
Network Pharmacology Exploration Reveals Gut Microbiota Modulation as a Common Therapeutic Mechanism for Anti-Fatigue Effect Treated with Maca Compounds Prescription

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. The Preparation of MCP
2.1.2. The Measurement of the Main Components in MCP
2.2. The Prediction of MCP Targets in Fatigue Using Network Pharmacology
2.2.1. The Screening of Active Ingredients and Potential Targets
2.2.2. Prediction of Anti-Fatigue Targets of the MCP
2.2.3. The Construction of Active Ingredients: Target Network
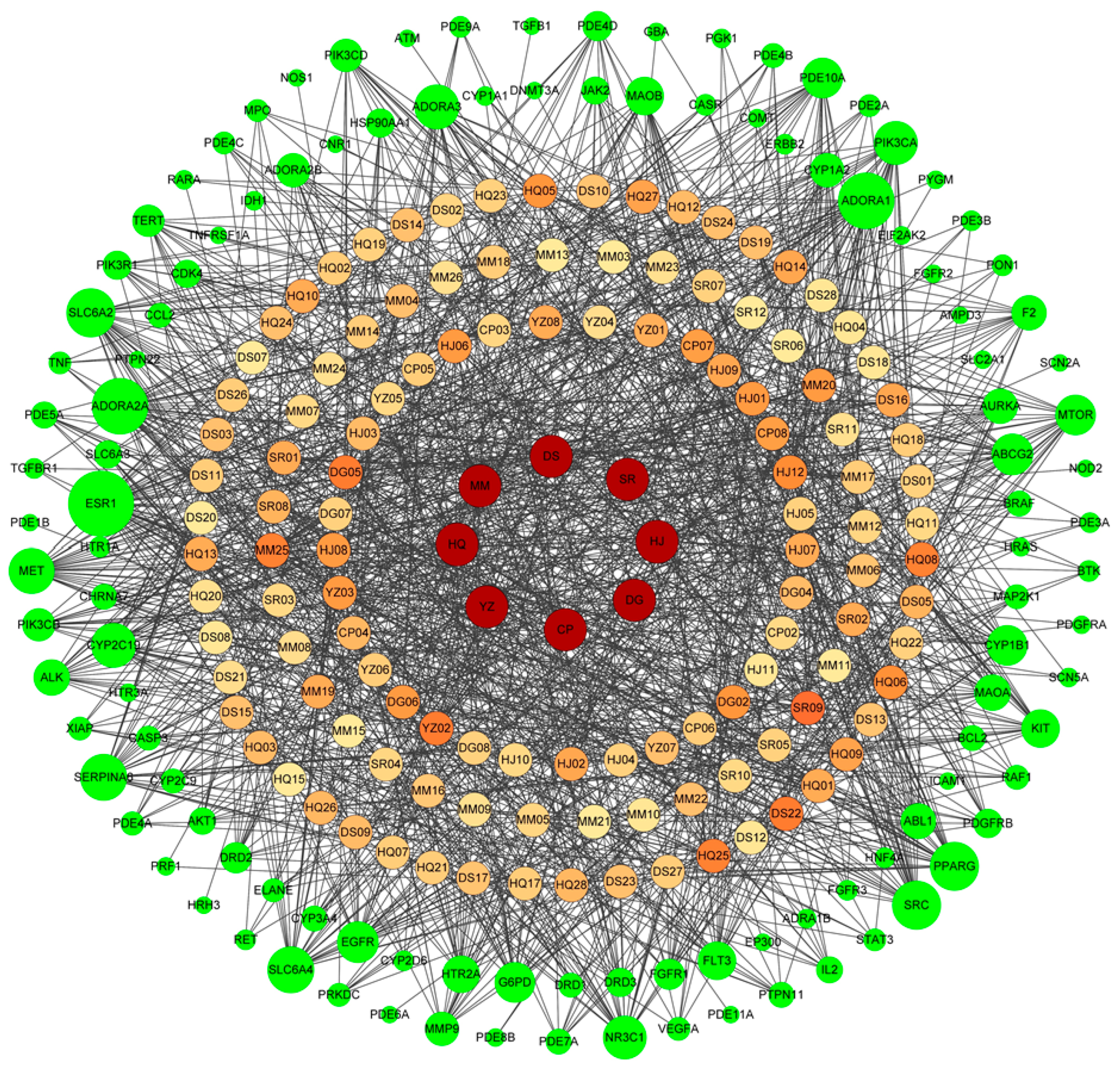
2.2.4. The Construction of the Protein Interaction Network
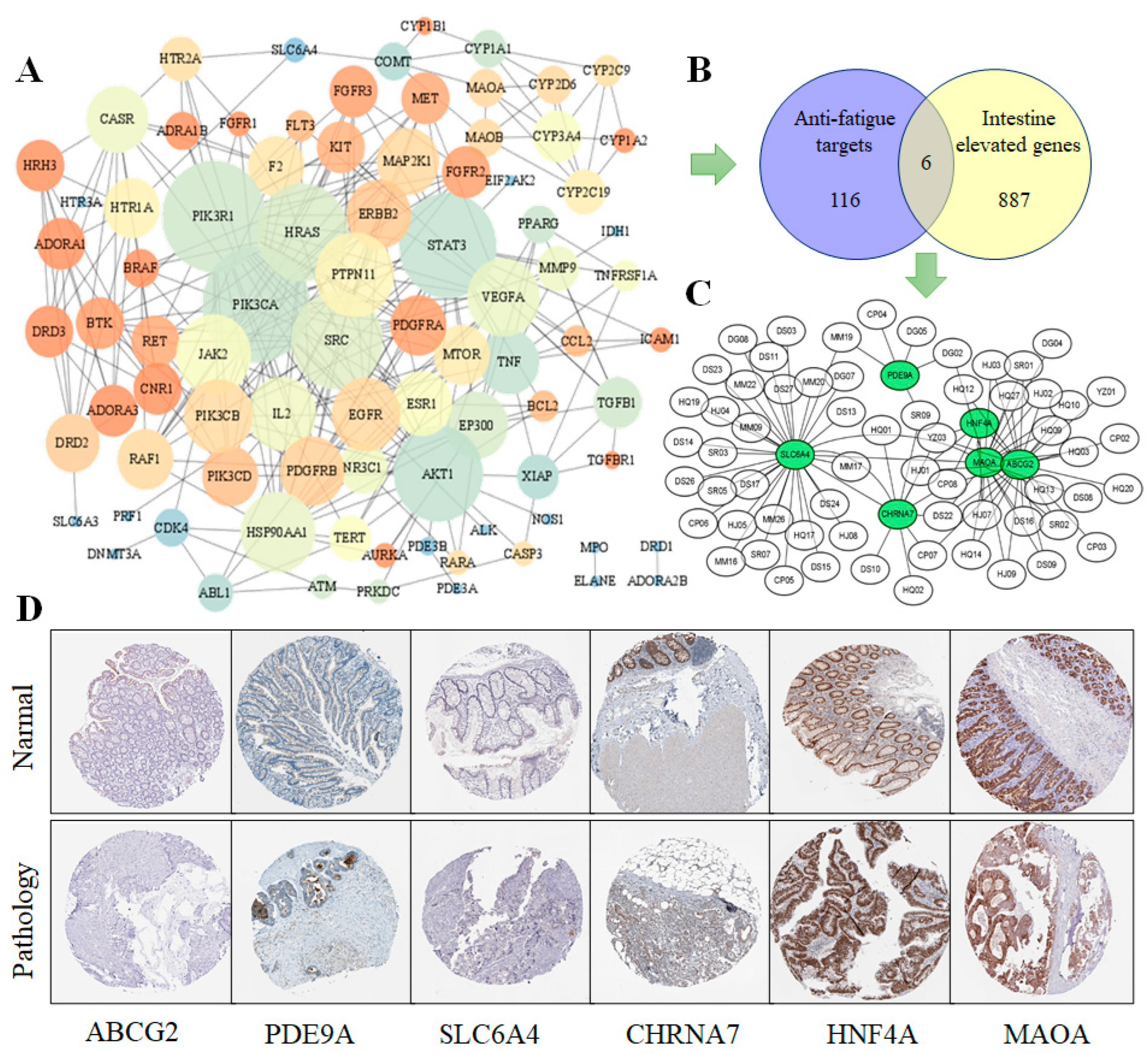
2.2.5. Tissue-Specific Gene Expression Analysis
2.2.6. Functional Enrichment Analysis
2.3. Gut Microbiota Composition in Swimming Mice
2.3.1. Experimental Animals
2.3.2. Weight-Loaded Forced Swimming Test (WFST)
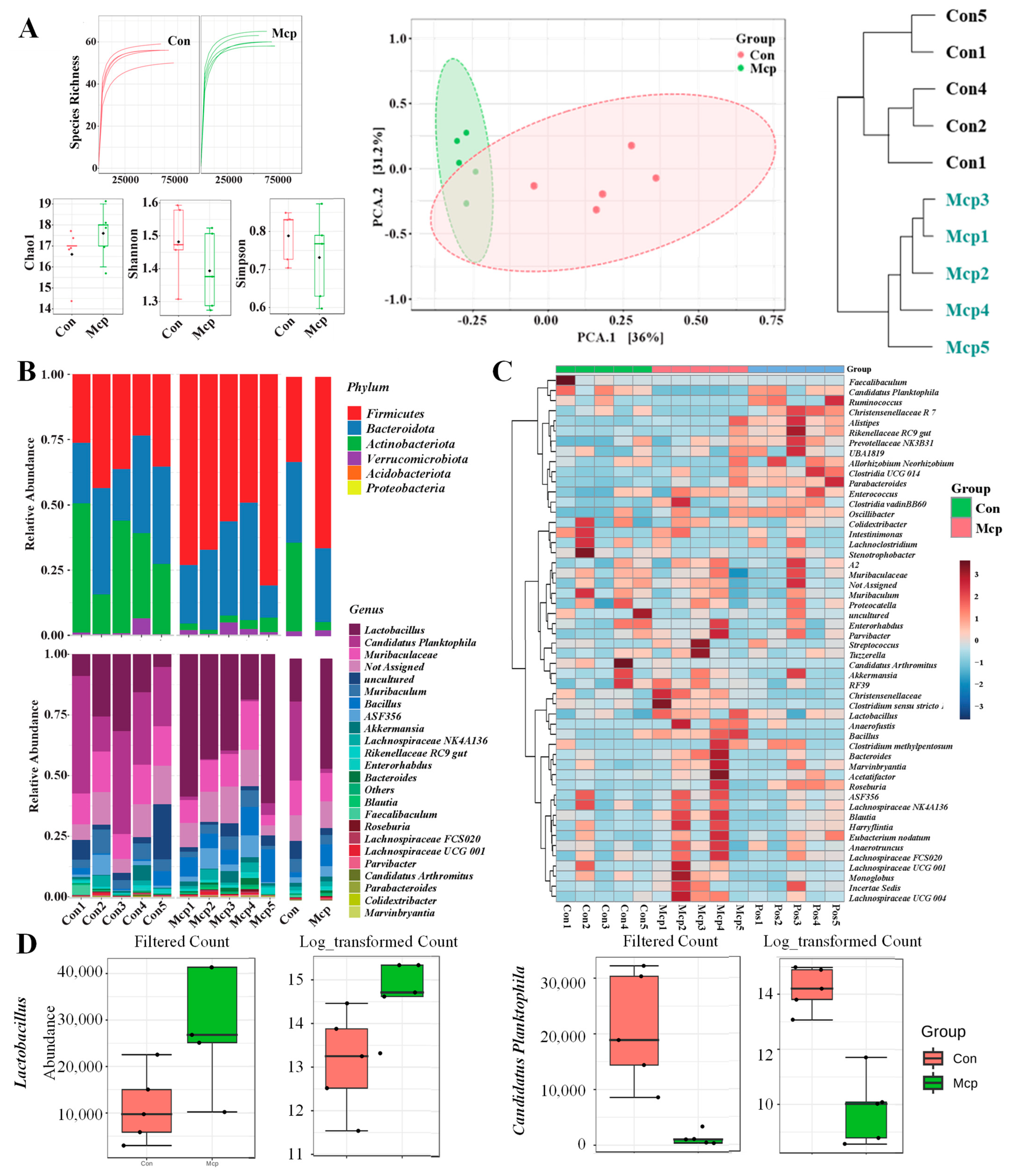
2.3.3. 16S rRNA Gene and Bioinformatics Analysis
2.4. Data Analysis
3. Results and Discussion
3.1. Measurement of the MCP Components
3.2. Network Pharmacology Analysis of the Main Components of the MCP
3.3. PPI Core Network Analysis and Specificity of Targets in Tissue Expression
3.4. Improvement on the Bacterial Diversity and Richness of Mice Gut Microbiota
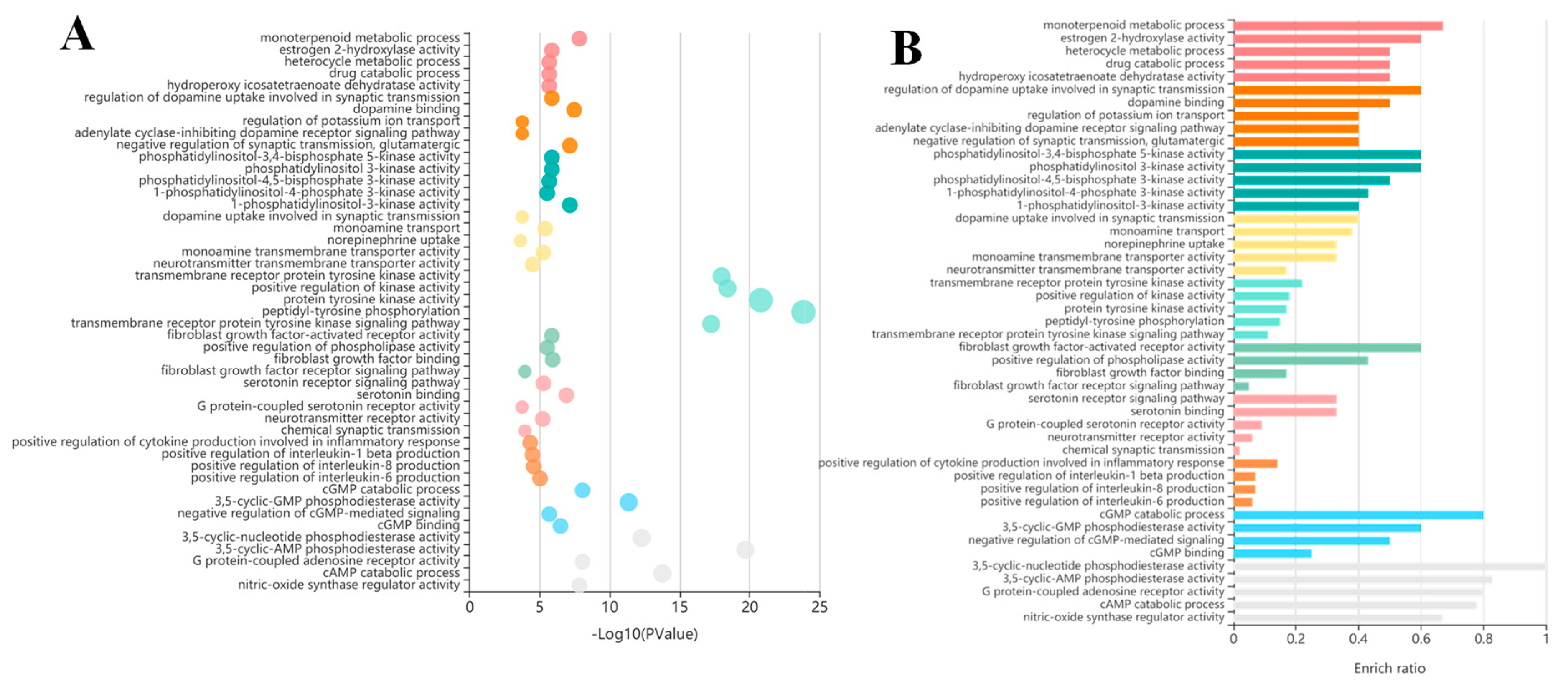
3.5. GO and KEGG Enrichment Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.Y.; Ryu, B.M.; Cui, Y.H.; Li, C.Y.; Zhou, C.X.; Hong, P.Z.; Lee, B.; Qian, Z.J. A peptide isolated from Hippocampus abdominalis improves exercise performance and exerts anti-fatigue effects via AMPK/PGC-1 alpha pathway in mice. J. Funct. Foods 2019, 61, 103489. [Google Scholar] [CrossRef]
