
Probiotic Supplementation and Micronutrient Status in Healthy Subjects: A Systematic Review of Clinical Trials

 ,
, 
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
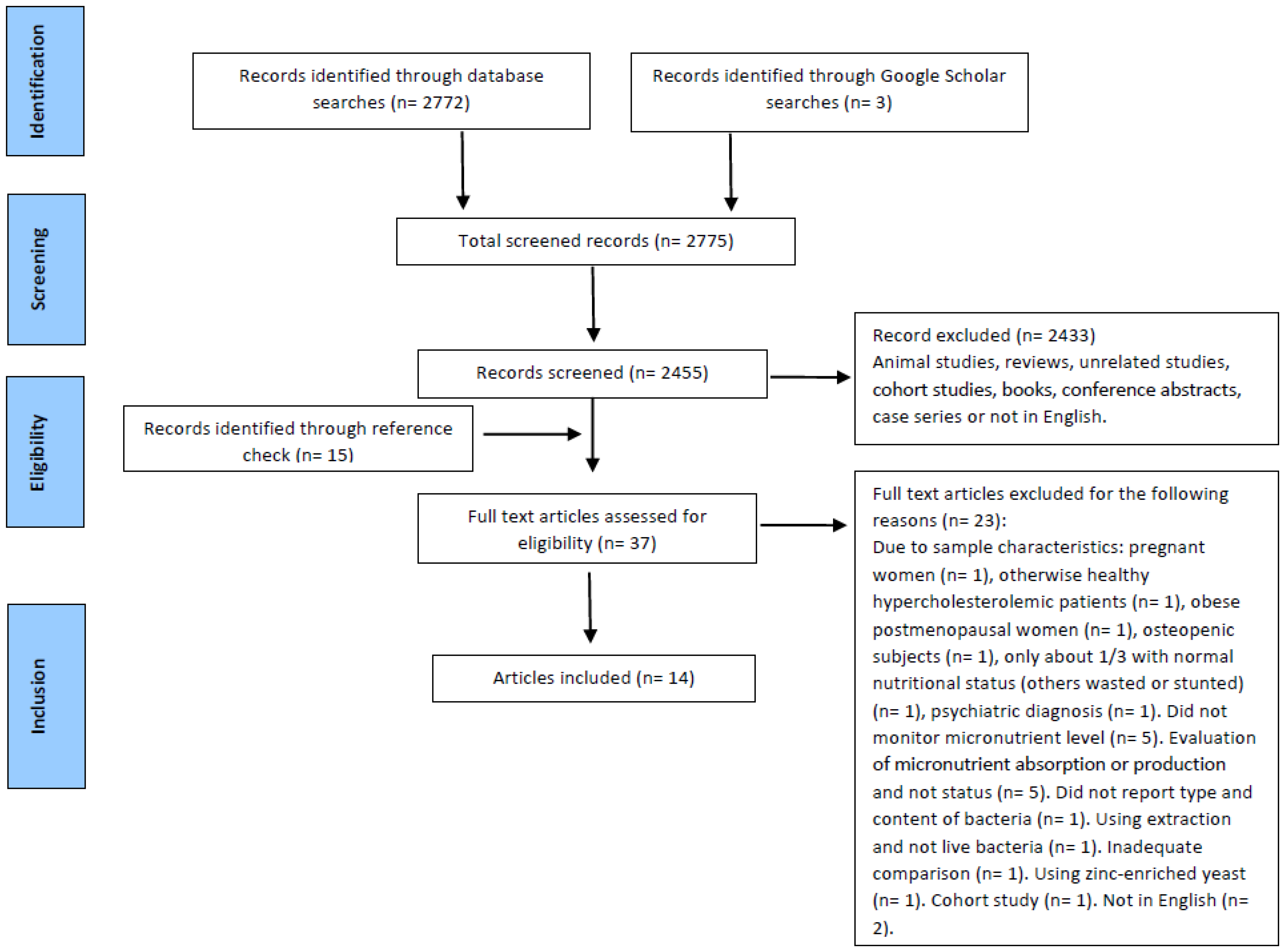
2.1. Literature Search Strategy
2.2. Eligibility Criteria and Study Selection
2.3. Data Abstraction
2.4. Quality Assessment
3. Results and Discussion
3.1. Study Selection
3.2. Quality Assessment
3.3. Characteristics of the Included Studies
3.3.1. Probiotic Supplementation and Fat-Soluble Vitamin and Carotenoid Status
3.3.2. Probiotic Supplementation and Vitamin B Group Status
3.3.3. Probiotic Supplementation and Mineral Status
3.4. Gaps in Current Research and Reasons for the Conflicting Results
4. Strengths and Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Appendix A
Appendix A.1. PubMed Search Strategies
Appendix A.2. Scopus Search Strategy
Appendix A.3. Embase Search Strategy
References
- Tulchinsky, T.H. Micronutrient deficiency conditions: Global health issues. Public Health Rev. 2010, 32, 243. [Google Scholar] [CrossRef] [Green Version]
- Diaz, J.; De Las Cagigas, A.; Rodriguez, R. Micronutrient deficiencies in developing and affluent countries. Eur. J. Clin. Nutr. 2003, 57, S70–S72. [Google Scholar] [CrossRef]
- Keats, E.C.; Neufeld, L.M.; Garrett, G.S.; Mbuya, M.N.N.; Bhutta, Z.A. Improved micronutrient status and health outcomes in low- and middle-income countries following large-scale fortification: Evidence from a systematic review and meta-analysis. Am. J. Clin. Nutr. 2019, 109, 1696–1708. [Google Scholar] [CrossRef] [Green Version]
- LeBlanc, J.G.; Milani, C.; De Giori, G.S.; Sesma, F.; Van Sinderen, D.; Ventura, M. Bacteria as vitamin suppliers to their host: A gut microbiota perspective. Curr. Opin. Biotechnol. 2013, 24, 160–168. [Google Scholar] [CrossRef]
- Willing, B.; Van Kessel, A. Host pathways for recognition: Establishing gastrointestinal microbiota as relevant in animal health and nutrition. Livest. Sci. 2010, 133, 82–91. [Google Scholar] [CrossRef]
- WHO; Food and Agriculture Organization of the United Nations. Report of a Joint FAO/WHO expert consultation of evaluations of health and nutritional properties of probiotics in food including powder milk and live lactic acid bacteria in Cordoba, Argentina. In Probiotics in Food: Health and Nutritional Properties and Guidelines for Evaluation; Amerian Córdoba Park Hotel: Córdoba, Argentina, 2001. [Google Scholar]
- Martín, R.; Langella, P. Emerging health concepts in the probiotics field: Streamlining the definitions. Front. Microbiol. 2019, 10, 1047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mokoena, M.P.; Mutanda, T.; Olaniran, A.O. Perspectives on the probiotic potential of lactic acid bacteria from African traditional fermented foods and beverages. J. Food Nutr. Res. 2016, 60, 29630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bezkorovainy, A. Probiotics: Determinants of survival and growth in the gut. Am. J. Clin. Nutr. 2001, 73, 399s–405s. [Google Scholar] [CrossRef] [PubMed]
- Pompei, A.; Cordisco, L.; Amaretti, A.; Zanoni, S.; Matteuzzi, D.; Rossi, M. Folate production by bifidobacteria as a potential probiotic property. Appl. Environ. Microbiol. 2007, 73, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Pompei, A.; Cordisco, L.; Amaretti, A.; Zanoni, S.; Raimondi, S.; Matteuzzi, D.; Rossi, M. Administration of folate-producing bifidobacteria enhances folate status in Wistar rats. J. Nutr. 2007, 137, 2742–2746. [Google Scholar] [CrossRef]
