
Adiponectin: Structure, Physiological Functions, Role in Diseases, and Effects of Nutrition

 , ,
, ,
Abstract
:1. Introduction
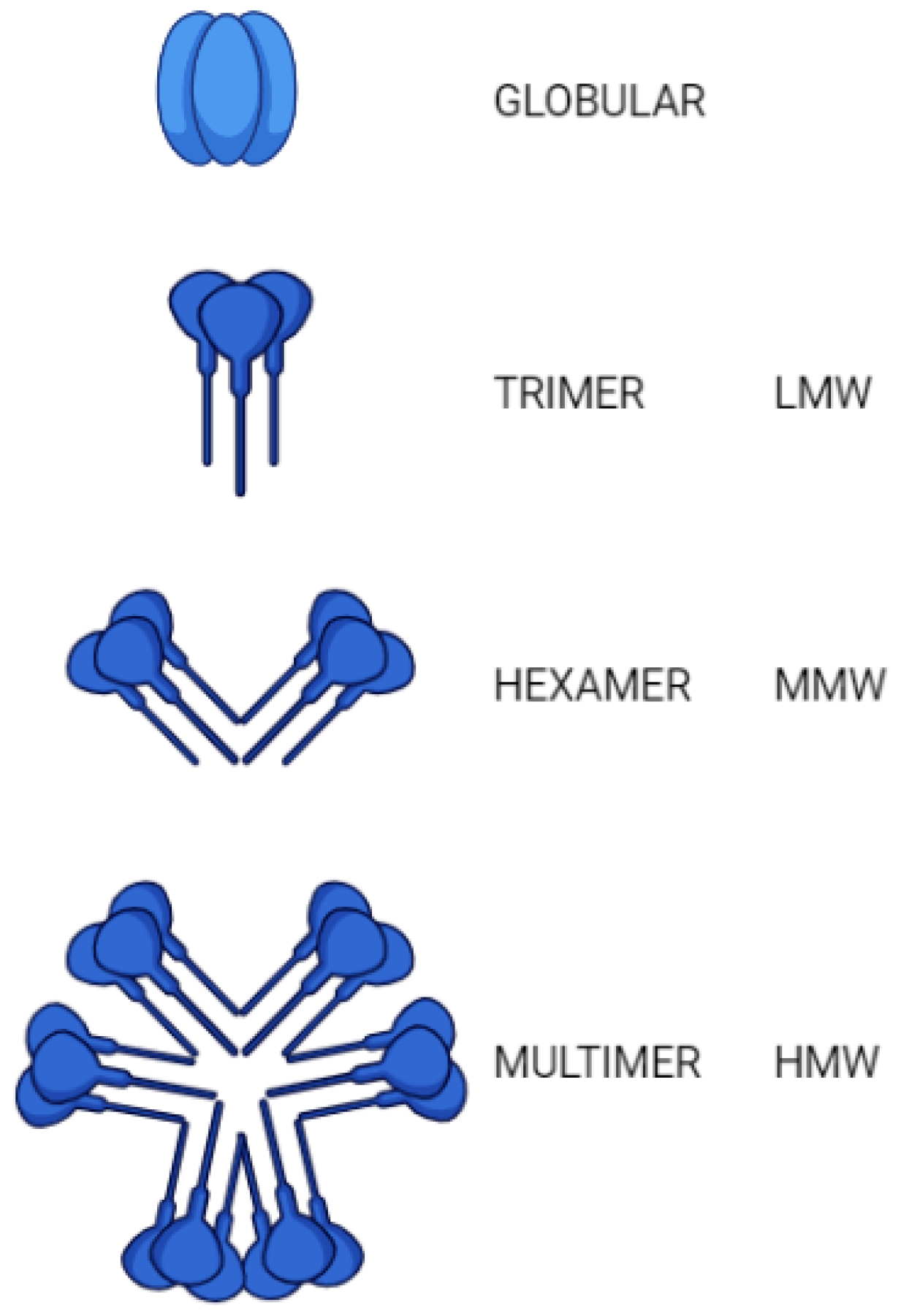
2. Structure
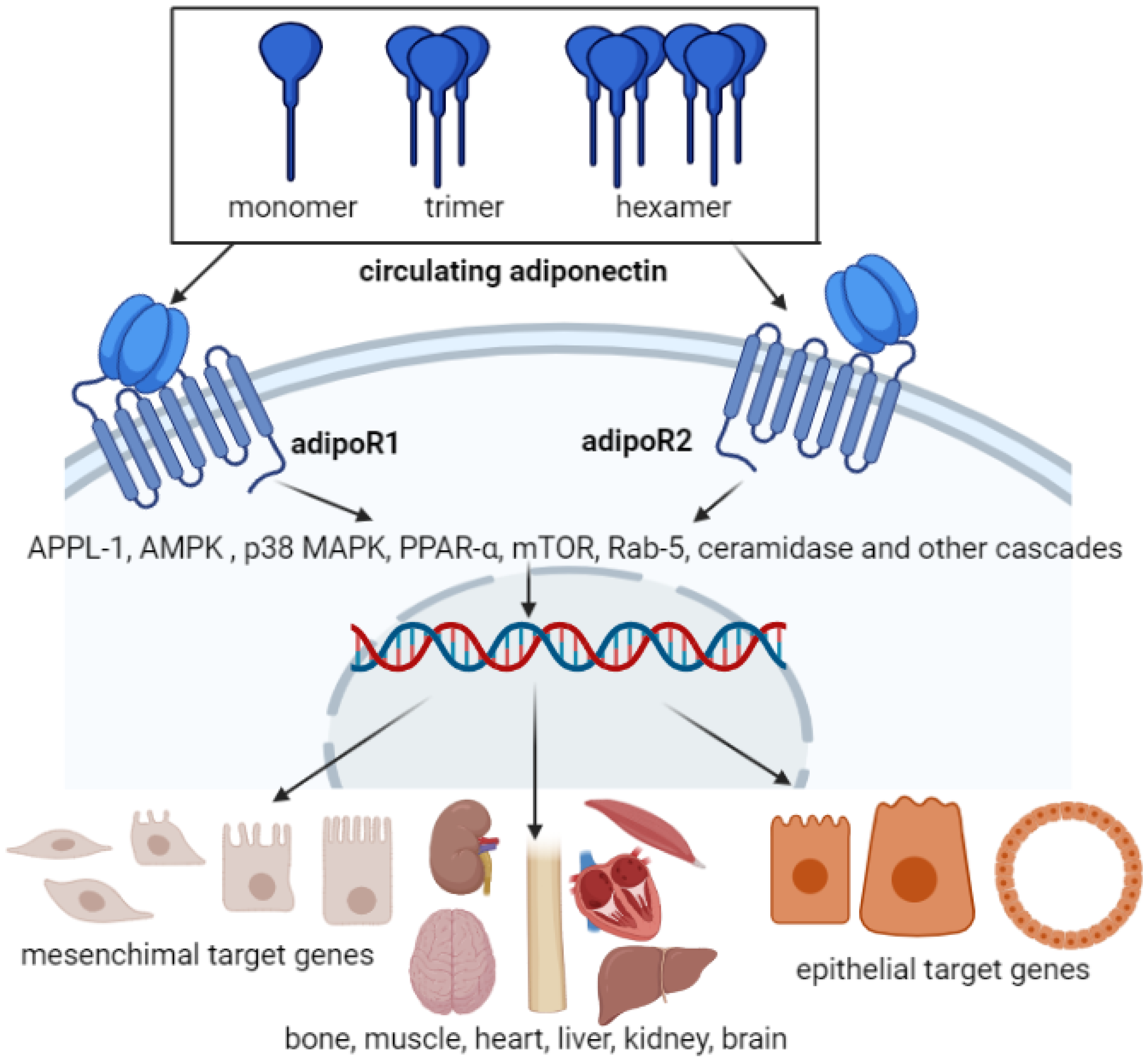
3. Adiponectin Receptors
4. Adiponectin Functions in Different Body Organs
4.1. Adiponectin Functions in the Central Nervous System
4.2. Adiponectin Functions in the Liver
4.3. Adiponectin Functions in the Muscle
4.4. Adiponectin Functions in the Heart
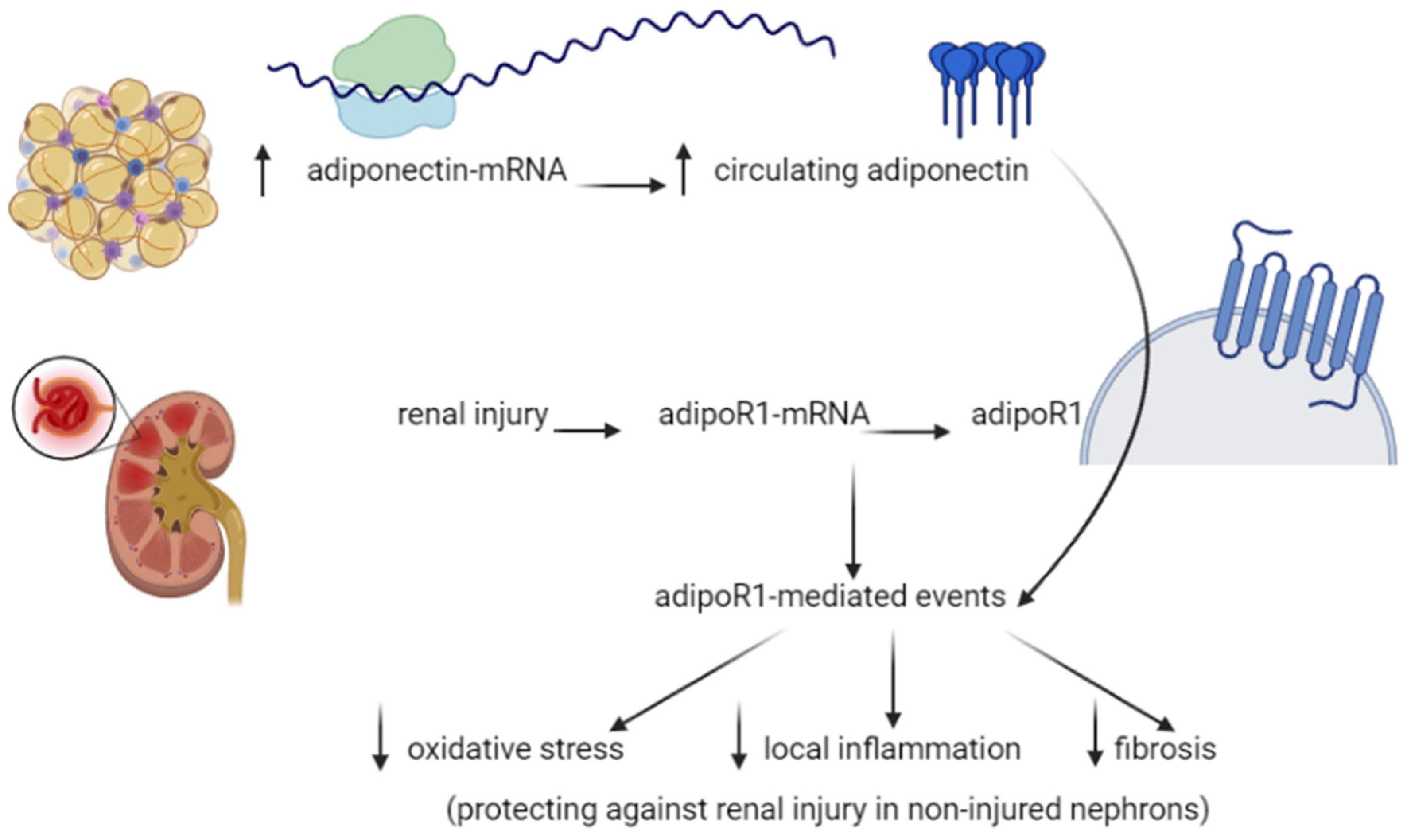
4.5. Adiponectin Functions in the Kidney
4.6. Adiponectin Function in the Bone
5. Adiponectin and Diseases
5.1. Adiponectin and Diabetes
5.2. Adiponectin and Cancer
5.3. Adiponectin and Cardiovascular Diseases
5.4. Adiponectin and Alzheimer’s Disease
5.5. Diet and Supplements
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kim, D.H.; Vanella, L.; Inoue, K.; Burgess, A.; Gotlinger, K.; Manthati, V.L.; Koduru, S.R.; Zeldin, D.C.; Falck, J.R.; Schwartzman, M.L.; et al. Epoxyeicosatrienoic acid agonist regulates human mesenchymal stem cell-derived adipocytes through activation of HO-1-pAKT signaling and a decrease in PPARgamma. Stem Cells Dev. 2010, 19, 1863–1873. [Google Scholar] [CrossRef]
- Zhao, L.; Fu, Z.; Liu, Z. Adiponectin and insulin cross talk: The microvascular connection. Trends Cardiovasc. Med. 2014, 24, 319–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takenouchi, Y.; Kobayashi, T.; Matsumoto, T.; Kamata, K.J.A. Gender differences in age-related endothelial function in the murine aorta. Atherosclerosis 2009, 206, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Schondorf, T.; Maiworm, A.; Emmison, N.; Forst, T.; Pfutzner, A. Biological background and role of adiponectin as marker for insulin resistance and cardiovascular risk. Clin. Lab. 2005, 51, 489–494. [Google Scholar]
- Achari, A.E.; Jain, S.K. Adiponectin, a Therapeutic Target for Obesity, Diabetes, and Endothelial Dysfunction. Int. J. Mol. Sci. 2017, 18, 1321. [Google Scholar] [CrossRef] [Green Version]
- Lau, W.B.; Tao, L.; Wang, Y.; Li, R.; Ma, X.L. Systemic adiponectin malfunction as a risk factor for cardiovascular disease. Antioxid. Redox Signal. 2011, 15, 1863–1873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, Y.; Nakanishi, K.; Tachibana, I.; Kumanogoh, A. Adiponectin: A novel link between adipocytes and COPD. Vitam. Horm. 2012, 90, 419–435. [Google Scholar] [PubMed]
- Fruebis, J.; Tsao, T.S.; Javorschi, S.; Ebbets-Reed, D.; Erickson, M.R.; Yen, F.T.; Bihain, B.E.; Lodish, H.F. Proteolytic cleavage product of 30-kDa adipocyte complement-related protein increases fatty acid oxidation in muscle and causes weight loss in mice. Proc. Natl. Acad. Sci. USA 2001, 98, 2005–2010. [Google Scholar] [CrossRef] [PubMed]
- Hara, K.; Horikoshi, M.; Yamauchi, T.; Yago, H.; Miyazaki, O.; Ebinuma, H.; Imai, Y.; Nagai, R.; Kadowaki, T.; et al. Measurement of the high-molecular weight form of adiponectin in plasma is useful for the prediction of insulin resistance and metabolic syndrome. Diabetes Care 2006, 29, 1357–1362. [Google Scholar] [CrossRef] [Green Version]
- Martin, L.J.; Woo, J.G.; Daniels, S.R.; Goodman, E.; Dolan, L.M. The relationships of adiponectin with insulin and lipids are strengthened with increasing adiposity. J. Clin. Endocrinol. Metab. 2005, 90, 4255–4259. [Google Scholar] [CrossRef] [Green Version]
- Kelly, M.; Ruderman, N.B.; Tomas, E. AMP-activated protein kinase and its regulation by adiponectin and interleukin-6. Scand. J. Food Nutr. 2006, (Suppl. 2), 85–91. [Google Scholar] [CrossRef]