- Howell, L.L.; Cunningham, K.A. Serotonin 5-HT2 Receptor Interactions with Dopamine Function: Implications for Therapeutics in Cocaine Use Disorder. Pharmacol. Rev. 2015, 67, 176–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Huang, L.Q.; Tang, X.C.; Zhang, H.Y. Retrospect and prospect of active principles from Chinese herbs in the treatment of dementia. Acta Pharm. Sin. 2010, 31, 649–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, C.; Xu, X.; Wei, X.; Feng, W.; Huang, H.; Liu, H.; Xu, R.; Lin, J.; Han, L.; Zhang, D. Natural medicines for the treatment of fatigue: Bioactive components, pharmacology, and mechanisms. Pharmacol. Res. 2019, 148, 104409. [Google Scholar] [CrossRef] [PubMed]
- Barabasi, A.-L.; Gulbahce, N.; Loscalzo, J. Network medicine: A network-based approach to human disease. Nat. Rev. Genet. 2011, 12, 56–68. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.F.; Hu, Y.Q.; Wu, Q.G.; Zhang, R. Virtual Screening of Potential Anti-fatigue Mechanism of Polygonati Rhizoma Based on Network Pharmacology. Comb. Chem. High Throughput Screen. 2019, 22, 612–624. [Google Scholar] [CrossRef] [PubMed]
- Giloteaux, L.; Goodrich, J.K.; Walters, W.A.; Levine, S.M.; Ley, R.E.; Hanson, M.R. Reduced diversity and altered composition of the gut microbiome in individuals with myalgic encephalomyelitis/chronic fatigue syndrome. Microbiome 2016, 4, 30. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.; Shi, C.; Xia, P.; Ning, K.; Xiang, H.; Xie, Q. Fermented Deer Blood Ameliorates Intense Exercise-Induced Fatigue via Modulating Small Intestine Microbiota and Metabolites in Mice. Nutrients 2021, 13, 1543. [Google Scholar] [CrossRef]
