
Peripancreatic Adipose Tissue Remodeling and Inflammation during High Fat Intake of Palm Oils or Lard in Rats

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Diets
2.2. Tissue Sampling and Blood Parameters
2.3. Isoprostanoid Quantification
2.4. Immunohistochemistry and Haematoxylin Staining
2.5. Statistical Analysis
3. Results
3.1. Body Weight, Glucose Homeostasis and Blood Lipids
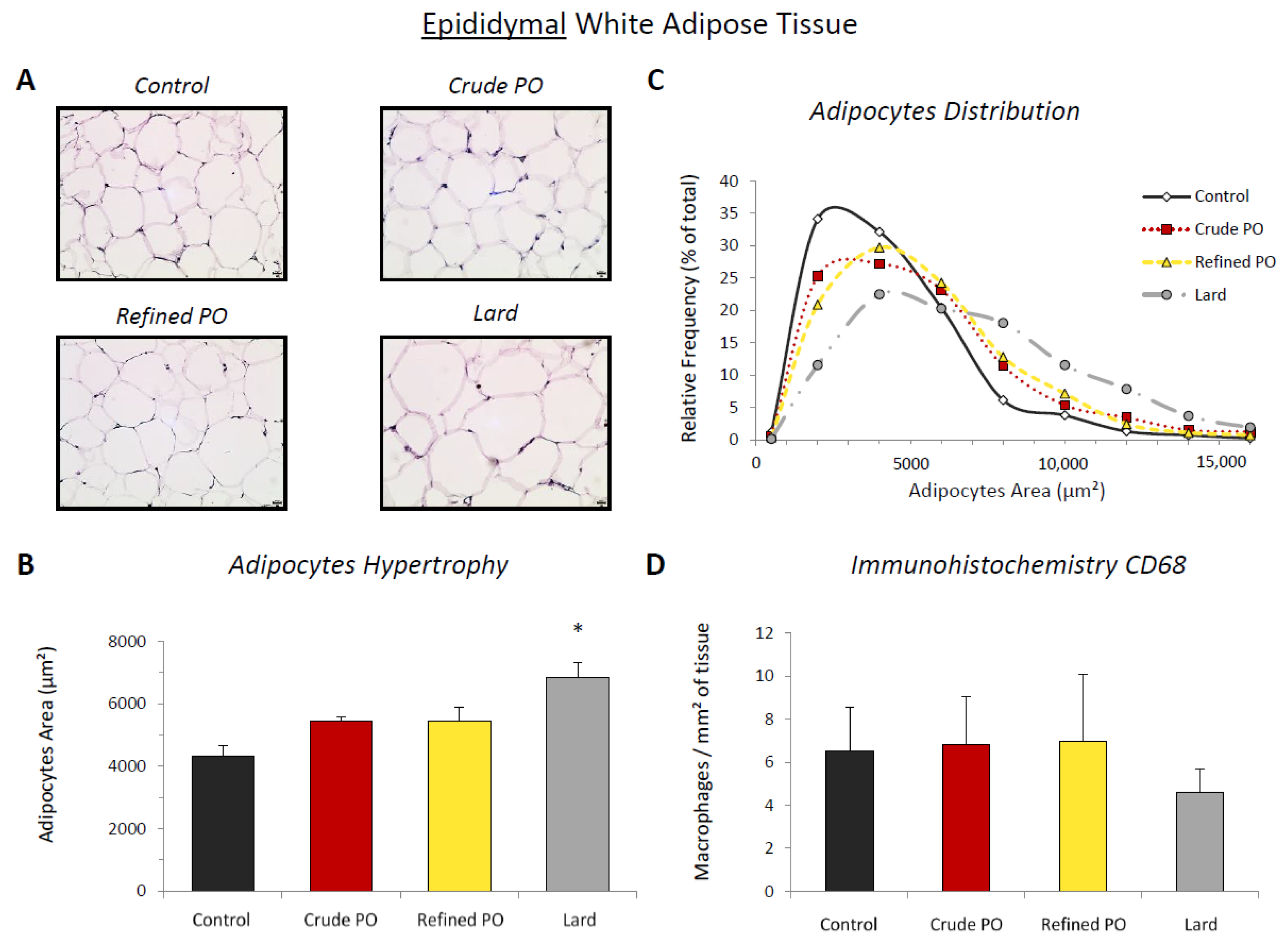
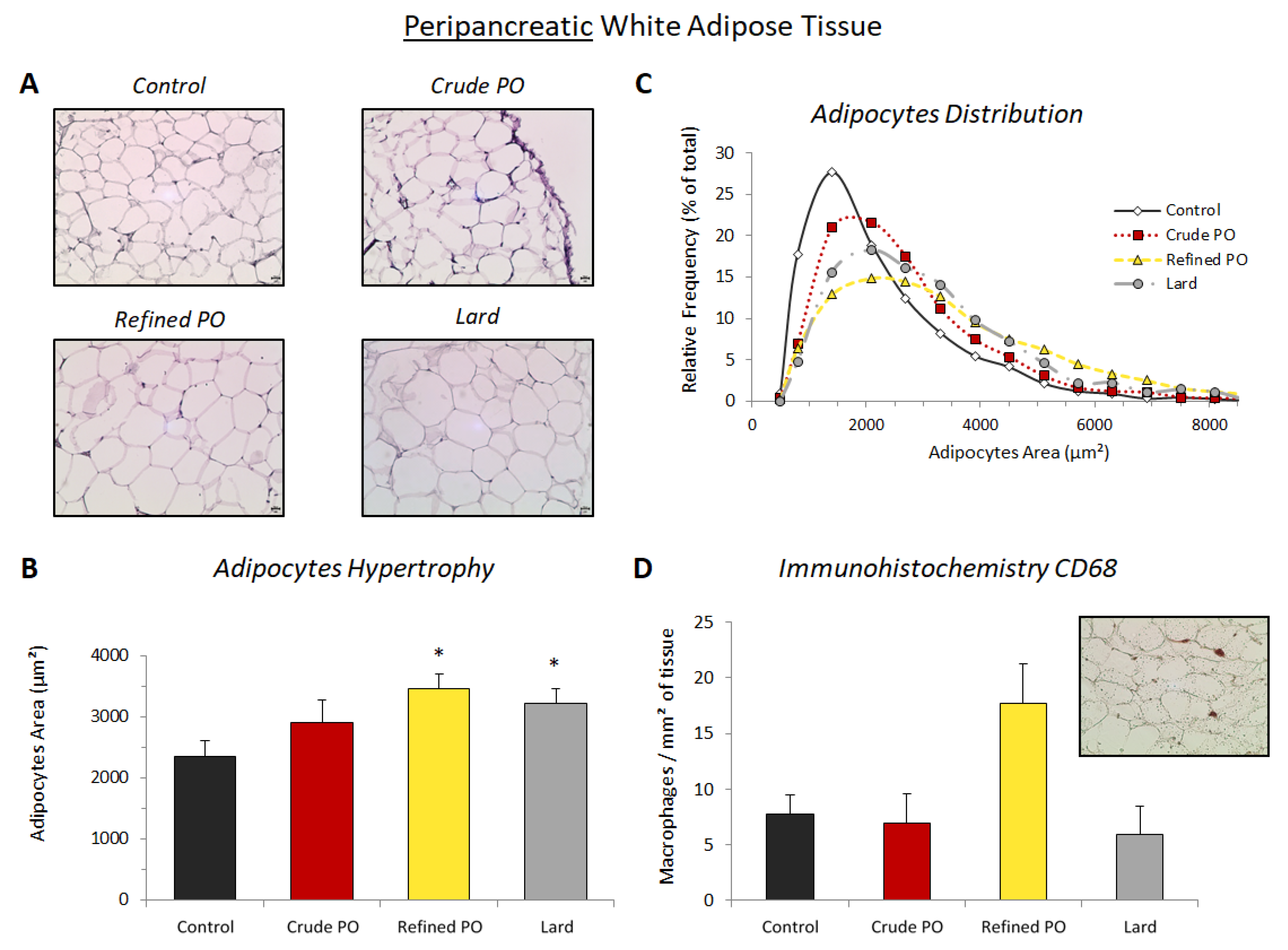
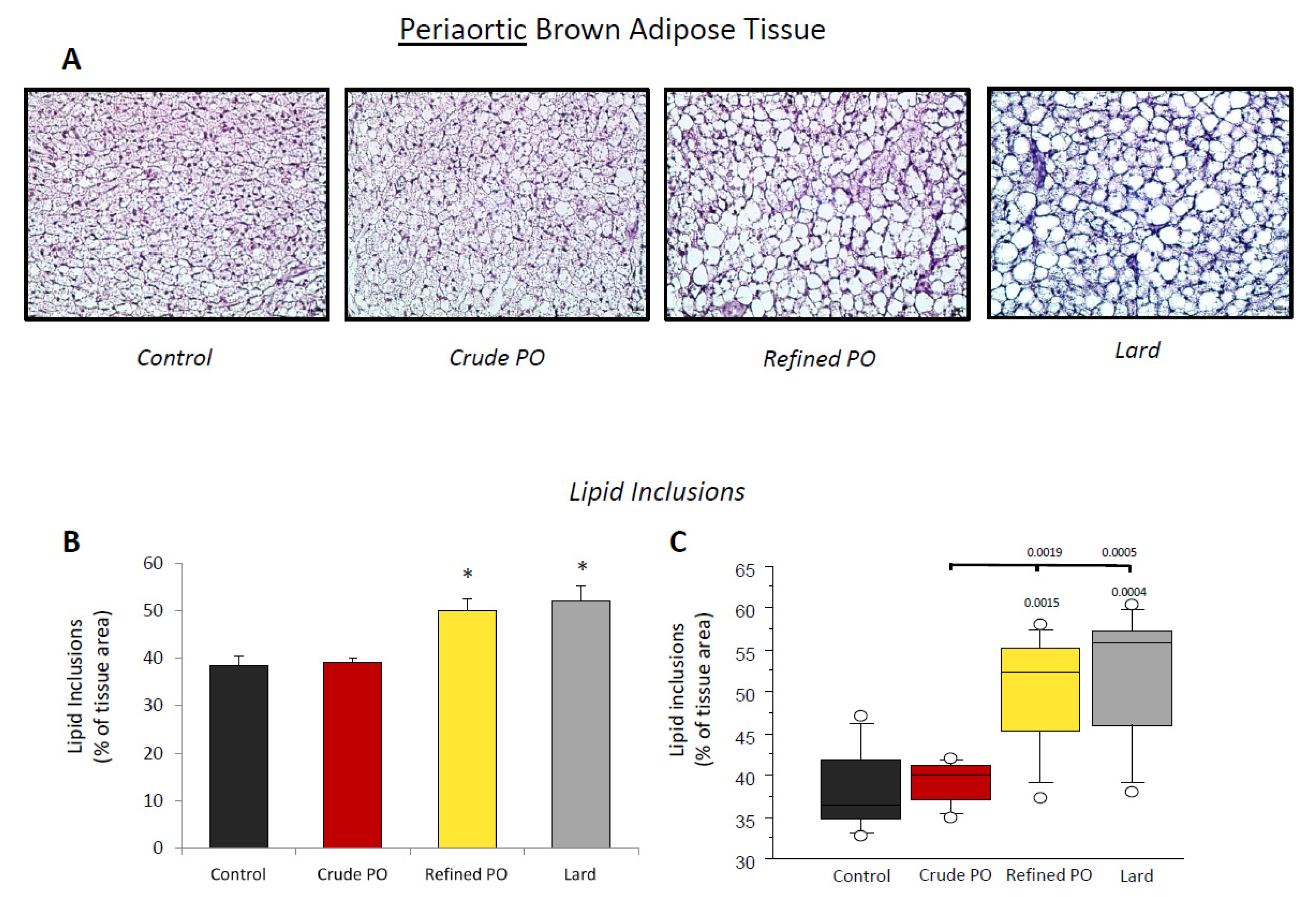
3.2. Lipid Storage in Adipose Tissues
3.3. Macrophage Recruitment in White Adipose Tissue
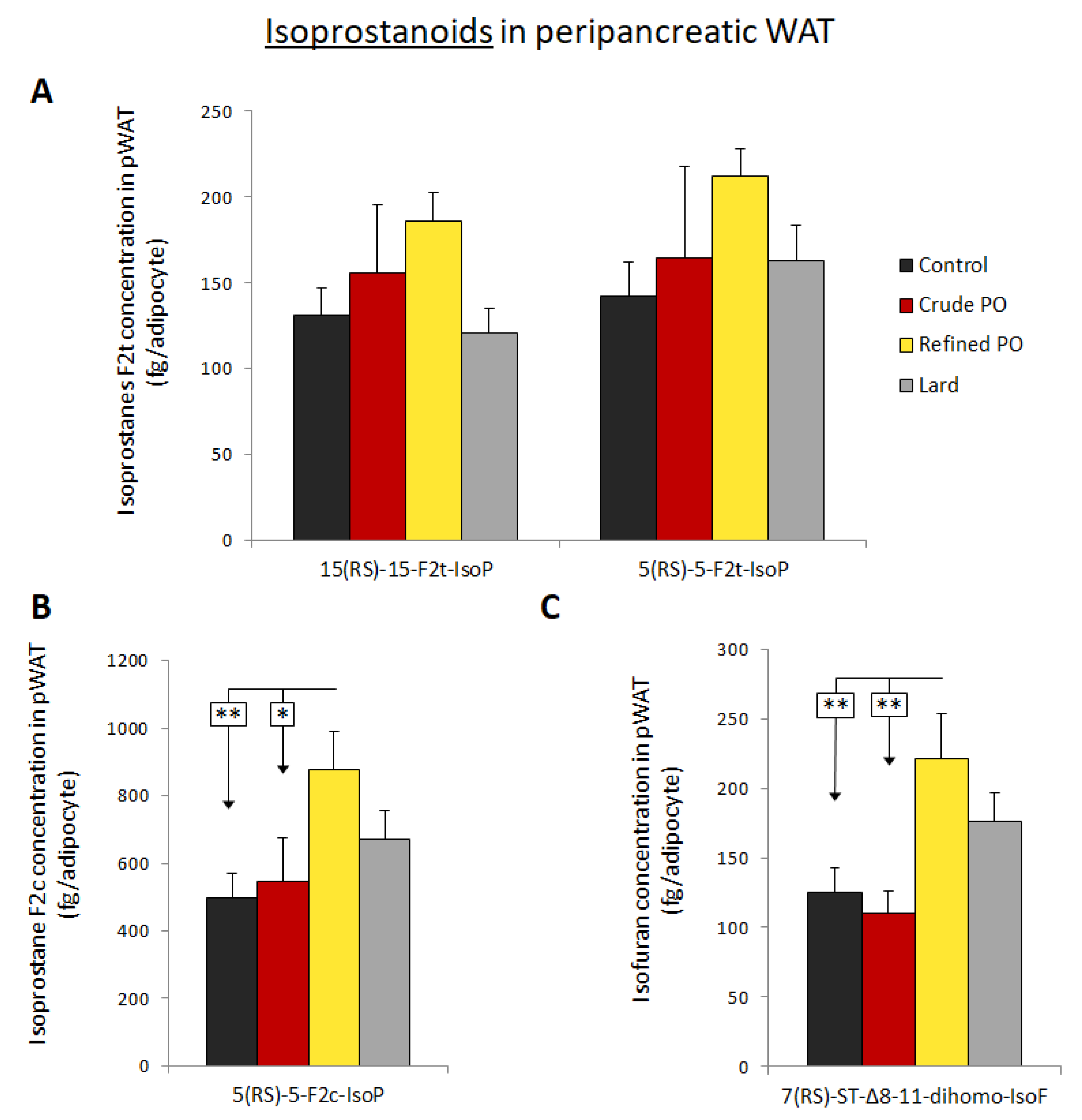
3.4. Isoprostanoid Formation in pWAT
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mba, O.I.; Dumont, M.-J.; Ngadi, M. Palm Oil: Processing, Characterization and Utilization in the Food Industry—A Review. Food Biosci. 2015, 10, 26–41. [Google Scholar] [CrossRef]
- Edem, D.O. Palm Oil: Biochemical, Physiological, Nutritional, Hematological, and Toxicological Aspects: A Review. Plant Foods Hum. Nutr. 2002, 57, 319–341. [Google Scholar] [CrossRef]
- Selvaduray, K.R.; Radhakrishnan, A.K.; Kutty, M.K.; Nesaretnam, K. Palm Tocotrienols Decrease Levels of Pro-Angiogenic Markers in Human Umbilical Vein Endothelial Cells (HUVEC) and Murine Mammary Cancer Cells. Genes Nutr. 2012, 7, 53–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odia, O.J.; Ofori, S.; Maduka, O. Palm Oil and the Heart: A Review. World J. Cardiol. 2015, 7, 144–149. [Google Scholar] [CrossRef]
- Gesteiro, E.; Guijarro, L.; Sánchez-Muniz, F.J.; Vidal-Carou, M.D.C.; Troncoso, A.; Venanci, L.; Jimeno, V.; Quilez, J.; Anadón, A.; González-Gross, M. Palm Oil on the Edge. Nutrients 2019, 11, 2008. [Google Scholar] [CrossRef] [Green Version]
- Ahsan, H.; Ahad, A.; Iqbal, J.; Siddiqui, W.A. Pharmacological Potential of Tocotrienols: A Review. Nutr. Metab. 2014, 11, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferguson, J.J.A.; Stojanovski, E.; MacDonald-Wicks, L.; Garg, M.L. Fat Type in Phytosterol Products Influence Their Cholesterol-Lowering Potential: A Systematic Review and Meta-Analysis of RCTs. Prog. Lipid Res. 2016, 64, 16–29. [Google Scholar] [CrossRef]
- Fattore, E.; Bosetti, C.; Brighenti, F.; Agostoni, C.; Fattore, G. Palm Oil and Blood Lipid-Related Markers of Cardiovascular Disease: A Systematic Review and Meta-Analysis of Dietary Intervention Trials. Am. J. Clin. Nutr. 2014, 99, 1331–1350. [Google Scholar] [CrossRef] [Green Version]
- Mancini, A.; Imperlini, E.; Nigro, E.; Montagnese, C.; Daniele, A.; Orrù, S.; Buono, P. Biological and Nutritional Properties of Palm Oil and Palmitic Acid: Effects on Health. Molecules 2015, 20, 17339–17361. [Google Scholar] [CrossRef]