- Kadowaki, T.; Yamauchi, T.; Kubota, N.; Hara, K.; Ueki, K.; Tobe, K. Adiponectin and adiponectin receptors in insulin resistance, diabetes, and the metabolic syndrome. J. Clin. Investig. 2006, 116, 1784–1792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satoh, H.; Nguyen, M.T.; Trujillo, M.; Imamura, T.; Usui, I.; Scherer, P.E.; Olefsky, J.M. Adenovirus-mediated adiponectin expression augments skeletal muscle insulin sensitivity in male Wistar rats. Diabetes 2005, 54, 1304–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, F.M.; Trujillo, M.E.; Hanif, W.; Barnett, A.H.; McTernan, P.G.; Scherer, P.E.; Kumar, S. Serum high molecular weight complex of adiponectin correlates better with glucose tolerance than total serum adiponectin in Indo-Asian males. Diabetologia 2005, 48, 1084–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.; Song, X.; Briggs, M.; Violand, B.; Salsgiver, W.; Gulve, E.A.; Luo, Y. Adiponectin represses gluconeogenesis independent of insulin in hepatocytes. Biochem. Biophys. Res. Commun. 2005, 338, 793–799. [Google Scholar] [CrossRef] [PubMed]
- Bobbert, T.; Rochlitz, H.; Wegewitz, U.; Akpulat, S.; Mai, K.; Weickert, M.O.; Möhlig, M.; Pfeiffer, A.F.; Spranger, J. Changes of adiponectin oligomer composition by moderate weight reduction. Diabetes 2005, 54, 2712–2719. [Google Scholar] [CrossRef] [Green Version]
- Watson, R.R.; Zibadi, S.; Preedy, V.R. Dietary Components and Immune Function; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Vionnet, N.; Hani, E.H.; Dupont, S.; Gallina, S.; Francke, S.; Dotte, S.; De Matos, F.; Durand, E.; Leprêtre, F.; Lecoeur, C.; et al. Genomewide search for type 2 diabetes-susceptibility genes in French whites: Evidence for a novel susceptibility locus for early-onset diabetes on chromosome 3q27-qter and independent replication of a type 2-diabetes locus on chromosome 1q21-q24. Am. J. Hum. Genet. 2000, 67, 1470–1480. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, L.; Scherer, P.E. The crystal structure of a complement-1q family protein suggests an evolutionary link to tumor necrosis factor. Curr. Biol. 1998, 8, 335–338. [Google Scholar] [CrossRef] [Green Version]
- de Luis, D.A.; Izaola, O.; Primo, D.; Gomez-Hoyos, E.; Ortola, A.; Lopez-Gomez, J.J.; Aller, R. Role of rs1501299 variant in the adiponectin gene on total adiponectin levels, insulin resistance and weight loss after a Mediterranean hypocaloric diet. Diabetes Res. Clin. Pract. 2019, 148, 262–267. [Google Scholar] [CrossRef]
- Yamauchi, T.; Iwabu, M.; Okada-Iwabu, M.; Kadowaki, T. Adiponectin receptors: A review of their structure, function and how they work. Best Pract. Res. Clin. Endocrinol. Metab. 2014, 28, 15–23. [Google Scholar] [CrossRef]
- Yu, D.; Yu, Z.; Sun, Q.; Sun, L.; Li, H.; Song, J.; Mi, M.; Wu, H.; Lu, L.; Liu, C.; et al. Effects of body fat on the associations of high-molecular-weight adiponectin, leptin and soluble leptin receptor with metabolic syndrome in Chinese. PLoS ONE 2011, 6, e16818. [Google Scholar] [CrossRef]
- Peng, Y.J.; Shen, T.L.; Chen, Y.S.; Mersmann, H.J.; Liu, B.H.; Ding, S.T. Adiponectin and adiponectin receptor 1 overexpression enhance inflammatory bowel disease. J. Biomed. Sci. 2018, 25, 24. [Google Scholar] [CrossRef]
- Wang, Y.; Ma, X.L.; Lau, W.B. Cardiovascular Adiponectin Resistance: The Critical Role of Adiponectin Receptor Modification. Trends Endocrinol. Metab. TEM 2017, 28, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Fu, C.; Xiao, J.; Ye, Z. Uric acid upregulates the adiponectin receptor 1 pathway in renal proximal tubule epithelial cells. Mol. Med. Rep. 2018, 17, 3545–3554. [Google Scholar] [PubMed]