- Zhou, S.-S.; Zhou, J.; Xu, J.-D.; Shen, H.; Kong, M.; Yip, K.-M.; Han, Q.-B.; Zhao, Z.-Z.; Xu, J.; Chen, H.-B.; et al. Ginseng ameliorates exercise-induced fatigue potentially by regulating the gut microbiota. Food Funct. 2021, 12, 3954–3964. [Google Scholar] [CrossRef]
- Li, Z.; Lin, Z.; Lu, Z.; Ying, Z. Effects of a traditional Chinese medicine formula containing the Coix seed and Lotus seed on the intestinal morphology and microbiota of local piglets. AMB Express 2021, 11, 159. [Google Scholar] [CrossRef]
- Liu, L.; Zou, Z.; Yang, J.; Li, X.; Zhu, B.; Zhang, H.; Sun, Y.; Zhang, Y.; Zhang, Z.-J.; Wang, W. Jianpi Jieyu Decoction, an Empirical Herbal Formula, Exerts Psychotropic Effects in Association with Modulation of Gut Microbial Diversity and GABA Activity. Front. Pharmacol. 2021, 12, 681. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Xu, W.; Wang, N.; Jiang, W.; Cheng, Y.; Guo, Y.; Yao, W.; Hu, B.; Du, P.; Qian, H. Anti-fatigue effect of Lepidium meyenii Walp. (Maca) on preventing mitochondria-mediated muscle damage and oxidative stress in vivo and vitro. Food Funct. 2021, 12, 3132–3141. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Wang, R.; Hua, H.; Cheng, Y.; Guo, Y.; Qian, H.; Du, P. The macamide relieves fatigue by acting as inhibitor of inflammatory response in exercising mice: From central to peripheral. Eur. J. Pharmacol. 2022, 917, 174758. [Google Scholar] [CrossRef]