- Rosen, E.D.; Spiegelman, B.M. Adipocytes as Regulators of Energy Balance and Glucose Homeostasis. Nature 2006, 444, 847–853. [Google Scholar] [CrossRef] [Green Version]
- Reilly, S.M.; Saltiel, A.R. Adapting to Obesity with Adipose Tissue Inflammation. Nat. Rev. Endocrinol. 2017, 13, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Lysaght, J.; van der Stok, E.P.; Allott, E.H.; Casey, R.; Donohoe, C.L.; Howard, J.M.; McGarrigle, S.A.; Ravi, N.; Reynolds, J.V.; Pidgeon, G.P. Pro-Inflammatory and Tumour Proliferative Properties of Excess Visceral Adipose Tissue. Cancer Lett. 2011, 312, 62–72. [Google Scholar] [CrossRef]
- Tchkonia, T.; Thomou, T.; Zhu, Y.; Karagiannides, I.; Pothoulakis, C.; Jensen, M.D.; Kirkland, J.L. Mechanisms and Metabolic Implications of Regional Differences among Fat Depots. Cell Metab. 2013, 17, 644–656. [Google Scholar] [CrossRef] [Green Version]
- Go, R.-E.; Hwang, K.-A.; Kim, Y.-S.; Kim, S.-H.; Nam, K.-H.; Choi, K.-C. Effects of Palm and Sunflower Oils on Serum Cholesterol and Fatty Liver in Rats. J. Med. Food 2015, 18, 363–369. [Google Scholar] [CrossRef]
- Djohan, Y.F.; Badia, E.; Bonafos, B.; Fouret, G.; Lauret, C.; Dupuy, A.-M.; Pinot, E.; Sutra, T.; Gaillet, S.; Lambert, K.; et al. High Dietary Intake of Palm Oils Compromises Glucose Tolerance Whereas High Dietary Intake of Olive Oil Compromises Liver Lipid Metabolism and Integrity. Eur. J. Nutr. 2019, 58, 3091–3107. [Google Scholar] [CrossRef]
- Cherian, S.; Lopaschuk, G.D.; Carvalho, E. Cellular Cross-Talk between Epicardial Adipose Tissue and Myocardium in Relation to the Pathogenesis of Cardiovascular Disease. Am. J. Physiol. Endoc. M 2012, 303, E937–E949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Dibouni, A.; Gaspar, R.; Ige, S.; Boateng, S.; Cagampang, F.R.; Gibbins, J.; Cox, R.D.; Sellayah, D. Unique Genetic and Histological Signatures of Mouse Pericardial Adipose Tissue. Nutrients 2020, 12, 1855. [Google Scholar] [CrossRef] [PubMed]
- Palau, N.; Rebuffat, S.A.; Altirriba, J.; Piquer, S.; Hanzu, F.A.; Gomis, R.; Barbera, A. Role of IGFBP-3 in the Regulation of β-Cell Mass during Obesity: Adipose Tissue/β-Cell Cross Talk. Endocrinology 2012, 153, 177–187. [Google Scholar] [CrossRef]
- Malpique, R.; Figueiredo, H.; Esteban, Y.; Rebuffat, S.A.; Hanzu, F.A.; Vinaixa, M.; Yanes, O.; Correig, X.; Barceló-Batllori, S.; Gasa, R.; et al. Integrative Analysis Reveals Novel Pathways Mediating the Interaction between Adipose Tissue and Pancreatic Islets in Obesity in Rats. Diabetologia 2014, 57, 1219–1231. [Google Scholar] [CrossRef]
- Rebuffat, S.A.; Sidot, E.; Guzman, C.; Azay-Milhau, J.; Jover, B.; Lajoix, A.-D.; Peraldi-Roux, S. Adipose Tissue Derived-Factors Impaired Pancreatic β-Cell Function in Diabetes. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 3378–3387. [Google Scholar] [CrossRef]
