
Probiotic Consumption Boosts Thymus in Obesity and Senescence Mouse Models

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain and Culture Conditions
2.2. Isolation of the Cell Wall of a Probiotic Bacterium
2.3. Animals, Feeding Procedures and Study Design for Obese and Aging Experimental Mouse Models
- 1.
- Normal control (NC): animals fed ad libitum with conventional food and water;
- 1.2.
- Normal control mice supplemented with probiotics (NC+P): mice received a conventional diet and a suspension of the probiotic bacterium in the drinking water;
- 2.
- Obese control (OC): animals fed ad libitum with the HFD and water;
- 2.1.
- Obese plus probiotics (OC+P): OC animals received a HFD and a suspension containing probiotic bacteria. See details in Figure S1B.
2.4. Serum and Small Intestinal Fluid Samples
2.5. Analysis of Intestinal Microbiota
2.6. Histological Studies
2.7. Determination of T-Cell Population in the Thymus
2.8. Ex Vivo Assays in Cell Cultures
- Normal controls of different ages;
- Normal controls of different ages with live probiotic bacteria (NC+B);
- Normal controls of different ages with probiotic cell wall (NC+W).
2.9. Determination of Cytokine Production
2.10. Statistical Analysis
3. Results
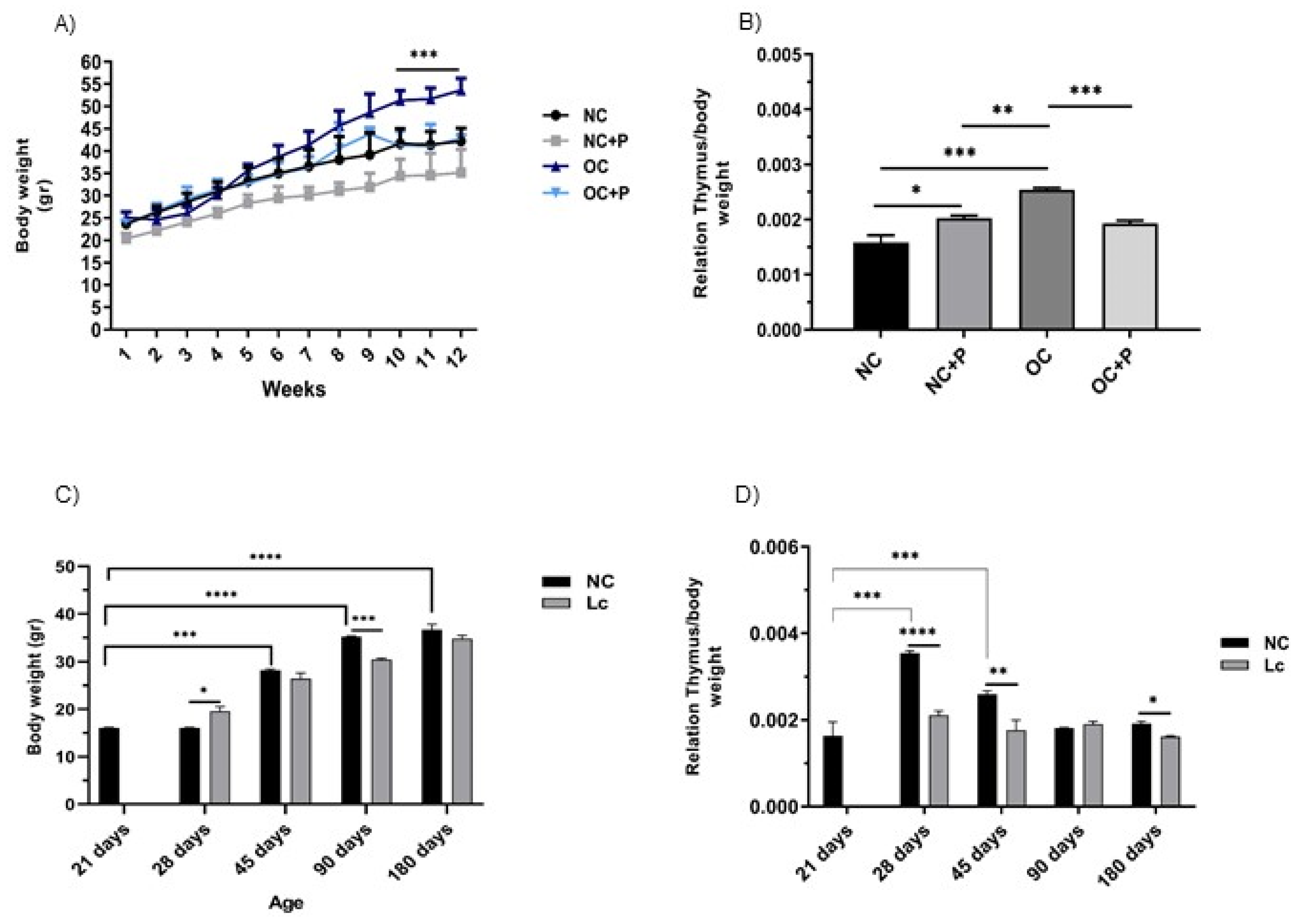
3.1. Changes in Body Weight and Thymus. Effects of Probiotic Supplementation on These Parameters in Obese and Senescent Mice
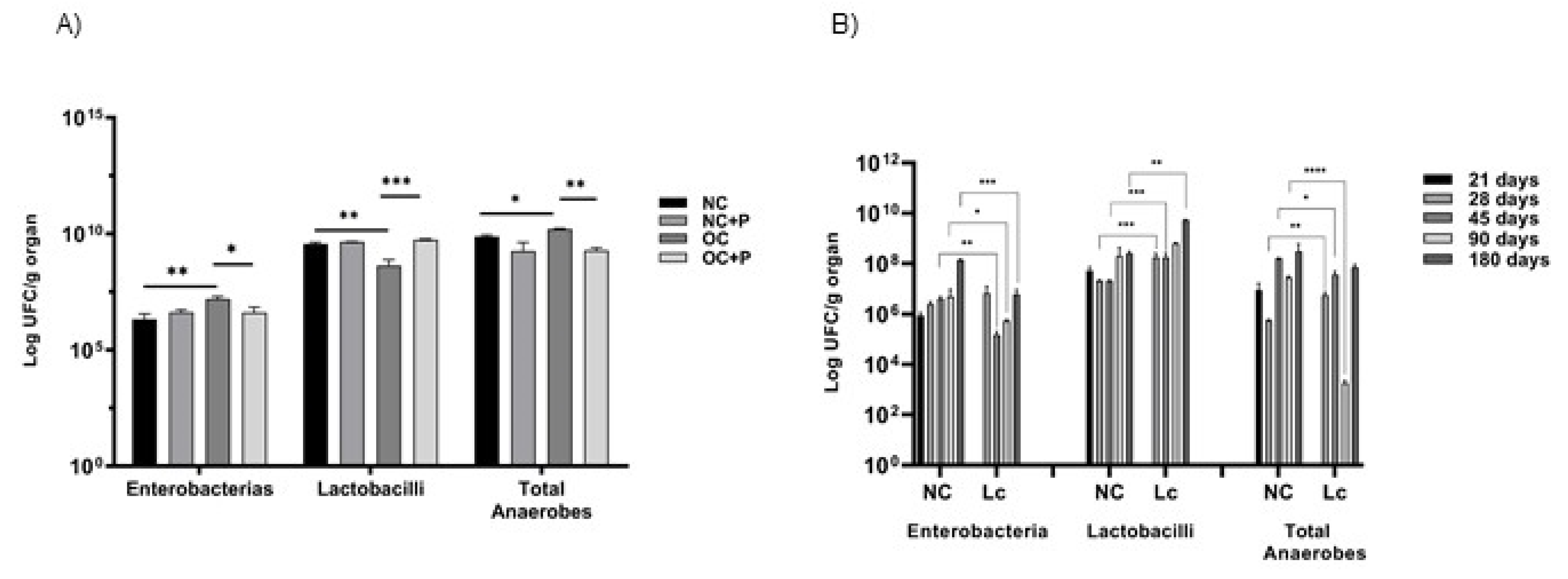
3.2. Effect of Probiotic Supplementation on the Intestinal Microbiota
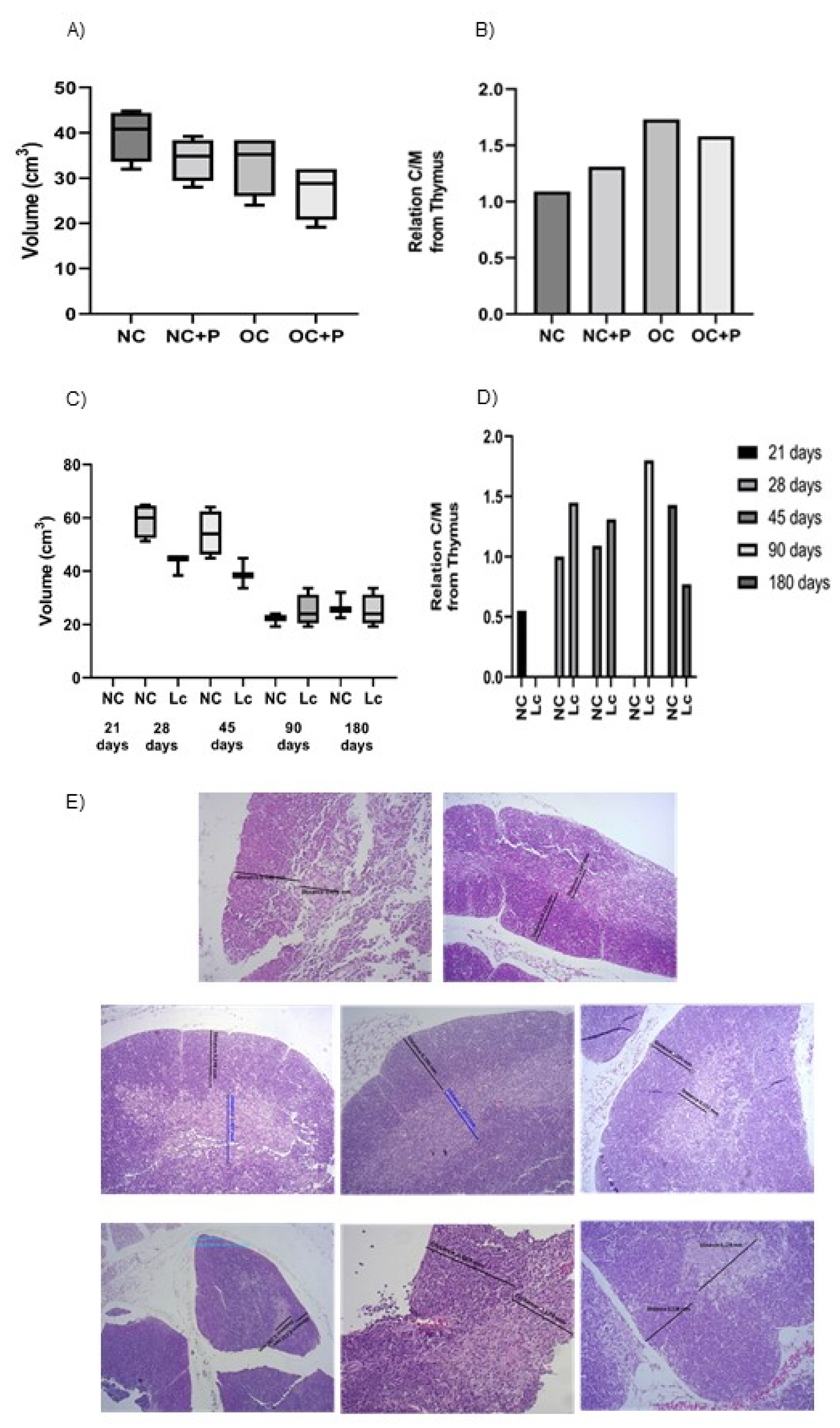
3.3. Histological Studies in Thymus
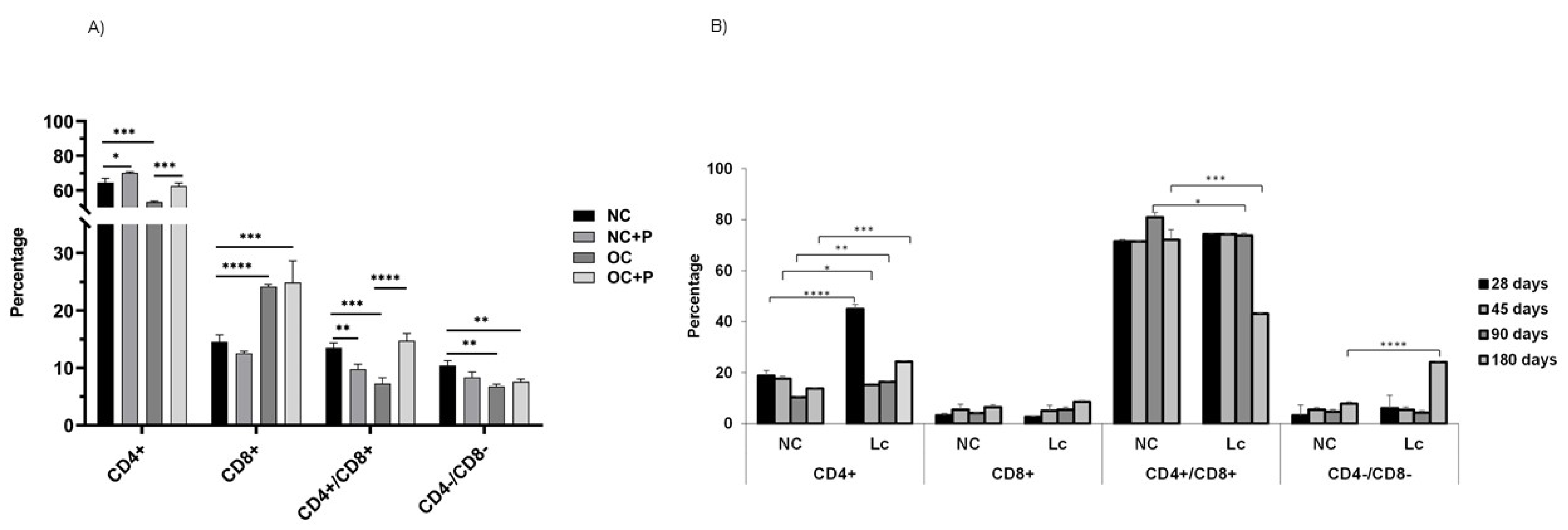
3.4. Determination of CD4+ and CD8+ T Cell Population in Thymus
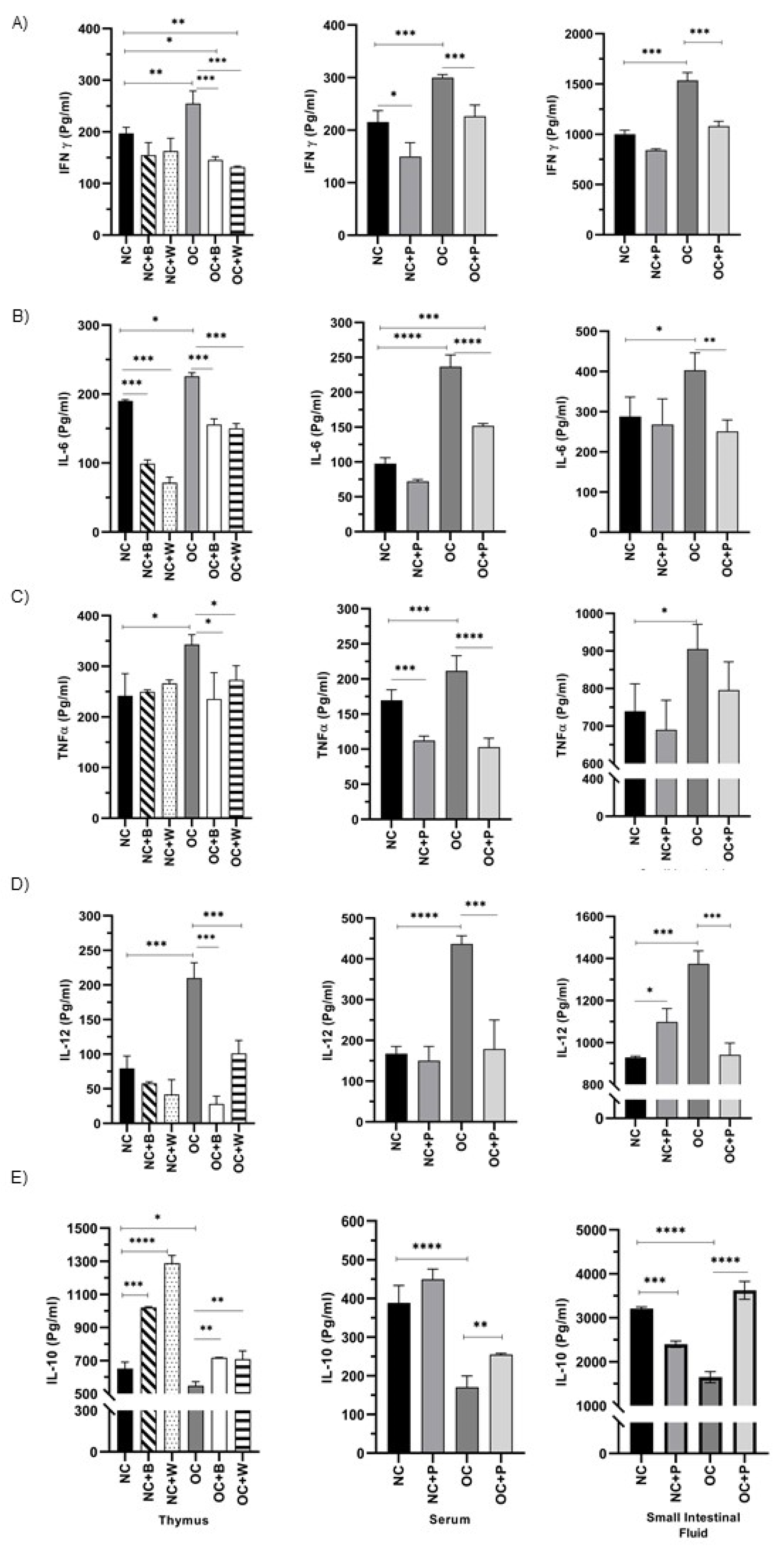
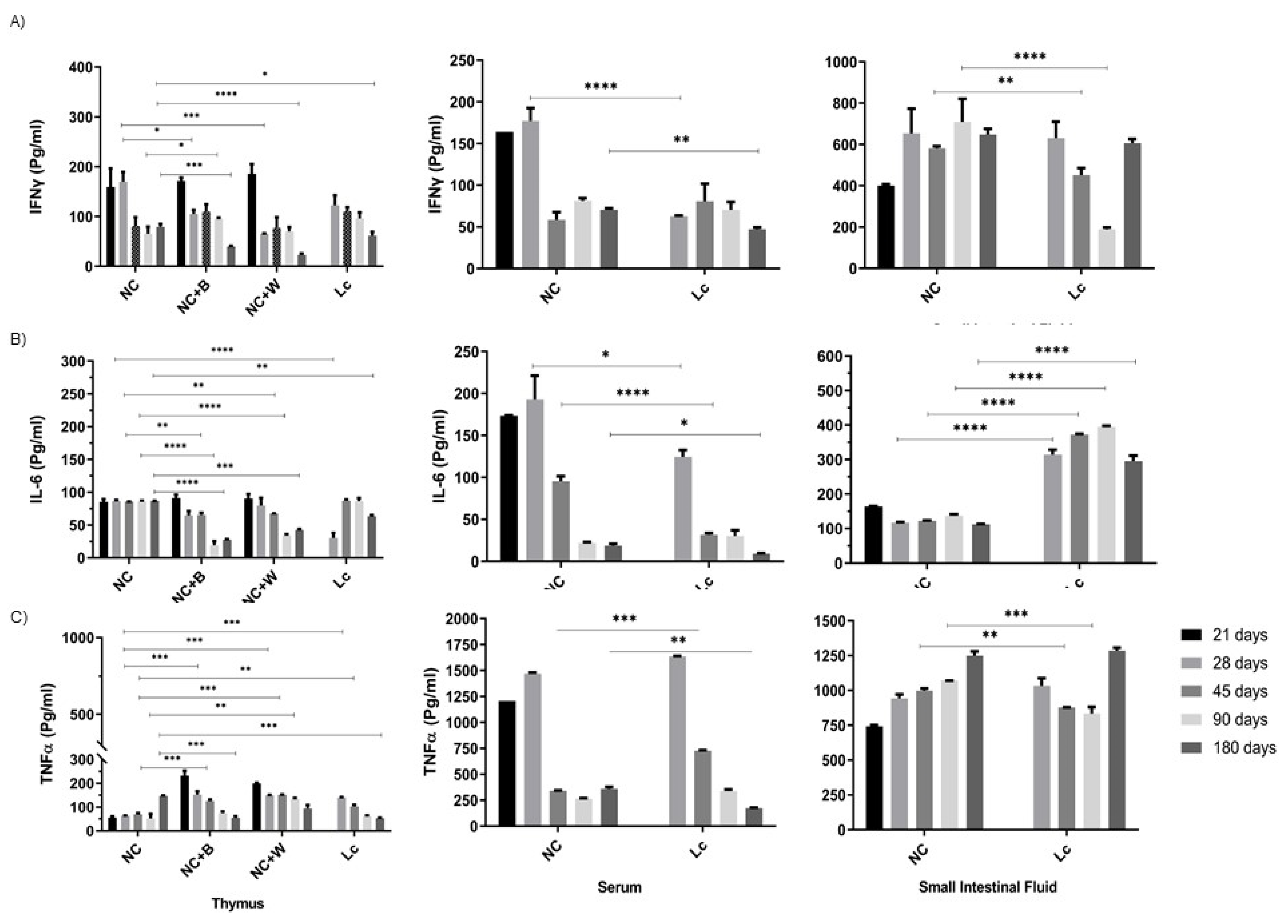
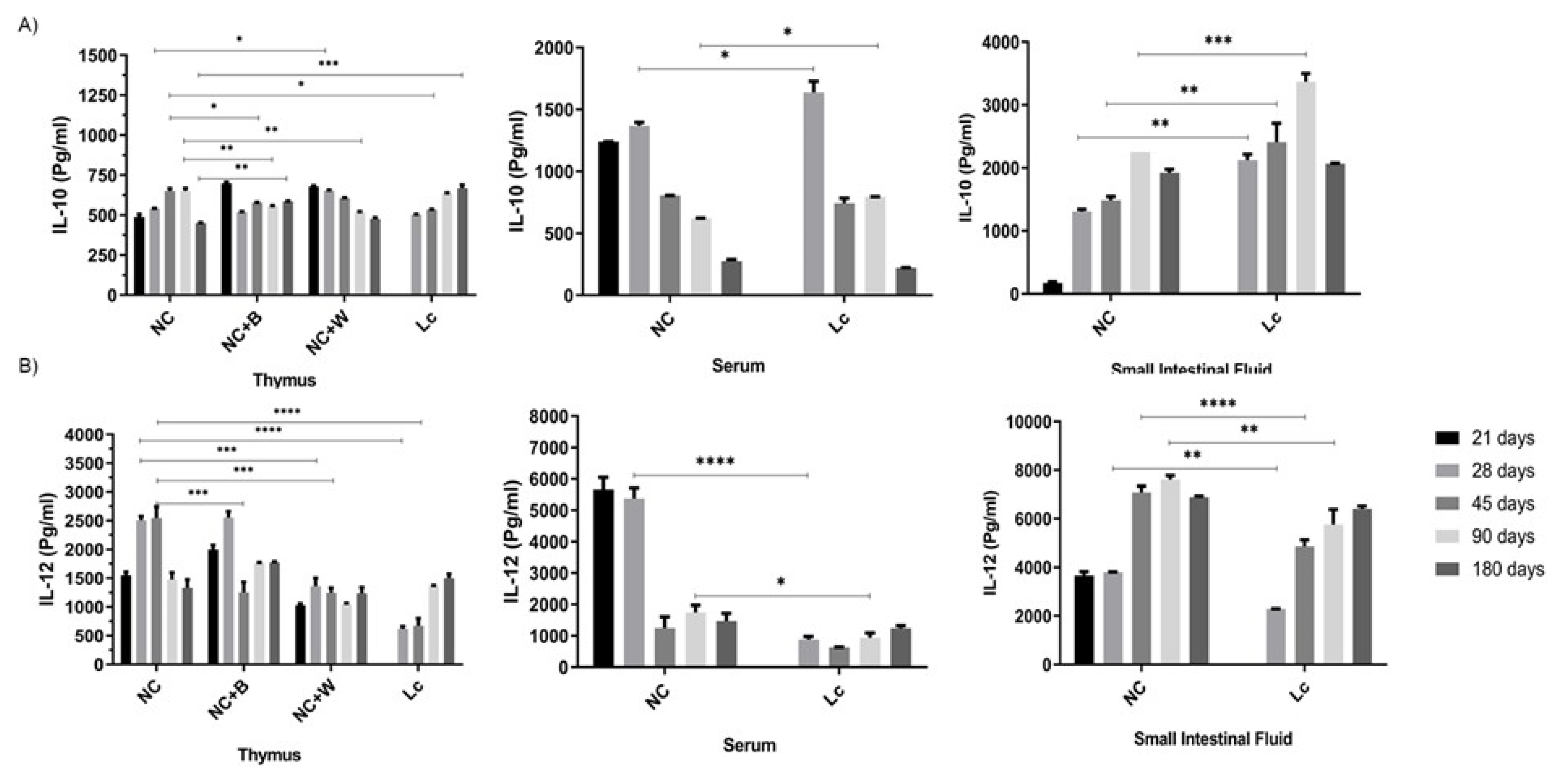
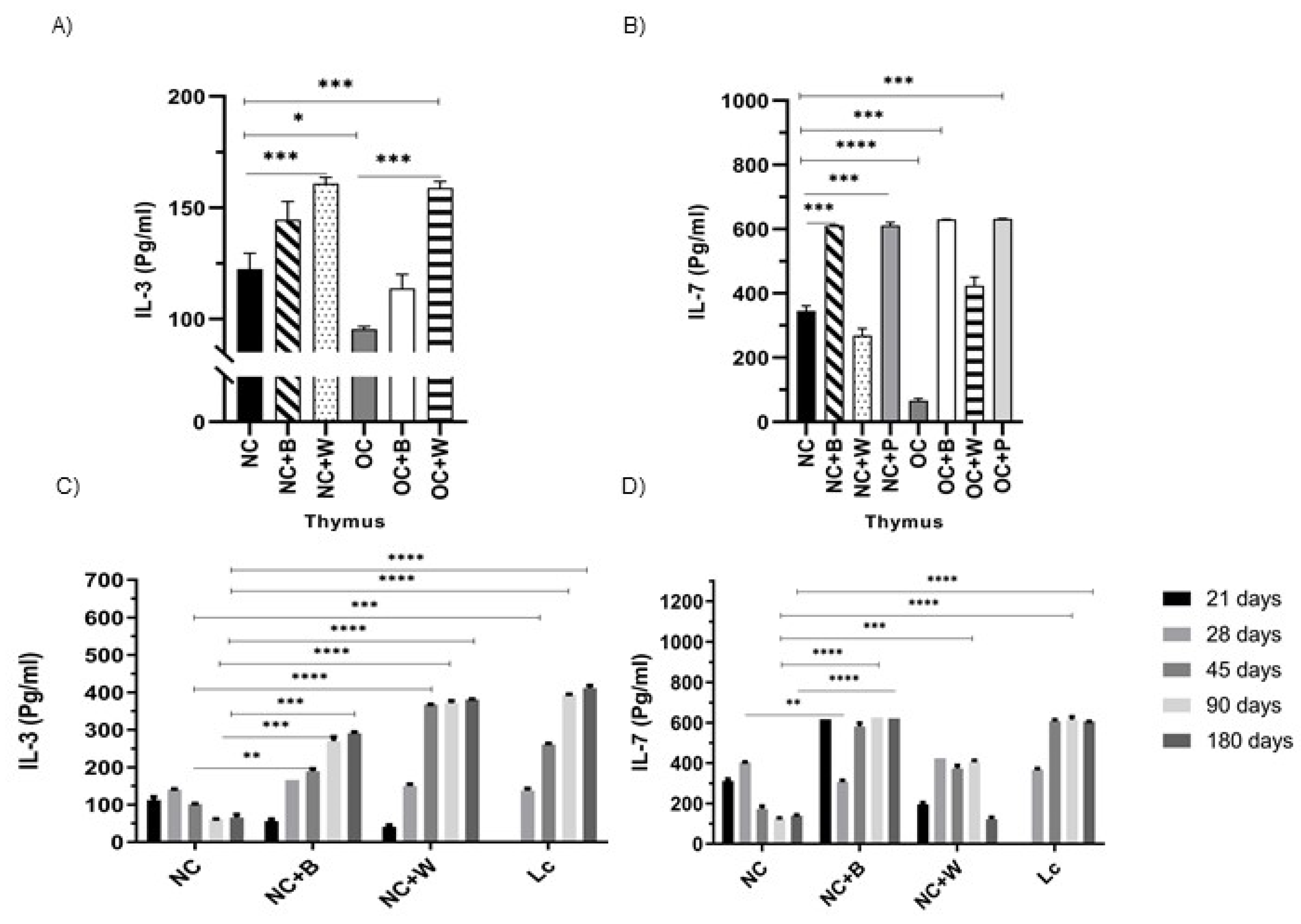