- Sheridan, P.O.; Bindels, L.B.; Saulnier, D.M.; Reid, G.; Nova, E.; Holmgren, K.; O’Toole, P.W.; Bunn, J.; Delzenne, N.; Scott, K.P. Can prebiotics and probiotics improve therapeutic outcomes for undernourished individuals? Gut Microbes 2014, 5, 74–82. [Google Scholar] [CrossRef] [Green Version]
- Bering, S.; Suchdev, S.; Sjøltov, L.; Berggren, A.; Tetens, I.; Bukhave, K. A lactic acid-fermented oat gruel increases non-haem iron absorption from a phytate-rich meal in healthy women of childbearing age. Br. J. Nutr. 2006, 96, 80–85. [Google Scholar] [CrossRef] [Green Version]
- Jones, M.L.; Martoni, C.J.; Prakash, S. Oral supplementation with probiotic L. reuteri NCIMB 30242 increases mean circulating 25-hydroxyvitamin D: A post hoc analysis of a randomized controlled trial. J. Clin. Endocrinol. Metab. 2013, 98, 2944–2951. [Google Scholar] [CrossRef] [Green Version]
- Hollander, D.; Muralidhara, K.; Zimmerman, A. Vitamin D-3 intestinal absorption in vivo: Influence of fatty acids, bile salts, and perfusate pH on absorption. Gut 1978, 19, 267–272. [Google Scholar] [CrossRef] [Green Version]
- Narva, M.; Nevala, R.; Poussa, T.; Korpela, R. The effect of Lactobacillus helveticus fermented milk on acute changes in calcium metabolism in postmenopausal women. Eur. J. Nutr. 2004, 43, 61–68. [Google Scholar] [CrossRef]
- Hemarajata, P.; Versalovic, J. Effects of probiotics on gut microbiota: Mechanisms of intestinal immunomodulation and neuromodulation. Ther. Adv. Gastroenterol. 2013, 6, 39–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campana, R.; van Hemert, S.; Baffone, W. Strain-specific probiotic properties of lactic acid bacteria and their interference with human intestinal pathogens invasion. Gut Pathog. 2017, 9, 12. [Google Scholar] [CrossRef] [Green Version]
- Capozzi, V.; Russo, P.; Dueñas, M.T.; López, P.; Spano, G. Lactic acid bacteria producing B-group vitamins: A great potential for functional cereals products. Appl. Microbiol. Biotechnol. 2012, 96, 1383–1394. [Google Scholar] [CrossRef]
- Crittenden, R.; Martinez, N.; Playne, M. Synthesis and utilisation of folate by yoghurt starter cultures and probiotic bacteria. Int. J. Food Microbiol. 2003, 80, 217–222. [Google Scholar] [CrossRef]
- LeBlanc, J.; Laiño, J.E.; del Valle, M.J.; Vannini, V.; van Sinderen, D.; Taranto, M.P.; de Valdez, G.F.; de Giori, G.S.; Sesma, F. B-Group vitamin production by lactic acid bacteria–current knowledge and potential applications. J. Appl. Microbiol. 2011, 111, 1297–1309. [Google Scholar] [CrossRef]
- LeBlanc, J.G.; Chain, F.; Martín, R.; Bermúdez-Humarán, L.G.; Courau, S.; Langella, P. Beneficial effects on host energy metabolism of short-chain fatty acids and vitamins produced by commensal and probiotic bacteria. Microb. Cell Factories 2017, 16, 79. [Google Scholar] [CrossRef] [Green Version]
- Rossi, M.; Amaretti, A.; Raimondi, S. Folate Production by Probiotic Bacteria. Nutrients 2011, 3, 118–134. [Google Scholar] [CrossRef] [Green Version]
- Hoppe, M.; Önning, G.; Hulthén, L. Freeze-dried Lactobacillus plantarum 299v increases iron absorption in young females—Double isotope sequential single-blind studies in menstruating women. PLoS ONE 2017, 12, e0189141. [Google Scholar] [CrossRef]
