- Ramel, S.E.; Gray, H.L.; Ode, K.L.; Younge, N.; Georgieff, M.K.; Demerath, E.W. Body composition changes in preterm infants following hospital discharge: Comparison with term infants. J. Pediatr. Gastroenterol. Nutr. 2011, 53, 333–338. [Google Scholar] [CrossRef]
- Sebastiani, G.; García-Beltran, C.; Pie, S.; Guerra, A.; López-Bermejo, A.; de Toledo, J.S.; de Zegher, F.; Rosés, F.; Ibáñez, L. The sequence of prenatal growth restraint and postnatal catch-up growth: Normal heart but thicker intima-media and more pre-peritoneal fat in late infancy. Pediatr. Obes. 2019, 14, e12476. [Google Scholar] [CrossRef]
- De Zegher, F.; Díaz, M.; Sebastiani, G.; Martín-Ancel, A.; Sánchez-Infantes, D.; López-Bermejo, A.; Ibáñez, L. Abundance of circulating preadipocyte factor 1 in early life. Diabetes Care 2012, 35, 848–849. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Kim, K.A.; Kim, J.H.; Sul, H.S. Pref-1, a preadipocyte secreted factor that inhibits adipogenesis. J. Nutr. 2006, 136, 2953–2956. [Google Scholar] [CrossRef] [PubMed]
- Vogelezang, S.; Santos, S.; Toemen, L.; Oei, E.H.; Felix, J.F.; Jaddoe, V.W. Associations of fetal and infant weight change with general, visceral, and organ adiposity at school age. JAMA Netw. Open 2019, 2, e192843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parra-Vargas, M.; Ramon-Krauel, M.; Lerin, C.; Jimenez-Chillaron, J.C. Size does matter: Litter size strongly determines adult metabolism in rodents. Cell Metab. 2020, 32, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Victora, C.G.; Bahl, R.; Barros, A.J.; França, G.V.; Horton, S.; Krasevec, J.; Murch, S.; Sankar, M.J.; Walker, N.; Rollins, N.C. The Lancet Breastfeeding Series Group. Breastfeeding in the 21st century: Epidemiology, mechanisms, and lifelong effect. Lancet 2016, 387, 475–490. [Google Scholar] [CrossRef] [Green Version]
- Rollins, N.C.; Bhandari, N.; Hajeebhoy, N.; Horton, S.; Lutter, C.K.; Martines, J.C.; Piwoz, E.G.; Richter, L.M.; Victora, C.G. The Lancet Breastfeeding Series Group. Why invest, and what it will take to improve breastfeeding practices? Lancet 2016, 387, 491–504. [Google Scholar] [CrossRef]
- Breij, L.M.; Abrahamse-Berkeveld, M.; Acton, D.; Rolfe, E.D.; Ong, K.K.; Hokken-Koelega, A.C. Impact of early infant growth, duration of breastfeeding and maternal factors on total body fat mass and visceral fat at 3 and 6 months of age. Ann. Nutr. Metab. 2017, 71, 203–210. [Google Scholar] [CrossRef]
- Rodríguez-Cano, A.M.; Mier-Cabrera, J.; Allegre-Dávalos, A.L.; Muñoz-Manrique, C.; Perichart-Perera, O. Higher fat mass and fat mass accretion during the first six months of life in exclusively breastfed infants. Pediatr. Res. 2020, 87, 588–594. [Google Scholar] [CrossRef]
- Bell, K.A.; Wagner, C.L.; Feldman, H.A.; Shypailo, R.J.; Belfort, M.B. Associations of infant feeding with trajectories of body composition and growth. Am. J. Clin. Nutr. 2017, 106, 491–498. [Google Scholar] [CrossRef]
- De Zegher, F.; Sebastiani, G.; Diaz, M.; Gómez-Roig, M.D.; López-Bermejo, A.; Ibáñez, L. Breast-feeding vs formula-feeding for infants born small-for-gestational-age: Divergent effects on fat mass and on circulating IGF-I and high-molecular-weight adiponectin in late infancy. J. Clin. Endocrinol. Metab. 2013, 98, 1242–1247. [Google Scholar] [CrossRef] [Green Version]
- Gridneva, Z.; Rea, A.; Tie, W.J.; Lai, C.T.; Kugananthan, S.; Ward, L.C.; Murray, K.; Hartmann, P.E.; Geddes, D.T. Carbohydrates in human milk and body composition of term infants during the first 12 months of lactation. Nutrients 2019, 11, 1472. [Google Scholar] [CrossRef] [Green Version]