3.5. Determination of Cytokine Production by Ex Vivo and In Vitro Studies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boehm, T. Thymus development and function. Curr. Opin. Immunol. 2008, 20, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Palmer, D.B. The effect of age on thymic function. Front. Immunol. 2013, 4, 316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pobezinsky, L.A.; Angelov, G.S.; Tai, X.; Jeurling, S.; Van Laethem, F.; Feigenbaum, L.; Park, J.H.; Singer, A. Clonal deletion and the fate of autoreactive thymocytes that survive negative selection. Nat. Immunol. 2012, 13, 569–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savino, W.; Dardenne, M. Neuroendocrine Control of Thymus Physiology. Endocr. Rev. 2000, 21, 412–443. [Google Scholar] [PubMed]
- Scalea, J.R.; Hickman, J.B.; Moore, D.J.; Brayman, K.L. An overview of the necessary thymic contributions to tolerance in transplantation. Clin. Immunol. 2016, 173, 1–9. [Google Scholar] [CrossRef]
- Ladi, E.; Yin, X.; Chtanova, T.; Robey, E.A. Thymic microenvironments for T cell differentiation and selection. Nat. Immunol. 2006, 7, 338–343. [Google Scholar] [CrossRef]
- Nitta, T.; Ohigashi, I.; Nakagawa, Y.; Takahama, Y. Cytokine crosstalk for thymic medulla formation. Curr. Opin. Immunol. 2011, 23, 190–197. [Google Scholar] [CrossRef]
- Romagnoli, P.; Van Meerwijk, J.P.M. Thymic selection and lineage commitment of CD4+Foxp3+ regulatory T lymphocytes. Prog. Mol. Biol. Transl. Sci. 2010, 92, 251–277. [Google Scholar] [CrossRef]
- Ladi, E.; Schwickert, T.A.; Chtanova, T.; Chen, Y.; Herzmark, P.; Yin, X.; Aaron, H.; Chan, S.W.; Lipp, M.; Roysam, B.; et al. Thymocyte-dendritic cell interactions near sources of CCR7 ligands in the thymic cortex. J. Immunol. 2008, 181, 7014–7023. [Google Scholar] [CrossRef] [Green Version]
- Yin, X.; Ladi, E.; Chan, S.W.; Li, O.; Killeen, N.; Kappes, D.J.; Robey, E.A. CCR7 expression in developing thymocytes is linked to the CD4 versus CD8 lineage decision. J. Immunol. 2007, 179, 7358–7364. [Google Scholar] [CrossRef]
