
Ghee Butter from Bovine Colostrum Reduces Inflammation in the Mouse Model of Acute Pancreatitis with Potential Involvement of Free Fatty Acid Receptors


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Induction of Acute Pancreatitis
2.3. Treatment with GBBC
2.4. FFAR1 and FFAR4 Antagonist Administration
2.5. Blood Serum and Tissue Collection
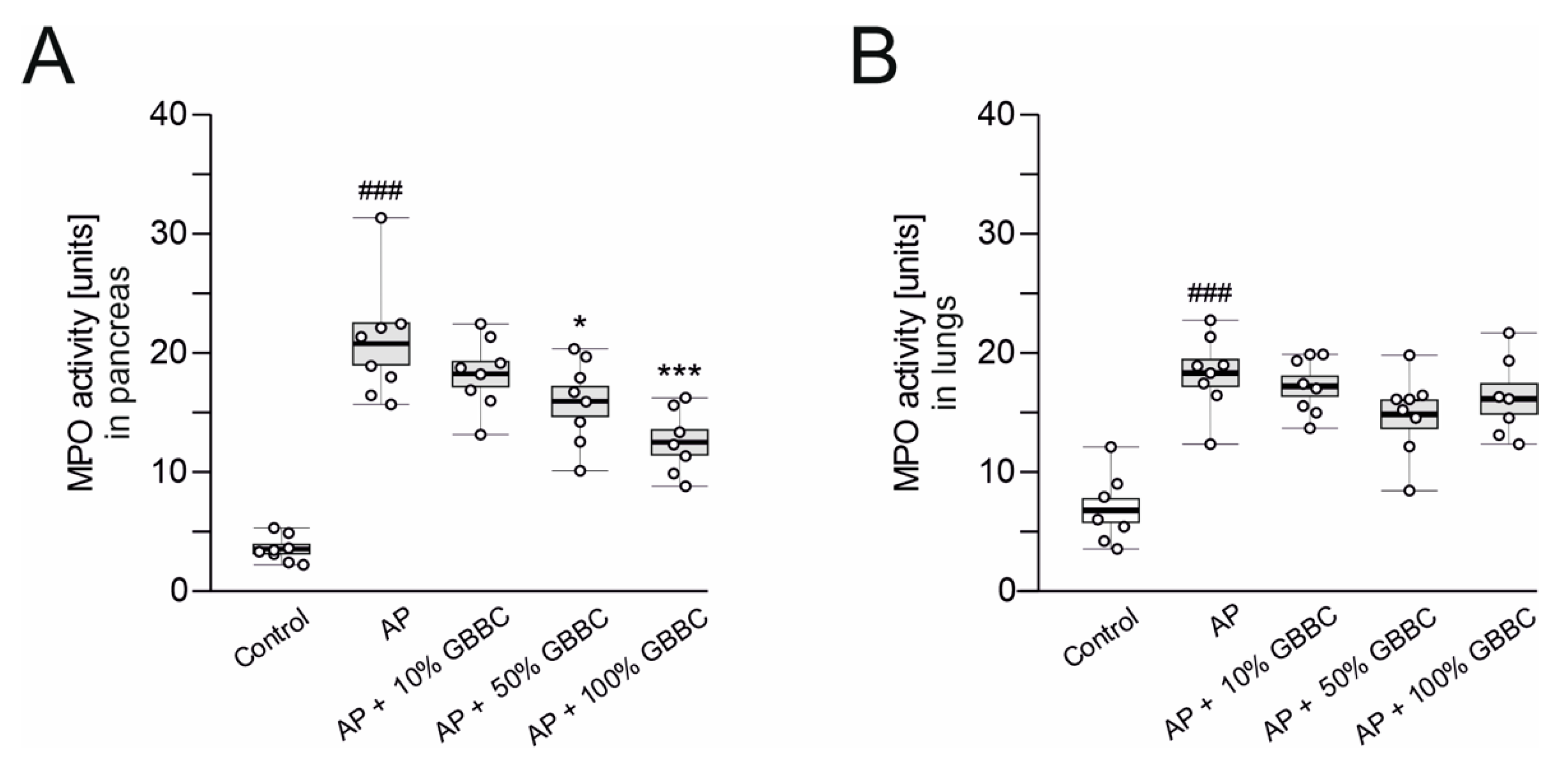
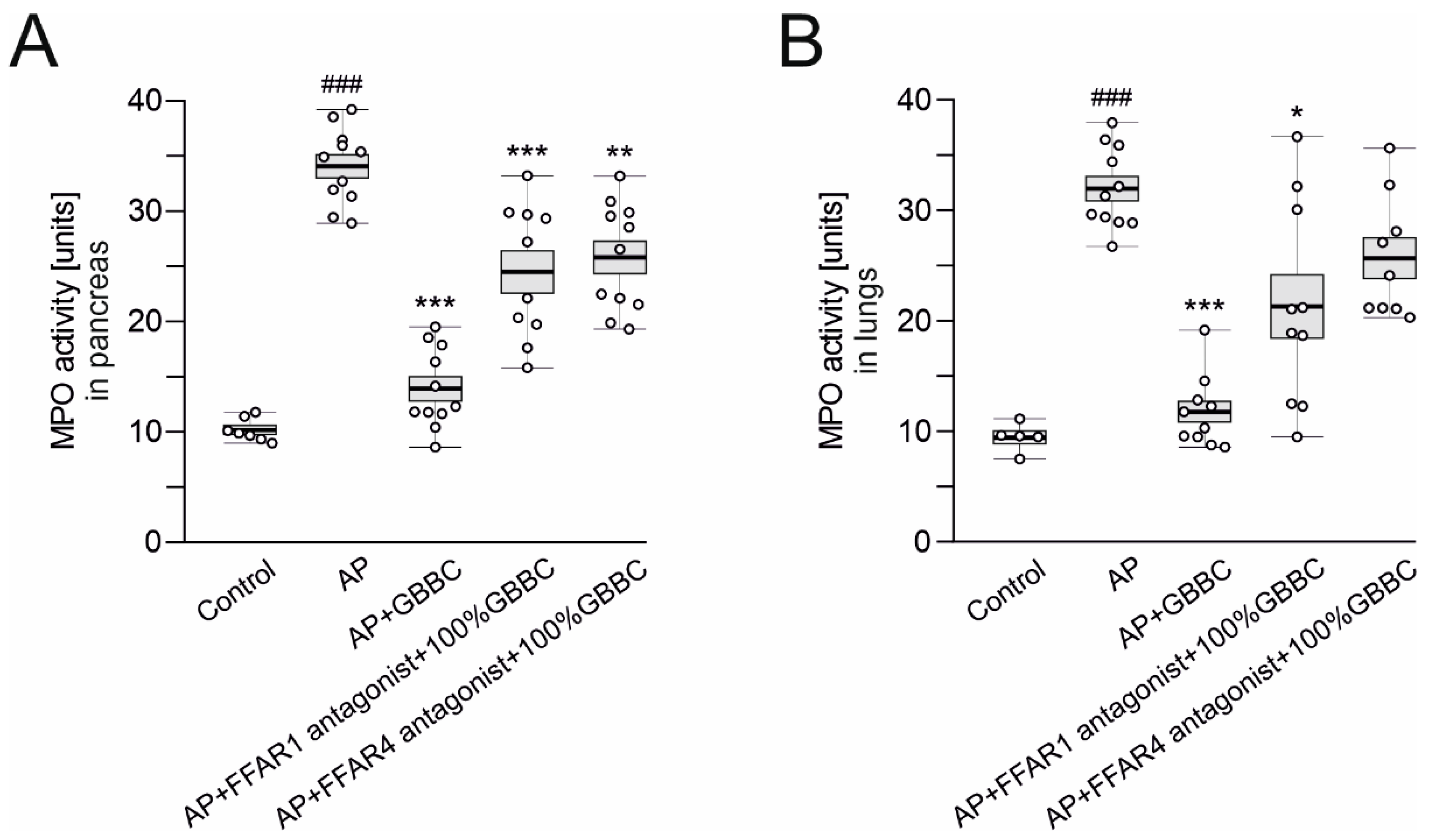
2.6. Determination of Myeloperoxidase Activity
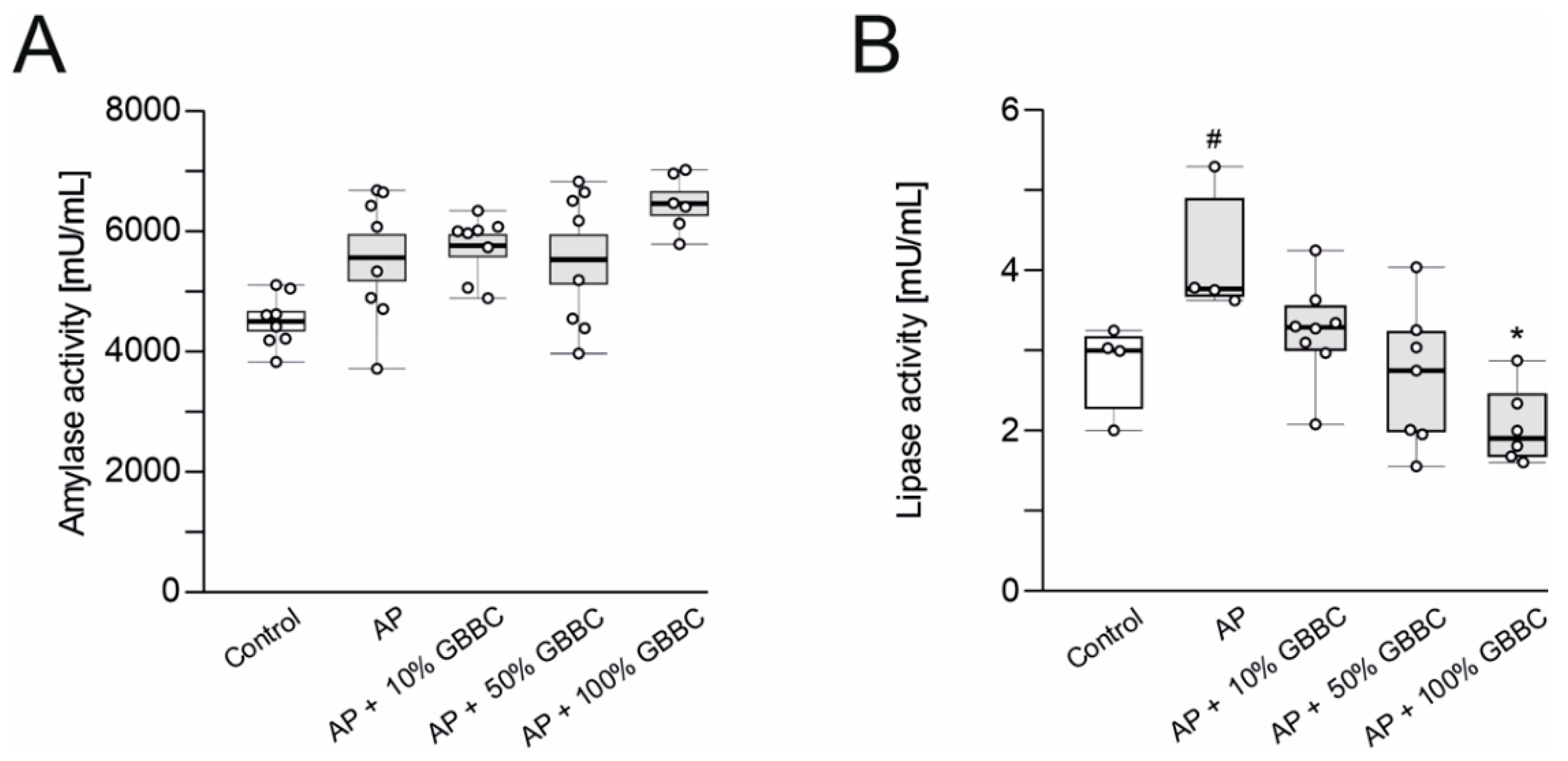
2.7. Determination of Amylase Activity in Blood Serum
- B = nitrophenol amount from the standard curve (in nmol);
- ΔT = reaction time (T2–T1) (min);
- V = pretreated sample volume added to the reaction well (in mL);
- D = sample dilution factor.
2.8. Determination of Lipase Activity in Blood Serum
- B = amount of glycerol in sample well calculated from standard curve (nmol);
- ΔT = reaction time (T2–T1) (min);
- V = original sample volume added into the reaction well (mL);
- D = sample dilution factor.
