
Creatine Monohydrate Supplementation Increases White Adipose Tissue Mitochondrial Markers in Male and Female Rats in a Depot Specific Manner

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Study Design
2.2. Tissue Collection and Homogenization
2.3. Western Blotting
2.4. Real-Time qPCR
2.5. Histology
2.6. Statistical Analysis
3. Results
3.1. Animal and Adipose Tissue Depot Characteristics
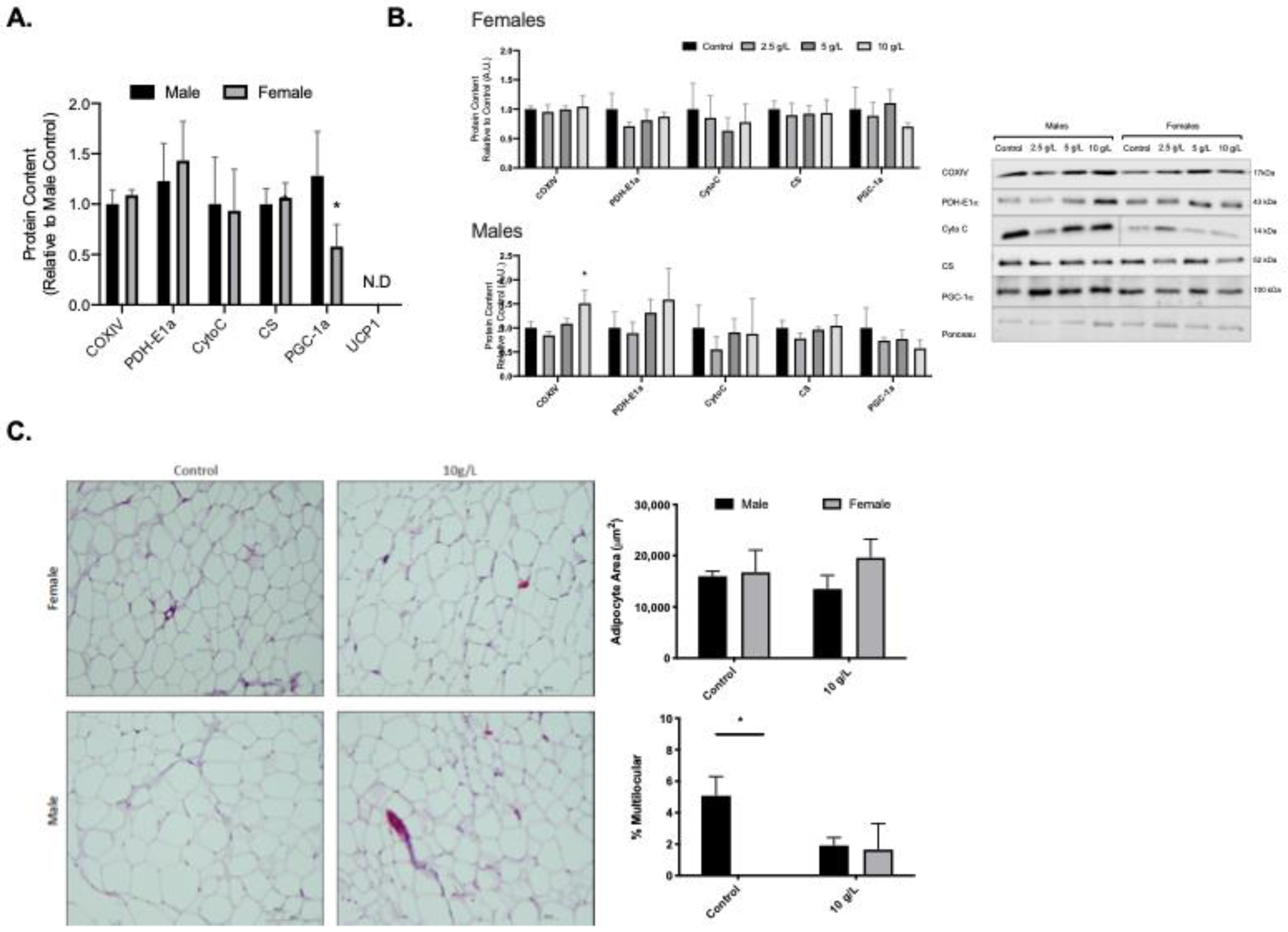
3.2. Creatine Supplementation Increases Mitochondrial Markers in Female Rat Inguinal White Adipose Tissue
3.3. Creatine Supplementation increases Mitochondrial Markers in Male Rat Gonadal White Adipose Tissue
3.4. Creatine Supplementation does Not increase Mitochondrial Markers Brown Adipose Tissue in either Male or Female Rats
3.5. Creatine Supplementation Alters Markers of Creatine Uptake and Metabolism
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Berlet, H.H.; Bonsmann, I.; Birringer, H. Occurrence of free creatine, phosphocreatine and creatine phosphokinase in adipose tissue. Biochim. Biophys. Acta 1976, 437, 166–174. [Google Scholar] [CrossRef]
- Bertholet, A.M.; Kazak, L.; Chouchani, E.T.; Bogaczynska, M.G.; Paranjpe, I.; Wainwright, G.L.; Betourne, A.; Kajimura, S.; Spiegelman, B.M.; Kirichok, Y. Mitochondrial Patch Clamp of Beige Adipocytes Reveals UCP1-Positive and UCP1-Negative Cells Both Exhibiting Futile Creatine Cycling. Cell Metab. 2017, 25, 811–822.e4. [Google Scholar] [CrossRef] [Green Version]
- Buford, T.W.; Kreider, R.B.; Stout, J.R.; Greenwood, M.; Campbell, B.; Spano, M.; Ziegenfuss, T.; Lopez, H.; Landis, J.; Antonio, J. International Society of Sports Nutrition position stand: Creatine supplementation and exercise. J. Int. Soc. Sports Nutr. 2007, 4, 6. [Google Scholar] [CrossRef] [Green Version]
- Cannon, B.; Nedergaard, J. Brown adipose tissue: Function and physiological significance. Physiol. Rev. 2004, 84, 277–359. [Google Scholar] [CrossRef] [PubMed]
- Chang, E.; Varghese, M.; Singer, K. Gender and Sex Differences in Adipose Tissue. Curr. Diabetes Rep. 2018, 18, 69. [Google Scholar] [CrossRef]
- Choi, C.H.J.; Cohen, P. Adipose crosstalk with other cell types in health and disease. Exp. Cell Res. 2017, 360, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Chouchani, E.T.; Kazak, L.; Spiegelman, B.M. New Advances in Adaptive Thermogenesis: UCP1 and Beyond. Cell Metab. 2019, 29, 27–37. [Google Scholar] [CrossRef]
- Cinti, S. Transdifferentiation properties of adipocytes in the adipose organ. Am. J. Physiol. Endocrinol. Metab. 2009, 297, E977–E986. [Google Scholar] [CrossRef] [PubMed]