- Rudolph, M.C.; Young, B.E.; Lemas, D.J.; Palmer, C.E.; Hernandez, T.L.; Barbour, L.A.; Friedman, J.E.; Krebs, N.F.; MacLean, P.S. Early infant adipose deposition is positively associated with the n-6 to n-3 fatty acid ratio in human milk independent of maternal BMI. Int. J. Obes. 2017, 41, 510–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaidya, H.; Cheema, S.K. Breastmilk with a high omega-6 to omega-3 fatty acid ratio induced cellular events similar to insulin resistance and obesity in 3T3-L1 adipocytes. Pediatric. Obes. 2018, 13, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Chan, D.; Goruk, S.; Becker, A.B.; Subbarao, P.; Mandhane, P.J.; Turvey, S.E.; Lefebvre, D.; Sears, M.R.; Field, C.J.; Azad, M.B. Adiponectin, leptin and insulin in breast milk: Associations with maternal characteristics and infant body composition in the first year of life. Int. J. Obes. (Lond.) 2018, 42, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Gingras, V.; Aris, I.M.; Rifas-Shiman, S.L.; Switkowski, K.M.; Oken, E.; Hivert, M.F. Timing of complementary feeding introduction and adiposity throughout childhood. Pediatrics 2019, 144, e20191320. [Google Scholar] [CrossRef] [PubMed]
- Bäckhed, F.; Roswall, J.; Peng, Y.; Feng, Q.; Jia, H.; Kovatcheva-Datchary, P.; Li, Y.; Xia, Y.; Xie, H.; Zhong, H.; et al. Dynamics and stabilization of the human gut microbiome during the first year of life. Cell Host Microbe 2015, 17, 690–703. [Google Scholar] [CrossRef] [Green Version]
- Mueller, N.T.; Whyatt, R.; Hoepner, L.; Oberfield, S.; Dominguez-Bello, M.G.; Widen, E.M.; Hassoun, A.; Perera, F.; Rundle, A. Prenatal exposure to antibiotics, cesarean section and risk of childhood obesity. Int. J. Obes. 2015, 39, 665–670. [Google Scholar] [CrossRef] [Green Version]
- Tun, H.M.; Bridgman, S.L.; Chari, R.; Field, C.J.; Guttman, D.S.; Becker, A.B.; Mandhane, P.J.; Turvey, S.E.; Subbarao, P.; Sears, M.R.; et al. Roles of birth mode and infant gut microbiota in intergenerational transmission of overweight and obesity from mother to offspring. JAMA Pediatr. 2018, 172, 368–377. [Google Scholar] [CrossRef]
- Miller, S.A.; Wu, R.K.S.; Oremus, M. The association between antibiotic use in infancy and childhood overweight or obesity: A systematic review and meta-analysis. Obes. Rev. 2018, 19, 1463–1475. [Google Scholar] [CrossRef]
- Forbes, J.D.; Azad, M.B.; Vehling, L.; Tun, H.M.; Konya, T.B.; Guttman, D.S.; Field, C.J.; Lefebvre, D.; Sears, M.R.; Becker, A.B.; et al. Association of exposure to formula in the hospital and subsequent infant feeding practices with gut microbiota and risk of overweight in the first year of life. JAMA Pediatr. 2018, 172, e181161. [Google Scholar] [CrossRef]
- Caesar, R.; Tremaroli, V.; Kovatcheva-Datchary, P.; Cani, P.D.; Bäckhed, F. Crosstalk between gut microbiota and dietary lipids aggravates WAT inflammation through TLR signaling. Cell Metab. 2015, 22, 658–668. [Google Scholar] [CrossRef] [Green Version]
- Cho, I.; Yamanishi, S.; Cox, L.; Methe, B.A.; Zavadil, J.; Li, K.; Gao, Z.; Mahana, D.; Raju, K.; Teitler, I.; et al. Antibiotics in early life alter the murine colonic microbiome and adiposity. Nature 2012, 488, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Kincaid, H.J.; Nagpal, R.; Yadav, H. Microbiome-immune-metabolic axis in the epidemic of childhood obesity: Evidence and opportunities. Obes. Rev. 2020, 21, e12963. [Google Scholar] [CrossRef] [PubMed]