- Gerst, F.; Wagner, R.; Kaiser, G.; Panse, M.; Heni, M.; Machann, J.; Bongers, M.N.; Sartorius, T.; Sipos, B.; Fend, F.; et al. Metabolic Crosstalk between Fatty Pancreas and Fatty Liver: Effects on Local Inflammation and Insulin Secretion. Diabetologia 2017, 60, 2240–2251. [Google Scholar] [CrossRef]
- Laugerette, F.; Furet, J.-P.; Debard, C.; Daira, P.; Loizon, E.; Géloën, A.; Soulage, C.O.; Simonet, C.; Lefils-Lacourtablaise, J.; Bernoud-Hubac, N.; et al. Oil Composition of High-Fat Diet Affects Metabolic Inflammation Differently in Connection with Endotoxin Receptors in Mice. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E374–E386. [Google Scholar] [CrossRef] [Green Version]
- Mounien, L.; Tourniaire, F.; Landrier, J.-F. Anti-Obesity Effect of Carotenoids: Direct Impact on Adipose Tissue and Adipose Tissue-Driven Indirect Effects. Nutrients 2019, 11, 1562. [Google Scholar] [CrossRef] [Green Version]
- Harari, A.; Coster, A.C.F.; Jenkins, A.; Xu, A.; Greenfield, J.R.; Harats, D.; Shaish, A.; Samocha-Bonet, D. Obesity and Insulin Resistance Are Inversely Associated with Serum and Adipose Tissue Carotenoid Concentrations in Adults. J. Nutr. 2020, 150, 38–46. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased Oxidative Stress in Obesity and Its Impact on Metabolic Syndrome. J. Clin. Investig. 2004, 114, 1752–1761. [Google Scholar] [CrossRef]
- Murdolo, G.; Piroddi, M.; Luchetti, F.; Tortoioli, C.; Canonico, B.; Zerbinati, C.; Galli, F.; Iuliano, L. Oxidative Stress and Lipid Peroxidation By-Products at the Crossroad between Adipose Organ Dysregulation and Obesity-Linked Insulin Resistance. Biochimie 2013, 95, 585–594. [Google Scholar] [CrossRef]
- Okuno, Y.; Fukuhara, A.; Hashimoto, E.; Kobayashi, H.; Kobayashi, S.; Otsuki, M.; Shimomura, I. Oxidative Stress Inhibits Healthy Adipose Expansion Through Suppression of SREBF1-Mediated Lipogenic Pathway. Diabetes 2018, 67, 1113–1127. [Google Scholar] [CrossRef] [Green Version]
- Morrow, J.D.; Hill, K.E.; Burk, R.F.; Nammour, T.M.; Badr, K.F.; Roberts, L.J. A Series of Prostaglandin F2-like Compounds Are Produced in Vivo in Humans by a Non-Cyclooxygenase, Free Radical-Catalyzed Mechanism. Proc. Natl. Acad. Sci. USA 1990, 87, 9383–9387. [Google Scholar] [CrossRef] [Green Version]
- Galano, J.-M.; Lee, Y.Y.; Oger, C.; Vigor, C.; Vercauteren, J.; Durand, T.; Giera, M.; Lee, J.C.-Y. Isoprostanes, Neuroprostanes and Phytoprostanes: An Overview of 25 Years of Research in Chemistry and Biology. Prog. Lipid Res. 2017, 68, 83–108. [Google Scholar] [CrossRef]