- Scheers, N.; Rossander-Hulthen, L.; Torsdottir, I.; Sandberg, A.-S. Increased iron bioavailability from lactic-fermented vegetables is likely an effect of promoting the formation of ferric iron (Fe3+). Eur. J. Nutr. 2016, 55, 373–382. [Google Scholar] [CrossRef] [Green Version]
- Skrypnik, K.; Bogdanski, P.; Schmidt, M.; Suliburska, J. The Effect of Multispecies Probiotic Supplementation on Iron Status in Rats. Biol. Trace Elem. Res. 2019, 192, 234–243. [Google Scholar] [CrossRef] [Green Version]
- Kelleher, S.L.; Casas, I.; Carbajal, N.; Lonnerdal, B. Supplementation of infant formula with the probiotic lactobacillus reuteri and zinc: Impact on enteric infection and nutrition in infant rhesus monkeys. J. Pediatr. Gastroenterol. Nutr. 2002, 35, 162–168. [Google Scholar] [CrossRef]
- Silva, M.R.; Dias, G.; Ferreira, C.L.; Franceschini, S.C.; Costa, N.M. Growth of preschool children was improved when fed an iron-fortified fermented milk beverage supplemented with Lactobacillus acidophilus. Nutr. Res. 2008, 28, 226–232. [Google Scholar] [CrossRef]
- Simeoni, M.; Citraro, M.L.; Cerantonio, A.; Deodato, F.; Provenzano, M.; Cianfrone, P.; Capria, M.; Corrado, S.; Libri, E.; Comi, A. An open-label, randomized, placebo-controlled study on the effectiveness of a novel probiotics administration protocol (ProbiotiCKD) in patients with mild renal insufficiency (stage 3a of CKD). Eur. J. Nutr. 2019, 58, 2145–2156. [Google Scholar] [CrossRef] [Green Version]
- Mokhtari, Z.; Karbaschian, Z.; Pazouki, A.; Kabir, A.; Hedayati, M.; Mirmiran, P.; Hekmatdoost, A. The Effects of Probiotic Supplements on Blood Markers of Endotoxin and Lipid Peroxidation in Patients Undergoing Gastric Bypass Surgery; a Randomized, Double-Blind, Placebo-Controlled, Clinical Trial with 13 Months Follow-Up. Obes. Surg. 2019, 29, 1248–1258. [Google Scholar] [CrossRef]
- Woodard, G.A.; Encarnacion, B.; Downey, J.R.; Peraza, J.; Chong, K.; Hernandez-Boussard, T.; Morton, J.M. Probiotics improve outcomes after Roux-en-Y gastric bypass surgery: A prospective randomized trial. J. Gastrointest. Surg. 2009, 13, 1198–1204. [Google Scholar] [CrossRef]
- Valentini, L.; Pinto, A.; Bourdel-Marchasson, I.; Ostan, R.; Brigidi, P.; Turroni, S.; Hrelia, S.; Hrelia, P.; Bereswill, S.; Fischer, A. Impact of personalized diet and probiotic supplementation on inflammation, nutritional parameters and intestinal microbiota–The “RISTOMED project”: Randomized controlled trial in healthy older people. Clin. Nutr. 2015, 34, 593–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capcarova, M.; Weiss, J.; Hrncar, C.; Kolesarova, A.; Pal, G. Effect of Lactobacillus fermentum and Enterococcus faecium strains on internal milieu, antioxidant status and body weight of broiler chickens. J. Anim. Physiol. Anim. Nutr. 2010, 94, e215–e224. [Google Scholar] [CrossRef] [PubMed]
- Capcarova, M.; Hascik, P.; Kolesarova, A.; Kacaniova, M.; Mihok, M.; Pal, G. The effect of selected microbial strains on internal milieu of broiler chickens after peroral administration. Vet. Sci. Res. J. 2011, 91, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Asemi, Z.; Bahmani, S.; Shakeri, H.; Jamal, A.; Faraji, A.-M. Effect of multispecies probiotic supplements on serum minerals, liver enzymes and blood pressure in patients with type 2 diabetes. Int. J. Diabetes Dev. Ctries. 2015, 35, 90–95. [Google Scholar] [CrossRef]
- PRISMA. Available online: http://www.prisma-statement.org/Protocols/ (accessed on 19 March 2020).