- Vansaun, M.N. Molecular pathways: Adiponectin and leptin signaling in cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2013, 19, 1926–1932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hug, C.; Wang, J.; Ahmad, N.S.; Bogan, J.S.; Tsao, T.S.; Lodish, H.F. T-cadherin is a receptor for hexameric and high-molecular-weight forms of Acrp30/adiponectin. Proc. Natl. Acad. Sci. USA 2004, 101, 10308–10313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker-Duffen, J.L.; Nakamura, K.; Silver, M.; Zuriaga, M.A.; MacLauchlan, S.; Aprahamian, T.R.; Walsh, K. Divergent roles for adiponectin receptor 1 (AdipoR1) and AdipoR2 in mediating revascularization and metabolic dysfunction in vivo. J. Biol. Chem. 2014, 289, 16200–16213. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Li, H.; Tai, L.W.; Gu, P.; Cheung, C.W. Adiponectin regulates thermal nociception in a mouse model of neuropathic pain. Br. J. Anaesth. 2018, 120, 1356–1367. [Google Scholar] [CrossRef] [Green Version]
- Yau, S.Y.; Li, A.; Hoo, R.L.; Ching, Y.P.; Christie, B.R.; Lee, T.M.; Xu, A.; So, K.-F. Physical exercise-induced hippocampal neurogenesis and antidepressant effects are mediated by the adipocyte hormone adiponectin. Proc. Natl. Acad. Sci. USA 2014, 111, 15810–15815. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Wang, X.; Lu, X.Y. Adiponectin Exerts Neurotrophic Effects on Dendritic Arborization, Spinogenesis, and Neurogenesis of the Dentate Gyrus of Male Mice. Endocrinology 2016, 157, 2853–2869. [Google Scholar] [CrossRef] [Green Version]
- Malberg, J.E. Implications of adult hippocampal neurogenesis in antidepressant action. J. Psychiatry Neurosci. JPN 2004, 29, 196–205. [Google Scholar] [PubMed]
- Park, S.; Kim, D.S.; Kwon, D.Y.; Yang, H.J. Long-term central infusion of adiponectin improves energy and glucose homeostasis by decreasing fat storage and suppressing hepatic gluconeogenesis without changing food intake. J. Neuroendocrinol. 2011, 23, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Mandal, P.; Pritchard, M.T.; Nagy, L.E. Anti-inflammatory pathways and alcoholic liver disease: Role of an adiponectin/interleukin-10/heme oxygenase-1 pathway. World J. Gastroenterol. 2010, 16, 1330–1336. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Deepa, S.S.; Etzler, J.C.; Ryu, J.; Mao, X.; Fang, Q.; Liu, D.D.; Torres, J.M.; Jia, W.; Lechleiter, J.D.; et al. Adiponectin activates AMP-activated protein kinase in muscle cells via APPL1/LKB1-dependent and phospholipase C/Ca2+/Ca2+/calmodulin-dependent protein kinase kinase-dependent pathways. J. Biol. Chem. 2009, 284, 22426–22435. [Google Scholar] [CrossRef] [Green Version]
- Tilg, H.; Moschen, A.R. Evolution of inflammation in nonalcoholic fatty liver disease: The multiple parallel hits hypothesis. Hepatology 2010, 52, 1836–1846. [Google Scholar] [CrossRef]
- Combs, T.P.; Marliss, E.B. Adiponectin signaling in the liver. Rev. Endocr. Metab. Disord. 2014, 15, 137–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elissa, L.A.; Elsherbiny, N.M.; Magmomah, A.O. Propolis restored adiponectin level in type 2 diabetes through PPARγ activation. Egypt. J. Basic Appl. Sci. 2015, 2, 318–326. [Google Scholar] [CrossRef] [Green Version]
- Aleidi, S.; Issa, A.; Bustanji, H.; Khalil, M.; Bustanji, Y. Adiponectin serum levels correlate with insulin resistance in type 2 diabetic patients. Saudi Pharm. J. SPJ Off. Publ. Saudi Pharm. Soc. 2015, 23, 250–256. [Google Scholar] [CrossRef] [Green Version]
- Yamauchi, T.; Kamon, J.; Minokoshi, Y.; Ito, Y.; Waki, H.; Uchida, S.; Yamashita, S.; Noda, M.; Kita, S.; Ueki, K.; et al. Adiponectin stimulates glucose utilization and fatty-acid oxidation by activating AMP-activated protein kinase. Nat. Med. 2002, 8, 1288–1295. [Google Scholar] [CrossRef]
- Lee, S.; Kwak, H.B. Role of adiponectin in metabolic and cardiovascular disease. J. Exerc. Rehabil. 2014, 10, 54–59. [Google Scholar] [CrossRef] [Green Version]