- Pawelec, G. Age and immunity: What is “immunosenescence”? Exp. Gerontol. 2018, 105, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Pera, A.; Campos, C.; Lopez, N.; Hassouneh, F.; Alonso, C.; Tarazona, R.; Solana, R. Immunosenescence: Implications for response to infection and vaccination in older people. Maturitas 2015, 82, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Fu, H.; Yang, H.; Wang, L.; He, Y. Clinical management of cervical ectopic thymus in children. J. Pediatr. Surg. 2011, 46, e33–e36. [Google Scholar] [CrossRef] [PubMed]
- Chowhan, A.K.; Kinnera, V.S.; Yootla, M.; Reddy, M.K. Cervical ectopic thymus masquerading as metastatic thyroid papillary carcinoma. Malays. J. Pathol. 2010, 32, 65–68. [Google Scholar] [PubMed]
- Herman, T.E.; Siegel, M.J. Cervical ectopic thymus. J. Perinatol. 2009, 29, 173–174. [Google Scholar] [CrossRef] [PubMed]
- Mizia-Malarz, A.; Sobol, G.; Maldyk, J.; Stolpa, W.; Szyszka, A.; Wos, H. Cervical ectopic thymus in a 9-month-old girl: Diagnostic difficulties. J. Pediatr. Hematol. Oncol. 2009, 31, 599–601. [Google Scholar] [CrossRef] [PubMed]
- Prasad, T.R.; Chui, C.H.; Ong, C.L.; Meenakshi, A. Cervical ectopic thymus in an infant. Singap. Med. J. 2006, 47, 68–70. [Google Scholar]
- Yang, H.; Youm, Y.H.; Dixit, V.D. Inhibition of thymic adipogenesis by caloric restriction is coupled with reduction in age-related thymic involution. J. Immunol. 2009, 183, 3040–3052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howard, J.K.; Lord, G.M.; Matarese, G.; Vendetti, S.; Ghatei, M.A.; Ritter, M.A.; Lechler, R.I.; Bloom, S.R. Leptin protects mice from starvation-induced lymphoid atrophy and increases thymic cellularity in ob/ob mice. J. Clin. Investig. 1999, 104, 1051–1059. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Yun-Hee, Y.; Bolormaa Vandanmagsar, J.R.; Ganesh Kumar, K.; Butler, A.A.; Dixit, V.D. Obesity accelerates thymic aging. Blood 2009, 114, 3803–3812. [Google Scholar] [CrossRef] [Green Version]
- Harrington, K.A.; Kennedy, D.S.; Tang, B.; Hickie, C.; Phelan, E.; Torreggiani, W.; Halpenny, D. Computed tomographic evaluation of the thymus—Does obesity affect thymic fatty involution in a healthy young adult population? Br. J. Radiol. 2018, 91, 20170609. [Google Scholar] [CrossRef] [PubMed]