- Haddaway, N.R.; Collins, A.M.; Coughlin, D.; Kirk, S. The role of Google Scholar in evidence reviews and its applicability to grey literature searching. PLoS ONE 2015, 10, e0138237. [Google Scholar] [CrossRef] [Green Version]
- Higgins, J.P.; Thomas, J.; Chandler, J.; Cumpston, M.; Li, T.; Page, M.J.; Welch, V.A. Cochrane Handbook for Systematic Reviews of Interventions, 2nd ed.; John Wiley & Sons: Chichester, UK, 2019. [Google Scholar]
- Sterne, J.A.; Savović, J.; Page, M.J.; Elbers, R.G.; Blencowe, N.S.; Boutron, I.; Cates, C.J.; Cheng, H.-Y.; Corbett, M.S.; Eldridge, S.M. RoB 2: A revised tool for assessing risk of bias in randomised trials. BMJ 2019, 366, l4898. [Google Scholar] [CrossRef] [Green Version]
- Ding, H.; Hu, G.L.; Zheng, X.Y.; Chen, Q.; Threapleton, D.E.; Zhou, Z.H. The method quality of cross-over studies involved in Cochrane Systematic Reviews. PLoS ONE 2015, 10, e0120519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sterne, J.A.; Hernán, M.A.; Reeves, B.C.; Savović, J.; Berkman, N.D.; Viswanathan, M.; Henry, D.; Altman, D.G.; Ansari, M.T.; Boutron, I. ROBINS-I: A tool for assessing risk of bias in non-randomised studies of interventions. BMJ 2016, 355, i4919. [Google Scholar] [CrossRef] [Green Version]
- Elmadfa, I.; Heinzle, C.; Majchrzak, D.; Foissy, H. Influence of a probiotic yoghurt on the status of vitamins B1, B2 and B6 in the healthy adult human. Ann. Nutr. Metab. 2001, 45, 13–18. [Google Scholar] [CrossRef]
- Fabian; Elmadfa. The effect of daily consumption of probiotic and conventional yoghurt on oxidant and antioxidant parameters in plasma of young healthy women. Int. J. Vitam. Nutr. Res. 2007, 77, 79–88. [Google Scholar] [CrossRef]
- Fabian, E.; Majchrzak, D.; Dieminger, B.; Meyer, E.; Elmadfa, I. Influence of probiotic and conventional yoghurt on the status of vitamins B1, B2 and B6 in young healthy women. Ann. Nutr. Metab. 2008, 52, 29–36. [Google Scholar] [CrossRef]
- Surono, I.S.; Martono, P.D.; Kameo, S.; Suradji, E.W.; Koyama, H. Effect of probiotic L. plantarum IS-10506 and zinc supplementation on humoral immune response and zinc status of Indonesian pre-school children. Trace Elem. Med. Biol. 2014, 28, 465–469. [Google Scholar] [CrossRef]
- Agustina, R.; Bovee-Oudenhoven, I.M.; Lukito, W.; Fahmida, U.; Van De Rest, O.; Zimmermann, M.B.; Firmansyah, A.; Wulanti, R.; Albers, R.; van den Heuvel, E.G.; et al. Probiotics Lactobacillus reuteri DSM 17938 and Lactobacillus casei CRL 431 modestly increase growth, but not iron and zinc status, among Indonesian children aged 1–6 years. J. Nutr. 2013, 143, 1184–1193. [Google Scholar] [CrossRef] [Green Version]
- Donaldson, M.S. Metabolic vitamin B12 status on a mostly raw vegan diet with follow-up using tablets, nutritional yeast, or probiotic supplements. Ann. Nutr. Metab. 2000, 44, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Gohel, M.K.; Prajapati, J.B.; Mudgal, S.V.; Pandya, H.V.; Singh, U.S.; Trivedi, S.S.; Phatak, A.G.; Patel, R.M. Effect of probiotic dietary intervention on calcium and haematological parameters in geriatrics. JCDR 2016, 10, LC05. [Google Scholar] [CrossRef] [PubMed]
- Axling, U.; Önning, G.; Combs, M.A.; Bogale, A.; Högström, M.; Svensson, M. The Effect of Lactobacillus plantarum 299v on Iron Status and Physical Performance in Female Iron-Deficient Athletes: A Randomized Controlled Trial. Nutrients 2020, 12, 1279. [Google Scholar] [CrossRef] [PubMed]
- Ballini, A.; Gnoni, A.; De Vito, D.; Dipalma, G.; Cantore, S.; Gargiulo Isacco, C.; Saini, R.; Santacroce, L.; Topi, S.; Scarano, A.; et al. Effect of probiotics on the occurrence of nutrition absorption capacities in healthy children: A randomized double-blinded placebo-controlled pilot study. Eur. Rev. Med. Pharm. Sci. 2019, 23, 8645–8657. [Google Scholar]
- Mohammad, M.A.; Molloy, A.; Scott, J.; Hussein, L. Plasma cobalamin and folate and their metabolic markers methylmalonic acid and total homocysteine among Egyptian children before and after nutritional supplementation with the probiotic bacteria Lactobacillus acidophilus in yoghurt matrix. Int. J. Food Sci. Nutr. 2006, 57, 470–480. [Google Scholar] [CrossRef]
- Korčok, D.J.; Tršić-Milanoviće, N.; Ivanović, N.; Đorđević, B. Development of probiotic formulation for the treatment of iron deficiency anemia. Chem. Pharm. Bull. 2018, 66, 347–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swanson, K.S.; Gibson, G.R.; Hutkins, R.; Reimer, R.A.; Reid, G.; Verbeke, K.; Scott, K.P.; Holscher, H.D.; Azad, M.B.; Delzenne, N.M. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of synbiotics. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 687–701. [Google Scholar] [CrossRef]