- Fang, X.; Palanivel, R.; Cresser, J.; Schram, K.; Ganguly, R.; Thong, F.S.; Tuinei, J.; Xu, A.; Abel, E.D.; Sweeney, G. An APPL1-AMPK signaling axis mediates beneficial metabolic effects of adiponectin in the heart. Am. J. Physiol. Endocrinol. Metab. 2010, 299, E721–E729. [Google Scholar] [CrossRef] [Green Version]
- Sharma, K.; Ramachandrarao, S.; Qiu, G.; Usui, H.K.; Zhu, Y.; Dunn, S.R.; Ouedraogo, R.; Hough, K.; McCue, P.; Chan, L.; et al. Adiponectin regulates albuminuria and podocyte function in mice. J. Clin. Investig. 2008, 118, 1645–1656. [Google Scholar] [CrossRef] [PubMed]
- Christou, G.A.; Kiortsis, D.N. The role of adiponectin in renal physiology and development of albuminuria. J. Endocrinol. 2014, 221, R49–R61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsioufis, C.; Dimitriadis, K.; Chatzis, D.; Vasiliadou, C.; Tousoulis, D.; Papademetriou, V.; Toutouzas, P.; Stefanadis, C.; Kallikazaros, I. Relation of microalbuminuria to adiponectin and augmented C-reactive protein levels in men with essential hypertension. Am. J. Cardiol. 2005, 96, 946–951. [Google Scholar] [CrossRef] [PubMed]
- Naot, D.; Musson, D.S.; Cornish, J. The Activity of Adiponectin in Bone. Calcif. Tissue Int. 2017, 100, 486–499. [Google Scholar] [CrossRef] [PubMed]
- Phillips, A.; Cobbold, C. A Comparison of the Effects of Aerobic and Intense Exercise on the Type 2 Diabetes Mellitus Risk Marker Adipokines, Adiponectin and Retinol Binding Protein-4. Int. J. Chronic Dis. 2014, 2014, 358058. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.K.; Lam, K.S.; Wang, B.; Xu, A. Signaling mechanisms underlying the insulin-sensitizing effects of adiponectin. Best Pract. Res. Clin. Endocrinol. Metab. 2014, 28, 3–13. [Google Scholar] [CrossRef]
- Hussain, M.K.; Deli, F.A.; Algenabi, A.H.A.; Abdul-Rudha, K.H. Adiponectin gene polymorphisms as a predictor for development of type 2 diabetes mellitus in Iraqi population. Gene 2018, 662, 118–122. [Google Scholar] [CrossRef]
- Siitonen, N.; Pulkkinen, L.; Lindstrom, J.; Kolehmainen, M.; Eriksson, J.G.; Venojarvi, M.; Ilanne-Parikka, P.; Keinänen-Kiukaanniemi, S.; Tuomilehto, J.; Uusitupa, M. Association of ADIPOQ gene variants with body weight, type 2 diabetes and serum adiponectin concentrations: The Finnish Diabetes Prevention Study. BMC Med Genet. 2011, 12, 5. [Google Scholar] [CrossRef]
- Gupta, V.; Mishra, S.; Mishra, S.; Kumar, S.; Gupta, V. Association of Leptin: Adiponectin ratio and metabolic risk markers in postmenopausal women. Immunol. Lett. 2018, 196, 63–67. [Google Scholar] [CrossRef]
- Yadav, A.; Kataria, M.A.; Saini, V.; Yadav, A. Role of leptin and adiponectin in insulin resistance. Clin. Chim. Acta Int. J. Clin. Chem. 2013, 417, 80–84. [Google Scholar] [CrossRef]
- Prates, R.E.; Beretta, M.V.; Nascimento, F.V.; Bernaud, F.R.; de Almeira, J.C.; Rodrigues, T.C. Saturated fatty acid intake decreases serum adiponectin levels in subjects with type 1 diabetes. Diabetes Res. Clin. Pract. 2016, 116, 205–211. [Google Scholar] [CrossRef]
- Tishinsky, J.M.; Robinson, L.E.; Dyck, D.J. Insulin-sensitizing properties of adiponectin. Biochimie 2012, 94, 2131–2136. [Google Scholar] [CrossRef]
- Katira, A.; Tan, P.H. Evolving role of adiponectin in cancer-controversies and update. Cancer Biol. Med. 2016, 13, 101–119. [Google Scholar] [CrossRef] [Green Version]