- Benndorf, R.A.; Schwedhelm, E.; Gnann, A.; Taheri, R.; Kom, G.; Didié, M.; Steenpass, A.; Ergün, S.; Böger, R.H. Isoprostanes Inhibit Vascular Endothelial Growth Factor-Induced Endothelial Cell Migration, Tube Formation, and Cardiac Vessel Sprouting in Vitro, as Well as Angiogenesis in Vivo via Activation of the Thromboxane A(2) Receptor: A Potential Link between Oxidative Stress and Impaired Angiogenesis. Circ. Res. 2008, 103, 1037–1046. [Google Scholar] [CrossRef] [Green Version]
- Proudfoot, J.M.; Murrey, M.W.; McLean, S.; Greenland, E.L.; Barden, A.E.; Croft, K.D.; Galano, J.-M.; Durand, T.; Mori, T.A.; Pixley, F.J. F2-Isoprostanes Affect Macrophage Migration and CSF-1 Signalling. Free Radic. Biol. Med. 2018, 126, 142–152. [Google Scholar] [CrossRef]
- Lee, C.-Y.J.; Huang, S.H.; Jenner, A.M.; Halliwell, B. Measurement of F2-Isoprostanes, Hydroxyeicosatetraenoic Products, and Oxysterols from a Single Plasma Sample. Free Radical. Bio. Med. 2008, 44, 1314–1322. [Google Scholar] [CrossRef]
- Vigor, C.; Reversat, G.; Rocher, A.; Oger, C.; Galano, J.-M.; Vercauteren, J.; Durand, T.; Tonon, T.; Leblanc, C.; Potin, P. Isoprostanoids Quantitative Profiling of Marine Red and Brown Macroalgae. Food Chem. 2018, 268, 452–462. [Google Scholar] [CrossRef] [Green Version]
- Rotondo, F.; Romero, M.D.M.; Ho-Palma, A.C.; Remesar, X.; Fernández-López, J.A.; Alemany, M. Quantitative Analysis of Rat Adipose Tissue Cell Recovery, and Non-Fat Cell Volume, in Primary Cell Cultures. PeerJ 2016, 4, e2725. [Google Scholar] [CrossRef] [Green Version]
- Parlee, S.D.; Lentz, S.I.; Mori, H.; MacDougald, O.A. Quantifying Size and Number of Adipocytes in Adipose Tissue. Meth. Enzymol. 2014, 537, 93–122. [Google Scholar] [CrossRef] [Green Version]
- de la Torre, A.; Lee, Y.Y.; Mazzoni, A.; Guy, A.; Bultel-Poncé, V.; Durand, T.; Oger, C.; Lee, J.C.-Y.; Galano, J.-M. Total Syntheses and in Vivo Quantitation of Novel Neurofuran and Dihomo-Isofuran Derived from Docosahexaenoic Acid and Adrenic Acid. Chemistry 2015, 21, 2442–2446. [Google Scholar] [CrossRef]
- Morrow, J.D.; Awad, J.A.; Boss, H.J.; Blair, I.A.; Roberts, L.J. Non-Cyclooxygenase-Derived Prostanoids (F2-Isoprostanes) Are Formed in Situ on Phospholipids. Proc. Natl. Acad. Sci. USA 1992, 89, 10721–10725. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, I.; Aprahamian, T.; Kikuchi, R.; Shimizu, A.; Papanicolaou, K.N.; MacLauchlan, S.; Maruyama, S.; Walsh, K. Vascular Rarefaction Mediates Whitening of Brown Fat in Obesity. J. Clin. Investig. 2014, 124, 2099–2112. [Google Scholar] [CrossRef] [Green Version]
- Klöting, N.; Fasshauer, M.; Dietrich, A.; Kovacs, P.; Schön, M.R.; Kern, M.; Stumvoll, M.; Blüher, M. Insulin-Sensitive Obesity. Am. J. Physiol. Endocrinol. Metab. 2010, 299, E506–E515. [Google Scholar] [CrossRef]
- Bódis, K.; Roden, M. Energy Metabolism of White Adipose Tissue and Insulin Resistance in Humans. Eur. J. Clin. Investig. 2018, 48, e13017. [Google Scholar] [CrossRef] [Green Version]