- Novotny Núñez, I.; Galdeano, C.M.; Carmuega, E.; Weill, R.; de Moreno De Leblanc, A.; Perdigón, G. Effect of a probiotic fermented milk on the thymus in Balb/c mice under non-severe protein-energy malnutrition. Br. J. Nutr. 2013, 110, 500–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maldonado Galdeano, C.; Novotny Núñez, I.; de Moreno de LeBlanc, A.; Carmuega, E.; Weill, R.; Perdigón, G. Impact of a probiotic fermented milk in the gut ecosystem and in the systemic immunity using a non-severe protein-energy-malnutrition model in mice. BMC Gastroenterol. 2011, 11, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, N.P. Functional cultures and health benefits. Int. Dairy J. 2007, 17, 1262–1277. [Google Scholar] [CrossRef]
- Galdeano, C.M.; Perdigon, G. The Probiotic Bacterium Lactobacillus casei Induces Activation of the Gut Mucosal Immune System through Innate Immunity. Clin. Vaccine Immunol. 2006, 13, 219–226. [Google Scholar] [CrossRef] [Green Version]
- Galdeano, C.M.; de Moreno de LeBlanc, A.; Vinderola, G.; Bibas Bonet, M.E.; Perdigon, G. Proposed model: Mechanisms of immunomodulation induced by probiotic bacteria. Clin. Vaccine Immunol. 2007, 14, 485–492. [Google Scholar] [CrossRef] [Green Version]
- de Moreno de LeBlanc, A.; Castillo, N.A.; Perdigon, G. Anti-infective mechanisms induced by a probiotic Lactobacillus strain against Salmonella enterica serovar Typhimurium infection. Int. J. Food Microbiol. 2010, 138, 223–231. [Google Scholar] [CrossRef]
- Velez, E.; Castillo, N.; Mesón, O.; Grau, A.; Bibas Bonet, M.E.; Perdigón, G. Study of the effect exerted by fructo-oligosaccharides from yacon (Smallanthus sonchifolius) root flour in an intestinal infection model with Salmonella Typhimurium. Br. J. Nutr. 2013, 109, 1971–1979. [Google Scholar] [CrossRef] [Green Version]
- Velez, E.M.M.; Maldonado Galdeano, C.; Carmuega, E.; Weill, R.; Bibas Bonet, M.E.; Perdigon, G. Probiotic fermented milk consumption modulates the allergic process induced by ovoalbumin I. in mice. Br. J. Nutr. 2015, 114, 566–576. [Google Scholar] [CrossRef] [Green Version]