- Gelsomino, L.; Naimo, G.D.; Catalano, S.; Mauro, L.; Andò, S. The emerging role of adiponectin in female malignancies. Int. J. Mol. Sci. 2019, 20, 2127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barb, D.; Williams, C.J.; Neuwirth, A.K.; Mantzoros, C.S. Adiponectin in relation to malignancies: A review of existing basic research and clinical evidence. Am. J. Clin. Nutr. 2007, 86, s858–s866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pais, R.; Silaghi, H.; Silaghi, A.-C.; Rusu, M.-L.; Dumitrascu, D.-L. Metabolic syndrome and risk of subsequent colorectal cancer. World J. Gastroenterol. 2009, 15, 5141–5148. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.-H.; Yu, B.-Y.; Youn, D.-S. Relationship of serum adiponectin and resistin levels with breast cancer risk. J. Korean Med. Sci. 2007, 22, 117–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaudet, M.M.; Falk, R.T.; Gierach, G.L.; Lacey, J.V., Jr.; Graubard, B.I.; Dorgan, J.F.; Brinton, L.A. Do adipokines underlie the association between known risk factors and breast cancer among a cohort of United States women? Cancer Epidemiol. 2010, 34, 580–586. [Google Scholar] [CrossRef] [Green Version]
- Petridou, E.; Mantzoros, C.; Dessypris, N.; Koukoulomatis, P.; Addy, C.; Voulgaris, Z.; Chrousos, G.; Trichopoulos, D. Plasma adiponectin concentrations in relation to endometrial cancer: A case-control study in Greece. J. Clin. Endocrinol. Metab. 2003, 88, 993–997. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, M.; Kitayama, J.; Yamauchi, T.; Kadowaki, T.; Maki, T.; Miyato, H.; Yamashita, H.; Nagawa, H. Adiponectin inhibits the growth and peritoneal metastasis of gastric cancer through its specific membrane receptors AdipoR1 and AdipoR2. Cancer Sci. 2007, 98, 1120–1127. [Google Scholar] [CrossRef]
- Aleksandrova, K.; Stelmach-Mardas, M.; Schlesinger, S. Recent results in cancer. In Cancer Obesity and Liver; Springer: Berlin/Heidelberg, Germany, 2016; Volume 208, pp. 177–198. [Google Scholar]
- Ebrahimi-Mamaeghani, M.; Mohammadi, S.; Arefhosseini, S.R.; Fallah, P.; Bazi, Z. Adiponectin as a potential biomarker of vascular disease. Vasc. Health Risk Manag. 2015, 11, 55–70. [Google Scholar]
- Kumada, M.; Kihara, S.; Sumitsuji, S.; Kawamoto, T.; Matsumoto, S.; Ouchi, N.; Arita, Y.; Okamoto, Y.; Shimomura, I.; Hiraoka, H.; Nakamura, T.; Funahashi, T.; et al. Association of hypoadiponectinemia with coronary artery disease in men. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 85–89. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, N.; Kanda, J.; Nakamura, T.; Horie, A.; Kurosawa, H.; Hashimoto, T.; Sato, K.; Kushida, S.; Suzuki, M.; Yano, S.; et al. Association of hypoadiponectinemia in men with early onset of coronary heart disease and multiple coronary artery stenoses. Metab. Clin. Exp. 2006, 55, 1653–1657. [Google Scholar] [CrossRef]
- Mitsuhashi, H.; Yatsuya, H.; Tamakoshi, K.; Matsushita, K.; Otsuka, R.; Wada, K.; Sugiura, K.; Takefuji, S.; Hotta, Y.; Kondo, T.; et al. Adiponectin level and left ventricular hypertrophy in Japanese men. Hypertension 2007, 49, 1448–1454. [Google Scholar] [CrossRef] [Green Version]
- Imatoh, T.; Miyazaki, M.; Momose, Y.; Tanihara, S.; Une, H. Adiponectin levels associated with the development of hypertension: A prospective study. Hypertens. Res. Off. J. Jpn. Soc. Hypertens. 2008, 31, 229–233. [Google Scholar] [CrossRef] [Green Version]
- Kozakova, M.; Muscelli, E.; Flyvbjerg, A.; Frystyk, J.; Morizzo, C.; Palombo, C.; Ferrannini, E. Adiponectin and left ventricular structure and function in healthy adults. J. Clin. Endocrinol. Metab. 2008, 93, 2811–2818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leon, B.M.; Maddox, T.M. Diabetes and cardiovascular disease: Epidemiology, biological mechanisms, treatment recommendations and future research. World J. Diabetes 2015, 6, 1246–1258. [Google Scholar] [CrossRef] [PubMed]
- Woodward, L.; Akoumianakis, I.; Antoniades, C. Unravelling the adiponectin paradox: Novel roles of adiponectin in the regulation of cardiovascular disease. Br. J. Pharmacol. 2017, 174, 4007–4020. [Google Scholar] [CrossRef] [PubMed]