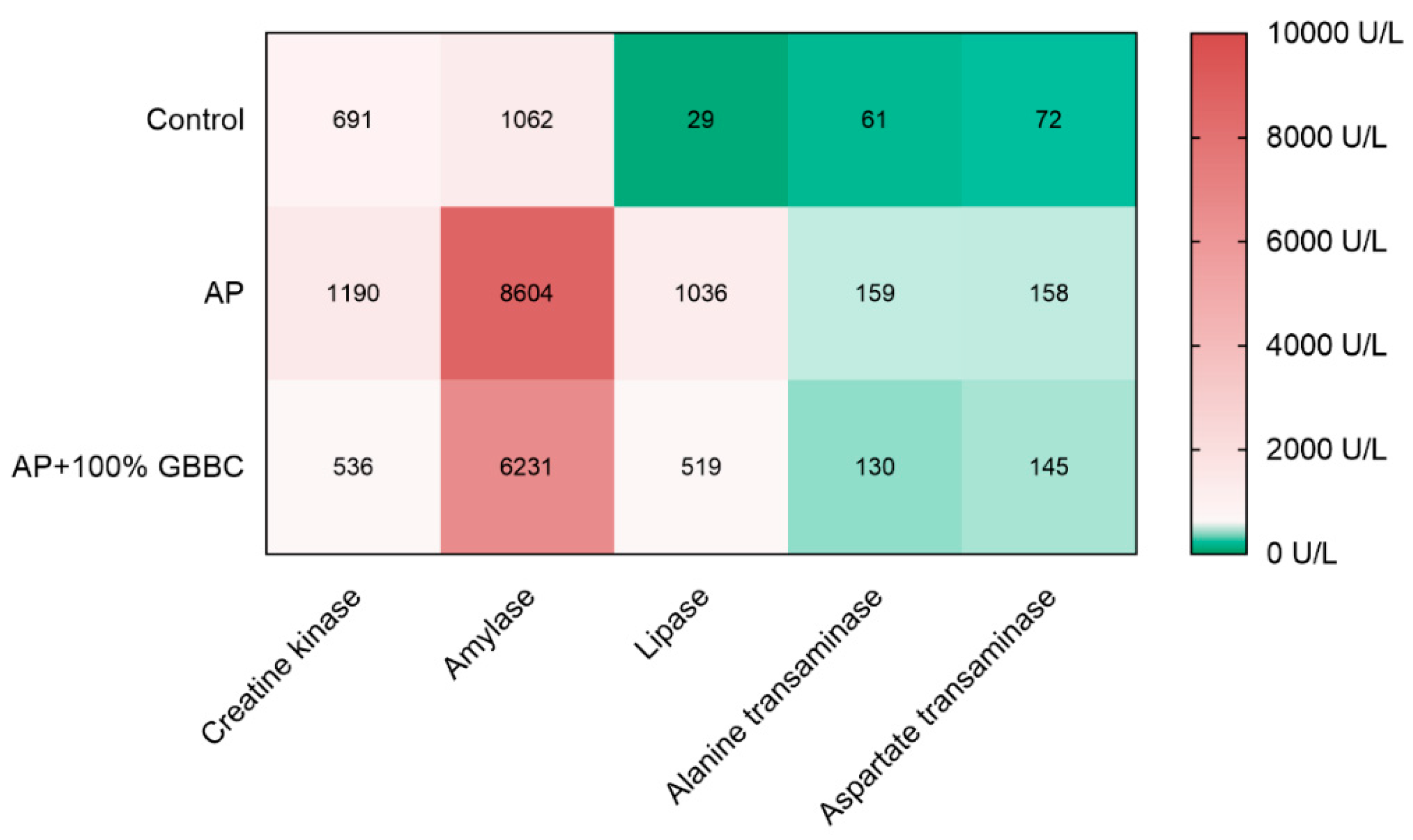
2.9. Serum Blood Tests
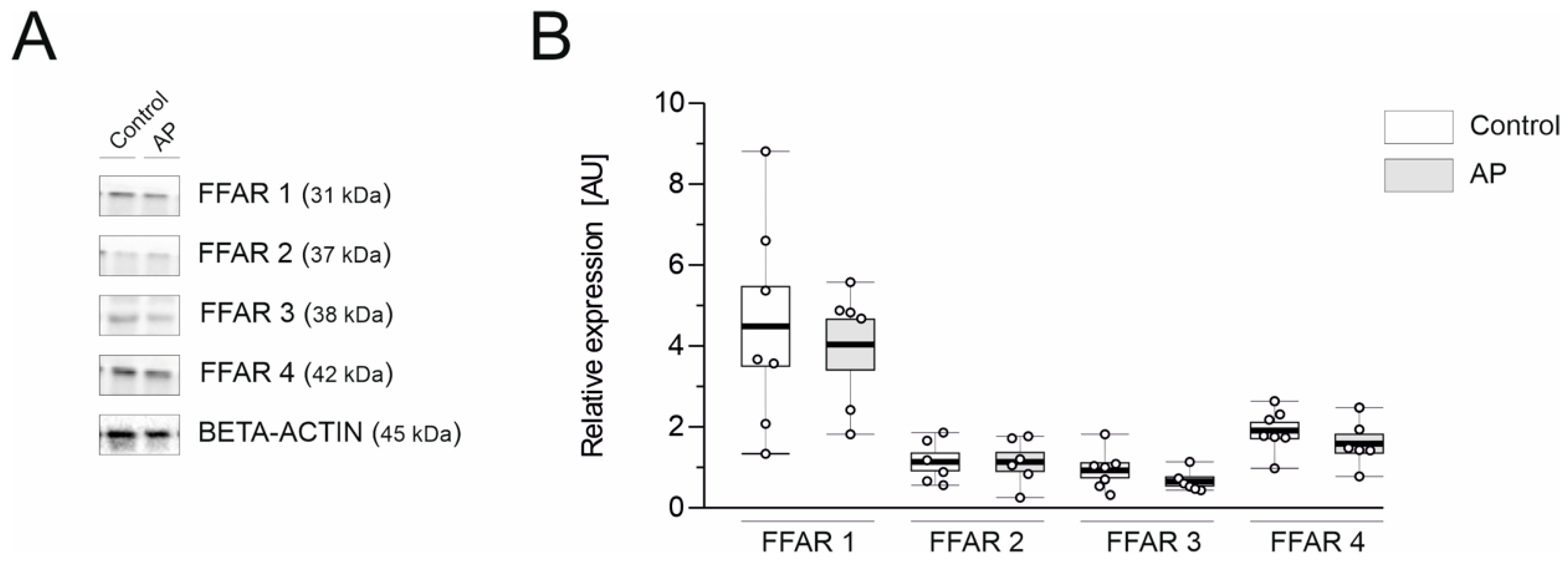
2.10. Western Blot Analysis
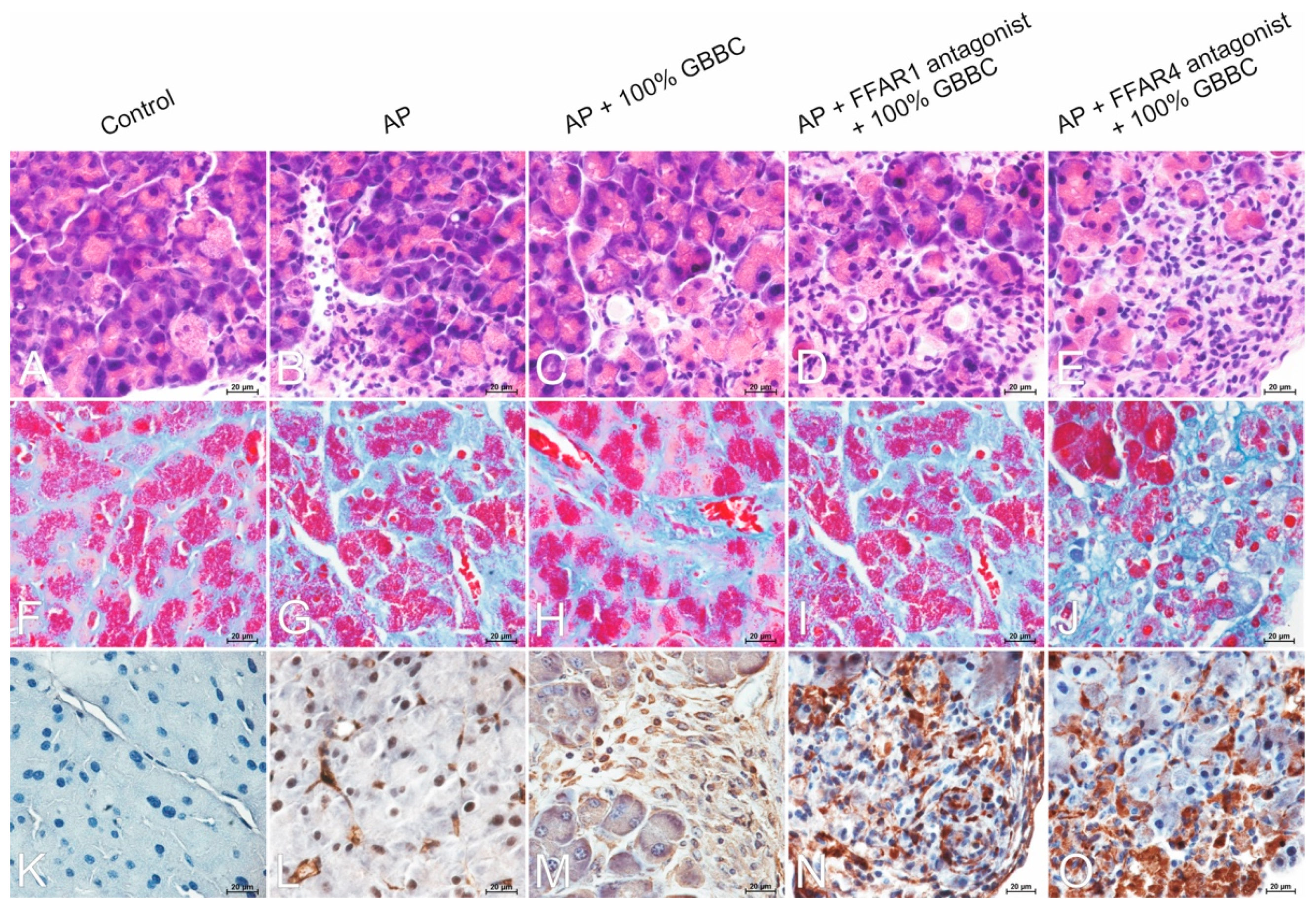
2.11. Histopathological Analysis
2.12. Drugs
2.13. Statistical Analysis
3. Results
3.1. Administration of GBBC Attenuated Pancreatitis in L-Arginine-Induced AP in Mice
3.2. Administration of GBBC Significantly Reduced Lipase Activity but Not Amylase Activity in the Blood Serum in L-Arginine-Induced AP in Mice
3.3. GBBC Normalized Various Metabolic Enzyme Activities Increased in L-Arginine-Induced AP in Mice
3.4. FFAR1 and FFAR4 Are Highly Expressed in the Mouse Pancreas
3.5. The Anti-Inflammatory Activity of GBBC in L-Arginine-Induced AP in Mice Could Depend on FFAR1 and FFAR4
3.6. Morphological Analysis Revealed Decreased Severity of Pancreatitis after GBBC Treatment in AP Mice
3.6.1. H&E Staining
3.6.2. Masson’s Trichrome Staining
3.6.3. Immunohistochemical Reaction with Anti-α-SMA
4. Discussion
5. Study Limitations
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Lankisch, P.G.; Apte, M.; Banks, P.A. Acute pancreatitis. Lancet 2015, 386, 85–96. [Google Scholar] [CrossRef]
- Hammad, A.Y.; Ditillo, M.; Castanon, L. Pancreatitis, Surg. Clin. North Am. 2018, 98, 895–913. [Google Scholar] [CrossRef]
- Moreira, M.; Matias, J.E.; Souza, C.J.; Nicoluzzi, J.E.; Caron, P.E.; Repka, J.C. Action of tacrolimus in arginine induced experimental acute pancreatitis. Rev. Col. Bras. Cir. 2011, 38, 260–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirkegård, J.; Cronin-Fenton, D.; Heide-Jørgensen, U.; Mortensen, F.V. Acute Pancreatitis and Pancreatic Cancer Risk: A Nationwide Matched-Cohort Study in Denmark. Gastroenterology 2018, 154, 1729–1736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geiger, A.J. Colostrum: Back to basics with immunoglobulins. J. Anim. Sci. 2020, 98, S126–S132. [Google Scholar] [CrossRef] [PubMed]
- Menchetti, L.; Traina, G.; Tomasello, G.; Casagrande-Proietti, P.; Leonardi, L.; Barbato, O.; Brecchia, G. Potential benefits of colostrum in gastrointestinal diseases. Front. Biosci. 2016, 8, 331–351. [Google Scholar]
- Playford, R.J.; Weiser, M.J. Bovine Colostrum: Its Constituents and Uses. Nutrients 2021, 13, 265. [Google Scholar] [CrossRef]