- Zhu, H.K.; Hu, B.; Hua, H.Y.; Liu, C.; Cheng, Y.L.; Guo, Y.H.; Yao, W.R.; Qian, H. Macamides: A review of structures, isolation, therapeutics and prospects. Food Res. Int. 2020, 138, 109819. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Y.; Zhang, C.Y.; Hui, H.Y.; Tan, Z.J. Effect of Gegenqinlian decoction on intestinal mucosal flora in mice with diarrhea induced by high temperature and humidity treatment. 3 Biotech 2021, 11, 83. [Google Scholar] [CrossRef]
- Liu, C.; Du, P.; Cheng, Y.; Guo, Y.; Hu, B.; Yao, W.; Zhu, X.; Qian, H. Study on fecal fermentation characteristics of aloe polysaccharides in vitro and their predictive modeling. Carbohydr. Polym. 2021, 256, 117571. [Google Scholar] [CrossRef]
- Fulle, S.; Mecocci, P.; Fano, G.; Vecchiet, I.; Vecchini, A.; Racciotti, D.; Cherubini, A.; Pizzigallo, E.; Vecchiet, L.; Senin, U.; et al. Specific oxidative alterations in vastus lateralis muscle of patients with the diagnosis of chronic fatigue syndrome. Free Radic. Bio Med. 2000, 29, 1252–1259. [Google Scholar] [CrossRef]
- Liu, C.; Hu, B.; Cheng, Y.L.; Guo, Y.H.; Yao, W.R.; Qian, H. In-depth analysis of the mechanisms of aloe polysaccharides on mitigating subacute colitis in mice via microbiota informatics. Carbohydr. Polym. 2021, 265, 11. [Google Scholar] [CrossRef]
- Sawada, D.; Kuwano, Y.; Tanaka, H.; Hara, S.; Uchiyama, Y.; Sugawara, T.; Fujiwara, S.; Rokutan, K.; Nishida, K. Daily intake of Lactobacillus gasseri CP2305 relieves fatigue and stress-related symptoms in male university Ekiden runners: A double-blind, randomized, and placebo-controlled clinical trial. J. Funct. Food. 2019, 57, 465–476. [Google Scholar] [CrossRef]