- Marchal, P.O.; Kavvadas, P.; Abed, A.; Kazazian, C.; Authier, F.; Koseki, H.; Hiraoka, S.; Boffa, J.J.; Martinerie, C.; Chadjichristos, C.E. Reduced NOV/CCN3 Expression Limits Inflammation and Interstitial Renal Fibrosis after Obstructive Nephropathy in Mice. PLoS ONE 2015, 10, e0137876. [Google Scholar] [CrossRef] [PubMed]
- Pakradouni, J.; Le Goff, W.; Calmel, C.; Antoine, B.; Villard, E.; Frisdal, E.; Abifadel, M.; Tordjman, J.; Poitou, C.; Bonnefont-Rousselot, D. Plasma NOV/CCN3 levels are closely associated with obesity in patients with metabolic disorders. PLoS ONE 2013, 8, e66788. [Google Scholar] [CrossRef] [Green Version]
- Twig, G.; Shirihai, O.S. The interplay between mitochondrial dynamics and mitophagy. Antioxid. Redox Signal. 2011, 14, 1939–1951. [Google Scholar] [CrossRef] [Green Version]
- Hull, T.D.; Boddu, R.; Guo, L.; Tisher, C.C.; Traylor, A.M.; Patel, B.; Joseph, R.; Prabhu, S.D.; Suliman, H.B.; Piantadosi, C.A. Heme oxygenase-1 regulates mitochondrial quality control in the heart. JCI Insight 2016, 1, e85817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.P.; Schragenheim, J.; Cao, J.; Falck, J.R.; Abraham, N.G.; Bellner, L. PGC-1 alpha regulates HO-1 expression, mitochondrial dynamics and biogenesis: Role of epoxyeicosatrienoic acid. Prostaglandins Other Lipid Mediat. 2016, 125, 8–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.P.; McClung, J.A.; Bellner, L.; Cao, J.; Waldman, M.; Schragenheim, J.; Arad, M.; Hochhauser, E.; Falck, J.R.; Weingarten, J.A.; et al. CYP-450 Epoxygenase Derived Epoxyeicosatrienoic Acid Contribute to Reversal of Heart Failure in Obesity-Induced Diabetic Cardiomyopathy via PGC-1 alpha Activation. Cardiovasc. Pharmacol. Open Access 2018, 7, 233. [Google Scholar]
- Rowe, G.C.; Jiang, A.; Arany, Z. PGC-1 coactivators in cardiac development and disease. Circ. Res. 2010, 107, 825–838. [Google Scholar] [CrossRef] [Green Version]
- Delaby, C.; Gabelle, A.; Blum, D.; Schraen-Maschke, S.; Moulinier, A.; Boulanghien, J.; Séverac, D.; Buée, L.; Rème, T.; Lehmann, S. Central Nervous System and Peripheral Inflammatory Processes in Alzheimer’s Disease: Biomarker Profiling Approach. Front. Neurol. 2015, 6, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalaria, R.N.; Maestre, G.E.; Arizaga, R.; Friedland, R.P.; Galasko, D.; Hall, K.; Luchsinger, J.A.; Ogunniyi, A.; Perry, E.K.; Potocnik, F.; et al. Alzheimer’s disease and vascular dementia in developing countries: Prevalence, management, and risk factors. Lancet Neurol. 2008, 7, 812–826. [Google Scholar] [CrossRef] [Green Version]
- Talbot, K.; Wang, H.Y.; Kazi, H.; Han, L.Y.; Bakshi, K.P.; Stucky, A.; Fuino, R.L.; Kawaguchi, K.R.; Samoyedny, A.J.; Wilson, R.S.; et al. Demonstrated brain insulin resistance in Alzheimer’s disease patients is associated with IGF-1 resistance, IRS-1 dysregulation, and cognitive decline. J. Clin. Investig. 2012, 122, 1316–1338. [Google Scholar] [CrossRef] [Green Version]
- Wan, Z.; Little, J.P.J.I.; Signaling, C. Adiponectin and Alzheimer’s disease: Is there a link? Inflamm. Cell Signal. 2014, 1. [Google Scholar] [CrossRef]
- Mullins, R.J.; Diehl, T.C.; Chia, C.W.; Kapogiannis, D. Insulin Resistance as a Link between Amyloid-Beta and Tau Pathologies in Alzheimer’s Disease. Front. Aging Neurosci. 2017, 9, 118. [Google Scholar] [CrossRef] [PubMed]