- Wibowo, N.; Bardosono, S.; Irwinda, R. Effects of Bifidobacterium animalis lactis HN019 (DR10TM), inulin, and micronutrient fortified milk on faecal DR10TM, immune markers, and maternal micronutrients among Indonesian pregnant women. Asia Pac. J. Clin. Nutr. 2016, 25, S102–s110. [Google Scholar] [PubMed]
- Karbaschian, Z.; Mokhtari, Z.; Pazouki, A.; Kabir, A.; Hedayati, M.; Moghadam, S.S.; Mirmiran, P.; Hekmatdoost, A. Probiotic Supplementation in Morbid Obese Patients Undergoing One Anastomosis Gastric Bypass-Mini Gastric Bypass (OAGB-MGB) Surgery: A Randomized, Double-Blind, Placebo-Controlled, Clinical Trial. Obes. Surg. 2018, 28, 2874–2885. [Google Scholar] [CrossRef] [PubMed]
- Yuki, N.; Watanabe, K.; Mike, A.; Tagami, Y.; Tanaka, R.; Ohwaki, M.; Morotomi, M. Survival of a probiotic, Lactobacillus casei strain Shirota, in the gastrointestinal tract: Selective isolation from faeces and identification using monoclonal antibodies. Int. J. Food Microbiol. 1999, 48, 51–57. [Google Scholar] [CrossRef]
- Magnúsdóttir, S.; Ravcheev, D.; de Crécy-Lagard, V.; Thiele, I. Systematic genome assessment of B-vitamin biosynthesis suggests co-operation among gut microbes. Front. Genet. 2015, 6, 148. [Google Scholar] [CrossRef] [Green Version]
- Cárdenas, N.; Laiño, J.E.; Delgado, S.; Jiménez, E.; Del Valle, M.J.; De Giori, G.S.; Sesma, F.; Mayo, B.; Fernández, L.; LeBlanc, J.G. Relationships between the genome and some phenotypical properties of Lactobacillus fermentum CECT 5716, a probiotic strain isolated from human milk. Appl. Microbiol. Biotechnol. 2015, 99, 4343–4353. [Google Scholar] [CrossRef]
- Hannibal, L.; Lysne, V.; Bjørke-Monsen, A.-L.; Behringer, S.; Grünert, S.C.; Spiekerkoetter, U.; Jacobsen, D.W.; Blom, H.J. Biomarkers and algorithms for the diagnosis of vitamin B12 deficiency. Front. Mol. Biosci. 2016, 3, 27. [Google Scholar] [CrossRef] [Green Version]
- de Benoist, B. Conclusions of a WHO Technical Consultation on folate and vitamin B12 deficiencies. Food Nutr Bull. 2008, 29, S238–S244. [Google Scholar] [CrossRef]
- Lin, M.; Young, C. Folate levels in cultures of lactic acid bacteria. Int. Dairy J. 2000, 10, 409–413. [Google Scholar] [CrossRef]
- Nor, N.M.; Mohamad, R.; Foo, H.L.; Rahim, R.A. Improvement of folate biosynthesis by lactic acid bacteria using response surface methodology. Food Technol. Biotechnol. 2010, 48, 243–250. [Google Scholar]
- Deguchi, Y.; Morishita, T.; Mutai, M. Comparative studies on synthesis of water-soluble vitamins among human species of bifidobacteria. Agric. Biol. Chem. 1985, 49, 13–19. [Google Scholar]
- Strozzi, G.P.; Mogna, L. Quantification of folic acid in human feces after administration of Bifidobacterium probiotic strains. J. Clin. Gastroenterol. 2008, 42, S179–S184. [Google Scholar] [CrossRef]
- D’Aimmo, M.R.; Mattarelli, P.; Biavati, B.; Carlsson, N.-G.; Andlid, T. The potential of bifidobacteria as a source of natural folate. J. Appl. Microbiol. 2012, 112, 975–984. [Google Scholar] [CrossRef]
- Eloe-Fadrosh, E.A.; Brady, A.; Crabtree, J.; Drabek, E.F.; Ma, B.; Mahurkar, A.; Ravel, J.; Haverkamp, M.; Fiorino, A.-M.; Botelho, C. Functional dynamics of the gut microbiome in elderly people during probiotic consumption. MBio 2015, 6, e00231-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez, B.; Delgado, S.; Blanco-Míguez, A.; Lourenço, A.; Gueimonde, M.; Margolles, A. Probiotics, gut microbiota, and their influence on host health and disease. Mol. Nutr. Food Res. 2017, 61, 1600240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lahtinen, S.J.; Tammela, L.; Korpela, J.; Parhiala, R.; Ahokoski, H.; Mykkänen, H.; Salminen, S.J. Probiotics modulate the Bifidobacterium microbiota of elderly nursing home residents. Age 2009, 31, 59–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madhu, A.N.; Giribhattanavar, P.; Narayan, M.S.; Prapulla, S.G. Probiotic lactic acid bacterium from kanjika as a potential source of vitamin B 12: Evidence from LC-MS, immunological and microbiological techniques. Biotechnol. Lett. 2010, 32, 503–506. [Google Scholar] [CrossRef]
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Role of the gut microbiota in nutrition and health. BMJ. 2018, 361, k2179. [Google Scholar] [CrossRef] [Green Version]