- Soderborg, T.K.; Clark, S.E.; Mulligan, C.E.; Janssen, R.C.; Babcock, L.; Ir, D.; Young, B.; Krebs, N.; Lemas, D.J.; Johnson, L.K.; et al. The gut microbiota in infants of obese mothers increases inflammation and susceptibility to NAFLD. Nat. Commun. 2018, 9, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Gallagher, D.; Andres, A.; Fields, D.A.; Evans, W.J.; Kuczmarski, R.; Lowe, W.L., Jr.; Lumeng, J.C.; Oken, E.; Shepherd, J.A.; Sun, S.; et al. Body composition measurements from birth through 5 years: Challenges, gaps, and existing & emerging technologies-A National Institutes of Health workshop. Obes. Rev. 2020, 21, e13033. [Google Scholar] [PubMed]
- Orsso, C.E.; Silva, M.I.; Gonzalez, M.C.; Rubin, D.A.; Heymsfield, S.B.; Prado, C.M.; Haqq, A.M. Assessment of body composition in pediatric overweight and obesity: A systematic review of the reliability and validity of common techniques. Obes. Rev. 2020, 21, e13041. [Google Scholar] [CrossRef] [PubMed]
- Wells, J.C.; Davies, P.S.; Fewtrell, M.S.; Cole, T.J. Body composition reference charts for UK infants and children aged 6 weeks to 5 years based on measurement of total body water by isotope dilution. Eur. J. Clin. Nutr. 2020, 74, 141–148. [Google Scholar] [CrossRef] [Green Version]
- McConnell-Nzunga, J.; Naylor, P.J.; Macdonald, H.M.; Rhodes, R.E.; Hofer, S.M.; McKay, H.A. Body fat accrual trajectories for a sample of Asian-Canadian and Caucasian-Canadian children and youth: A longitudinal DXA-based study. Pediatr. Obes. 2020, 15, e12570. [Google Scholar] [CrossRef]
- McCarthy, H.D.; Cole, T.J.; Fry, T.; Jebb, S.A.; Prentice, A.M. Body fat reference curves for children. Int. J. Obes. 2006, 30, 598–602. [Google Scholar] [CrossRef] [Green Version]
- Ripka, W.L.; Orsso, C.E.; Haqq, A.M.; Luz, T.G.; Prado, C.M.; Ulbricht, L. Lean mass reference curves in adolescents using dual-energy x-ray absorptiometry (DXA). PLoS ONE 2020, 15, e0228646. [Google Scholar] [CrossRef]
- Taylor, R.W.; Grant, A.M.; Williams, S.M.; Goulding, A. Sex differences in regional body fat distribution from pre- to postpuberty. Obesity 2010, 18, 1410–1416. [Google Scholar] [CrossRef]
- Shen, W.; Punyanitya, M.; Silva, A.M.; Chen, J.; Gallagher, D.; Sardinha, L.B.; Allison, D.B.; Heymsfield, S.B. Sexual dimorphism of adipose tissue distribution across the lifespan: A cross-sectional whole-body magnetic resonance imaging study. Nutr. Metab. (Lond.) 2009, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, H.S.; Lancaster, J.B. An evolutionary and ecological analysis of human fertility, mating patterns, and parental investment. In Offspring: Human Fertility Behavior in Biodemographic Perspective Panel; Wachter, K., Bulatao, R., Eds.; National Academy of Sciences: Washington, DC, USA, 2003; pp. 170–223. [Google Scholar]
- Chang, E.; Varghese, M.; Singer, K. Gender and sex differences in adipose tissue. Curr. Diabetes Rep. 2018, 18, 69. [Google Scholar] [CrossRef]
- Widen, E.M.; Gallagher, D. Body composition changes in pregnancy: Measurement, predictors and outcomes. Eur. J. Clin. Nutr. 2014, 68, 643–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopp-Hoolihan, L.E.; Van Loan, M.D.; Wong, W.W.; King, J.C. Fat mass deposition during pregnancy using a four-component model. J. Appl. Physiol. 1999, 87, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Lederman, S.A.; Paxton, A.; Heymsfield, S.B.; Wang, J.; Thornton, J.; Pierson, R.N., Jr. Body fat and water changes during pregnancy in women with different body weight and weight gain. Obstet. Gynecol. 1997, 90, 483–488. [Google Scholar] [CrossRef]