- Cova, I.; Clerici, F.; Rossi, A.; Cucumo, V.; Ghiretti, R.; Maggiore, L.; Pomati, S.; Galimberti, D.; Scarpini, E.; Mariani, C.; et al. Weight loss predicts progression of mild cognitive impairment to Alzheimer’s disease. PLoS ONE 2016, 11, e0151710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pakaski, M.; Feher, A.; Juhasz, A.; Drotos, G.; Fazekas, O.C.; Kovacs, J.; Janka, Z.; Kálmán, J. Serum adipokine levels modified by donepezil treatment in Alzheimer’s disease. J. Alzheimer’s Dis. JAD 2014, 38, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.G.; Javorschi, S.; Hevener, A.L.; Kruszynska, Y.T.; Norman, R.A.; Sinha, M.; Olefsky, J.M. The effect of thiazolidinediones on plasma adiponectin levels in normal, obese, and type 2 diabetic subjects. Diabetes 2002, 51, 2968–2974. [Google Scholar] [CrossRef] [Green Version]
- Amin, R.H.; Mathews, S.T.; Camp, H.S.; Ding, L.; Leff, T. Selective activation of PPARgamma in skeletal muscle induces endogenous production of adiponectin and protects mice from diet-induced insulin resistance. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E28–E37. [Google Scholar] [CrossRef]
- Plaisance, E.P.; Lukasova, M.; Offermanns, S.; Zhang, Y.; Cao, G.; Judd, R.L. Niacin stimulates adiponectin secretion through the GPR109A receptor. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E549–E558. [Google Scholar] [CrossRef] [Green Version]
- Rosenson, R.S. Effect of fenofibrate on adiponectin and inflammatory biomarkers in metabolic syndrome patients. Obesity 2009, 17, 504–509. [Google Scholar] [CrossRef]
- Katsiki, N.; Mantzoros, C.S. Statins in relation to adiponectin: A significant association with clinical implications. Atherosclerosis 2016, 253, 270–272. [Google Scholar] [CrossRef] [Green Version]
- Ng, R.C.; Chan, K.H. Potential Neuroprotective Effects of Adiponectin in Alzheimer’s Disease. Int. J. Mol. Sci. 2017, 18, 592. [Google Scholar] [CrossRef] [Green Version]
- Bloemer, J.; Pinky, P.D.; Govindarajulu, M.; Hong, H.; Judd, R.; Amin, R.H.; Moore, T.; Dhanasekaran, M.; Reed, M.N.; Suppiramaniam, V. Role of Adiponectin in Central Nervous System Disorders. Neural Plast. 2018, 2018, 4593530. [Google Scholar] [CrossRef] [Green Version]
- van Himbergen, T.M.; Beiser, A.S.; Ai, M.; Seshadri, S.; Otokozawa, S.; Au, R.; Thongtang, N.; Wolf, P.A.; Schaefer, E. Biomarkers for insulin resistance and inflammation and the risk for all-cause dementia and alzheimer disease: Results from the Framingham Heart Study. Arch. Neurol. 2012, 69, 594–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letra, L.; Rodrigues, T.; Matafome, P.; Santana, I.; Seica, R. Adiponectin and sporadic Alzheimer’s disease: Clinical and molecular links. Front. Neuroendocrinol. 2019, 52, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Izadi, V.; Azadbakht, L. Specific dietary patterns and concentrations of adiponectin. J. Res. Med. Sci. Off. J. Isfahan Univ. Med. Sci. 2015, 20, 178–184. [Google Scholar]
- Paschos, G.K.; Zampelas, A.; Panagiotakos, D.B.; Katsiougiannis, S.; Griffin, B.A.; Votteas, V.; Skopouli, F.N. Effects of flaxseed oil supplementation on plasma adiponectin levels in dyslipidemic men. Eur. J. Nutr. 2007, 46, 315–320. [Google Scholar] [CrossRef]
- Flachs, P.; Mohamed-Ali, V.; Horakova, O.; Rossmeisl, M.; Hosseinzadeh-Attar, M.J.; Hensler, M.; Ruzickova, J.; Kopecky, J. Polyunsaturated fatty acids of marine origin induce adiponectin in mice fed a high-fat diet. Diabetologia 2006, 49, 394–397. [Google Scholar] [CrossRef] [Green Version]
- Bahreini, M.; Ramezani, A.H.; Shishehbor, F.; Mansoori, A. The Effect of Omega-3 on Circulating Adiponectin in Adults with Type 2 Diabetes Mellitus: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Can. J. Diabetes 2018, 42, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Oster, R.T.; Tishinsky, J.M.; Yuan, Z.; Robinson, L.E. Docosahexaenoic acid increases cellular adiponectin mRNA and secreted adiponectin protein, as well as PPARgamma mRNA.; in 3T3-L1 adipocytes. Appl. Physiol. Nutr. Metab. 2010, 35, 783–789. [Google Scholar] [CrossRef]