- Asemi, Z.; Esmaillzadeh, A. Effect of daily consumption of probiotic yoghurt on serum levels of calcium, iron and liver enzymes in pregnant women. Int. J. Prev. Med. 2013, 4, 949. [Google Scholar]
- Trinidad, T.P.; Wolever, T.; Thompson, L.U. Effect of acetate and propionate on calcium absorption from the rectum and distal colon of humans. Am. J. Clin. Nutr. 1996, 63, 574–578. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.-C.; Wu, J.-B.; Lu, T.-J.; Lin, W.-C. The prebiotic effect of Anoectochilus formosanus and its consequences on bone health. Br. J. Nutr. 2013, 109, 1779–1788. [Google Scholar] [CrossRef] [Green Version]
- Cheung, A.L.T.F.; Wilcox, G.; Walker, K.Z.; Shah, N.P.; Strauss, B.; Ashton, J.F.; Stojanovska, L. Fermentation of calcium-fortified soya milk does not appear to enhance acute calcium absorption in osteopenic post-menopausal women. Br. J. Nutr. 2011, 105, 282–286. [Google Scholar] [CrossRef] [Green Version]
- Sanggaard, K.; Holst, J.; Rehfeld, J.; Sandström, B.; Raben, A.; Tholstrup, T. Different effects of whole milk and a fermented milk with the same fat and lactose content on gastric emptying and postprandial lipaemia, but not on glycaemic response and appetite. Br. J. Nutr. 2004, 92, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Mahé, S.; Marteau, P.; Huneau, J.-F.; Thuillier, F.; Tomé, D. Intestinal nitrogen and electrolyte movements following fermented milk ingestion in man. Br. J. Nutr. 1994, 71, 169–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.S.; Noguchi, T.; Naito, H. Phosphopeptides and soluble calcium in the small intestine of rats given a casein diet. Br. J. Nutr. 1980, 43, 457–467. [Google Scholar] [CrossRef] [Green Version]
- Kitts, D.D.; Yuan, Y.V.; Nagasawa, T.; Moriyama, Y. Effect of casein, casein phosphopeptides and calcium intake on ileal 45 Ca disappearance and temporal systolic blood pressure in spontaneously hypertensive rats. Br. J. Nutr. 1992, 68, 765–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, M.; Sandström, B.; Jensen, M.; Sørensen, S.S. Casein phosphopeptides improve zinc and calcium absorption from rice-based but not from whole-grain infant cereal. J. Pediatr. Gastroenterol. Nutr. 1997, 24, 56–62. [Google Scholar] [CrossRef]
- Hansen, M.; Sandström, B.; Jensen, M.; Sørensen, S. Effect of casein phosphopeptides on zinc and calcium absorption from bread meals. J. Trace Elem. Med. Biol. 1997, 11, 143–149. [Google Scholar] [CrossRef]
- Heaney, R.P.; Saito, Y.; Orimo, H. Effect of caseinphosphopeptide on absorbability of co-ingested calcium in normal postmenopausal women. J. Bone Miner. Metab. 1994, 12, 77–81. [Google Scholar] [CrossRef]
- Bomba, A.; Nemcová, R.r.; Mudroňová, D.; Guba, P. The possibilities of potentiating the efficacy of probiotics. Trends Food Sci. Technol. 2002, 13, 121–126. [Google Scholar] [CrossRef]
- Nemcova, R.; Bomba, A.; Gancarcikova, S.; Herich, R.; Guba, P. Study of the effect of Lactobacillus paracasei and fructooligosaccharides on the faecal microflora in weanling piglets. Berl. Munch. Tierarztl. Wochenschr. 1999, 112, 225–228. [Google Scholar]
- Nakashima, A. Stimulatory effect of phytin on acid production by Lactobacillus casei. J. Nutr. Sci. Vitaminol. 1997, 43, 419–424. [Google Scholar] [CrossRef] [Green Version]
- Bomba, A.; Nemcová, R.; Gancarčíková, S.; Herich, R.; Kaštel, R. Potentiation of the Effectiveness of Lactobacillus Casei in the Prevention of E. Coli Induced Diarrhea in Conventional and Gnotobiotic Pigs. In Mechanisms in the Pathogenesis of Enteric Diseases 2; Paul, P.S., Francis, D.H., Eds.; Springer: Boston, MA, USA, 1999; pp. 185–190. [Google Scholar]
- Holm, A.; Poulsen, H. Zinc oxide in treating E. coli diarrhea in pigs after weaning. Compend. Contin. Educ. Pract. Vet. 1996, 18, s26–s29. [Google Scholar]
- Tesán, F.; Hernández, F.; Torti, H.; Massot, F.; Huarte, M.; de Celis, E.R.; Barreiro, M.; Weill, R.; Cremaschi, G.; Boccio, J. Glycine-stabilized zinc gluconate has similar bioavailability than zinc sulfate in a zinc fortified probiotic food. Open Nutraceuticals J. 2011, 4, 136–143. [Google Scholar] [CrossRef] [Green Version]
- Shah, M.; Zaneb, H.; Masood, S.; Khan, R.U.; Ashraf, S.; Sikandar, A.; Rehman, H.F.U.; Rehman, H.U. Effect of dietary supplementation of zinc and multi-microbe probiotic on growth traits and alteration of intestinal architecture in broiler. Probiotics Antimicrob. Proteins 2019, 11, 931–937. [Google Scholar] [CrossRef] [PubMed]
- Gill, H.; Prasad, J. Probiotics, immunomodulation, and health benefits. In Bioactive Components of Milk; Springer: New York, NY, USA, 2008; pp. 423–454. [Google Scholar]