- Lee, M.C.; Hsu, Y.J.; Ho, H.H.; Hsieh, S.H.; Kuo, Y.W.; Sung, H.C.; Huang, C.C. Lactobacillus salivarius Subspecies salicinius SA-03 is a New Probiotic Capable of Enhancing Exercise Performance and Decreasing Fatigue. Microorganisms 2020, 8, 545. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-M.; Wei, L.; Chiu, Y.-S.; Hsu, Y.-J.; Tsai, T.-Y.; Wang, M.-F.; Huang, C.-C. Lactobacillus plantarum TWK10 Supplementation Improves Exercise Performance and Increases Muscle Mass in Mice. Nutrients 2016, 8, 205. [Google Scholar] [CrossRef] [PubMed]
- Jezbera, J.; Sharma, A.K.; Brandt, U.; Doolittle, W.F.; Hahn, M.W. ‘Candidatus Planktophila limnetica’, an actinobacterium representing one of the most numerically important taxa in freshwater bacterioplankton. Int. J. Syst. Evol. Microbiol. 2009, 59, 2864–2869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.-R.; Chen, H.-W.; Li, Y.; Zhou, M.-Y.; Wong, V.K.-W.; Jiang, Z.-H.; Zhang, W. Network Pharmacology Exploration Reveals Anti-Apoptosis as a Common Therapeutic Mechanism for Non-Alcoholic Fatty Liver Disease Treated with Blueberry Leaf Polyphenols. Nutrients 2021, 13, 4060. [Google Scholar] [CrossRef] [PubMed]
- Higgins, M.R.; Izadi, A.; Kaviani, M. Antioxidants and Exercise Performance: With a Focus on Vitamin E and C Supplementation. Int. J. Environ. Res. Public Health 2020, 17, 8452. [Google Scholar] [CrossRef] [PubMed]
- Wyckelsma, V.L.; Venckunas, T.; Brazaitis, M.; Gastaldello, S.; Snieckus, A.; Eimantas, N.; Baranauskiene, N.; Subocius, A.; Skurvydas, A.; Paasuke, M.; et al. Vitamin C and E Treatment Blunts Sprint Interval Training-Induced Changes in Inflammatory Mediator-, Calcium-, and Mitochondria-Related Signaling in Recreationally Active Elderly Humans. Antioxidants 2020, 9, 879. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Yang, Q.; Yang, B.; Xu, H.; Nasif, O.; Muruganantham, S.; Chen, J. Phyllanthin Averts Oxidative Stress and Neuroinflammation in Cerebral Ischemic-Reperfusion Injury through Modulation of the NF-kappa B and AMPK/Nrf2 Pathways. J. Environ. Pathol. Toxicol. Oncol. 2021, 40, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Huang, C.; Liu, Y.; Chen, T.; Huang, R.; Liang, M.; Zhang, J.; Xu, H. A Network Pharmacology Approach to Uncover the Potential Mechanism of Yinchensini Decoction. Evid.-Based Complementary Altern. Med. 2018, 2018. [Google Scholar] [CrossRef]
- Kim, J.-H.; Kang, D.-M.; Cho, Y.-J.; Hyun, J.-W.; Ahn, M.-J. Medicarpin Increases Antioxidant Genes by Inducing NRF2 Transcriptional Level in HeLa Cells. Antioxidants 2022, 11, 421. [Google Scholar] [CrossRef]
- Xia, C.; Chen, J.; Deng, J.-L.; Zhu, Y.-Q.; Li, W.-Y.; Jie, B.; Chen, T.-Y. Novel macamides from maca (Lepidium meyenii Walpers) root and their cytotoxicity. Phytochem. Lett. 2018, 25, 65–69. [Google Scholar] [CrossRef]
- Boolani, A.; Gallivan, K.M.; Ondrak, K.S.; Christopher, C.J.; Castro, H.F.; Campagna, S.R.; Taylor, C.M.; Luo, M.; Dowd, S.E.; Smith, M.L.; et al. Trait Energy and Fatigue May Be Connected to Gut Bacteria among Young Physically Active Adults: An Exploratory Study. Nutrients 2022, 14, 466. [Google Scholar] [CrossRef]
- Hsu, Y.J.; Huang, W.C.; Lin, J.S.; Chen, Y.M.; Ho, S.T.; Huang, C.C.; Tung, Y.T. Kefir Supplementation Modifies Gut Microbiota Composition, Reduces Physical Fatigue, and Improves Exercise Performance in Mice. Nutrients 2018, 10, 862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, M.; Lin, L.; Chen, H.; Ge, X.; Huang, Y.; Zheng, Z.; Li, S.; Pan, Y.; Liu, B.; Zeng, F. Anti-fatigue property of the oyster polypeptide fraction and its effect on gut microbiota in mice. Food Funct. 2020, 11, 8659–8669. [Google Scholar] [CrossRef] [PubMed]