- Esmaillzadeh, A.; Azadbakht, L. Legume consumption is inversely associated with serum concentrations of adhesion molecules and inflammatory biomarkers among Iranian women. J. Nutr. 2012, 142, 334–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rokling-Andersen, M.H.; Reseland, J.E.; Veierod, M.B.; Anderssen, S.A.; Jacobs, D.R., Jr.; Urdal, P.; Jansson, J.O.; Drevon, C.A. Effects of long-term exercise and diet intervention on plasma adipokine concentrations. Am. J. Clin. Nutr. 2007, 86, 1293–1301. [Google Scholar] [CrossRef] [Green Version]
- Tsukinoki, R.; Morimoto, K.; Nakayama, K. Association between lifestyle factors and plasma adiponectin levels in Japanese men. Lipids Health Dis. 2005, 4, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa, M.M.; Melo, A.L.; Damasceno, N.R. The benefits of omega-3 supplementation depend on adiponectin basal level and adiponectin increase after the supplementation: A randomized clinical trial. Nutrition 2017, 34, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Mejia-Montilla, J.; Reyna-Villasmil, E.; Dominguez-Brito, L.; Naranjo-Rodriguez, C.; Noriega-Verdugo, D.; Padilla-Samaniego, M.; Vargas-Olalla, V. Supplementation with omega-3 fatty acids and plasma adiponectin in women with polycystic ovary syndrome. Endocrinol. Diabetes Y Nutr. 2018, 65, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Rizza, S.; Tesauro, M.; Cardillo, C.; Galli, A.; Iantorno, M.; Gigli, F.; Sbraccia, P.; Federici, M.; Quon, M.J.; Lauro, D. Fish oil supplementation improves endothelial function in normoglycemic offspring of patients with type 2 diabetes. Atherosclerosis 2009, 206, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Mostowik, M.; Gajos, G.; Zalewski, J.; Nessler, J.; Undas, A. Omega-3 polyunsaturated fatty acids increase plasma adiponectin to leptin ratio in stable coronary artery disease. Cardiovasc. Drugs Ther. 2013, 27, 289–295. [Google Scholar] [CrossRef] [Green Version]
- Silva, F.M.; de Almeida, J.C.; Feoli, A.M. Effect of diet on adiponectin levels in blood. Nutr. Rev. 2011, 69, 599–612. [Google Scholar] [CrossRef]
- Bergqvist, S.; Ghosh, G.; Komives, E.A. The IkappaBalpha/NF-kappaB complex has two hot spots, one at either end of the interface. Protein Sci. A Publ. Protein Soc. 2008, 17, 2051–2058. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Insulin | Adiponectin | |
|---|---|---|
| Lipogenesis | ↑ | ↓ |
| Fat oxidation | ↓ | ↑ |
| Glycogenolysis | ↓ | ↓ |
| Gluconeogenesis | ↓ | ↓ |
| Glycolysis | ↑ | - |
| Glycogen synthesis | ↑ | - |
| Organ | Adiponectin Functions |
|---|---|
| Brain |
|
| Liver |
|
| Muscle |
|
| Heart |
|
| Kidney |
|
| Bone |
|
| Disease | Effect on Adiponectin |
|---|---|
| Diabetes |
|
| Cancer |
|
| Cardiovascular Diseases |
|
| Alzheimer’s disease |
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khoramipour, K.; Chamari, K.; Hekmatikar, A.A.; Ziyaiyan, A.; Taherkhani, S.; Elguindy, N.M.; Bragazzi, N.L. Adiponectin: Structure, Physiological Functions, Role in Diseases, and Effects of Nutrition. Nutrients 2021, 13, 1180. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13041180
Khoramipour K, Chamari K, Hekmatikar AA, Ziyaiyan A, Taherkhani S, Elguindy NM, Bragazzi NL. Adiponectin: Structure, Physiological Functions, Role in Diseases, and Effects of Nutrition. Nutrients. 2021; 13(4):1180. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13041180
Chicago/Turabian StyleKhoramipour, Kayvan, Karim Chamari, Amirhosein Ahmadi Hekmatikar, Amirhosein Ziyaiyan, Shima Taherkhani, Nihal M. Elguindy, and Nicola Luigi Bragazzi. 2021. "Adiponectin: Structure, Physiological Functions, Role in Diseases, and Effects of Nutrition" Nutrients 13, no. 4: 1180. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13041180