- Novotny Núñez, I.; Maldonado Galdeano, C.; de Moreno de LeBlanc, A.; Perdigón, G. Lactobacillus casei CRL 431 administration decreases inflammatory cytokines in a diet-induced obese mouse model. Nutrition 2015, 31, 1000–1007. [Google Scholar] [CrossRef]
- Castillo, N.A.; de Moreno de LeBlanc, A.; Galdeano, C.M.; Perdigón, G. Comparative study of the protective capacity against Salmonella infection between probiotic and nonprobiotic Lactobacilli. J. Appl. Microbiol. 2013, 114, 861–876. [Google Scholar] [CrossRef] [PubMed]
- Castillo, N.A.; Perdigón, G.; de Moreno de LeBlanc, A. Oral administration of a probiotic Lactobacillus modulates cytokine production and TLR expression improving the immune response against Salmonella enterica serovar Typhimurium infection in mice. BMC Microbiol. 2011, 11, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balcells, F.; Mariani, C.; Weill, R.; Perdigon, G.; Maldonado Galdeano, M.C. Effect of Yogurt With or Without Probiotic Addition on Body Composition Changes and Immune System in an Obese Model. Food Sci. Nutr. 2017, 3, 1–9. [Google Scholar]
- Lemme-Dumit, J.M.; Polti, M.A.; Perdigón, G.; Maldonado-Galdeano, C. Probiotic bacteria cell walls stimulate the activity of the intestinal epithelial cells and macrophage functionality. Benef. Microbes. 2018, 9, 153–164. [Google Scholar] [CrossRef]
- Lemme-Dumit, J.M.; Cazorla, S.I.; Perdigón, G.; Maldonado-Galdeano, C. Probiotic bacteria and their cell walls induce Th1-Type immunity against Salmonella Typhimurium challenge. Front. Immunol. 2021, 12, 660854. [Google Scholar] [CrossRef]
- Thomas, R.; Wang, W.; Su, D.-M. Contributions of Age-Related Thymic Involution to Immunosenescence and Inflammaging. Immun. Ageing 2020, 17, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Haynes, B.F.; Sempowski, G.D.; Wells, A.F.; Hale, L.P. The human thymus during aging. Immunol. Res. 2000, 22, 253–261. [Google Scholar] [CrossRef]