- Vonderheid, S.C.; Tussing-Humphreys, L.; Park, C.; Pauls, H.; OjiNjideka Hemphill, N.; LaBomascus, B.; McLeod, A.; Koenig, M.D. A Systematic Review and Meta-Analysis on the Effects of Probiotic Species on Iron Absorption and Iron Status. Nutrients 2019, 11, 2938. [Google Scholar] [CrossRef] [Green Version]
- Johansson, M.; Molin, G.; Jeppsson, B.; Nobaek, S.; Ahrne, S.; Bengmark, S. Administration of different Lactobacillus strains in fermented oatmeal soup: In vivo colonization of human intestinal mucosa and effect on the indigenous flora. Appl. Env. Microbiol. 1993, 59, 15–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derman, D.; Bothwell, T.; Torrance, J.; Bezwoda, W.; MacPhail, A.; Kew, M.; Sayers, M.; Disler, P.; Charlton, R. Iron absorption from maize (Zea mays) and sorghum (Sorghum vulgare) beer. Br. J. Nutr. 1980, 43, 271–279. [Google Scholar] [CrossRef] [Green Version]
- Liljeberg, H.; Björck, I. Delayed gastric emptying rate may explain improved glycaemia in healthy subjects to a starchy meal with added vinegar. Eur. J. Clin. Nutr. 1998, 52, 368–371. [Google Scholar] [CrossRef] [Green Version]
- Salovaara, S.; Larsson Alminger, M.; Eklund-Jonsson, C.; Andlid, T.; Sandberg, A.-S. Prolonged transit time through the stomach and small intestine improves iron dialyzability and uptake in vitro. J. Agric. Food Chem. 2003, 51, 5131–5136. [Google Scholar] [CrossRef]
- Salovaara, S.; Sandberg, A.-S.; Andlid, T. Organic acids influence iron uptake in the human epithelial cell line Caco-2. J. Agric. Food Chem. 2002, 50, 6233–6238. [Google Scholar] [CrossRef]
- Sandberg, A.-S.; Önning, G.; Engström, N.; Scheers, N. Iron Supplements Containing Lactobacillus plantarum 299v Increase Ferric Iron and Up-regulate the Ferric Reductase DCYTB in Human Caco-2/HT29 MTX Co-Cultures. Nutrients 2018, 10, 1949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douglas, L.C.; Sanders, M.E. Probiotics and prebiotics in dietetics practice. J. Am. Diet. Assoc. 2008, 108, 510–521. [Google Scholar] [CrossRef]
- Chapman, C.; Gibson, G.R.; Rowland, I. Health benefits of probiotics: Are mixtures more effective than single strains? Eur. J. Nutr. 2011, 50, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Spinler, J.K.; Taweechotipatr, M.; Rognerud, C.L.; Ou, C.N.; Tumwasorn, S.; Versalovic, J. Human-derived probiotic Lactobacillus reuteri demonstrate antimicrobial activities targeting diverse enteric bacterial pathogens. Anaerobe 2008, 14, 166–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Shea, E.F.; Cotter, P.D.; Stanton, C.; Ross, R.P.; Hill, C. Production of bioactive substances by intestinal bacteria as a basis for explaining probiotic mechanisms: Bacteriocins and conjugated linoleic acid. Int. J. Food Microbiol. 2012, 152, 189–205. [Google Scholar] [CrossRef]
- Collado, M.; Meriluoto, J.; Salminen, S. Role of commercial probiotic strains against human pathogen adhesion to intestinal mucus. Lett. Appl. Microbiol. 2007, 45, 454–460. [Google Scholar] [CrossRef]
- Lee, B.J.; Bak, Y.-T. Irritable bowel syndrome, gut microbiota and probiotics. J. Neurogastroenterol. Motil. 2011, 17, 252. [Google Scholar] [CrossRef] [Green Version]
- Arthur, J.C.; Gharaibeh, R.Z.; Uronis, J.M.; Perez-Chanona, E.; Sha, W.; Tomkovich, S.; Mühlbauer, M.; Fodor, A.A.; Jobin, C. VSL# 3 probiotic modifies mucosal microbial composition but does not reduce colitis-associated colorectal cancer. Sci. Rep. 2013, 3, 2868. [Google Scholar]
- Sugahara, H.; Odamaki, T.; Fukuda, S.; Kato, T.; Xiao, J.-z.; Abe, F.; Kikuchi, J.; Ohno, H. Probiotic Bifidobacterium longum alters gut luminal metabolism through modification of the gut microbial community. Sci. Rep. 2015, 5, 13548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turroni, F.; Milani, C.; Duranti, S.; Mancabelli, L.; Mangifesta, M.; Viappiani, A.; Lugli, G.A.; Ferrario, C.; Gioiosa, L.; Ferrarini, A. Deciphering bifidobacterial-mediated metabolic interactions and their impact on gut microbiota by a multi-omics approach. ISME J. 2016, 10, 1656–1668. [Google Scholar] [CrossRef]
- Preidis, G.A.; Saulnier, D.M.; Blutt, S.E.; Mistretta, T.A.; Riehle, K.P.; Major, A.M.; Venable, S.F.; Finegold, M.J.; Petrosino, J.F.; Conner, M.E. Probiotics stimulate enterocyte migration and microbial diversity in the neonatal mouse intestine. FASEB J. 2012, 26, 1960–1969. [Google Scholar] [CrossRef] [Green Version]