- McGrath, B.A.; Fox, P.F.; McSweeney, P.L.H.; Kelly, A.L. Composition and properties of bovine colostrum: A review. Dairy Sci. Technol. 2016, 96, 133–158. [Google Scholar] [CrossRef]
- Sienkiewicz, M.; Jaśkiewicz, A.; Tarasiuk, A.; Fichna, J. Lactoferrin: An overview of its main functions, immunomodulatory and antimicrobial role, and clinical significance. Crit. Rev. Food Sci. Nutr. 2021, Mar 8, 1–18. [Google Scholar] [CrossRef]
- Noel, P.; Patel, K.; Durgampudi, C.; Trivedi, R.N.; de Oliveira, C.; Crowell, M.D.; Pannala, R.; Lee, K.; Brand, R.; Chennat, J.; et al. Peripancreatic fat necrosis worsens acute pancreatitis independent of pancreatic necrosis via unsaturated fatty acids increased in human pancreatic necrosis collections. Gut 2016, 65, 100–111. [Google Scholar] [CrossRef] [Green Version]
- Yadav, R.; Angolkar, T.; Kaur, G.; Buttar, H.S. Antibacterial and Antiinflammatory Properties of Bovine Colostrum. Recent Pat. Inflamm. Allergy Drug Discov. 2016, 10, 49–53. [Google Scholar] [CrossRef]
- van Hooijdonk, A.C.; Kussendrager, K.D.; Steijns, J.M. In vivo antimicrobial and antiviral activity of components in bovine milk and colostrum involved in non-specific defence. Br. J. Nutr. 2000, 84, 127–134. [Google Scholar] [CrossRef] [Green Version]
- Benson, K.F.; Carter, S.G.; Patterson, K.M.; Patel, D.; Jensen, G.S. A novel extract from bovine colostrum whey supports anti-bacterial and anti-viral innate immune functions in vitro and in vivo: I. Enhanced immune activity in vitro translates to improved microbial clearance in animal infection models. Prev. Med. 2012, 54, S116–S123. [Google Scholar] [CrossRef]
- Berlutti, F.; Pantanella, F.; Natalizi, T.; Frioni, A.; Paesano, R.; Polimeni, A.; Valenti, P. Antiviral properties of lactoferrin\-a natural immunity molecule. Molecules 2011, 16, 6992–7012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, K.E.; Carter, D.A. The Antifungal Activity of Lactoferrin and Its Derived Peptides: Mechanisms of Action and Synergy with Drugs against Fungal Pathogens. Front. Microbiol. 2017, 8, 2. [Google Scholar] [CrossRef] [Green Version]