- Cohen, P.; Levy, J.D.; Zhang, Y.; Frontini, A.; Kolodin, D.P.; Svensson, K.J.; Lo, J.C.; Zeng, X.; Ye, L.; Khandekar, M.J.; et al. Ablation of PRDM16 and beige adipose causes metabolic dysfunction and a subcutaneous to visceral fat switch. Cell 2014, 156, 304–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, R.; Naclerio, F.; Allgrove, J.; Jimenez, A. Creatine supplementation with specific view to exercise/sports performance: An update. J. Int. Soc. Sports Nutr. 2012, 9, 33. [Google Scholar] [CrossRef] [Green Version]
- Enerback, S. The origins of brown adipose tissue. N. Engl. J. Med. 2009, 360, 2021–2023. [Google Scholar] [CrossRef] [PubMed]
- Fedorenko, A.; Lishko, P.V.; Kirichok, Y. Mechanism of fatty-acid-dependent UCP1 uncoupling in brown fat mitochondria. Cell 2012, 151, 400–413. [Google Scholar] [CrossRef] [Green Version]
- Gesta, S.; Bluher, M.; Yamamoto, Y.; Norris, A.W.; Berndt, J.; Kralisch, S.; Boucher, J.; Lewis, C.; Kahn, C.R. Evidence for a role of developmental genes in the origin of obesity and body fat distribution. Proc. Natl. Acad. Sci. USA 2006, 103, 6676–6681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granneman, J.G.; Burnazi, M.; Zhu, Z.; Schwamb, L.A. White adipose tissue contributes to UCP1-independent thermogenesis. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E1230–E1236. [Google Scholar] [CrossRef] [Green Version]
- Grimpo, K.; Volker, M.N.; Heppe, E.N.; Braun, S.; Heverhagen, J.T.; Heldmaier, G. Brown adipose tissue dynamics in wild-type and UCP1-knockout mice: In vivo insights with magnetic resonance. J. Lipid Res. 2014, 55, 398–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirshman, M.F.; Wardzala, L.J.; Goodyear, L.J.; Fuller, S.P.; Horton, E.D.; Horton, E.S. Exercise training increases the number of glucose transporters in rat adipose cells. Am. J. Physiol. 1989, 257, E520–E530. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, K.; Kang, Q.; Yoneshiro, T.; Camporez, J.P.; Maki, H.; Homma, M.; Shinoda, K.; Chen, Y.; Lu, X.; Maretich, P.; et al. UCP1-independent signaling involving SERCA2b-mediated calcium cycling regulates beige fat thermogenesis and systemic glucose homeostasis. Nat. Med. 2017, 23, 1454–1465. [Google Scholar] [CrossRef] [Green Version]
- Ipsiroglu, O.S.; Stromberger, C.; Ilas, J.; Hoger, H.; Muhl, A.; Stockler-Ipsiroglu, S. Changes of tissue creatine concentrations upon oral supplementation of creatine-monohydrate in various animal species. Life Sci. 2001, 69, 1805–1815. [Google Scholar] [CrossRef]
- Kazak, L.; Chouchani, E.T.; Jedrychowski, M.P.; Erickson, B.K.; Shinoda, K.; Cohen, P.; Vetrivelan, R.; Lu, G.Z.; Laznik-Bogoslavski, D.; Hasenfuss, S.C.; et al. A creatine-driven substrate cycle enhances energy expenditure and thermogenesis in beige fat. Cell 2015, 163, 643–655. [Google Scholar] [CrossRef] [Green Version]