- Bosaeus, M.; Andersson-Hall, U.; Andersson, L.; Karlsson, T.; Ellegård, L.; Holmäng, A. Body composition during pregnancy: Longitudinal changes and method comparisons. Reprod. Sci. 2020, 28, 1–3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lassek, W.D.; Gaulin, S.J. Changes in body fat distribution in relation to parity in American women: A covert form of maternal depletion. Am. J. Phys. Anthropol. 2006, 131, 295–302. [Google Scholar] [CrossRef]
- Scholl, T.O.; Hediger, M.L.; Schall, J.I.; Khoo, C.S.; Fischer, R.L. Maternal growth during pregnancy and the competition for nutrients. Am. J. Clin. Nutr. 1994, 60, 183–188. [Google Scholar] [CrossRef]
- Palmer, B.F.; Clegg, D.J. The sexual dimorphism of obesity. Mol. Cell Endocrinol. 2015, 15, 113–119. [Google Scholar] [CrossRef] [Green Version]
- Brown, L.; Clegg, D. Central effects of estradiol in the regulation of adiposity. J. Steroid Biochem. Mol. Biol. 2010, 122, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Currie, C.; Ahluwalia, N.; Godeau, E.; Gabhainn, S.N.; Due, P.; Currie, D.B. Is obesity at individual and national level associated with lower age at menarche? Evidence from 34 countries in the health behaviour in school-aged children study. J. Adolesc. Health 2012, 50, 621–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieuwenhuis, D.; Pujol-Gualdo, N.; Arnoldussen, I.A.; Kiliaan, A.J. Adipokines: A gear shift in puberty. Obes. Rev. 2020, 21, e13005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wosje, K.S.; Khoury, P.R.; Claytor, R.P.; Copeland, K.A.; Hornung, R.W.; Daniels, S.R.; Kalkwarf, H.J. Dietary patterns associated with fat and bone mass in young children. Am. J. Clin. Nutr. 2010, 92, 294–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suissa, K.; Benedetti, A.; Henderson, M.; Gray-Donald, K.; Paradis, G. Effects of dietary glycemic index and load on children’s cardiovascular risk factors. Ann. Epidemiol. 2019, 40, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Setayeshgar, S.; Maximova, K.; Ekwaru, J.P.; Gray-Donald, K.; Henderson, M.; Paradis, G.; Tremblay, A.; Veugelers, P. Diet quality as measured by the Diet Quality Index-International is associated with prospective changes in body fat among Canadian children. Public Health Nutr. 2017, 20, 456–463. [Google Scholar] [CrossRef] [Green Version]
- Nguyen-Rodriguez, S.T.; Gallo, L.C.; Isasi, C.R.; Buxton, O.M.; Thomas, K.S.; Sotres-Alvarez, D.; Redline, S.; Castañeda, S.F.; Carnethon, M.R.; Daviglus, M.L.; et al. Adiposity, depression symptoms and inflammation in Hispanic/Latino youth: Results from HCHS/SOL youth. Ann. Behav. Med. 2020, 54, 529–534. [Google Scholar] [CrossRef]
- Boswell, N.; Byrne, R.; Davies, P.S.W. Aetiology of eating behaviours: A possible mechanism to understand obesity development in early childhood. Neurosci. Biobehav. Rev. 2018, 95, 438–448. [Google Scholar] [CrossRef]
- Appelhans, B.M. Neurobehavioral inhibition of reward-driven feeding: Implications for dieting and obesity. Obesity 2009, 17, 640–647. [Google Scholar] [CrossRef]
- Campbell, K.J.; Crawford, D.A.; Salmon, J.; Carver, A.; Garnett, S.P.; Baur, L.A. Associations between the home food environment and obesity-promoting eating behaviors in adolescence. Obesity 2007, 15, 719–730. [Google Scholar] [CrossRef]
- Hill, D.C.; Moss, R.H.; Sykes-Muskett, B.; Conner, M.; O’Connor, D.B. Stress and eating behaviors in children and adolescents: Systematic review and meta-analysis. Appetite 2018, 123, 14–22. [Google Scholar] [CrossRef]