- Arrowood, M.J.; Mead, J.R.; Mahrt, J.L.; Sterling, C.R. Effects of immune colostrum and orally administered antisporozoite monoclonal antibodies on the outcome of Cryptosporidium parvum infections in neonatal mice. Infect. Immun. 1989, 57, 2283–2288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jauhiainen, T.; Korpela, R. Milk Peptides and Blood Pressure. J. Nutr. 2007, 137, S825–S829. [Google Scholar] [CrossRef] [Green Version]
- Doursout, M.-F.; Horton, H.; Hoang, L.; Liang, Y.; Hwang, S.-A.; Boyd, S.; Actor, J.K.; Kruzel, M.L. Lactoferrin moderates LPS-induced hypotensive response and gut injury in rats. Int. Immunopharmacol. 2004, 15, 227–231. [Google Scholar] [CrossRef]
- Hayashida, K.-I.; Takeuchi, T.; Ozaki, T.; Shimizu, H.; Ando, K.; Miyamoto, A.; Harada, E. Bovine lactoferrin has a nitric oxide-dependent hypotensive effect in rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 286, R359–R365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, I.; Ichimura, A.; Ohue-Kitano, R.; Igarashi, M. Free fatty acid receptors in health and disease. Physiol. Rev. 2020, 100, 171–210. [Google Scholar] [CrossRef] [PubMed]
- Dawra, R.; Sharif, R.; Phillips, P.; Dudeja, V.; Dhaulakhandi, D.; Saluja, A.K. Development of a new mouse model of acute pancreatitis induced by administration of L-arginine. AJP Gastrointest. Liver Physiol. 2006, 292, G1009–G1018. [Google Scholar] [CrossRef] [Green Version]
- Kui, B.; Balla, Z.; Vasas, B.; Vegh, E.T.; Pallagi, P.; Kormanyos, E.S.; Venglovecz, V.; Ivanyi, B.; Takacs, T.; Hegyi, P.; et al. New insights into the methodology of L-arginine-induced acute pancreatitis. PLoS ONE 2015, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Masood, O. Cellular Mechanisms of L- arginine Induced Experimental Acute Pancreatitis. Ph.D. Thesis, University of Manchester, Manchester, UK, 2013. [Google Scholar]
- Fichna, J.; Dicay, M.; Lewellyn, K.; Janecka, A.; Zjawiony, J.K.; MacNaughton, W.K.; Storr, M.A. Salvinorin A has antiinflammatory and antinociceptive effects in experimental models of colitis in mice mediated by KOR and CB1 receptors. Inflamm. Bowel Dis. 2012, 18, 1137–1145. [Google Scholar] [CrossRef]
- Cardiff, R.D.; Miller, C.H.; Munn, R.J. Manual hematoxylin and eosin staining of mouse tissue sections. Cold Spring Harb. Protoc. 2014, 2014, 655–658. [Google Scholar] [CrossRef]
- Bancroft, J.D.; Gamble, M. Theory and Practice of Histological Techniques; Churchill Livingstone: London, UK, 2008. [Google Scholar]