- Kazak, L.; Chouchani, E.T.; Lu, G.Z.; Jedrychowski, M.P.; Bare, C.J.; Mina, A.I.; Kumari, M.; Zhang, S.; Vuckovic, I.; Laznik-Bogoslavski, D.; et al. Genetic Depletion of Adipocyte Creatine Metabolism Inhibits Diet-Induced Thermogenesis and Drives Obesity. Cell Metab. 2017, 26, 693. [Google Scholar] [CrossRef]
- Kazak, L.; Rahbani, J.F.; Samborska, B.; Lu, G.Z.; Jedrychowski, M.P.; Lajoie, M.; Zhang, S.; Ramsay, L.C.; Dou, F.Y.; Tenen, D.; et al. Ablation of adipocyte creatine transport impairs thermogenesis and causes diet-induced obesity. Nat. Metab. 2019, 1, 360–370. [Google Scholar] [CrossRef]
- Kopecky, J.; Clarke, G.; Enerback, S.; Spiegelman, B.; Kozak, L.P. Expression of the mitochondrial uncoupling protein gene from the aP2 gene promoter prevents genetic obesity. J. Clin. Investig. 1995, 96, 2914–2923. [Google Scholar] [CrossRef]
- Kreider, R.B. Effects of creatine supplementation on performance and training adaptations. Mol. Cell. Biochem. 2003, 244, 89–94. [Google Scholar] [CrossRef]
- Kreider, R.B.; Melton, C.; Rasmussen, C.J.; Greenwood, M.; Lancaster, S.; Cantler, E.C.; Milnor, P.; Almada, A.L. Long-term creatine supplementation does not significantly affect clinical markers of health in athletes. Mol. Cell. Biochem. 2003, 244, 95–104. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Lowell, B.B.; S-Susulic, V.; Hamann, A.; Lawitts, J.A.; Himms-Hagen, J.; Boyer, B.B.; Kozak, L.P.; Flier, J.S. Development of obesity in transgenic mice after genetic ablation of brown adipose tissue. Nature 1993, 366, 740–742. [Google Scholar] [CrossRef]
- Macotela, Y.; Boucher, J.; Tran, T.T.; Kahn, C.R. Sex and depot differences in adipocyte insulin sensitivity and glucose metabolism. Diabetes 2009, 58, 803–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacPherson, R.E.; Baumeister, P.; Peppler, W.T.; Wright, D.C.; Little, J.P. Reduced cortical BACE1 content with one bout of exercise is accompanied by declines in AMPK, Akt, and MAPK signaling in obese, glucose-intolerant mice. J. Appl. Physiol. 2015, 119, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- McKie, G.L.; Wright, D.C. Biochemical adaptations in white adipose tissue following aerobic exercise: From mitochondrial biogenesis to browning. Biochem. J. 2020, 477, 1061–1081. [Google Scholar] [CrossRef]
- Misra, A.; Garg, A.; Abate, N.; Peshock, R.M.; Stray-Gundersen, J.; Grundy, S.M. Relationship of anterior and posterior subcutaneous abdominal fat to insulin sensitivity in nondiabetic men. Obes. Res. 1997, 5, 93–99. [Google Scholar] [CrossRef]