- Palmer, G.; Aurrand-Lions, M.; Contassot, E.; Talabot-Ayer, D.; Ducrest-Gay, D.; Vesin, C.; Chobaz-Péclat, V.; Busso, N.; Gabay, C. Indirect Effects of Leptin Receptor Deficiency on Lymphocyte Populations and Immune Response in db/db Mice. J. Immunol. 2006, 177, 2899–2907. [Google Scholar] [CrossRef] [Green Version]
- Youm, Y.-H.; Kanneganti, T.-D.; Vandanmagsar, B.; Zhu, X.; Ravussin, A.; Adijiang, A.; Owen, J.S.; Thomas, M.J.; Francis, J.; Parks, J.S.; et al. The NLRP3 Inflammasome Promotes Age-Related Thymic Demise and Immunosenescence. Cell Rep. 2012, 1, 56–68. [Google Scholar] [CrossRef] [Green Version]
- Rezzani, R.; Nardo, L.; Favero, G.; Peroni, M.; Rodella, L.R. Thymus and aging: Morphological, radiological, and functional overview. Age 2014, 36, 313–351. [Google Scholar] [CrossRef] [Green Version]
- Savino, W.; Dardenne, M.; Velloso, L.A.; Silva-Barbosa, S.D. The thymus is a common target in malnutrition and infection. Br. J. Nutr. 2007, 98, S11–S16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez, A.R.; Berbert, L.R.; Lepletier, A.; Revelli, S.; Bottasso, O.; Silva-Barbosa, S.D.; Savino, W. TNF-α Is Involved in the Abnormal Thymocyte Migration during experimental Trypanosoma cruzi Infection and favors the export of immature cells. PLoS ONE 2012, 7, e34360. [Google Scholar] [CrossRef] [PubMed]
- Jolly, C.A. Dietary Restriction and Immune Function. J. Nutr. 2004, 134, 1853–1856. [Google Scholar] [CrossRef] [PubMed]
- Kuwahara, M.; Suzuki, J.; Tofukuji, S.; Yamada, T.; Kanoh, M.; Matsumoto, A.; Maruyama, S.; Kometani, K.; Kurosaki, T.; Ohara, O.; et al. The Menin-Bach2 axis is critical for regulating CD4 T-cell senescence and cytokine homeostasis. Nat. Commun. 2014, 5, 3555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, K.D.; Vodyanik, M.; Slukvin, I.I. Hematopoietic differentiation and production of mature myeloid cells from human pluripotent stem cells. Nat. Protoc. 2011, 6, 296–313. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, M. Effects of hemopoietic growth factors on stem cells in vitro. Hematol. Oncol. Clin. N. Am. 1989, 3, 453–464. [Google Scholar] [CrossRef]