- Abel, B.; Murakami, M.; Tosh, D.K.; Yu, J.; Lusvarghi, S.; Campbell, R.G.; Gao, Z.-G.; Jacobson, K.A.; Ambudkar, S.V. Interaction of A3 adenosine receptor ligands with the human multidrug transporter ABCG2. Eur. J. Med. Chem. 2022, 231, 114103. [Google Scholar] [CrossRef] [PubMed]
- Burns, G.L.; Hoedt, E.C.; Walker, M.M.; Talley, N.J.; Keely, S. Physiological mechanisms of unexplained (functional) gastrointestinal disorders. J. Physiol.-Lond. 2021, 599, 5141–5161. [Google Scholar] [CrossRef] [PubMed]
- Pryor, R.; Norvaisas, P.; Marinos, G.; Best, L.; Thingholm, L.B.; Quintaneiro, L.M.; De Haes, W.; Esser, D.; Waschina, S.; Lujan, C.; et al. Host-Microbe-Drug-Nutrient Screen Identifies Bacterial Effectors of Metformin Therapy. Cell 2019, 178, 1299–1312. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Major Bioactive Components | Contents |
|---|---|
| Total polysaccharides (mg/mL) | 34.78 |
| Reducing sugar (mg/mL) | 8.64 |
| Total proteins (mg/mL) | 0.812 |
| Total amino acids (mg/mL) | 1845.27 |
| Total fatty acids (μg/mL) | 110.59 |
| Total flavonoids (mg/mL) | 0.157 |
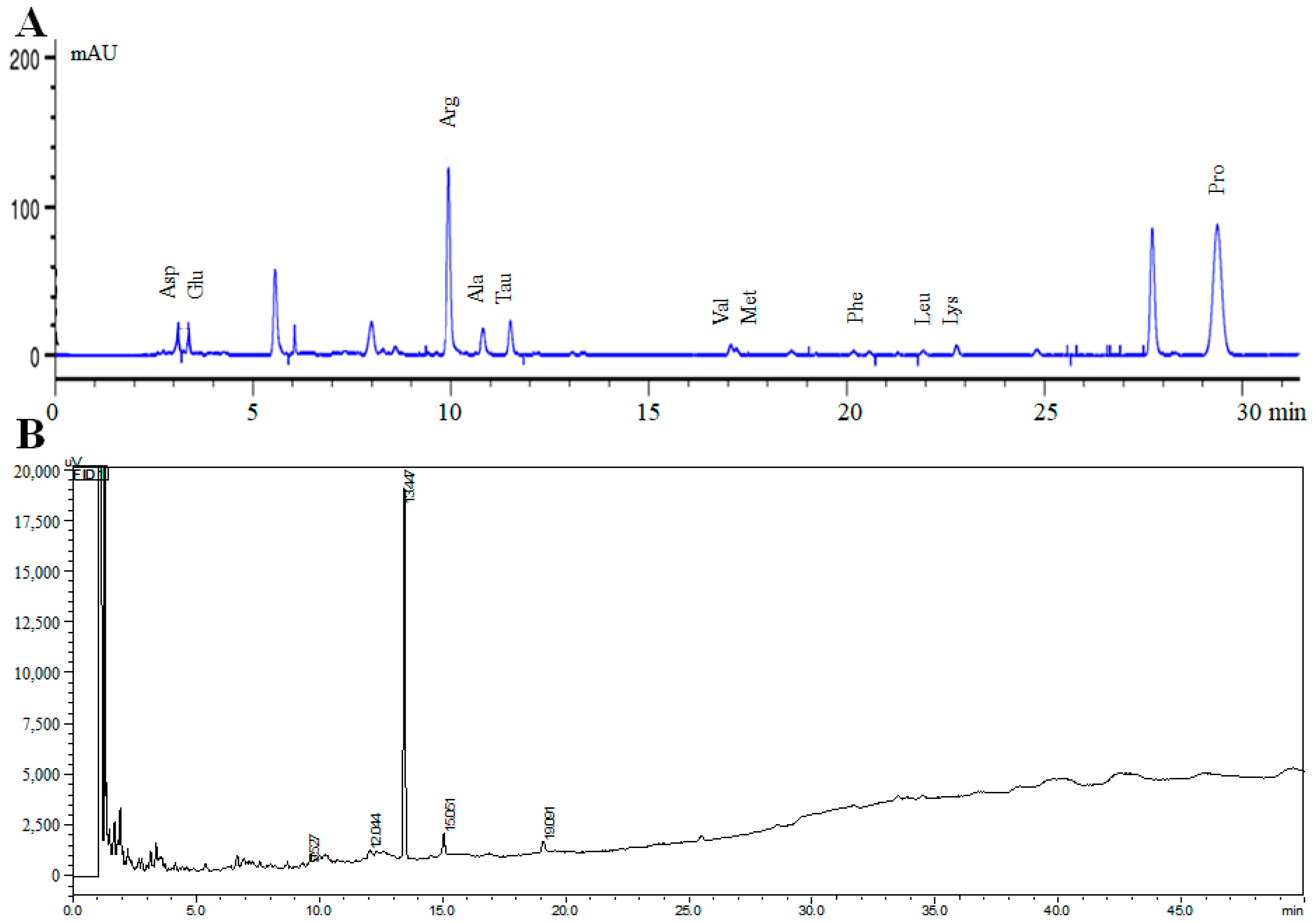
| Amino Acids | Ret. Time (min) | Peak Area (mAU`S) | Contents (μg/mL) | |
|---|---|---|---|---|
| Amino Acids | Asp | 3.123 | 267.597 | 3.486 |
| Glu | 3.377 | 133.185 | 4.081 | |
| Ser | 6.403 | 3.658 | 2.706 | |
| His | 7.342 | 82.776 | 5.315 | |
| Gly | 8.303 | 51.938 | 1.955 | |
| Thr | 8.614 | 267.514 | 3.230 | |
| Arg | 9.943 | 3420.104 | 4.611 | |