- Muller, S.; Balaz, M.; Stefanicka, P.; Varga, L.; Amri, E.Z.; Ukropec, J.; Wollscheid, B.; Wolfrum, C. Proteomic Analysis of Human Brown Adipose Tissue Reveals Utilization of Coupled and Uncoupled Energy Expenditure Pathways. Sci. Rep. 2016, 6, 30030. [Google Scholar] [CrossRef] [Green Version]
- Dunham, T.C.; Murphy, J.E.; MacPherson, R.E.; Fajardo, V.A.; Ward, W.E.; Roy, B.D. Sex-and tissue-dependent creatine uptake in response to different creatine monohydrate doses in male and female Sprague-Dawley rats. Appl. Physiol. Nutr. Metab. 2021. [Google Scholar] [CrossRef]
- Petrovic, N.; Walden, T.B.; Shabalina, I.G.; Timmons, J.A.; Cannon, B.; Nedergaard, J. Chronic peroxisome proliferator-activated receptor γ (PPARγ) activation of epididymally derived white adipocyte cultures reveals a population of thermogenically competent, UCP1-containing adipocytes molecularly distinct from classic brown adipocytes. J. Biol. Chem. 2010, 285, 7153–7164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahbani, J.F.; Roesler, A.; Hussain, M.F.; Samborska, B.; Dykstra, C.B.; Tsai, L.; Jedrychowski, M.P.; Vergnes, L.; Reue, K.; Spiegelman, B.M.; et al. Creatine kinase B controls futile creatine cycling in thermogenic fat. Nature 2021, 590, 480–485. [Google Scholar] [CrossRef] [PubMed]
- Roesler, A.; Kazak, L. UCP1-independent thermogenesis. Biochem. J. 2020, 477, 709–725. [Google Scholar] [CrossRef] [PubMed]
- Rosenwald, M.; Perdikari, A.; Rulicke, T.; Wolfrum, C. Bi-directional interconversion of brite and white adipocytes. Nat. Cell Biol. 2013, 15, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Sebo, Z.L.; Rodeheffer, M.S. Assembling the adipose organ: Adipocyte lineage segregation and adipogenesis in vivo. Development 2019, 146, dev172098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snijder, M.B.; Dekker, J.M.; Visser, M.; Bouter, L.M.; Stehouwer, C.D.; Kostense, P.J.; Yudkin, J.S.; Heine, R.J.; Nijpels, G.; Seidell, J.C. Associations of hip and thigh circumferences independent of waist circumference with the incidence of type 2 diabetes: The Hoorn Study. Am. J. Clin. Nutr. 2003, 77, 1192–1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somjen, D.; Tordjman, K.; Waisman, A.; Mor, G.; Amir-Zaltsman, Y.; Kohen, F.; Kaye, A.M. Estrogen stimulation of creatine kinase B specific activity in 3T3L1 adipocytes after their differentiation in culture: Dependence on estrogen receptor. J. Steroid Biochem. Mol. Biol. 1997, 62, 401–408. [Google Scholar] [CrossRef]