- Smith, C. Hematopoietic stem cells and hematopoiesis. Cancer Control 2003, 1, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Ventevogel, M.S.; Sempowski, G.D. Thymic rejuvenation and aging. Curr. Opin. Immunol. 2013, 25, 516–522. [Google Scholar] [CrossRef] [Green Version]
- Mackall, C.L.; Fry, T.J.; Bare, C.; Morgan, P.; Galbraith, A.; Gress, R.E. IL-7 increases both thymic-dependent and thymic independent T-cell regeneration after bone marrow transplantation. Blood 2001, 97, 1491–1497. [Google Scholar] [CrossRef] [Green Version]
- Mackall, C.L.; Fry, T.J.; Gress, R.E. Harnessing the biology of IL-7 for therapeutic application. Nat. Rev. Immunol. 2011, 11, 330–342. [Google Scholar] [CrossRef]
- Barata, J.T.; Durum, S.K.; Seddon, B. Flip the coin: IL-7 and IL-7R in health and disease. Nat. Immunol. 2019, 20, 1584–1593. [Google Scholar] [CrossRef] [PubMed]
- Henson, S.M.; Snelgrove, R.; Hussell, T.; Wells, D.J.; Aspinall, R. An IL-7 Fusion Protein That Shows Increased Thymopoietic Ability. J. Immunol. 2005, 175, 4112–4118. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1131. [Google Scholar] [CrossRef] [PubMed]
- Kasai, C.; Sugimoto, K.; Moritani, I.; Tanaka, J.; Oya, Y.; Inoue, H.; Tameda, M.; Shiraki, K.; Ito, M.; Takei, Y.; et al. Comparison of the gut microbiota composition between obese and non-obese individuals in a Japanese population, as analyzed by terminal restriction fragment length polymorphism and next-generation sequencing. BMC Gastroenterol. 2015, 15, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langille, M.G.I.; Meehan, C.J.; Koenig, J.E.; Dhanani, A.S.; Rose, R.A.; Howlett, S.E.; Beiko, R.G. Microbial shifts in the aging mouse gut. Microbiome 2014, 2, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P.; et al. Human gut microbiome viewed across age and geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Musso, G.; Gambino, R.; Cassader, M. Obesity, Diabetes, and Gut Microbiota: The hygiene hypothesis expanded? Diabetes Care 2010, 33, 2277–2284. [Google Scholar] [CrossRef] [Green Version]