- Chusyd, D.E.; Wang, D.; Huffman, D.M.; Nagy, T.R. Relationships between Rodent White Adipose Fat Pads and Human White Adipose Fat Depots. Front. Nutr. 2016, 3, 10. [Google Scholar] [CrossRef] [Green Version]
- Konrad, D.; Rudich, A.; Schoenle, E.J. Improved Glucose Tolerance in Mice Receiving Intraperitoneal Transplantation of Normal Fat Tissue. Diabetologia 2007, 50, 833–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssens, S.; Heemskerk, M.M.; van den Berg, S.A.; van Riel, N.A.; Nicolay, K.; van Dijk, K.W.; Prompers, J.J. Effects of Low-Stearate Palm Oil and High-Stearate Lard High-Fat Diets on Rat Liver Lipid Metabolism and Glucose Tolerance. Nutr. Metab. 2015, 12, 1–11. [Google Scholar] [CrossRef] [Green Version]
- van Dijk, S.J.; Feskens, E.J.M.; Bos, M.B.; Hoelen, D.W.M.; Heijligenberg, R.; Bromhaar, M.G.; de Groot, L.C.P.G.M.; de Vries, J.H.M.; Müller, M.; Afman, L.A. A Saturated Fatty Acid-Rich Diet Induces an Obesity-Linked Proinflammatory Gene Expression Profile in Adipose Tissue of Subjects at Risk of Metabolic Syndrome. Am. J. Clin. Nutr. 2009, 90, 1656–1664. [Google Scholar] [CrossRef] [Green Version]
- Itariu, B.K.; Zeyda, M.; Hochbrugger, E.E.; Neuhofer, A.; Prager, G.; Schindler, K.; Bohdjalian, A.; Mascher, D.; Vangala, S.; Schranz, M.; et al. Long-Chain n-3 PUFAs Reduce Adipose Tissue and Systemic Inflammation in Severely Obese Nondiabetic Patients: A Randomized Controlled Trial. Am. J. Clin. Nutr. 2012, 96, 1137–1149. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, R.K.; Brito, G.A.P.; Coelho, I.; Pequitto, D.C.T.; Yamaguchi, A.A.; Borghetti, G.; Schiessel, D.L.; Kryczyk, M.; Machado, J.; Rocha, R.E.R.; et al. Low Fish Oil Intake Improves Insulin Sensitivity, Lipid Profile and Muscle Metabolism on Insulin Resistant MSG-Obese Rats. Lipids Health Dis. 2011, 10, 66. [Google Scholar] [CrossRef] [Green Version]
- Ramaiyan, B.; Bettadahalli, S.; Talahalli, R.R. Dietary Omega-3 but Not Omega-6 Fatty Acids down-Regulate Maternal Dyslipidemia Induced Oxidative Stress: A Three Generation Study in Rats. Biochem. Biophys. Res. Commun. 2016, 477, 887–894. [Google Scholar] [CrossRef]
- Sampaio, K.A.; Ayala, J.V.; Van Hoed, V.; Monteiro, S.; Ceriani, R.; Verhé, R.; Meirelles, A.J.A. Impact of Crude Oil Quality on the Refining Conditions and Composition of Nutraceuticals in Refined Palm Oil. J. Food Sci. 2017, 82, 1842–1850. [Google Scholar] [CrossRef]
- Bonet, M.L.; Canas, J.A.; Ribot, J.; Palou, A. Carotenoids and Their Conversion Products in the Control of Adipocyte Function, Adiposity and Obesity. Arch. Biochem. Biophys. 2015, 572, 112–125. [Google Scholar] [CrossRef] [Green Version]