- Stallknecht, B.; Vinten, J.; Ploug, T.; Galbo, H. Increased activities of mitochondrial enzymes in white adipose tissue in trained rats. Am. J. Physiol. 1991, 261, E410–E414. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Rahbani, J.F.; Jedrychowski, M.P.; Riley, C.L.; Vidoni, S.; Bogoslavski, D.; Hu, B.; Dumesic, P.A.; Zeng, X.; Wang, A.B.; et al. Mitochondrial TNAP controls thermogenesis by hydrolysis of phosphocreatine. Nature 2021, 593, 580–585. [Google Scholar] [CrossRef]
- Tchkonia, T.; Lenburg, M.; Thomou, T.; Giorgadze, N.; Frampton, G.; Pirtskhalava, T.; Cartwright, A.; Cartwright, M.; Flanagan, J.; Karagiannides, I.; et al. Identification of depot-specific human fat cell progenitors through distinct expression profiles and developmental gene patterns. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E298–E307. [Google Scholar] [CrossRef] [Green Version]
- Townsend, L.K.; Wright, D.C. Looking on the “brite” side exercise-induced browning of white adipose tissue. Pflug. Arch. Eur. J. Physiol. 2019, 471, 455–465. [Google Scholar] [CrossRef]
- Tran, T.T.; Yamamoto, Y.; Gesta, S.; Kahn, C.R. Beneficial effects of subcutaneous fat transplantation on metabolism. Cell Metab. 2008, 7, 410–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trayhurn, P. Endocrine and signalling role of adipose tissue: New perspectives on fat. Acta Physiol. Scand. 2005, 184, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Ukropec, J.; Anunciado, R.P.; Ravussin, Y.; Hulver, M.W.; Kozak, L.P. UCP1-independent thermogenesis in white adipose tissue of cold-acclimated Ucp1−/− mice. J. Biol. Chem. 2006, 281, 31894–31908. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, L.J.; Oosterlaar, A.M.; Hartgens, F.; Hesselink, M.K.; Snow, R.J.; Wagenmakers, A.J. Effects of creatine loading and prolonged creatine supplementation on body composition, fuel selection, sprint and endurance performance in humans. Clin. Sci. 2003, 104, 153–162. [Google Scholar] [CrossRef]
- Veniant, M.M.; Sivits, G.; Helmering, J.; Komorowski, R.; Lee, J.; Fan, W.; Moyer, C.; Lloyd, D.J. Pharmacologic Effects of FGF21 Are Independent of the “Browning” of White Adipose Tissue. Cell Metab. 2015, 21, 731–738. [Google Scholar] [CrossRef] [Green Version]
- Wakatsuki, T.; Hirata, F.; Ohno, H.; Yamamoto, M.; Sato, Y.; Ohira, Y. Thermogenic responses to high-energy phosphate contents and/or hindlimb suspension in rats. Jpn. J. Physiol. 1996, 46, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Rimm, E.B.; Stampfer, M.J.; Willett, W.C.; Hu, F.B. Comparison of abdominal adiposity and overall obesity in predicting risk of type 2 diabetes among men. Am. J. Clin. Nutr. 2005, 81, 555–563. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, H.; Ohira, Y.; Wakatsuki, T.; Yamamoto, M.; Kizaki, T.; Oh-ishi, S.; Ohno, H. Increased growth of brown adipose tissue but its reduced thermogenic activity in creatine-depleted rats fed β-guanidinopropionic acid. Biochim. Biophys. Acta 1995, 1230, 69–73. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Rexrode, K.M.; van Dam, R.M.; Li, T.Y.; Hu, F.B. Abdominal obesity and the risk of all-cause, cardiovascular, and cancer mortality: Sixteen years of follow-up in US women. Circulation 2008, 117, 1658–1667. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal Characteristics | ||||
| Creatine (g·L−1) | 0 | 2.5 $ | 5 & | 10 |
| End point body mass (g) | ||||
| Males | 587.8 ± 9.1 | 602.8 ± 14.1 | 610.3 ± 25.3 | 584.5 ±23.0 |
| Females | 376.8 ± 13.5 * | 416.3 ±13.5 * | 397.0 ± 7.1 * | 404.8 ± 28.3 * |
| Food Intake (g·day−1) | ||||
| Males | 24.4 ± 0.3 | 24.3 ± 0.9 | 23.7 ± 1.7 | 23.8 ± 1.3 |
| Females | 17.7 ± 1.2 * | 19.0 ± 0.7 * | 18.3 ± 0.1 * | 18.75 ± 1.2 * |
| Water Intake (mL·day−1) | ||||
| Males | 57.9 ± 14.0 | 44.9 ± 8.7 | 36.1 ± 2.2 # | 43.5 ± 1.0 |
| Females | 34.7 ± 4.6 * | 35.8 ± 5.4 * | 35.4 ± 1.9 * | 27.9 ± 1.3 * |
| Gonadal Adipose Tissue Protein Content (ug protein/mg tissue) | ||||
| Males | 19.4 ± 1.3 | 19.4 ± 1.2 | 18.2 ± 0.3 | 19.1 ± 1.3 |
| Females | 19.5 ± 1.0 | 18.0 ± 1.1 | 18.1 ± 0.3 | 18.1 ± 0.7 |
| Inguinal Adipose Tissue Protein Content (ug protein/mg tissue) | ||||
| Males | 28.2 ± 1.2 | 27.5 ± 3.9 | 33.9 ± 6.0 | 29.0 ± 0.9 |
| Females | 29.0 ± 4.6 | 25.1 ± 1.0 | 29.8 ± 1.7 | 29.9 ± 0.9 |
| Interscapular Brown Adipose Tissue Protein Content (ug protein/mg tissue) | ||||
| Males | 93.4 ± 5.4 | 111.6 ± 12.2 | 99.6 ± 9.4 | 107.2 ± 10.8 |
| Females | 112.0 ± 9.4 | 106.3 ± 12.2 | 117.2 ± 7.6 | 120.9 ± 3.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ryan, C.R.; Finch, M.S.; Dunham, T.C.; Murphy, J.E.; Roy, B.D.; MacPherson, R.E.K. Creatine Monohydrate Supplementation Increases White Adipose Tissue Mitochondrial Markers in Male and Female Rats in a Depot Specific Manner. Nutrients 2021, 13, 2406. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13072406
Ryan CR, Finch MS, Dunham TC, Murphy JE, Roy BD, MacPherson REK. Creatine Monohydrate Supplementation Increases White Adipose Tissue Mitochondrial Markers in Male and Female Rats in a Depot Specific Manner. Nutrients. 2021; 13(7):2406. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13072406
Chicago/Turabian StyleRyan, Chantal R., Michael S. Finch, Tyler C. Dunham, Jensen E. Murphy, Brian D. Roy, and Rebecca E. K. MacPherson. 2021. "Creatine Monohydrate Supplementation Increases White Adipose Tissue Mitochondrial Markers in Male and Female Rats in a Depot Specific Manner" Nutrients 13, no. 7: 2406. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13072406