- Fuemmeler, B.F.; Sheng, Y.; Schechter, J.C.; Do, E.; Zucker, N.; Majors, A.; Maguire, R.; Murphy, S.K.; Hoyo, C.; Kollins, S.H. Associations between attention deficit hyperactivity disorder symptoms and eating behaviors in early childhood. Pediatr. Obes. 2020, 15, e12631. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Cai, Z.; Fan, X. Prevalence of binge and loss of control eating among children and adolescents with overweight and obesity: An exploratory meta-analysis. Int. J. Eat. Disord. 2017, 50, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Nicolucci, A.C.; Hume, M.P.; Martínez, I.; Mayengbam, S.; Walter, J.; Reimer, R.A. Prebiotics reduce body fat and alter intestinal microbiota in children who are overweight or with obesity. Gastroenterology 2017, 153, 711–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, E.S.; Viardot, A.; Psichas, A.; Morrison, D.J.; Murphy, K.G.; Zac-Varghese, S.E.; MacDougall, K.; Preston, T.; Tedford, C.; Finlayson, G.S.; et al. Effects of targeted delivery of propionate to the human colon on appetite regulation, body weight maintenance and adiposity in overweight adults. Gut 2015, 64, 1744–1754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larraufie, P.; Martin-Gallausiaux, C.; Lapaque, N.; Dore, J.; Gribble, F.M.; Reimann, F.; Blottiere, H.M. SCFAs strongly stimulate PYY production in human enteroendocrine cells. Sci. Rep. 2018, 8, 74. [Google Scholar] [CrossRef]
- Willemsen, L.E.; Koetsier, M.A.; Van Deventer, S.J.; Van Tol, E.A. Short chain fatty acids stimulate epithelial mucin 2 expression through differential effects on prostaglandin E(1) and E(2) production by intestinal myofibroblasts. Gut 2003, 52, 1442–1447. [Google Scholar] [CrossRef] [Green Version]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Fava, F.; Tuohy, K.M.; Chabo, C.; et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007, 56, 1232–1242. [Google Scholar] [CrossRef] [Green Version]
- Borgo, F.; Verduci, E.; Riva, A.; Lassandro, C.; Riva, E.; Morace, G.; Borghi, E. Relative abundance in bacterial and fungal gut microbes in obese children: A case control study. Child. Obes. 2017, 13, 78–84. [Google Scholar] [CrossRef]
- López-Contreras, B.E.; Morán-Ramos, S.; Villarruel-Vázquez, R.; Macías-Kauffer, L.; Villamil-Ramírez, H.; León-Mimila, P.; Vega-Badillo, J.; Sánchez-Muñoz, F.; Llanos-Moreno, L.E.; Canizalez-Román, A.; et al. Composition of gut microbiota in obese and normal-weight Mexican school-age children and its association with metabolic traits. Pediatr. Obes. 2018, 13, 381–388. [Google Scholar] [CrossRef]
- Indiani, C.M.; Rizzardi, K.F.; Castelo, P.M.; Ferraz, L.F.; Darrieux, M.; Parisotto, T.M. Childhood obesity and Firmicutes/Bacteroidetes ratio in the gut microbiota: A systematic review. Child. Obes. 2018, 14, 501–509. [Google Scholar] [CrossRef]
- Mbakwa, C.A.; Hermes, G.D.; Penders, J.; Savelkoul, P.H.; Thijs, C.; Dagnelie, P.C.; Mommers, M.; Zoetendal, E.G.; Smidt, H.; Arts, I.C. Gut microbiota and body weight in school-aged children: The KOALA birth cohort study. Obesity 2018, 26, 1767–1776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamo, K.B.; Colley, R.C.; Hadjiyannakis, S.; Goldfield, G.S. Physical activity and sedentary behavior in obese youth. J. Pediatr. 2015, 166, 1270–1275. [Google Scholar] [CrossRef]
- Herman, K.M.; Sabiston, C.M.; Mathieu, M.E.; Tremblay, A.; Paradis, G. Sedentary behavior in a cohort of 8- to 10-year-old children at elevated risk of obesity. Prev. Med. (Baltim) 2014, 60, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Wiersma, R.; Haverkamp, B.F.; van Beek, J.H.; Riemersma, A.M.; Boezen, H.M.; Smidt, N.; Corpeleijn, E.; Hartman, E. Unravelling the association between accelerometer-derived physical activity and adiposity among preschool children: A systematic review and meta-analyses. Obes. Rev. 2020, 21, e12936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Hermoso, A.; Alonso-Martinez, A.M.; Ramírez-Vélez, R.; Izquierdo, M. Effects of exercise intervention on health-related physical fitness and blood pressure in preschool children: A systematic review and meta-analysis of randomized controlled trials. Sports Med. 2020, 50, 187–203. [Google Scholar] [CrossRef]