- Macarulla, M.T.; Alberdi, G.; Gómez, S.; Tueros, I.; Bald, C.; Rodríguez, V.M.; Martínez, J.A.; Portillo, M.P. Effects of Different Doses of Resveratrol on Body Fat and Serum Parameters in Rats Fed a Hypercaloric Diet. J. Physiol. Biochem. 2009, 65, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Poudyal, H.; Campbell, F.; Brown, L. Olive Leaf Extract Attenuates Cardiac, Hepatic, and Metabolic Changes in High Carbohydrate–, High Fat–Fed Rats. J. Nutr. 2010, 140, 946–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claro-Cala, C.M.; Quintela, J.C.; Pérez-Montero, M.; Miñano, J.; de Sotomayor, M.A.; Herrera, M.D.; Rodríguez-Rodríguez, A.R. Pomace Olive Oil Concentrated in Triterpenic Acids Restores Vascular Function, Glucose Tolerance and Obesity Progression in Mice. Nutrients 2020, 12, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jernås, M.; Palming, J.; Sjöholm, K.; Jennische, E.; Svensson, P.-A.; Gabrielsson, B.G.; Levin, M.; Sjögren, A.; Rudemo, M.; Lystig, T.C.; et al. Separation of Human Adipocytes by Size: Hypertrophic Fat Cells Display Distinct Gene Expression. FASEB J. 2006, 20, 1540–1542. [Google Scholar] [CrossRef]
- Nguyen, M.T.A.; Favelyukis, S.; Nguyen, A.-K.; Reichart, D.; Scott, P.A.; Jenn, A.; Liu-Bryan, R.; Glass, C.K.; Neels, J.G.; Olefsky, J.M. A Subpopulation of Macrophages Infiltrates Hypertrophic Adipose Tissue and Is Activated by Free Fatty Acids via Toll-like Receptors 2 and 4 and JNK-Dependent Pathways. J. Biol. Chem. 2007, 282, 35279–35292. [Google Scholar] [CrossRef] [Green Version]
- Yamada, M.; Maintinguer Norde, M.; Borges, M.C.; Mieko de Meneses Fujii, T.; Silva Jacob, P.; Fonseca-Alaniz, M.H.; Cardoso Alonso-Vale, M.I.; Torres-Leal, F.L.; Tirapegui, J.; Fock, R.A.; et al. Nutrient-Adjusted High-Fat Diet Is Associated with Absence of Periepididymal Adipose Tissue Inflammation: Is There a Link with Adequate Micronutrient Levels? Int. J. Vitam. Nutr. Res. 2013, 83, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Guo, J.; Liu, F.; Wang, M.; Li, C.; Jia, L.; Zhai, L.; Wei, W.; Bai, Y. Depot-Specific Inflammation with Decreased Expression of ATM2 in White Adipose Tissues Induced by High-Margarine/Lard Intake. PLoS ONE 2017, 12, e0188007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poret, J.M.; Souza-Smith, F.; Marcell, S.J.; Gaudet, D.A.; Tzeng, T.H.; Braymer, H.D.; Harrison-Bernard, L.M.; Primeaux, S.D. High Fat Diet Consumption Differentially Affects Adipose Tissue Inflammation and Adipocyte Size in Obesity-Prone and Obesity-Resistant Rats. Int. J. Obes. 2018, 42, 535–541. [Google Scholar] [CrossRef] [Green Version]