| Ala | 10.820 | 144.076 | 2.256 | |
| Tau | 11.503 | 466.864 | 3.310 | |
| Tyr | 13.076 | 26.769 | 5.064 | |
| Cys | 16.360 | 6.677 | 4.948 | |
| Val | 17.056 | 34.216 | 3.037 | |
| Met | 17.50 | 11.686 | 3.760 | |
| Trp | 19.216 | 19.450 | 6.350 | |
| Phe | 20.175 | 36.052 | 4.441 | |
| Ile | 20.552 | 46.827 | 3.363 | |
| Leu | 21.916 | 53.758 | 3.335 | |
| Lys | 22.752 | 211.235 | 2.224 | |
| Pro | 29.295 | 8255.328 | 2.010 | |
| Fatty Acids | C12:0 | 9.527 | 828 | 0.769 |
| C14:0 | 12.044 | 3292 | 3.059 | |
| C15:0 | 13.447 | 92632 | 86.08 | |
| C16:0 | 15.051 | 6117 | 5.685 | |
| C18:0 | 19.091 | 4742 | 4.407 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, H.; Wang, R.; Hua, H.; Cheng, Y.; Guo, Y.; Qian, H.; Du, P. Network Pharmacology Exploration Reveals Gut Microbiota Modulation as a Common Therapeutic Mechanism for Anti-Fatigue Effect Treated with Maca Compounds Prescription. Nutrients 2022, 14, 1533. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14081533
Zhu H, Wang R, Hua H, Cheng Y, Guo Y, Qian H, Du P. Network Pharmacology Exploration Reveals Gut Microbiota Modulation as a Common Therapeutic Mechanism for Anti-Fatigue Effect Treated with Maca Compounds Prescription. Nutrients. 2022; 14(8):1533. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14081533
Chicago/Turabian StyleZhu, Hongkang, Ruoyong Wang, Hanyi Hua, Yuliang Cheng, Yahui Guo, He Qian, and Peng Du. 2022. "Network Pharmacology Exploration Reveals Gut Microbiota Modulation as a Common Therapeutic Mechanism for Anti-Fatigue Effect Treated with Maca Compounds Prescription" Nutrients 14, no. 8: 1533. https://0-doi-org.brum.beds.ac.uk/10.3390/nu14081533