- Matull, W.R.; Pereira, S.P.; O’Donohue, J.W. Biochemical markers of acute pancreatitis. J. Clin. Pathol. 2006, 59, 340–344. [Google Scholar] [CrossRef] [Green Version]
- Tenner, S.; Baillie, J.; DeWitt, J.; Vege, S.S. American College of Gastroenterology guideline: Management of acute pancreatitis. Am. J. Gastroenterol. 2013, 108, 1400–1415. [Google Scholar] [CrossRef] [PubMed]
- Hegyi, P.; Rakonczay, Z., Jr.; Sari, R.; Gog, C.; Lonovics, J.; Takacs, T.; Czako, L. L-arginine-induced experimental pancreatitis. World J. Gastroenterol. 2004, 10, 2003–2009. [Google Scholar] [CrossRef] [PubMed]
- Kui, B.; Balla, Z.; Vegh, E.T.; Pallagi, P.; Venglovecz, V.; Ivanyi, B.; Takacs, T.; Hegyi, P.; Rakonczay, Z., Jr. Recent advances in the investigation of pancreatic inflammation induced by large doses of basic amino acids in rodents. Lab. Investig. 2014, 94, 138–149. [Google Scholar] [CrossRef] [Green Version]
- Houthuijzen, J.M. For Better or Worse: FFAR1 and FFAR4 Signaling in Cancer and Diabetes. Mol Pharmacol. 2016, 90, 738–743. [Google Scholar] [CrossRef] [Green Version]
- Hidalgo, M.A.; Nahuelpan, C.; Manosalva, C.; Jara, E.; Carretta, M.D.; Conejeros, I.; Loaiza, A.; Chihuailaf, R.; Burgos, R.A. Oleic acid induces intracellular calcium mobilization, MAPK phosphorylation, superoxide production and granule release in bovine neutrophils. Biochem. Biophys. Res. Commun. 2011, 409, 280–286. [Google Scholar] [CrossRef] [PubMed]
- Fujita, T.; Matsuoka, T.; Honda, T.; Kabashima, K.; Hirata, T.; Narumiya, S. A GPR40 agonist GW9508 suppresses CCL5, CCL17, and CXCL10 induction in keratinocytes and attenuates cutaneous immune inflammation. J. Investig. Dermatol. 2011, 131, 1660–1667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagatake, T.; Shiogama, Y.; Inoue, A.; Kikuta, J.; Honda, T.; Tiwari, P.; Kishi, T.; Yanagisawa, A.; Isobe, Y.; Matsumoto, N.; et al. The 17,18-epoxyeicosatetraenoic acid-G protein-coupled receptor 40 axis ameliorates contact hypersensitivity by inhibiting neutrophil mobility in mice and cynomolgus macaques. J. Allergy Clin. Immunol. 2018, 142, 470–484. [Google Scholar] [CrossRef] [Green Version]