- McNabney, S.; Henagan, T. Short Chain Fatty Acids in the Colon and Peripheral Tissues: A Focus on Butyrate, Colon Cancer, Obesity and Insulin Resistance. Nutrients 2017, 9, 1348. [Google Scholar] [CrossRef] [Green Version]
- Bischoff, S.C.; Barbara, G.; Buurman, W.; Ockhuizen, T.; Schulzke, J.-D.; Serino, M.; Tilg, H.; Watson, A.; Wells, J.M. Intestinal permeability—A new target for disease prevention and therapy. BMC Gastroenterol. 2014, 14, 189. [Google Scholar] [CrossRef] [Green Version]
- Ragonnaud, E.; Biragyn, A. Gut microbiota as the key controllers of “healthy” aging of elderly people. Ragonnaud and Biragyn. Immun. Ageing 2021, 18, 2. [Google Scholar] [CrossRef]
- Biagi, E.; Nylund, L.; Candela, M.; Ostan, R.; Bucci, L.; Elisa Pini, E.; Janne Nikkïla, J.; Monti, D.; Satokari, R.; Franceschi, C.; et al. Through Ageing, and Beyond: Gut Microbiota and Inflammatory Status in Seniors and Centenarians. PLoS ONE 2010, 5, e10667. [Google Scholar] [CrossRef]
- de Moreno de LeBlanc, A.; Maldonado Galdeano, C.; Chaves, S.; Perdigon, G. Oral administration of L. casei CRL 431 increases immunity in bronchus and mammary glands. Eur. J. Inflamm. 2005, 3, 23–28. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balcells, F.; Martínez Monteros, M.J.; Gómez, A.L.; Cazorla, S.I.; Perdigón, G.; Maldonado-Galdeano, C. Probiotic Consumption Boosts Thymus in Obesity and Senescence Mouse Models. Nutrients 2022, 14, 616. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14030616
Balcells F, Martínez Monteros MJ, Gómez AL, Cazorla SI, Perdigón G, Maldonado-Galdeano C. Probiotic Consumption Boosts Thymus in Obesity and Senescence Mouse Models. Nutrients. 2022; 14(3):616. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14030616
Chicago/Turabian StyleBalcells, Florencia, María José Martínez Monteros, Alba Lorena Gómez, Silvia Inés Cazorla, Gabriela Perdigón, and Carolina Maldonado-Galdeano. 2022. "Probiotic Consumption Boosts Thymus in Obesity and Senescence Mouse Models" Nutrients 14, no. 3: 616. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14030616