- Chanclón, B.; Wu, Y.; Vujičić, M.; Bauzá-Thorbrügge, M.; Banke, E.; Micallef, P.; Kanerva, J.; Wilder, B.; Rorsman, P.; Wernstedt Asterholm, I. Peripancreatic Adipose Tissue Protects against High-Fat-Diet-Induced Hepatic Steatosis and Insulin Resistance in Mice. Int. J. Obes. 2020, 44, 2323–2334. [Google Scholar] [CrossRef]
- Rebuffat, S.A.; Oliveira, J.M.; Altirriba, J.; Palau, N.; Garcia, A.; Esteban, Y.; Nadal, B.; Gomis, R. Downregulation of Sfrp5 Promotes Beta Cell Proliferation during Obesity in the Rat. Diabetologia 2013, 56, 2446–2455. [Google Scholar] [CrossRef] [Green Version]
- Brinkley, T.E.; Leng, X.; Chughtai, H.L.; Nicklas, B.J.; Kritchevsky, S.B.; Ding, J.; Kitzman, D.W.; Hundley, W.G. Periaortic Fat and Cardiovascular Risk: A Comparison of High-Risk Older Adults and Age-Matched Healthy Controls. Int. J. Obes. 2014, 38, 1397–1402. [Google Scholar] [CrossRef] [Green Version]
- Costa, R.M.; Neves, K.B.; Tostes, R.C.; Lobato, N.S. Perivascular Adipose Tissue as a Relevant Fat Depot for Cardiovascular Risk in Obesity. Front. Physiol. 2018, 9, 253. [Google Scholar] [CrossRef] [PubMed]
- Masi, L.N.; Crisma, A.R.; Martins, A.R.; do Amaral, C.L.; Torres, R.P.; Mancini Filho, J.; Hirabara, S.M.; Curi, R. Inflammatory State of Periaortic Adipose Tissue in Mice under Obesogenic Dietary Regimens. J. Nutr. Intermed. Metab. 2016, 6, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Oi-Kano, Y.; Iwasaki, Y.; Nakamura, T.; Watanabe, T.; Goto, T.; Kawada, T.; Watanabe, K.; Iwai, K. Oleuropein Aglycone Enhances UCP1 Expression in Brown Adipose Tissue in High-Fat-Diet-Induced Obese Rats by Activating β-Adrenergic Signaling. J. Nutr. Biochem. 2017, 40, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Fessel, J.P.; Porter, N.A.; Moore, K.P.; Sheller, J.R.; Roberts, L.J. Discovery of Lipid Peroxidation Products Formed in Vivo with a Substituted Tetrahydrofuran Ring (Isofurans) That Are Favored by Increased Oxygen Tension. Proc. Natl. Acad. Sci. USA 2002, 99, 16713–16718. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Chen, X. Reducing Selenoprotein P Expression Suppresses Adipocyte Differentiation as a Result of Increased Preadipocyte Inflammation. Am. J. Physiol. Endocrinol. Metab. 2011, 300, E77–E85. [Google Scholar] [CrossRef] [Green Version]
- Marseglia, L.; Manti, S.; D’Angelo, G.; Nicotera, A.; Parisi, E.; Di Rosa, G.; Gitto, E.; Arrigo, T. Oxidative Stress in Obesity: A Critical Component in Human Diseases. Int. J. Mol. Sci. 2014, 16, 378–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chattopadhyay, M.; Khemka, V.K.; Chatterjee, G.; Ganguly, A.; Mukhopadhyay, S.; Chakrabarti, S. Enhanced ROS Production and Oxidative Damage in Subcutaneous White Adipose Tissue Mitochondria in Obese and Type 2 Diabetes Subjects. Mol. Cell Biochem. 2015, 399, 95–103. [Google Scholar] [CrossRef]