- Oh, D.Y.; Talukdar, S.; Bae, W.J.; Imamura, T.; Morinaga, H.; Fan, W.Q.; Li, P.; Lu, W.J.; Watkins, S.M.; Olefsky, J.M. GPR120 is an omega-3 fatty acid receptor mediating potent anti-inflammatory and insulin-sensitizing effects. Cell 2010, 142, 687–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, D.Y.; Olefsky, J.M. Omega 3 fatty acids and GPR120. Cell Metab. 2012, 15, 564–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, H.; Umemoto, T.; Kakei, M.; Momomura, S.-i.; Kawakami, M.; Ishikawa, S.-e.; Hara, K. Eicosapentaenoic acid shows anti-inflammatory effect via GPR120 in 3T3-L1 adipocytes and attenuates adipose tissue inflammation in diet-induced obese mice. Nutr. Metab. 2017, 14, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Zha, D.; Wang, L.; Qiao, L.; Lu, L.; Mei, L.; Chen, C.; Qiu, J. Phenotypic characterization of GPR120-expressing cells in the interstitial tissue of pancreas. Tissue Cell 2013, 45, 421–427. [Google Scholar] [CrossRef]
- Konno, Y.; Ueki, S.; Takeda, M.; Kobayashi, Y.; Tamaki, M.; Moritoki, Y.; Oyamada, H.; Itoga, M.; Kayaba, H.; Omokawa, A.; et al. Functional analysis of free fatty acid receptor GPR120 in human eosinophils: Implications in metabolic homeostasis. PLoS ONE 2015, 10, e0120386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, P.E.; Scales, M.K.; Allen, B.L.; di Magliano, M.P. Pancreatic Fibroblast Heterogeneity: From Development to Cancer. Cells 2020, 9, 2464. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tarasiuk, A.; Talar, M.; Bulak, K.; Fichna, J. Ghee Butter from Bovine Colostrum Reduces Inflammation in the Mouse Model of Acute Pancreatitis with Potential Involvement of Free Fatty Acid Receptors. Nutrients 2021, 13, 3271. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13093271
Tarasiuk A, Talar M, Bulak K, Fichna J. Ghee Butter from Bovine Colostrum Reduces Inflammation in the Mouse Model of Acute Pancreatitis with Potential Involvement of Free Fatty Acid Receptors. Nutrients. 2021; 13(9):3271. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13093271
Chicago/Turabian StyleTarasiuk, Aleksandra, Marcin Talar, Kamila Bulak, and Jakub Fichna. 2021. "Ghee Butter from Bovine Colostrum Reduces Inflammation in the Mouse Model of Acute Pancreatitis with Potential Involvement of Free Fatty Acid Receptors" Nutrients 13, no. 9: 3271. https://0-doi-org.brum.beds.ac.uk/10.3390/nu13093271