- Kelley, G.A.; Kelley, K.S.; Pate, R.R. Exercise and adiposity in overweight and obese children and adolescents: A systematic review with network meta-analysis of randomised trials. BMJ Open 2019, 9. [Google Scholar] [CrossRef] [Green Version]
- González-Ruiz, K.; Ramirez-Velez, R.; Correa-Bautista, J.E.; Peterson, M.D.; García-Hermoso, A. The effects of exercise on abdominal fat and liver enzymes in pediatric obesity: A systematic review and meta-analysis. Child Obes. 2017, 13, 272–282. [Google Scholar] [CrossRef]
- Dias, K.A.; Ingul, C.B.; Tjønna, A.E.; Keating, S.E.; Gomersall, S.R.; Follestad, T.; Hosseini, M.S.; Hollekim-Strand, S.M.; Ro, T.B.; Haram, M.; et al. Effect of high-intensity interval training on fitness, fat mass and cardiometabolic biomarkers in children with obesity: A randomised controlled trial. Sport Med. 2018, 48, 733–746. [Google Scholar] [CrossRef]
- Hay, J.; Wittmeier, K.; MacIntosh, A.; Wicklow, B.; Duhamel, T.; Sellers, E.; Dean, H.; Ready, E.; Berard, L.; Kriellaars, D.; et al. Physical activity intensity and type 2 diabetes risk in overweight youth: A randomized trial. Int. J. Obes. (Lond.) 2016, 40, 607–614. [Google Scholar] [CrossRef]
- Pontzer, H. Constrained total energy expenditure and the evolutionary biology of energy balance. Exerc. Sport Sci. Rev. 2015, 43, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.K.; Johnson, W.D.; Myers, C.A.; Apolzan, J.W.; Earnest, C.P.; Thomas, D.M.; Rood, J.C.; Johannsen, N.M.; Tudor-Locke, C.; Harris, M.; et al. Effect of different doses of supervised exercise on food intake, metabolism, and non-exercise physical activity: The E-MECHANIC randomized controlled trial. Am. J. Clin. Nutr. 2019, 110, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Riou, M.È.; Jomphe-Tremblay, S.; Lamothe, G.; Finlayson, G.S.; Blundell, J.E.; Décarie-Spain, L.; Gagnon, J.C.; Doucet, É. Energy compensation following a supervised exercise intervention in women living with overweight/obesity is accompanied by an early and sustained decrease in non-structured physical activity. Front. Physiol. 2019, 10, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thivel, D.; Julian, V.; Miguet, M.; Pereira, B.; Beaulieu, K.; Finlayson, G.; Richard, R.; Duclos, M. Introducing eccentric cycling during a multidisciplinary weight loss intervention might prevent adolescents with obesity from increasing their food intake: The TEXTOO study. Physiol. Behav. 2020, 214, 112744. [Google Scholar] [CrossRef] [PubMed]
- Miguet, M.; Fearnbach, N.S.; Metz, L.; Khammassi, M.; Julian, V.; Cardenoux, C.; Pereira, B.; Boirie, Y.; Duclos, M.; Thivel, D. Effect of HIIT versus MICT on body composition and energy intake in dietary restrained and unrestrained adolescents with obesity. Appl. Physiol. Nutr. Metab. 2020, 45, 437–445. [Google Scholar] [CrossRef]
- Fillon, A.; Mathieu, M.E.; Masurier, J.; Roche, J.; Miguet, M.; Khammassi, M.; Finlayson, G.; Beaulieu, K.; Pereira, B.; Duclos, M.; et al. Effect of exercise-meal timing on energy intake, appetite and food reward in adolescents with obesity: The TIMEX study. Appetite 2020, 146, 104506. [Google Scholar] [CrossRef]
- Bleich, S.N.; Vercammen, K.A.; Zatz, L.Y.; Frelier, J.M.; Ebbeling, C.B.; Peeters, A. Interventions to prevent global childhood overweight and obesity: A systematic review. Lancet Diabetes Endocrinol. 2018, 6, 332–346. [Google Scholar] [CrossRef]