- Johnson, A.R.; Wilkerson, M.D.; Sampey, B.P.; Troester, M.A.; Hayes, D.N.; Makowski, L. Cafeteria Diet-Induced Obesity Causes Oxidative Damage in White Adipose. Biochem. Biophys. Res. Commun. 2016, 473, 545–550. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Berg, A.H.; Iyengar, P.; Lam, T.K.T.; Giacca, A.; Combs, T.P.; Rajala, M.W.; Du, X.; Rollman, B.; Li, W.; et al. The Hyperglycemia-Induced Inflammatory Response in Adipocytes: The Role of Reactive Oxygen Species. J. Biol. Chem. 2005, 280, 4617–4626. [Google Scholar] [CrossRef] [Green Version]
- Laget, J. Biocommunication between Visceral Adipose Tissue and Pancreatic Beta-Cells: Isoprostanes and MiRNAs. Ph.D. Thesis, Université de Montpellier, Montpellier, France, 2019. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Crude PO | Refined PO | Lard | p-Value | |
|---|---|---|---|---|---|
| Initial body weight (g) | 209 ± 4 | 203 ± 1 | 204 ± 2 | 208 ± 3 | NS |
| Final body weight (g) | 527 ± 11 | 609 ± 14 ** | 612 ± 20 ** | 611 ± 23 ** | 0.0065 |
| Body weight gain (g) | 318 ± 9 | 406 ± 14 ** | 408 ± 19 ** | 403 ± 22 ** | 0.0022 |
| eWAT weight (g) | 12.6 ± 1.8 | 20.1 ± 2.1 | 16.6 ± 3.5 | 21.4 ± 2.9 | NS |
| Fasting glucose (mg/dL) | 136.0 ± 3.5 | 155.0 ± 5.8 * | 157.1 ± 3.7 ** | 131.6 ± 5.7 | 0.0015 |
| Fasting insulin (mU/L) | 53.5 ± 7.6 | 108.6 ± 22.9 | 147.0 ± 29.9 | 93.6 ± 23.6 | NS |
| HOMA-IR | 18.1 ± 2.8 | 41.7 ± 9.1 | 57.6 ± 12.0 ** | 31.6 ± 8.7 | 0.0344 |
| Blood TG (mM) | 1.03 ± 0.15 | 0.99 ± 0.11 | 1.33 ± 0.21 | 1.01 ± 0.15 | NS |
| Total cholesterol (mM) | 1.97 ± 0.16 | 1.75 ± 0.15 | 2.00 ± 0.14 | 1.55 ± 0.09 | NS |
| HDL cholesterol (mM) | 1.48 ± 0.12 | 1.34 ± 0.85 | 1.56 ± 0.12 | 1.06 ± 0.12 * | 0.0179 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laget, J.; Djohan, Y.F.; Jeanson, L.; Muyor, K.; Badia, E.; Cristol, J.P.; Coudray, C.; Feillet-Coudray, C.; Vigor, C.; Oger, C.; et al. Peripancreatic Adipose Tissue Remodeling and Inflammation during High Fat Intake of Palm Oils or Lard in Rats. Nutrients 2021, 13, 1134. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13041134
Laget J, Djohan YF, Jeanson L, Muyor K, Badia E, Cristol JP, Coudray C, Feillet-Coudray C, Vigor C, Oger C, et al. Peripancreatic Adipose Tissue Remodeling and Inflammation during High Fat Intake of Palm Oils or Lard in Rats. Nutrients. 2021; 13(4):1134. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13041134
Chicago/Turabian StyleLaget, Jonas, Youzan Ferdinand Djohan, Laura Jeanson, Karen Muyor, Eric Badia, Jean Paul Cristol, Charles Coudray, Christine Feillet-Coudray, Claire Vigor, Camille Oger, and et al. 2021. "Peripancreatic Adipose Tissue Remodeling and Inflammation during High Fat Intake of Palm Oils or Lard in Rats" Nutrients 13, no. 4: 1134. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13041134