- Nobaek, S.; Johansson, M.-L.; Molin, G.; Ahrné, S.; Jeppsson, B. Alteration of intestinal microflora is associated with reduction in abdominal bloating and pain in patients with irritable bowel syndrome. Am. J. Gastroenterol. Suppl. 2000, 95, 1231–1238. [Google Scholar] [CrossRef] [PubMed]
- Cha, B.K.; Jung, S.M.; Choi, C.H.; Song, I.-D.; Lee, H.W.; Kim, H.J.; Hyuk, J.; Chang, S.K.; Kim, K.; Chung, W.-S. The effect of a multispecies probiotic mixture on the symptoms and fecal microbiota in diarrhea-dominant irritable bowel syndrome: A randomized, double-blind, placebo-controlled trial. J. Clin. Gastroenterol. 2012, 46, 220–227. [Google Scholar] [CrossRef]
- McNulty, N.P.; Yatsunenko, T.; Hsiao, A.; Faith, J.J.; Muegge, B.D.; Goodman, A.L.; Henrissat, B.; Oozeer, R.; Cools-Portier, S.; Gobert, G. The impact of a consortium of fermented milk strains on the gut microbiome of gnotobiotic mice and monozygotic twins. Sci. Transl. Med. 2011, 3, ra106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lahtinen, S.J.; Forssten, S.; Aakko, J.; Granlund, L.; Rautonen, N.; Salminen, S.; Viitanen, M.; Ouwehand, A.C. Probiotic cheese containing Lactobacillus rhamnosus HN001 and Lactobacillus acidophilus NCFM® modifies subpopulations of fecal lactobacilli and Clostridium difficile in the elderly. Age 2012, 34, 133–143. [Google Scholar] [CrossRef] [Green Version]
- Ouwehand, A.C.; Kirjavainen, P.V.; Shortt, C.; Salminen, S. Probiotics: Mechanisms and established effects. Int. Dairy J. 1999, 9, 43–52. [Google Scholar] [CrossRef]
- Morelli, L.; Cesena, C.; Lucchini, F.; Callegari, M.L.; Alander, M.; Mattila-Sandholm, T.; von Wright, A.; Salminen, S.; Lehto, E.; Vilpponen-Salmela, T. Role of cell aggregation protein in adhesion in vitro and in vivo. In Proceedings of the 2nd Workshop Demonstation of the Nutritional Functionality of Probiotic Foods FAIR CT96-1028, Espoo, Finland, 3 October 1997; pp. 63–64. [Google Scholar]
- Surono, I.S. In vitro probiotic properties of indigenous dadih lactic acid bacteria. Asian Australas. J. Anim. Sci. 2003, 16, 726–731. [Google Scholar] [CrossRef]
- Collado, M.C.; Surono, I.S.; Meriluoto, J.; Salminen, S. Potential probiotic characteristics of Lactobacillus and Enterococcus strains isolated from traditional dadih fermented milk against pathogen intestinal colonization. J. Food Prot. 2007, 70, 700–705. [Google Scholar] [CrossRef] [PubMed]
- Khedkar, C.; Dave, J.; Sannabhadti, S. Antibacterial activity of human strains of Lactobacillus acidophilus grown in milk against selected pathogenic and spoilage type bacteria. Cult. Dairy Prod. J. 1990, 25, 29–31. [Google Scholar]
- Ashar, M.; Prajapati, J. Bile tolerance, bile deconjugation and cholesterol reducing properties of dietary lactobacilli. Indian J. Microbiol. 1998, 38, 145–148. [Google Scholar]
- Kodaikkal, V.; Prajapati, J.; Ljungh, A. Evaluation of adhesion of Lactobacillus strains to HT-29 cells by a flow cytometric assay. Int. J. Appl. Anim. Sci. 2012, 1, 1–7. [Google Scholar]
- Prajapati, J.B.; Khedkar, C.D.; Chitra, J.; Suja, S.; Mishra, V.; Sreeja, V.; Patel, R.K.; Ahir, V.B.; Bhatt, V.D.; Sajnani, M.R.; et al. Whole-Genome Shotgun Sequencing of an Indian-Origin Lactobacillus helveticus Strain, MTCC 5463, with Probiotic Potential. J. Bacteriol. Res. 2011, 193, 4282–4283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Chen, D.-C. Facing a new challenge: The adverse effects of antibiotics on gut microbiota and host immunity. Chin. Med. J. 2019, 132, 1135. [Google Scholar] [CrossRef] [PubMed]
- Jernberg, C.; Löfmark, S.; Edlund, C.; Jansson, J.K. Long-term impacts of antibiotic exposure on the human intestinal microbiota. Microbiology 2010, 156, 3216–3223. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barkhidarian, B.; Roldos, L.; Iskandar, M.M.; Saedisomeolia, A.; Kubow, S. Probiotic Supplementation and Micronutrient Status in Healthy Subjects: A Systematic Review of Clinical Trials. Nutrients 2021, 13, 3001. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13093001
Barkhidarian B, Roldos L, Iskandar MM, Saedisomeolia A, Kubow S. Probiotic Supplementation and Micronutrient Status in Healthy Subjects: A Systematic Review of Clinical Trials. Nutrients. 2021; 13(9):3001. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13093001
Chicago/Turabian StyleBarkhidarian, Bahareh, Lucas Roldos, Michèle M. Iskandar, Ahmad Saedisomeolia, and Stan Kubow. 2021. "Probiotic Supplementation and Micronutrient Status in Healthy Subjects: A Systematic Review of Clinical Trials" Nutrients 13, no. 9: 3001. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13093001