- Gómez, S.F.; Esteve, R.C.; Subirana, I.; Serra-Majem, L.; Torrent, M.F.; Homs, C.; Bawaked, R.A.; Estrada, L.; Fíto, M.; Schröder, H. Effect of a community-based childhood obesity intervention program on changes in anthropometric variables, incidence of obesity, and lifestyle choices in Spanish children aged 8 to 10 years. Eur. J. Pediatr. 2018, 177, 1531–1539. [Google Scholar] [CrossRef]
- Barbour-Tuck, E.; Erlandson, M.; Muhajarine, N.; Foulds, H.; Baxter-Jones, A. Influence of childhood and adolescent fat development on fat mass accrual during emerging adulthood: A 20-year longitudinal study. Obesity 2018, 26, 613–620. [Google Scholar] [CrossRef] [Green Version]
- Knittle, J.L.; Timmers, K.; Ginsberg-Fellner, F.; Brown, R.E.; Katz, D.P. The growth of adipose tissue in children and adolescents: Cross-sectional and longitudinal studies of adipose cell number and size. J. Clin. Investig. 1979, 63, 239–246. [Google Scholar] [CrossRef] [Green Version]
- Landgraf, K.; Rockstroh, D.; Wagner, I.V.; Weise, S.; Tauscher, R.; Schwartze, J.T.; Löffler, D.; Bühligen, U.; Wojan, M.; Till, H.; et al. Evidence of early alterations in adipose tissue biology and function and its association with obesity-related inflammation and insulin resistance in children. Diabetes 2015, 64, 1249–1261. [Google Scholar] [CrossRef] [Green Version]
- Spalding, K.L.; Arner, E.; Westermark, P.O.; Bernard, S.; Buchholz, B.A.; Bergmann, O.; Blomqvist, L.; Hoffstedt, J.; Näslund, E.; Britton, T.; et al. Dynamics of fat cell turnover in humans. Nature 2008, 453, 783–787. [Google Scholar] [CrossRef] [PubMed]
- Tarabra, E.; Nouws, J.; Vash-Margita, A.; Nadzam, G.S.; Goldberg, R.; Van Name, M.; Pierpont, B.; Knight, J.R.; Shulman, G.I.; Caprio, S. The omentum of obese girls harbors small adipocytes and browning transcripts. JCI Insight 2020, 5, e135448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffery, E.; Church, C.D.; Holtrup, B.; Colman, L.; Rodeheffer, M.S. Rapid depot-specific activation of adipocyte precursor cells at the onset of obesity. Nat. Cell Biol. 2015, 17, 376–385. [Google Scholar] [CrossRef] [PubMed]
- Glastonbury, C.A.; Alves, A.C.; Moustafa, J.S.; Small, K.S. Cell-type heterogeneity in adipose tissue is associated with complex traits and reveals disease-relevant cell-specific eQTLs. Am. J. Hum. Genet. 2019, 104, 1013–1024. [Google Scholar] [CrossRef] [Green Version]
- Laforest, S.; Michaud, A.; Paris, G.; Pelletier, M.; Vidal, H.; Géloën, A.; Tchernof, A. Comparative analysis of three human adipocyte size measurement methods and their relevance for cardiometabolic risk. Obesity 2017, 25, 122–131. [Google Scholar] [CrossRef]
- Lenz, M.; Roumans, N.J.; Vink, R.G.; van Baak, M.A.; Mariman, E.C.; Arts, I.C.; de Kok, T.M.; Ertaylan, G. Estimating real cell size distribution from cross-section microscopy imaging. Bioinformatics 2016, 32, i396–i404. [Google Scholar] [CrossRef] [Green Version]
- Cifuentes-Zúñiga, F.; Arroyo-Jousse, V.; Soto-Carrasco, G.; Casanello, P.; Uauy, R.; Krause, B.J.; Castro-Rodríguez, J.A. IL-10 expression in macrophages from neonates born from obese mothers is suppressed by IL-4 and LPS/INFγ. J. Cell Physiol. 2017, 232, 3693–3701. [Google Scholar] [CrossRef]
- Kursawe, R.; Dixit, V.D.; Scherer, P.E.; Santoro, N.; Narayan, D.; Gordillo, R.; Giannini, C.; Lopez, X.; Pierpont, B.; Nouws, J.; et al. A role of the inflammasome in the low storage capacity of the abdominal subcutaneous adipose tissue in obese adolescents. Diabetes 2016, 65, 610–618. [Google Scholar] [CrossRef] [Green Version]
- Michaud, A.; Drolet, R.; Noël, S.; Paris, G.; Tchernof, A. Visceral fat accumulation is an indicator of adipose tissue macrophage infiltration in women. Metabolism 2012, 61, 689–698. [Google Scholar] [CrossRef]
- Klimcakova, E.; Roussel, B.; Kovacova, Z.; Kovacikova, M.; Siklova-Vitkova, M.; Combes, M.; Hejnova, J.; Decaunes, P.; Maoret, J.J.; Vedral, T.; et al. Macrophage gene expression is related to obesity and the metabolic syndrome in human subcutaneous fat as well as in visceral fat. Diabetologia 2011, 54, 876–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardy, O.T.; Perugini, R.A.; Nicoloro, S.M.; Gallagher-Dorval, K.; Puri, V.; Straubhaar, J.; Czech, M.P. Body mass index-independent inflammation in omental adipose tissue associated with insulin resistance in morbid obesity. Surg. Obes. Relat. Dis. 2011, 7, 60–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girón-Ulloa, A.; González-Domínguez, E.; Klimek, R.S.; Patiño-Martínez, E.; Vargas-Ayala, G.; Segovia-Gamboa, N.C.; Campos-Peña, V.; Rodríguez-Arellano, M.E.; Meraz-Ríos, M.A.; Campos-Campos, S.F.; et al. Specific macrophage subsets accumulate in human subcutaneous and omental fat depots during obesity. Immunol. Cell Biol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Christensen, R.H.; von Scholten, B.J.; Lehrskov, L.L.; Rossing, P.; Jørgensen, P.G. Epicardial adipose tissue: An emerging biomarker of cardiovascular complications in type 2 diabetes? Ther. Adv. Endocrinol. Metab. 2020, 11, 1–16. [Google Scholar] [CrossRef]
- Villasante Fricke, A.C.; Iacobellis, G. Epicardial adipose tissue: Clinical biomarker of cardio-metabolic risk. Int. J. Mol. Sci. 2019, 20, 5989. [Google Scholar] [CrossRef] [Green Version]
- Naukkarinen, J.; Heinonen, S.; Hakkarainen, A.; Lundbom, J.; Vuolteenaho, K.; Saarinen, L.; Hautaniemi, S.; Rodriguez, A.; Frühbeck, G.; Pajunen, P.; et al. Characterising metabolically healthy obesity in weight-discordant monozygotic twins. Diabetologia 2014, 57, 167–176. [Google Scholar] [CrossRef] [Green Version]
- Muniandy, M.; Heinonen, S.; Yki-Järvinen, H.; Hakkarainen, A.; Lundbom, J.; Lundbom, N.; Kaprio, J.; Rissanen, A.; Ollikainen, M.; Pietiläinen, K.H. Gene expression profile of subcutaneous adipose tissue in BMI-discordant monozygotic twin pairs unravels molecular and clinical changes associated with sub-types of obesity. Int. J. Obes. (Lond.) 2017, 41, 1176–1184. [Google Scholar] [CrossRef] [Green Version]



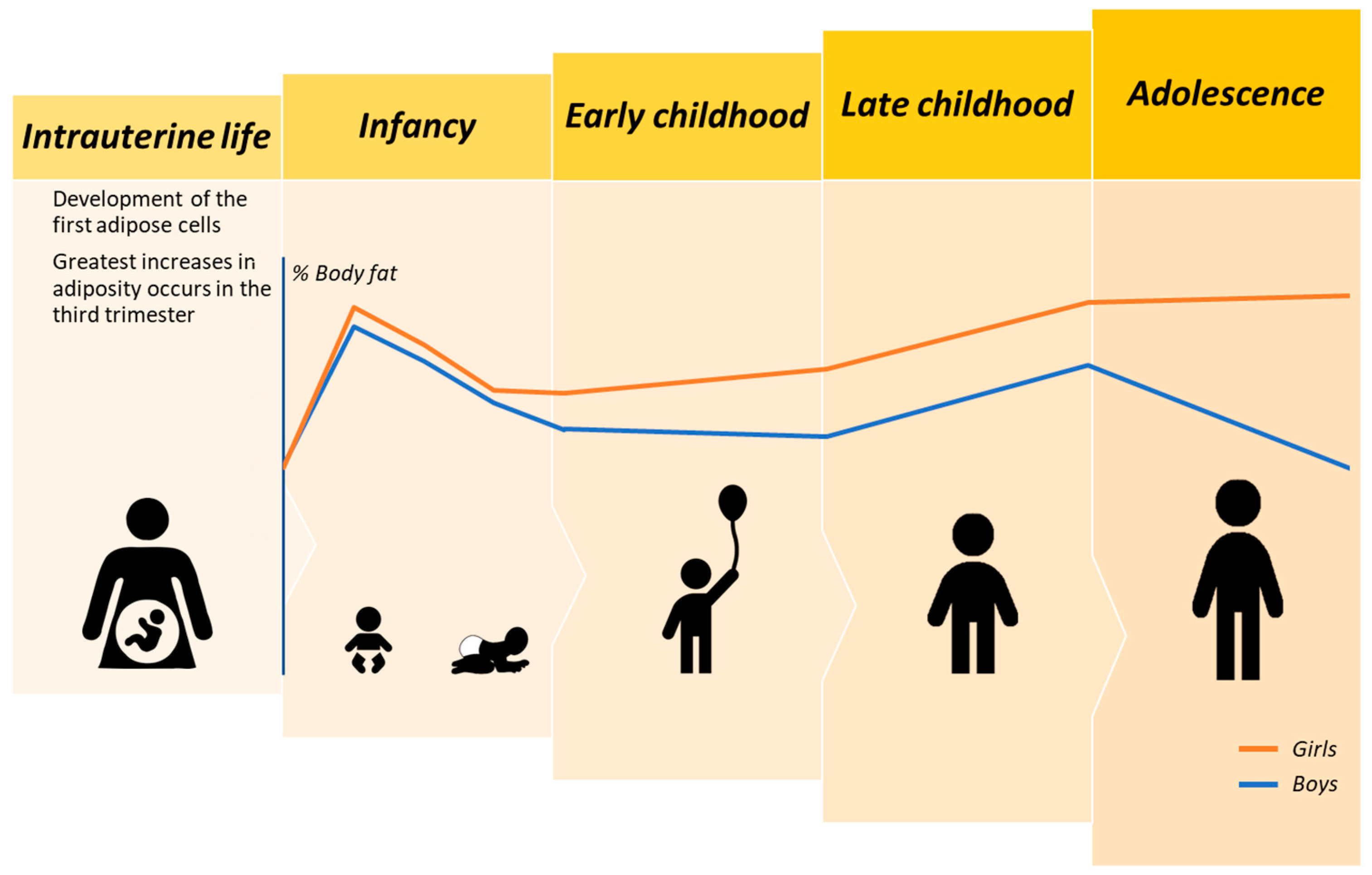{kind=link}
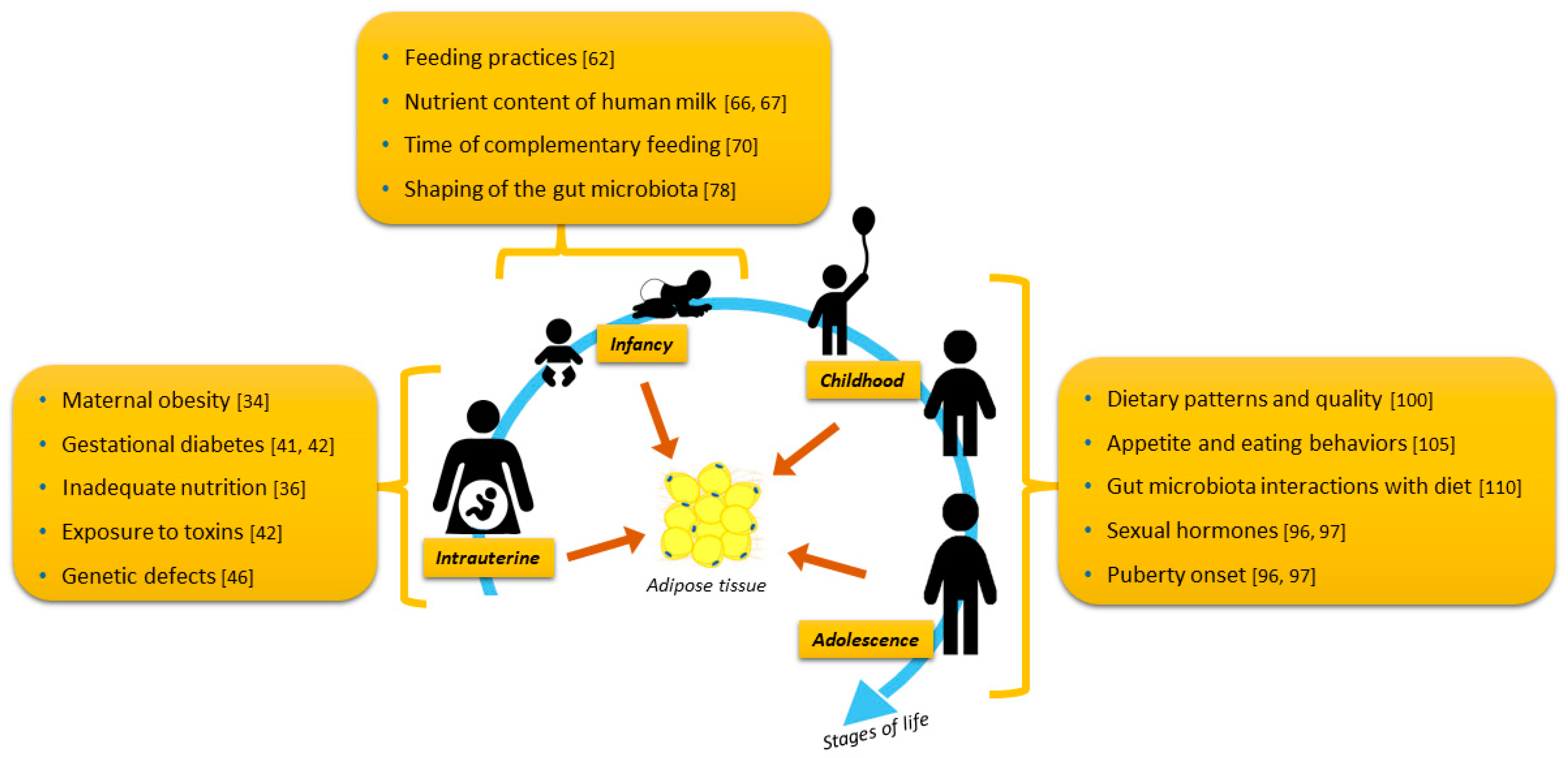{kind=link}
| White Adipose Tissue [3,19] | Brown Adipose Tissue [20] | |
|---|---|---|
| Morphology |
|
|
| Distribution |
|
|
| Primary function |
|
|
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orsso, C.E.; Colin-Ramirez, E.; Field, C.J.; Madsen, K.L.; Prado, C.M.; Haqq, A.M. Adipose Tissue Development and Expansion from the Womb to Adolescence: An Overview. Nutrients 2020, 12, 2735. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12092735
Orsso CE, Colin-Ramirez E, Field CJ, Madsen KL, Prado CM, Haqq AM. Adipose Tissue Development and Expansion from the Womb to Adolescence: An Overview. Nutrients. 2020; 12(9):2735. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12092735
Chicago/Turabian StyleOrsso, Camila E., Eloisa Colin-Ramirez, Catherine J. Field, Karen L. Madsen, Carla M. Prado, and Andrea M. Haqq. 2020. "Adipose Tissue Development and Expansion from the Womb to Adolescence: An Overview" Nutrients 12, no. 9: 2735. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12092735